
Cell-Based Regeneration and Treatment of Liver Diseases

 , , ,
, , ,  ,
,  ,
, {kind=link}
{kind=link}
Abstract
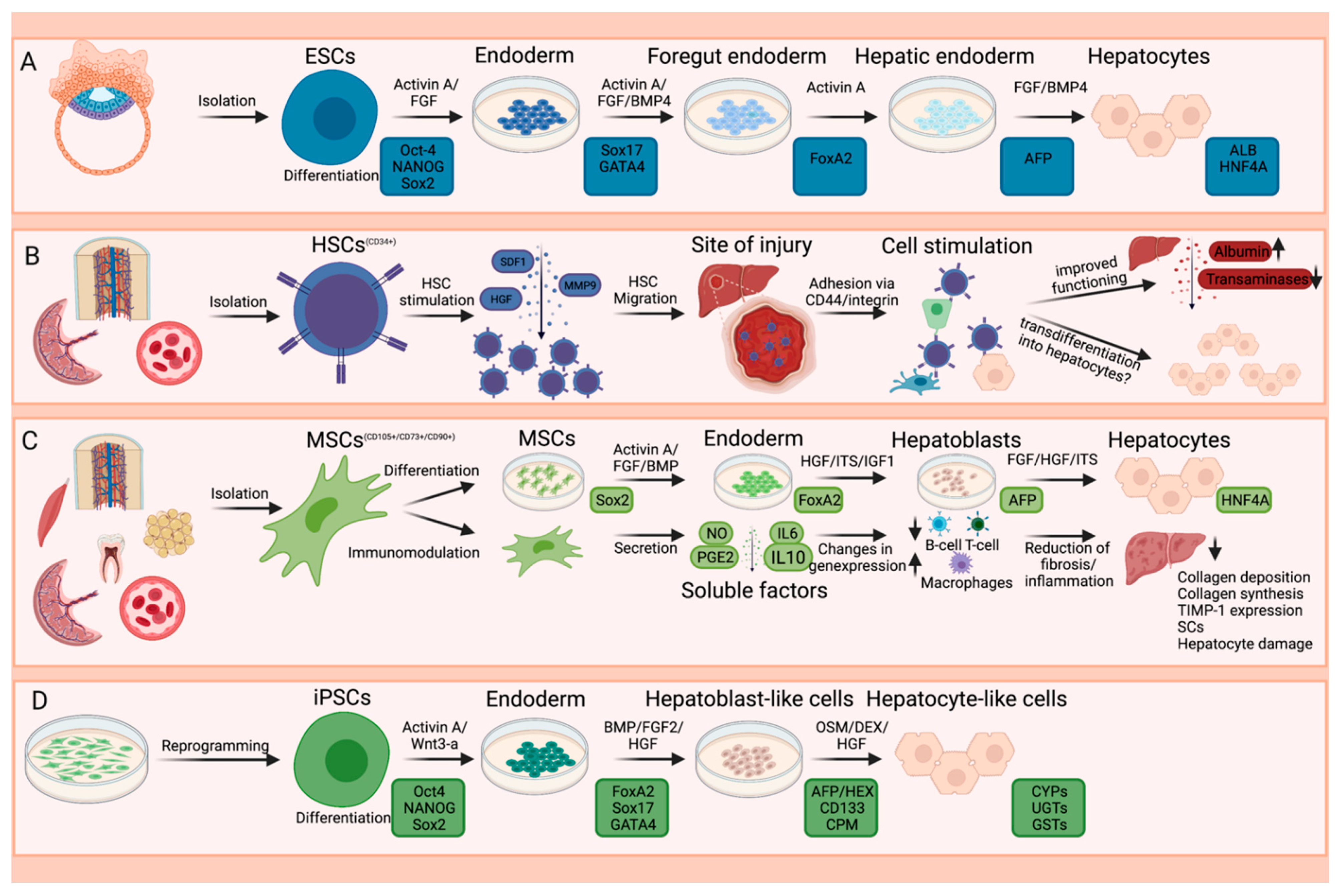
:1. Introduction
2. Liver Cell Types and Their Potential of Regeneration
2.1. Microanatomy of the Liver
2.2. Biliary Epithelial Cells—The Lining of the Biliary Tree
2.3. Mechanisms of Liver Cell Regeneration
3. Cellular Therapies for Treatment of Liver Diseases and Induction of Regeneration
3.1. Stem Cells: Sources, Mechanism of Action, and Applications
3.1.1. Embryonal Stem Cells (ESCs)
3.1.2. Hematopoietic Stem Cells (HSCs)

3.1.3. Induced Pluripotent Stem Cells (iPSCs)
3.1.4. Mesenchymal Stem Cells (MSCs)
3.2. Immune Cell Therapies: A Novel Paradigm in the Development of Therapies
3.2.1. Regulatory T-Cells (Tregs)
3.2.2. Chimeric Antigen Receptor-Engineered T-Cells (CAR-Ts)
3.2.3. Dendritic Cells (DCs)
3.2.4. Macrophages, Monocytes, and Kupffer Cells (KCs)
4. Technological Developments for Improved Delivery and Engraftment of Cell-Based Therapeutics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AFP | Alpha-1-fetoprotein |
| AIH | Autoimmune hepatitis |
| BEC | Biliary epithelial cells |
| bm | Bone marrow |
| BMP | Bone morphogenic protein |
| CAR-T | Chimeric antigen receptor-engineered T cells |
| CCL4 | Carbon tetrachloride |
| CDH11 | Cadherin 11 |
| CPM | Carboxypeptidase M |
| CTLA4 | Cytotoxic T lymphocyte-associated antigen 4 |
| CXCR | C-X-C motif chemokine receptor |
| DC | Dendritic cells |
| DCreg | Regulatory DC |
| DEX | Dexamethasone |
| DLL4 | Delta-ligand like 4 |
| dp | Dental pulp |
| EGFR | Epidermal growth factor receptor |
| ESC | Embryonic stem cell |
| EV | Extracellular vesicles |
| FGF | Fibroblast growth factor |
| FoxA2 | Forkhead-box protein A2 |
| G-CSF | Granulocyte colony-stimulating factor |
| GATA4 | Gata-binding protein 4 |
| GM-CSF | Granulocyte-monocyte colony-stimulating factor |
| GPC3 | Glypican 3 |
| GvHD | Graft-versus-host disease |
| HA | Hyaluronic acid |
| HBC | Hepatitis B |
| HCC | Hepatocellular carcinoma |
| HCV | Hepatitis C |
| HGF | Hepatocyte growth factor |
| HLC | Hepatoblast-like cells |
| HNF4α | Hepatocyte nuclear factor 4α |
| HSC | Hematopoietic stem cell |
| IGF1 | Insulin growth factor 1 |
| iHep | Induced hepatocyte-like cells |
| IL | Interleukin |
| INFγ | Interferon gamma |
| INR | International normalized ratio |
| iPSC | Induced pluripotent stem cell |
| ITS | Insulin-transferrin-sodium selenite |
| KC | Kupffer cells |
| LSEC | Liver sinusoidal endothelial cells |
| MMP | Matrix metallopeptidase |
| MoMF | Monocyte-derived macrophage |
| MSC | Mesenchymal stem cell |
| MUC | Mucin 1 |
| NMP | Normothermic machine perfusion |
| NO | Nitric oxide |
| NPC | Non-parenchymal cells |
| OLT | Orthoptic liver transplantation |
| Oct4 | Octamer binding transcription factor 4 |
| PEG2 | Prostaglansin E2 |
| RFP | Red flourescene protein |
| SC | Hepatic stellate cells |
| scFv | Single chain variable fragment |
| SDF1 | Stromal-cell-derived factor 1 |
| Sox | SRY-Box Transcription Factor |
| TAA | Tumor-assocated antigens |
| TAZ | PDZ-minding motif |
| TET1 | Ten-eleven translocation 1 |
| TGFα | Transforming growth factor alpha |
| TIMP1 | Inhibitor of metalloprotease 1 |
| TNFα | Tumor necrosis factor alpha |
| Tregs | Regulatory T cells |
| uc | umbilical cord |
| VEGF | Vascular endothelial growth factor |
| YAP | Yes-associated protein |
References
- Trefts, E.; Gannon, M.; Wasserman, D.H. The liver. Curr. Biol. 2017, 27, R1147–R1151. [Google Scholar] [CrossRef]
- Racanelli, V.; Rehermann, B. The liver as an immunological organ. Hepatology 2006, 43, S54–S62. [Google Scholar] [CrossRef]
- Guillot, A.; Tacke, F. Liver Macrophages: Old Dogmas and New Insights. Hepatol. Commun. 2019, 3, 730–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Chang-Yong, E.; Gong, Z.W.; Liu, S.; Wang, Z.X.; Yang, Y.S.; Zhang, X.W. Chimeric antigen receptor-engineered T-cell therapy for liver cancer. Hepatobiliary Pancreat. Dis. Int. 2018, 17, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Aravalli, R.N.; Steer, C.J. Immune-Mediated Therapies for Liver Cancer. Genes 2017, 8, 76. [Google Scholar] [CrossRef] [Green Version]
- Pinato, D.J.; Guerra, N.; Fessas, P.; Murphy, R.; Mineo, T.; Mauri, F.A.; Mukherjee, S.K.; Thursz, M.; Wong, C.N.; Sharma, R.; et al. Immune-based therapies for hepatocellular carcinoma. Oncogene 2020, 39, 3620–3637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asrani, S.K.; Devarbhavi, H.; Eaton, J.; Kamath, P.S. Burden of liver diseases in the world. J. Hepatol. 2019, 70, 151–171. [Google Scholar] [CrossRef]
- Ding, C.; Li, Y.; Guo, F.; Jiang, Y.; Ying, W.; Li, D.; Yang, D.; Xia, X.; Liu, W.; Zhao, Y.; et al. A Cell-type-resolved Liver Proteome. Mol. Cell Proteom. 2016, 15, 3190–3202. [Google Scholar] [CrossRef] [Green Version]
- Strazzabosco, M.; Fabris, L. Functional anatomy of normal bile ducts. Anat. Rec. 2008, 291, 653–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubes, P.; Jenne, C. Immune Responses in the Liver. Annu. Rev. Immunol. 2018, 36, 247–277. [Google Scholar] [CrossRef]
- Doherty, D.G. Immunity, tolerance and autoimmunity in the liver: A comprehensive review. J. Autoimmun. 2016, 66, 60–75. [Google Scholar] [CrossRef]
- Lu, W.Y.; Bird, T.G.; Boulter, L.; Tsuchiya, A.; Cole, A.M.; Hay, T.; Guest, R.V.; Wojtacha, D.; Man, T.Y.; Mackinnon, A.; et al. Hepatic progenitor cells of biliary origin with liver repopulation capacity. Nat. Cell Biol. 2015, 17, 971–983. [Google Scholar] [CrossRef]
- Brunner, S.M.; Junger, H.; Ruemmele, P.; Schnitzbauer, A.A.; Doenecke, A.; Kirchner, G.I.; Farkas, S.A.; Loss, M.; Scherer, M.N.; Schlitt, H.J.; et al. Bile duct damage after cold storage of deceased donor livers predicts biliary complications after liver transplantation. J. Hepatol. 2013, 58, 1133–1139. [Google Scholar] [CrossRef]
- Tabibian, J.H.; Masyuk, A.I.; Masyuk, T.V.; O’Hara, S.P.; LaRusso, N.F. Physiology of cholangiocytes. Compr. Physiol. 2013, 3, 541–565. [Google Scholar]
- Keplinger, K.M.; Bloomston, M. Anatomy and embryology of the biliary tract. Surg. Clin. N. Am. 2014, 94, 203–217. [Google Scholar] [CrossRef]
- Alpini, G.; Lenzi, R.; Sarkozi, L.; Tavoloni, N. Biliary physiology in rats with bile ductular cell hyperplasia. Evidence for a secretory function of proliferated bile ductules. J. Clin. Investig. 1988, 81, 569–578. [Google Scholar] [CrossRef] [Green Version]
- Alpini, G.; Lenzi, R.; Zhai, W.R.; Slott, P.A.; Liu, M.H.; Sarkozi, L.; Tavoloni, N. Bile secretory function of intrahepatic biliary epithelium in the rat. Am. J. Physiol. 1989, 257, G124–G133. [Google Scholar] [CrossRef] [PubMed]
- Alpini, G.; Ueno, Y.; Glaser, S.S.; Marzioni, M.; Phinizy, J.L.; Francis, H.; Lesage, G. Bile acid feeding increased proliferative activity and apical bile acid transporter expression in both small and large rat cholangiocytes. Hepatology 2001, 34, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Alpini, G.; Roberts, S.; Kuntz, S.M.; Ueno, Y.; Gubba, S.; Podila, P.V.; LeSage, G.; LaRusso, N.F. Morphological, molecular, and functional heterogeneity of cholangiocytes from normal rat liver. Gastroenterology 1996, 110, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Alpini, G.; Glaser, S.; Robertson, W.; Rodgers, R.E.; Phinizy, J.L.; Lasater, J.; LeSage, G.D. Large but not small intrahepatic bile ducts are involved in secretin-regulated ductal bile secretion. Am. J. Physiol. 1997, 272, G1064–G1074. [Google Scholar] [CrossRef]
- Martinez-Anso, E.; Castillo, J.E.; Diez, J.; Medina, J.F.; Prieto, J. Immunohistochemical detection of chloride/bicarbonate anion exchangers in human liver. Hepatology 1994, 19, 1400–1406. [Google Scholar] [CrossRef]
- Glaser, S.S.; Gaudio, E.; Rao, A.; Pierce, L.M.; Onori, P.; Franchitto, A.; Francis, H.L.; Dostal, D.E.; Venter, J.K.; DeMorrow, S.; et al. Morphological and functional heterogeneity of the mouse intrahepatic biliary epithelium. Lab. Investig. 2009, 89, 456–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazaridis, K.N.; Pham, L.; Vroman, B.; de Groen, P.C.; LaRusso, N.F. Kinetic and molecular identification of sodium-dependent glucose transporter in normal rat cholangiocytes. Am. J. Physiol. 1997, 272, G1168–G1174. [Google Scholar] [CrossRef]
- Ballatori, N.; Jacob, R.; Barrett, C.; Boyer, J.L. Biliary catabolism of glutathione and differential reabsorption of its amino acid constituents. Am. J. Physiol. 1988, 254, G1–G7. [Google Scholar] [CrossRef]
- Hohenester, S.; Wenniger, L.M.; Paulusma, C.C.; van Vliet, S.J.; Jefferson, D.M.; Elferink, R.P.; Beuers, U. A biliary HCO3- umbrella constitutes a protective mechanism against bile acid-induced injury in human cholangiocytes. Hepatology 2012, 55, 173–183. [Google Scholar] [CrossRef]
- Hohenester, S.; Maillette de Buy Wenniger, L.; Jefferson, D.M.; Oude Elferink, R.P.; Beuers, U. Biliary bicarbonate secretion constitutes a protective mechanism against bile acid-induced injury in man. Dig. Dis. 2011, 29, 62–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhushan, B.; Apte, U. Liver Regeneration after Acetaminophen Hepatotoxicity: Mechanisms and Therapeutic Opportunities. Am. J. Pathol. 2019, 189, 719–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalopoulos, G.K. Liver regeneration. J. Cell Physiol. 2007, 213, 286–300. [Google Scholar] [CrossRef]
- Michalopoulos, G.K.; Bhushan, B. Liver regeneration: Biological and pathological mechanisms and implications. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 40–55. [Google Scholar] [CrossRef]
- Campana, L.; Esser, H.; Huch, M.; Forbes, S. Liver regeneration and inflammation: From fundamental science to clinical applications. Nat. Rev. Mol. Cell Biol. 2021. [Google Scholar] [CrossRef]
- Fausto, N.; Campbell, J.S.; Riehle, K.J. Liver regeneration. Hepatology 2006, 43, S45–S53. [Google Scholar] [CrossRef] [PubMed]
- Michalopoulos, G.K. Principles of liver regeneration and growth homeostasis. Compr. Physiol. 2013, 3, 485–513. [Google Scholar]
- Marubashi, S.; Sakon, M.; Nagano, H.; Gotoh, K.; Hashimoto, K.; Kubota, M.; Kobayashi, S.; Yamamoto, S.; Miyamoto, A.; Dono, K.; et al. Effect of portal hemodynamics on liver regeneration studied in a novel portohepatic shunt rat model. Surgery 2004, 136, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Mars, W.M.; Liu, M.-L.; Kitson, R.P.; Goldfarb, R.H.; Gabauer, M.K.; Michalopoulos, G.K. Immediate Early Detection of Urokinase Receptor After Partial Hepatectomy and Its Implications for Initiation of Liver Regeneration. Hepatology 1995, 21, 1695–1701. [Google Scholar] [PubMed]
- Kim, T.-H.; Mars, W.M.; Stolz, D.B.; Michalopoulos, G.K. Expression and Activation of Pro-MMP-2 and Pro-MMP-9 during Rat Liver Regeneration. Hepatology 2000, 31, 75–82. [Google Scholar] [CrossRef]
- Lindroos, P.M.; Zarnegar, R.; Michalopoulos, G.K. Hepatocyte Growth Factor (Hepatopoietin A) Rapidly Increases in Plasma before DNA Synthesis and Liver Regeneration Stimulated by Partial Hepatectomy and Carbon Tetrachloride Administration. Hepatology 1991, 13, 744–750. [Google Scholar] [CrossRef]
- Patijn, G.A.; Lieber, A.; Schowalter, D.B.; Schwall, R.; Kay, M.A. Hepatocyte Growth Factor Induces Hepatocyte Proliferation In Vivo and Allows for Effiecient Retroviral-Mediated Gene Transfer in Mice. Hepatology 1998, 28, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Gilgenkrantz, H.; Collin de l’Hortet, A. Understanding Liver Regeneration: From Mechanisms to Regenerative Medicine. Am. J. Pathol. 2018, 188, 1316–1327. [Google Scholar] [CrossRef] [Green Version]
- Pepe-Mooney, B.J.; Dill, M.T.; Alemany, A.; Ordovas-Montanes, J.; Matsushita, Y.; Rao, A.; Sen, A.; Miyazaki, M.; Anakk, S.; Dawson, P.A.; et al. Single-Cell Analysis of the Liver Epithelium Reveals Dynamic Heterogeneity and an Essential Role for YAP in Homeostasis and Regeneration. Cell Stem Cell 2019, 25, 23–38.e28. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Lu, H.; Zou, Q.; Luo, L. Regeneration of liver after extreme hepatocyte loss occurs mainly via biliary transdifferentiation in zebrafish. Gastroenterology 2014, 146, 789–800.e788. [Google Scholar] [CrossRef]
- Raven, A.; Lu, W.Y.; Man, T.Y.; Ferreira-Gonzalez, S.; O’Duibhir, E.; Dwyer, B.J.; Thomson, J.P.; Meehan, R.R.; Bogorad, R.; Koteliansky, V.; et al. Cholangiocytes act as facultative liver stem cells during impaired hepatocyte regeneration. Nature 2017, 547, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Speicher, T.; Siegenthaler, B.; Bogorad, R.L.; Ruppert, R.; Petzold, T.; Padrissa-Altes, S.; Bachofner, M.; Anderson, D.G.; Koteliansky, V.; Fässler, R.; et al. Knockdown and knockout of β1-integrin in hepatocytes impairs liver regeneration through inhibition of growth factor signalling. Nat. Commun. 2014, 5, 3862. [Google Scholar] [CrossRef] [Green Version]
- Huch, M.; Bonfanti, P.; Boj, S.F.; Sato, T.; Loomans, C.J.M.; van de Wetering, M.; Sojoodi, M.; Li, V.S.W.; Schuijers, J.; Gracanin, A.; et al. Unlimited in vitro expansion of adult bi-potent pancreas progenitors through the Lgr5/R-spondin axis. EMBO J. 2013, 32, 2708–2721. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.; Upadhyay, N.; Greenbaum, L.E.; Kaestner, K.H. Ablation of Foxl1-Cre-labeled hepatic progenitor cells and their descendants impairs recovery of mice from liver injury. Gastroenterology 2015, 148, 192–202.e193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espanol-Suner, R.; Carpentier, R.; Van Hul, N.; Legry, V.; Achouri, Y.; Cordi, S.; Jacquemin, P.; Lemaigre, F.; Leclercq, I.A. Liver progenitor cells yield functional hepatocytes in response to chronic liver injury in mice. Gastroenterology 2012, 143, 1564–1575.e1567. [Google Scholar] [CrossRef]
- Suzuki, A.; Sekiya, S.; Buscher, D.; Izpisua Belmonte, J.C.; Taniguchi, H. Tbx3 controls the fate of hepatic progenitor cells in liver development by suppressing p19ARF expression. Development 2008, 135, 1589–1595. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Zhou, Y.; Hu, T.; Zhang, H.; Shen, M.; Cheng, P.; Dai, W.; Wang, F.; Chen, K.; Zhang, Y.; et al. Notch Signaling Coordinates Progenitor Cell-Mediated Biliary Regeneration Following Partial Hepatectomy. Sci. Rep. 2016, 6, 22754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulter, L.; Govaere, O.; Bird, T.G.; Radulescu, S.; Ramachandran, P.; Pellicoro, A.; Ridgway, R.A.; Seo, S.S.; Spee, B.; Van Rooijen, N.; et al. Macrophage-derived Wnt opposes Notch signaling to specify hepatic progenitor cell fate in chronic liver disease. Nat. Med. 2012, 18, 572–579. [Google Scholar] [CrossRef] [Green Version]
- Aloia, L.; McKie, M.A.; Vernaz, G.; Cordero-Espinoza, L.; Aleksieva, N.; van den Ameele, J.; Antonica, F.; Font-Cunill, B.; Raven, A.; Aiese Cigliano, R.; et al. Epigenetic remodelling licences adult cholangiocytes for organoid formation and liver regeneration. Nat. Cell Biol. 2019, 21, 1321–1333. [Google Scholar] [CrossRef]
- Yimlamai, D.; Christodoulou, C.; Galli, G.G.; Yanger, K.; Pepe-Mooney, B.; Gurung, B.; Shrestha, K.; Cahan, P.; Stanger, B.Z.; Camargo, F.D. Hippo pathway activity influences liver cell fate. Cell 2014, 157, 1324–1338. [Google Scholar] [CrossRef] [Green Version]
- Fukano, S.; Saitoh, Y.; Uchida, K.; Akiyoshi, T.; Takeda, K.i. Bile acid metabolism in partially hepatectomized rats. Steroids 1985, 45, 209–227. [Google Scholar] [CrossRef]
- Lesage, G.; Glaser, S.S.; Gubba, S.; Robertson, W.E.; Phinizy, J.L.; Lasater, J.; Rodgers, R.E.; Alpini, G. Regrowth of the rat biliary tree after 70% partial hepatectomy is coupled to increased secretin-induced ductal secretion. Gastroenterology 1996, 111, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Planas-Paz, L.; Sun, T.; Pikiolek, M.; Cochran, N.R.; Bergling, S.; Orsini, V.; Yang, Z.; Sigoillot, F.; Jetzer, J.; Syed, M.; et al. YAP, but Not RSPO-LGR4/5, Signaling in Biliary Epithelial Cells Promotes a Ductular Reaction in Response to Liver Injury. Cell Stem Cell 2019, 25, 39–53.e10. [Google Scholar] [CrossRef] [PubMed]
- Font-Burgada, J.; Shalapour, S.; Ramaswamy, S.; Hsueh, B.; Rossell, D.; Umemura, A.; Taniguchi, K.; Nakagawa, H.; Valasek, M.A.; Ye, L.; et al. Hybrid Periportal Hepatocytes Regenerate the Injured Liver without Giving Rise to Cancer. Cell 2015, 162, 766–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Wang, Y.; Pu, W.; Huang, X.; Qiu, L.; Li, Y.; Yu, W.; Zhao, H.; Liu, X.; He, L.; et al. Lineage Tracing Reveals the Bipotency of SOX9+ Hepatocytes during Liver Regeneration. Stem Cell Rep. 2019, 12, 624–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yimlamai, D.; Fowl, B.H.; Camargo, F.D. Emerging evidence on the role of the Hippo/YAP pathway in liver physiology and cancer. J. Hepatol. 2015, 63, 1491–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iansante, V.; Mitry, R.R.; Filippi, C.; Fitzpatrick, E.; Dhawan, A. Human hepatocyte transplantation for liver disease: Current status and future perspectives. Pediatr. Res. 2018, 83, 232–240. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.P.; Jiang, Y.Z.; Sun, L.Y.; Zhu, Z.J. Therapeutic effect and safety of stem cell therapy for chronic liver disease: A systematic review and meta-analysis of randomized controlled trials. Stem Cell Res. Ther. 2020, 11, 419. [Google Scholar] [CrossRef]
- Cantz, T.; Manns, M.P.; Ott, M. Stem cells in liver regeneration and therapy. Cell Tissue Res. 2008, 331, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Tsolaki, E.; Yannaki, E. Stem cell-based regenerative opportunities for the liver: State of the art and beyond. World J. Gastroenterol. 2015, 21, 12334–12350. [Google Scholar] [CrossRef]
- Boyd, A.; Newsome, P.; Lu, W.Y. The role of stem cells in liver injury and repair. Expert Rev. Gastroenterol. Hepatol. 2019, 13, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Sun, M.; Liu, W.; Li, Y.; Li, M. Stem Cell-Based Therapies for Liver Diseases: An Overview and Update. Tissue Eng. Regen. Med. 2019, 16, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Hannan, N.R.; Segeritz, C.P.; Touboul, T.; Vallier, L. Production of hepatocyte-like cells from human pluripotent stem cells. Nat. Protoc. 2013, 8, 430–437. [Google Scholar] [CrossRef] [Green Version]
- Tolosa, L.; Caron, J.; Hannoun, Z.; Antoni, M.; López, S.; Burks, D.; Castell, J.V.; Weber, A.; Gomez-Lechon, M.J.; Dubart-Kupperschmitt, A. Transplantation of hESC-derived hepatocytes protects mice from liver injury. Stem Cell Res. Ther. 2015, 6, 246. [Google Scholar] [CrossRef] [Green Version]
- Haideri, S.S.; McKinnon, A.C.; Taylor, A.H.; Kirkwood, P.; Starkey Lewis, P.J.; O’Duibhir, E.; Vernay, B.; Forbes, S.; Forrester, L.M. Injection of embryonic stem cell derived macrophages ameliorates fibrosis in a murine model of liver injury. NPJ Regen. Med. 2017, 2, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prior, N.; Inacio, P.; Huch, M. Liver organoids: From basic research to therapeutic applications. Gut 2019, 68, 2228–2237. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Wang, X.; Tan, Z.; Su, Y.; Liu, J.; Chang, M.; Yan, F.; Chen, J.; Chen, T.; Li, C.; et al. Human ESC-derived expandable hepatic organoids enable therapeutic liver repopulation and pathophysiological modeling of alcoholic liver injury. Cell Res. 2019, 29, 1009–1026. [Google Scholar] [CrossRef] [PubMed]
- Ramli, M.N.B.; Lim, Y.S.; Koe, C.T.; Demircioglu, D.; Tng, W.; Gonzales, K.A.U.; Tan, C.P.; Szczerbinska, I.; Liang, H.; Soe, E.L.; et al. Human Pluripotent Stem Cell-Derived Organoids as Models of Liver Disease. Gastroenterology 2020, 159, 1471–1486.e1412. [Google Scholar] [CrossRef]
- Mun, S.J.; Ryu, J.S.; Lee, M.O.; Son, Y.S.; Oh, S.J.; Cho, H.S.; Son, M.Y.; Kim, D.S.; Kim, S.J.; Yoo, H.J.; et al. Generation of expandable human pluripotent stem cell-derived hepatocyte-like liver organoids. J. Hepatol. 2019, 71, 970–985. [Google Scholar] [CrossRef]
- Kollet, O.; Shivtiel, S.; Chen, Y.-Q.; Suriawinata, J.; Thung, S.N.; Dabeva, M.D.; Kahn, J.; Spiegel, A.; Dar, A.; Samira, S.; et al. HGF, SDF-1, and MMP-9 are involved in stress-induced human CD34+ stem cell recruitment to the liver. J. Clin. Investig. 2003, 112, 160–169. [Google Scholar] [CrossRef]
- Crosby, H.A.; Lalor, P.F.; Ross, E.; Newsome, P.N.; Adams, D.H. Adhesion of human haematopoietic (CD34+) stem cells to human liver compartments is integrin and CD44 dependent and modulated by CXCR3 and CXCR4. J. Hepatol. 2009, 51, 734–749. [Google Scholar] [CrossRef]
- Lee, J.Y.; Hong, S.H. Hematopoietic Stem Cells and Their Roles in Tissue Regeneration. Int. J. Stem Cells 2020, 13, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lagasse, E.; Connors, H.; Al-Dhalimy, M.; Reitsma, M.; Dohse, M.; Osborne, L.; Wang, X.; Finegold, M.; Weissman, I.L.; Grompe, M. Purified hematopoietic stem cells can differentiate into hepatocytes in vivo. Nat. Med. 2000, 6, 1229–1234. [Google Scholar] [CrossRef]
- Khurana, S.; Jaiswal, A.K.; Mukhopadhyay, A. Hepatocyte nuclear factor-4alpha induces transdifferentiation of hematopoietic cells into hepatocytes. J. Biol. Chem. 2010, 285, 4725–4731. [Google Scholar] [CrossRef] [Green Version]
- Sellamuthu, S.; Manikandan, R.; Thiagarajan, R.; Babu, G.; Dinesh, D.; Prabhu, D.; Arulvasu, C. In vitro trans-differentiation of human umbilical cord derived hematopoietic stem cells into hepatocyte like cells using combination of growth factors for cell based therapy. Cytotechnology 2011, 63, 259–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazaro, C.A.; Croager, E.J.; Mitchell, C.; Campbell, J.S.; Yu, C.; Foraker, J.; Rhim, J.A.; Yeoh, G.C.; Fausto, N. Establishment, characterization, and long-term maintenance of cultures of human fetal hepatocytes. Hepatology 2003, 38, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Vassilopoulos, G.; Wang, P.-R.; Russel, D. Transplanted Bone Marrow Regenerates Liver By Cell Fusion. Nature 2003, 422, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Willenbring, H.; Akkari, Y.; Torimaru, Y.; Foster, M.; Al-Dhalimy, M.; Lagasse, E.; Finegold, M.; Olson, S.; Grompe, M. Cell Fusion Is The Principal Source Of Bone-Marrow-Derived Hepatocytes. Nature 2003, 422, 893–897. [Google Scholar] [CrossRef]
- Lizier, M.; Castelli, A.; Montagna, C.; Lucchini, F.; Vezzoni, P.; Faggioli, F. Cell fusion in the liver, revisited. World J. Hepatol. 2018, 10, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Sakaida, I.; Terai, S.; Yamamoto, N.; Aoyama, K.; Ishikawa, T.; Nishina, H.; Okita, K. Transplantation of bone marrow cells reduces CCl4-induced liver fibrosis in mice. Hepatology 2004, 40, 1304–1311. [Google Scholar] [CrossRef]
- Salama, H.; Zekri, A.R.; Zern, M.; Bahnassy, A.; Loutfy, S.; Shalaby, S.; Vigen, C.; Burke, W.; Mostafa, M.; Medhat, E.; et al. Autologous hematopoietic stem cell transplantation in 48 patients with end-stage chronic liver diseases. Cell Transplant. 2010, 19, 1475–1486. [Google Scholar] [CrossRef] [PubMed]
- Zekri, A.R.; Salama, H.; Medhat, E.; Musa, S.; Abdel-Haleem, H.; Ahmed, O.S.; Khedr, H.A.; Lotfy, M.M.; Zachariah, K.S.; Bahnassy, A.A. The impact of repeated autologous infusion of haematopoietic stem cells in patients with liver insufficiency. Stem Cell Res. Ther. 2015, 6, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pittenger, M.F.; Discher, D.E.; Peault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Asgari, S.; Moslem, M.; Bagheri-Lankarani, K.; Pournasr, B.; Miryounesi, M.; Baharvand, H. Differentiation and transplantation of human induced pluripotent stem cell-derived hepatocyte-like cells. Stem Cell Rev. Rep. 2013, 9, 493–504. [Google Scholar] [CrossRef]
- Choi, J.S.; Jeong, I.S.; Park, Y.J.; Kim, S.W. HGF and IL-10 expressing ALB::GFP reporter cells generated from iPSCs show robust anti-fibrotic property in acute fibrotic liver model. Stem Cell Res. Ther. 2020, 11, 332. [Google Scholar] [CrossRef]
- Tasnim, F.; Xing, J.; Huang, X.; Mo, S.; Wei, X.; Tan, M.H.; Yu, H. Generation of mature kupffer cells from human induced pluripotent stem cells. Biomaterials 2019, 192, 377–391. [Google Scholar] [CrossRef]
- Povero, D.; Pinatel, E.M.; Leszczynska, A.; Goyal, N.P.; Nishio, T.; Kim, J.; Kneiber, D.; de Araujo Horcel, L.; Eguchi, A.; Ordonez, P.M.; et al. Human induced pluripotent stem cell-derived extracellular vesicles reduce hepatic stellate cell activation and liver fibrosis. JCI Insight 2019, 5, e125652. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Snykers, S.; Vanhaecke, T.; Papeleu, P.; Luttun, A.; Jiang, Y.; Vander Heyden, Y.; Verfaillie, C.; Rogiers, V. Sequential exposure to cytokines reflecting embryogenesis: The key for in vitro differentiation of adult bone marrow stem cells into functional hepatocyte-like cells. Toxicol. Sci. 2006, 94, 330–341, discussion 235–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayatollahi, M.; Soleimani, M.; Tabei, S.Z.; Kabir Salmani, M. Hepatogenic differentiation of mesenchymal stem cells induced by insulin like growth factor-I. World J. Stem Cells 2011, 3, 113–121. [Google Scholar] [CrossRef]
- Afshari, A.; Shamdani, S.; Uzan, G.; Naserian, S.; Azarpira, N. Different approaches for transformation of mesenchymal stem cells into hepatocyte-like cells. Stem Cell Res. Ther. 2020, 11, 54. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Yuan, L.; An, Z.; Shi, D.; Xin, J.; Jiang, J.; Ren, K.; Chen, J.; Guo, B.; Zhou, X.; et al. DLL4 restores damaged liver by enhancing hBMSC differentiation into cholangiocytes. Stem Cell Res. 2020, 47, 101900. [Google Scholar] [CrossRef] [PubMed]
- Iwanaka, T.; Yamaza, T.; Sonoda, S.; Yoshimaru, K.; Matsuura, T.; Yamaza, H.; Ohga, S.; Oda, Y.; Taguchi, T. A model study for the manufacture and validation of clinical-grade deciduous dental pulp stem cells for chronic liver fibrosis treatment. Stem Cell Res. Ther. 2020, 11, 134. [Google Scholar] [CrossRef]
- Lee, J.; Choi, J.; Kang, S.; Kim, J.; Lee, R.; So, S.; Yoon, Y.I.; Kirchner, V.A.; Song, G.W.; Hwang, S.; et al. Hepatogenic Potential and Liver Regeneration Effect of Human Liver-derived Mesenchymal-Like Stem Cells. Cells 2020, 9, 1521. [Google Scholar] [CrossRef]
- Li, T.; Xia, M.; Gao, Y.; Chen, Y.; Xu, Y. Human umbilical cord mesenchymal stem cells: An overview of their potential in cell-based therapy. Expert Opin Biol. Ther. 2015, 15, 1293–1306. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Wu, Z.; Li, L. Mesenchymal stromal cells promote liver regeneration through regulation of immune cells. Int. J. Biol. Sci. 2020, 16, 893–903. [Google Scholar] [CrossRef]
- Kang, S.H.; Kim, M.Y.; Eom, Y.W.; Baik, S.K. Mesenchymal Stem Cells for the Treatment of Liver Disease: Present and Perspectives. Gut Liver 2020, 14, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, A.; Takeuchi, S.; Watanabe, T.; Yoshida, T.; Nojiri, S.; Ogawa, M.; Terai, S. Mesenchymal stem cell therapies for liver cirrhosis: MSCs as "conducting cells" for improvement of liver fibrosis and regeneration. Inflamm. Regen. 2019, 39, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eom, Y.W.; Shim, K.Y.; Baik, S.K. Mesenchymal stem cell therapy for liver fibrosis. Korean J. Intern. Med. 2015, 30, 580–589. [Google Scholar] [CrossRef]
- Shi, M.; Zhang, Z.; Xu, R.; Lin, H.; Fu, J.; Zou, Z.; Zhang, A.; Shi, J.; Chen, L.; Lv, S.; et al. Human mesenchymal stem cell transfusion is safe and improves liver function in acute-on-chronic liver failure patients. Stem Cells Transl. Med. 2012, 1, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, J.; Liu, H.; Li, Y.; Fu, J.; Sun, Y.; Xu, R.; Lin, H.; Wang, S.; Lv, S.; et al. Pilot study of umbilical cord-derived mesenchymal stem cell transfusion in patients with primary biliary cirrhosis. J. Gastroenterol. Hepatol. 2013, 28 (Suppl. S1), 85–92. [Google Scholar] [CrossRef] [PubMed]
- Lou, G.; Chen, Z.; Zheng, M.; Liu, Y. Mesenchymal stem cell-derived exosomes as a new therapeutic strategy for liver diseases. Exp. Mol. Med. 2017, 49, e346. [Google Scholar] [CrossRef]
- Forbes, S.J.; Gupta, S.; Dhawan, A. Cell therapy for liver disease: From liver transplantation to cell factory. J. Hepatol. 2015, 62, S157–S169. [Google Scholar] [CrossRef] [Green Version]
- Puppi, J.; Tan, N.; Mitry, R.R.; Hughes, R.D.; Lehec, S.; Mieli-Vergani, G.; Karani, J.; Champion, M.P.; Heaton, N.; Mohamed, R.; et al. Hepatocyte transplantation followed by auxiliary liver transplantation--a novel treatment for ornithine transcarbamylase deficiency. Am. J. Transplant. 2008, 8, 452–457. [Google Scholar] [CrossRef]
- Soltys, K.A.; Setoyama, K.; Tafaleng, E.N.; Soto Gutierrez, A.; Fong, J.; Fukumitsu, K.; Nishikawa, T.; Nagaya, M.; Sada, R.; Haberman, K.; et al. Host conditioning and rejection monitoring in hepatocyte transplantation in humans. J. Hepatol. 2017, 66, 987–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyburg, J.; Das, A.M.; Hoerster, F.; Lindner, M.; Kriegbaum, H.; Engelmann, G.; Schmidt, J.; Ott, M.; Pettenazzo, A.; Luecke, T.; et al. One liver for four children: First clinical series of liver cell transplantation for severe neonatal urea cycle defects. Transplantation 2009, 87, 636–641. [Google Scholar] [CrossRef]
- Stephenne, X.; Najimi, M.; Sibille, C.; Nassogne, M.C.; Smets, F.; Sokal, E.M. Sustained engraftment and tissue enzyme activity after liver cell transplantation for argininosuccinate lyase deficiency. Gastroenterology 2006, 130, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Meyburg, J.; Opladen, T.; Spiekerkotter, U.; Schlune, A.; Schenk, J.P.; Schmidt, J.; Weitz, J.; Okun, J.; Burger, F.; Omran, T.B.; et al. Human heterologous liver cells transiently improve hyperammonemia and ureagenesis in individuals with severe urea cycle disorders. J. Inherit. Metab. Dis. 2018, 41, 81–90. [Google Scholar] [CrossRef]
- Ambrosino, G.; Varotto, S.; Strom, S.C.; Guariso, G.; Franchin, E.; Miotto, D.; Caenazzo, L.; Basso, S.; Carraro, P.; Valente, M.L.; et al. Isolated Hepatocyte Transplantation for Crigler-Najjar Syndrome Type 1. Cell Transplant. 2005, 14, 151–157. [Google Scholar] [CrossRef]
- Fox, I.J.; Chowdhury, J.R.; Kaufman, S.S.; Goertzen, T.C.; Chowdhury, N.R.; Warkentin, P.I.; Dorko, K.; Sauter, B.V.; Strom, S.C. Treatment Of The Crigler-Najjar Syndrome Type I With Hepatocyte Transplantation. N. Engl. J. Med. 1998, 338, 1422–1426. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.J.; Mifsud, N.A.; Williamson, R.; Bertolino, P.; Hardikar, W. Cell-mediated rejection results in allograft loss after liver cell transplantation. Liver Transpl. 2008, 14, 688–694. [Google Scholar] [CrossRef]
- Hansel, M.C.; Gramignoli, R.; Skvorak, K.J.; Dorko, K.; Marongiu, F.; Blake, W.; Davila, J.; Strom, S.C. The history and use of human hepatocytes for the treatment of liver diseases: The first 100 patients. Curr. Protoc Toxicol. 2014, 62, 14.12.1-23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smets, F.; Dobbelaere, D.; McKiernan, P.; Dionisi-Vici, C.; Broue, P.; Jacquemin, E.; Lopes, A.I.; Goncalves, I.; Mandel, H.; Pawlowska, J.; et al. Phase I/II Trial of Liver-derived Mesenchymal Stem Cells in Pediatric Liver-based Metabolic Disorders: A Prospective, Open Label, Multicenter, Partially Randomized, Safety Study of One Cycle of Heterologous Human Adult Liver-derived Progenitor Cells (HepaStem) in Urea Cycle Disorders and Crigler-Najjar Syndrome Patients. Transplantation 2019, 103, 1903–1915. [Google Scholar] [PubMed]
- Lee, S.M.; Lee, S.D.; Wang, S.Z.; Sarkar, D.; Lee, H.M.; Khan, A.; Bhati, C.; Sharma, A.; Kumaran, V.; Bruno, D.; et al. Effect of mesenchymal stem cell in liver regeneration and clinical applications. Hepatoma Res. 2021, 7, 53. [Google Scholar] [CrossRef]
- Norris, S.; Collins, C.; Doherty, D.G.; Smith, F.; McEntee, G.; Traynor, O.; Nolan, N.; Hegarty, J.; O’Farrelly, C. Resident human hepatic lymphocytes are phenotypically different from circulating lymphocytes. J. Hepatol. 1998, 28, 84–90. [Google Scholar] [CrossRef]
- Oo, Y.H.; Sakaguchi, S. Regulatory T-cell directed therapies in liver diseases. J. Hepatol. 2013, 59, 1127–1134. [Google Scholar] [CrossRef] [Green Version]
- Mieli-Vergani, G.; Vergani, D. Autoimmune hepatitis. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.M.R.; Muller, Y.D.; Bluestone, J.A.; Tang, Q. Next-generation regulatory T cell therapy. Nat. Rev. Drug Discov. 2019, 18, 749–769. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Liu, Z.; Li, C.; Wei, Q.; Zheng, S.; Saeb-Parsy, K.; Xu, X. Regulatory T Cell Therapy Following Liver Transplantation. Liver Transpl. 2021, 27, 264–280. [Google Scholar] [CrossRef]
- Longhi, M.S.; Ma, Y.; Grant, C.R.; Samyn, M.; Gordon, P.; Mieli-Vergani, G.; Vergani, D. T-regs in autoimmune hepatitis-systemic lupus erythematosus/mixed connective tissue disease overlap syndrome are functionally defective and display a Th1 cytokine profile. J. Autoimmun. 2013, 41, 146–151. [Google Scholar] [CrossRef]
- Longhi, M.S.; Ma, Y.; Bogdanos, D.P.; Cheeseman, P.; Mieli-Vergani, G.; Vergani, D. Impairment of CD4(+)CD25(+) regulatory T-cells in autoimmune liver disease. J. Hepatol. 2004, 41, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Curbishley, S.M.; Eksteen, B.; Gladue, R.P.; Lalor, P.; Adams, D.H. CXCR 3 activation promotes lymphocyte transendothelial migration across human hepatic endothelium under fluid flow. Am. J. Pathol. 2005, 167, 887–899. [Google Scholar] [CrossRef] [Green Version]
- Létourneau, S.; van Leeuwen, E.M.; Krieg, C.; Martin, C.; Pantaleo, G.; Sprent, J.; Surh, C.D.; Boyman, O. IL-2/anti-IL-2 antibody complexes show strong biological activity by avoiding interaction with IL-2 receptor alpha subunit CD25. Proc. Natl. Acad. Sci. USA 2010, 107, 2171–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stross, L.; Günther, J.; Gasteiger, G.; Asen, T.; Graf, S.; Aichler, M.; Esposito, I.; Busch, D.H.; Knolle, P.; Sparwasser, T.; et al. Foxp3+ regulatory T cells protect the liver from immune damage and compromise virus control during acute experimental hepatitis B virus infection in mice. Hepatology 2012, 56, 873–883. [Google Scholar] [CrossRef]
- Safinia, N.; Grageda, N.; Scottà, C.; Thirkell, S.; Fry, L.J.; Vaikunthanathan, T.; Lechler, R.I.; Lombardi, G. Cell Therapy in Organ Transplantation: Our Experience on the Clinical Translation of Regulatory T Cells. Front. Immunol. 2018, 9, 354. [Google Scholar] [CrossRef]
- Fu, J.; Xu, D.; Liu, Z.; Shi, M.; Zhao, P.; Fu, B.; Zhang, Z.; Yang, H.; Zhang, H.; Zhou, C.; et al. Increased regulatory T cells correlate with CD8 T-cell impairment and poor survival in hepatocellular carcinoma patients. Gastroenterology 2007, 132, 2328–2339. [Google Scholar] [CrossRef]
- Rech, A.J.; Mick, R.; Martin, S.; Recio, A.; Aqui, N.A.; Powell, D.J.; Colligon, T.A.; Trosko, J.A.; Leinbach, L.I.; Pletcher, C.H.; et al. CD25 blockade depletes and selectively reprograms regulatory T cells in concert with immunotherapy in cancer patients. Sci. Transl. Med. 2012, 4, 134ra162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutmuller, R.P.; van Duivenvoorde, L.M.; van Elsas, A.; Schumacher, T.N.; Wildenberg, M.E.; Allison, J.P.; Toes, R.E.; Offringa, R.; Melief, C.J. Synergism of cytotoxic T lymphocyte-associated antigen 4 blockade and depletion of CD25(+) regulatory T cells in antitumor therapy reveals alternative pathways for suppression of autoreactive cytotoxic T lymphocyte responses. J. Exp. Med. 2001, 194, 823–832. [Google Scholar] [CrossRef] [Green Version]
- Dal Bo, M.; De Mattia, E.; Baboci, L.; Mezzalira, S.; Cecchin, E.; Assaraf, Y.G.; Toffoli, G. New insights into the pharmacological, immunological, and CAR-T-cell approaches in the treatment of hepatocellular carcinoma. Drug Resist. Update 2020, 51, 100702. [Google Scholar] [CrossRef]
- Wang, Z.; Wu, Z.; Liu, Y.; Han, W. New development in CAR-T cell therapy. J. Hematol. Oncol. 2017, 10, 53. [Google Scholar] [CrossRef] [Green Version]
- Burga, R.A.; Thorn, M.; Point, G.R.; Guha, P.; Nguyen, C.T.; Licata, L.A.; DeMatteo, R.P.; Ayala, A.; Joseph Espat, N.; Junghans, R.P.; et al. Liver myeloid-derived suppressor cells expand in response to liver metastases in mice and inhibit the anti-tumor efficacy of anti-CEA CAR-T. Cancer Immunol. Immunother. 2015, 64, 817–829. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Li, K.; Tu, H.; Pan, X.; Jiang, H.; Shi, B.; Kong, J.; Wang, H.; Yang, S.; Gu, J.; et al. Development of T cells redirected to glypican-3 for the treatment of hepatocellular carcinoma. Clin. Cancer Res. 2014, 20, 6418–6428. [Google Scholar] [CrossRef] [Green Version]
- Thomson, A.W.; Ezzelarab, M.B. Regulatory dendritic cells: Profiling, targeting, and therapeutic application. Curr. Opin. Organ. Transplant. 2018, 23, 538–545. [Google Scholar] [CrossRef]
- Morelli, A.E.; Thomson, A.W. Orchestration of transplantation tolerance by regulatory dendritic cell therapy or in-situ targeting of dendritic cells. Curr. Opin Organ. Transplant. 2014, 19, 348–356. [Google Scholar] [CrossRef] [Green Version]
- Thomson, A.W.; Humar, A.; Lakkis, F.G.; Metes, D.M. Regulatory dendritic cells for promotion of liver transplant operational tolerance: Rationale for a clinical trial and accompanying mechanistic studies. Hum. Immunol. 2018, 79, 314–321. [Google Scholar] [CrossRef] [PubMed]
- van der Heide, D.; Weiskirchen, R.; Bansal, R. Therapeutic Targeting of Hepatic Macrophages for the Treatment of Liver Diseases. Front. Immunol. 2019, 10, 2852. [Google Scholar] [CrossRef] [Green Version]
- Tacke, F.; Zimmermann, H.W. Macrophage heterogeneity in liver injury and fibrosis. J. Hepatol. 2014, 60, 1090–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triantafyllou, E.; Woollard, K.J.; McPhail, M.J.W.; Antoniades, C.G.; Possamai, L.A. The Role of Monocytes and Macrophages in Acute and Acute-on-Chronic Liver Failure. Front. Immunol. 2018, 9, 2948. [Google Scholar] [CrossRef]
- Loomba, R.; Lawitz, E.; Mantry, P.S.; Jayakumar, S.; Caldwell, S.H.; Arnold, H.; Diehl, A.M.; Djedjos, C.S.; Han, L.; Myers, R.P.; et al. The ASK1 inhibitor selonsertib in patients with nonalcoholic steatohepatitis: A randomized, phase 2 trial. Hepatology 2018, 67, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Pedroza, M.; To, S.; Smith, J.; Agarwal, S.K. Cadherin-11 contributes to liver fibrosis induced by carbon tetrachloride. PLoS ONE 2019, 14, e0218971. [Google Scholar] [CrossRef] [Green Version]
- Maradana, M.R.; Yekollu, S.K.; Zeng, B.; Ellis, J.; Clouston, A.; Miller, G.; Talekar, M.; Bhuyan, Z.A.; Mahadevaiah, S.; Powell, E.E.; et al. Immunomodulatory liposomes targeting liver macrophages arrest progression of nonalcoholic steatohepatitis. Metabolism 2018, 78, 80–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochigneux, P.; Chanez, B.; De Rauglaudre, B.; Mitry, E.; Chabannon, C.; Gilabert, M. Adoptive Cell Therapy in Hepatocellular Carcinoma: Biological Rationale and First Results in Early Phase Clinical Trials. Cancers 2021, 13, 271. [Google Scholar] [CrossRef] [PubMed]
- Siefert, J.; Hillebrandt, K.H.; Moosburner, S.; Podrabsky, P.; Geisel, D.; Denecke, T.; Unger, J.K.; Sawitzki, B.; Gul-Klein, S.; Lippert, S.; et al. Hepatocyte Transplantation to the Liver via the Splenic Artery in a Juvenile Large Animal Model. Cell Transplant. 2019, 28, 14S–24S. [Google Scholar] [CrossRef] [PubMed]
- Ridola, L.; Bragazzi, M.C.; Cardinale, V.; Carpino, G.; Gaudio, E.; Alvaro, D. Cholangiocytes: Cell transplantation. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 1516–1523. [Google Scholar] [CrossRef] [PubMed]
- Nevi, L.; Carpino, G.; Costantini, D.; Cardinale, V.; Riccioni, O.; Di Matteo, S.; Melandro, F.; Berloco, P.B.; Reid, L.; Gaudio, E.; et al. Hyaluronan coating improves liver engraftment of transplanted human biliary tree stem/progenitor cells. Stem Cell Res. Ther. 2017, 8, 68. [Google Scholar] [CrossRef] [PubMed]
- Kuijk, E.W.; Rasmussen, S.; Blokzijl, F.; Huch, M.; Gehart, H.; Toonen, P.; Begthel, H.; Clevers, H.; Geurts, A.M.; Cuppen, E. Generation and characterization of rat liver stem cell lines and their engraftment in a rat model of liver failure. Sci. Rep. 2016, 6, 22154. [Google Scholar] [CrossRef]
- Sampaziotis, F.; Justin, A.W.; Tysoe, O.C.; Sawiak, S.; Godfrey, E.M.; Upponi, S.S.; Gieseck, R.L., 3rd; de Brito, M.C.; Berntsen, N.L.; Gomez-Vazquez, M.J.; et al. Reconstruction of the mouse extrahepatic biliary tree using primary human extrahepatic cholangiocyte organoids. Nat. Med. 2017, 23, 954–963. [Google Scholar] [CrossRef]
- Yang, L.; Cao, H.; Sun, D.; Hou, B.; Lin, L.; Shen, Z.Y.; Song, H.L. Bone marrow mesenchymal stem cells combine with normothermic machine perfusion to improve rat donor liver quality-the important role of hepatic microcirculation in donation after circulatory death. Cell Tissue Res. 2020, 381, 239–254. [Google Scholar] [CrossRef] [PubMed]
- Rigo, F.; De Stefano, N.; Navarro-Tableros, V.; David, E.; Rizza, G.; Catalano, G.; Gilbo, N.; Maione, F.; Gonella, F.; Roggio, D.; et al. Extracellular Vesicles from Human Liver Stem Cells Reduce Injury in an Ex Vivo Normothermic Hypoxic Rat Liver Perfusion Model. Transplantation 2018, 102, e205–e210. [Google Scholar] [CrossRef]
- Laing, R.W.; Stubblefield, S.; Wallace, L.; Roobrouck, V.D.; Bhogal, R.H.; Schlegel, A.; Boteon, Y.L.; Reynolds, G.M.; Ting, A.E.; Mirza, D.F.; et al. The Delivery of Multipotent Adult Progenitor Cells to Extended Criteria Human Donor Livers Using Normothermic Machine Perfusion. Front. Immunol. 2020, 11, 1226. [Google Scholar] [CrossRef]
- Sampaziotis, F.; Muraro, D.; Tysoe, O.C.; Sawiak, S.; Beach, T.E.; Godfrey, E.M.; Upponi, S.S.; Brevini, T.; Wesley, B.T.; Garcia-Bernardo, J.; et al. Cholangiocyte organoids can repair bile ducts after transplantation in the human liver. Science 2021, 371, 839–846. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hofmann, J.; Hackl, V.; Esser, H.; Meszaros, A.T.; Fodor, M.; Öfner, D.; Troppmair, J.; Schneeberger, S.; Hautz, T. Cell-Based Regeneration and Treatment of Liver Diseases. Int. J. Mol. Sci. 2021, 22, 10276. https://doi.org/10.3390/ijms221910276
Hofmann J, Hackl V, Esser H, Meszaros AT, Fodor M, Öfner D, Troppmair J, Schneeberger S, Hautz T. Cell-Based Regeneration and Treatment of Liver Diseases. International Journal of Molecular Sciences. 2021; 22(19):10276. https://doi.org/10.3390/ijms221910276
Chicago/Turabian StyleHofmann, Julia, Verena Hackl, Hannah Esser, Andras T. Meszaros, Margot Fodor, Dietmar Öfner, Jakob Troppmair, Stefan Schneeberger, and Theresa Hautz. 2021. "Cell-Based Regeneration and Treatment of Liver Diseases" International Journal of Molecular Sciences 22, no. 19: 10276. https://doi.org/10.3390/ijms221910276
APA StyleHofmann, J., Hackl, V., Esser, H., Meszaros, A. T., Fodor, M., Öfner, D., Troppmair, J., Schneeberger, S., & Hautz, T. (2021). Cell-Based Regeneration and Treatment of Liver Diseases. International Journal of Molecular Sciences, 22(19), 10276. https://doi.org/10.3390/ijms221910276