
Changes in Stress-Mediated Markers in a Human Cardiomyocyte Cell Line under Hyperglycemia



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
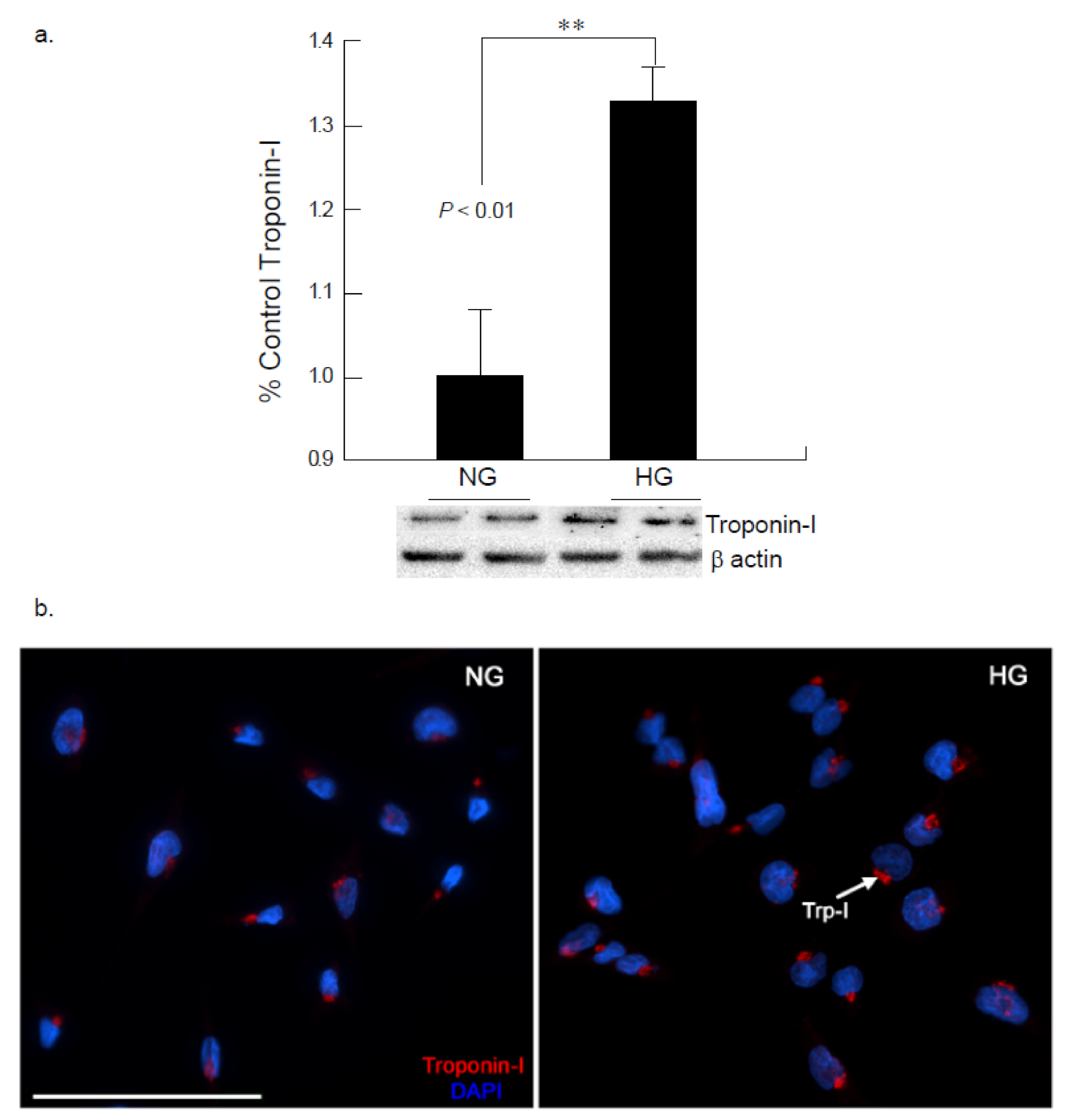
2.1. High Glucose Insult Results in Alterations in Connexin 43 and Troponin I Expression
2.2. Hyperglycemia Leads to Altered Signaling of HSP60
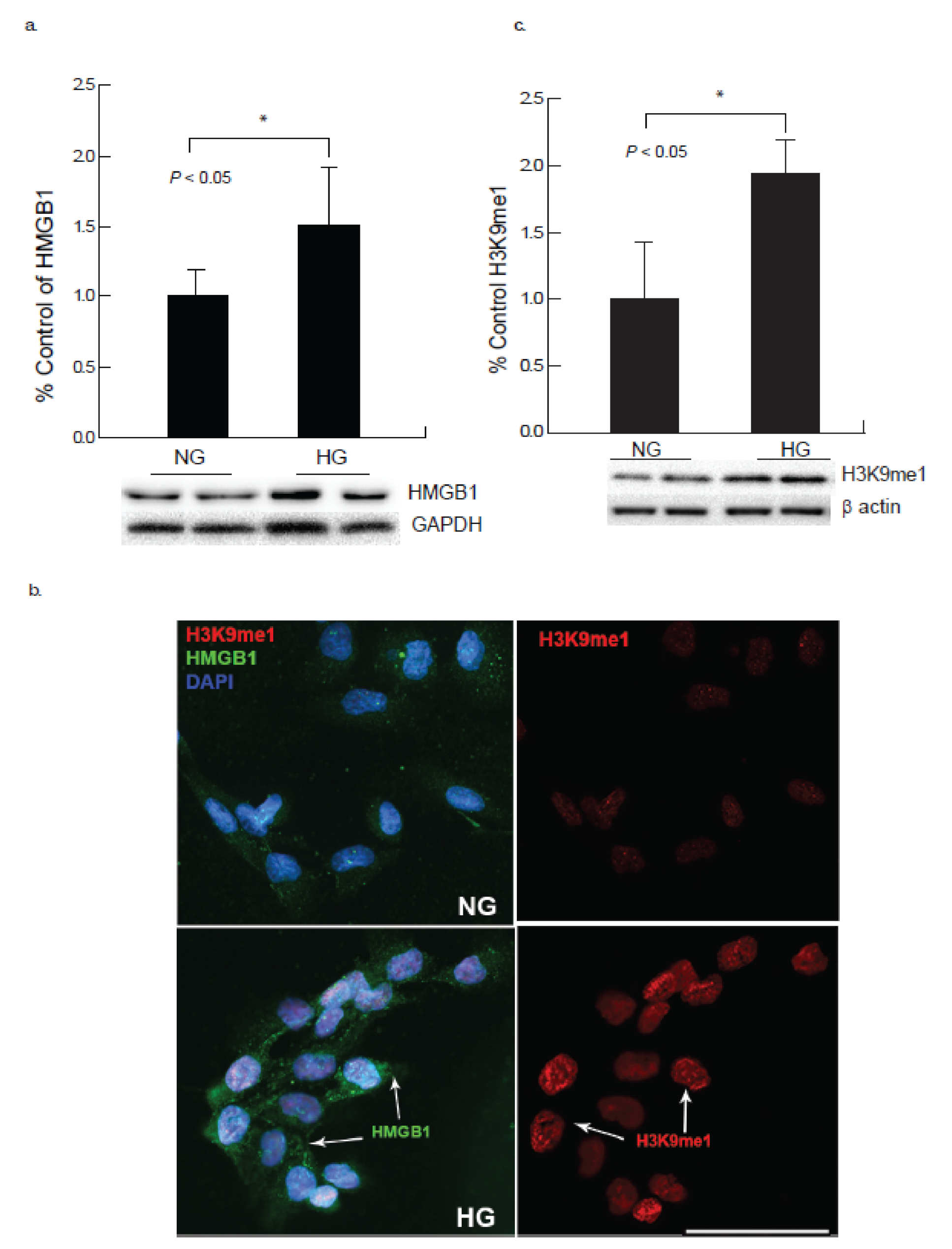
2.3. Hyperglycemia Induces Elevated Expression of HMGB1 and H3K9me1 in Cardiomyocyte
2.4. AC-16 Cells under Hyperglycemic Conditions Upregulates RAGE and TLR4 Expression
2.5. Hyperglycemia Modifies Chemokine Receptor CXCR4 Expression in Cardiomyocytes
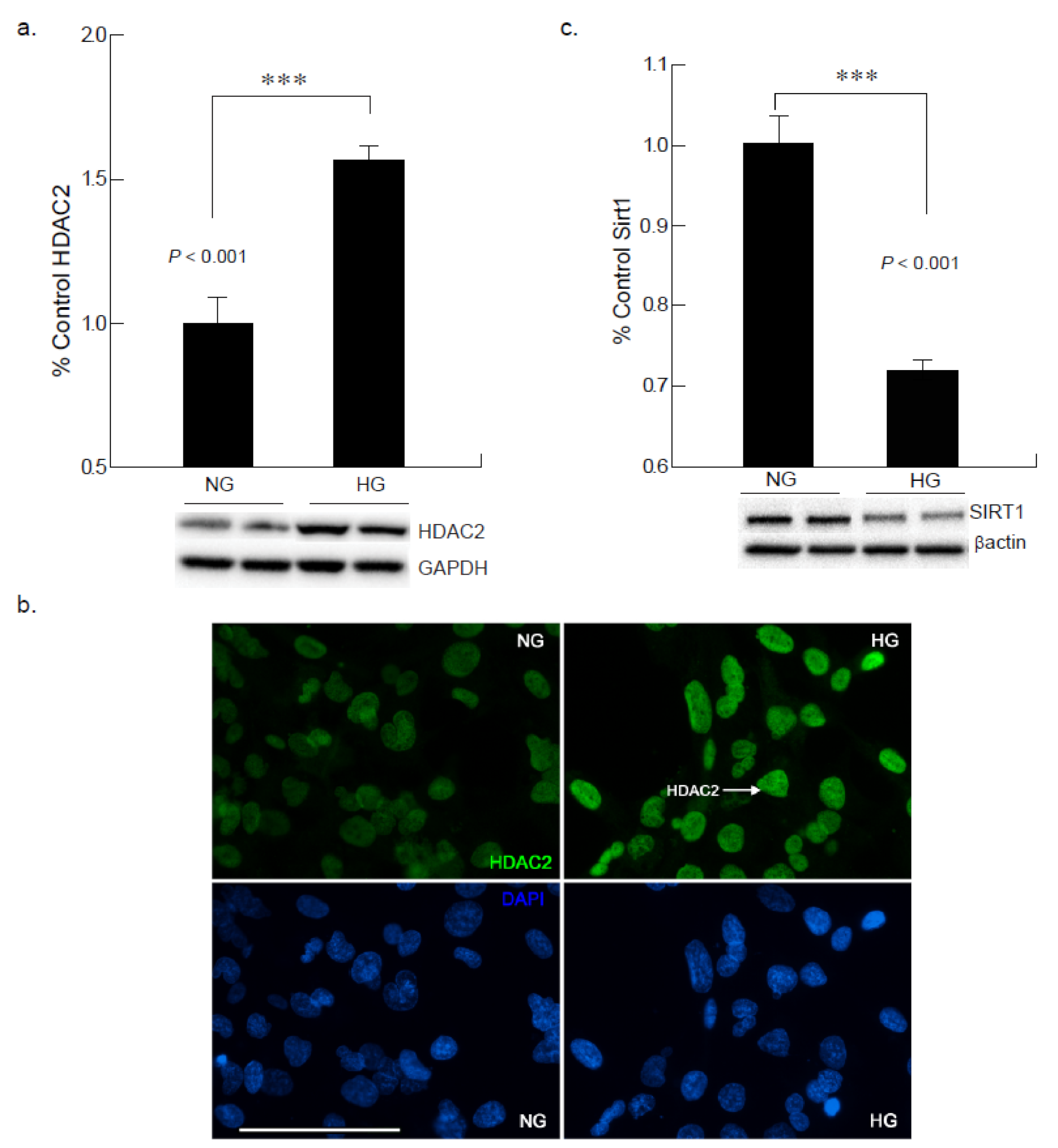
2.6. Hyperglycemia Leads to Increased HDAC2 Expression in Cardiomyocyte
2.7. Hyperglycemic Condition Downregulates Sirtuin-1 Expression in CM
3. Discussion
4. Materials and Methods
4.1. AC16 Human Cardiomyocyte Cell Line Culture
4.2. Protein Extraction and Western Blot Analysis
4.3. Immunocytochemistry
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CXCR4 | chemokine C-X-C motif receptor 4 |
| RAGE | receptor for advanced glycation end products |
| Toll-like receptor 4 | TLR4 |
| CX43 | Connexin-43 |
| Trp-I: | Troponin I |
| Sirtuin 1 | SIRT1 |
| Histone deacetylase 2 | HDAC2 |
| Heat shock protein 60 | HSP60 |
| Histone 3 lysine 9 mono-methylation | H3K9me1 |
| High mobility group box 1 | HMGB1 |
References
- Braunwald, E. Cardiomyopathies: An Overview. Circ. Res. 2017, 121, 711–721. [Google Scholar] [CrossRef]
- Zhou, P.; Pu, W.T. Recounting Cardiac Cellular Composition. Circ. Res. 2016, 118, 368–370. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Jackson, P.G.; Makan, J. Cardiac troponins. J. Clin. Pathol. 2004, 57, 1025–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, D.; Brownlee, M. Hyperglycemia-induced reactive oxygen species increase expression of the receptor for advanced glycation end products (RAGE) and RAGE ligands. Diabetes 2010, 59, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Tsan, M.F.; Gao, B. Endogenous ligands of Toll-like receptors. J. Leukoc. Biol. 2004, 76, 514–519. [Google Scholar] [CrossRef]
- Tobias, P.S.; Curtiss, L.K. Toll-like receptors in atherosclerosis. Biochem. Soc. Trans. 2007, 35, 1453–1455. [Google Scholar] [CrossRef]
- Willis, M.S.; Patterson, C. Hold me tight: Role of the heat shock protein family of chaperones in cardiac disease. Circulation 2010, 122, 1740–1751. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.S.; Shan, Y.X.; Yang, T.L.; Lin, H.D.; Chen, J.W.; Lin, S.J.; Wang, P.H. Insulin deficiency downregulated heat shock protein 60 and IGF-1 receptor signaling in diabetic myocardium. Diabetes 2005, 54, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Schafler, A.E.; Kirmanoglou, K.; Balbach, J.; Pecher, P.; Hannekum, A.; Schumacher, B. The expression of heat shock protein 60 in myocardium of patients with chronic atrial fibrillation. Basic Res. Cardiol. 2002, 97, 258–261. [Google Scholar] [CrossRef]
- Yerra, V.G.; Advani, A. Histones and heart failure in diabetes. Cell. Mol. Life Sci. 2018, 75, 3193–3213. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.Y.; Geng, Y.J.; Liang, J.L.; Lin, Q.X.; Lin, S.G.; Zhang, S.; Li, Y. High levels of glucose induce apoptosis in cardiomyocyte via epigenetic regulation of the insulin-like growth factor receptor. Exp. Cell Res. 2010, 316, 2903–2909. [Google Scholar] [CrossRef]
- Weeks, K.L.; Avkiran, M. Roles and post-translational regulation of cardiac class IIa histone deacetylase isoforms. J. Physiol. 2015, 593, 1785–1797. [Google Scholar] [CrossRef] [Green Version]
- Boutant, M.; Canto, C. SIRT1 metabolic actions: Integrating recent advances from mouse models. Mol. Metab. 2014, 3, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Canto, C.; Gerhart-Hines, Z.; Feige, J.N.; Lagouge, M.; Noriega, L.; Milne, J.C.; Elliott, P.J.; Puigserver, P.; Auwerx, J. AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity. Nature 2009, 458, 1056–1060. [Google Scholar] [CrossRef]
- Canto, C.; Auwerx, J. Interference between PARPs and SIRT1: A novel approach to healthy ageing? Aging 2011, 3, 543–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheu, J.J.; Chang, L.T.; Chiang, C.H.; Sun, C.K.; Chang, N.K.; Youssef, A.A.; Wu, C.J.; Lee, F.Y.; Yip, H.K. Impact of diabetes on cardiomyocyte apoptosis and connexin43 gap junction integrity: Role of pharmacological modulation. Int. Heart J. 2007, 48, 233–245. [Google Scholar] [CrossRef] [Green Version]
- Tirziu, D.; Giordano, F.J.; Simons, M. Cell communications in the heart. Circulation 2010, 122, 928–937. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Ye, P.; Luo, L.; Xiao, W.; Xu, R.; Wu, H. Association between blood glucose levels and high-sensitivity cardiac troponin T in an overt cardiovascular disease-free community-based study. Diabetes Res. Clin. Pract. 2012, 97, 139–145. [Google Scholar] [CrossRef]
- Rubin, J.; Matsushita, K.; Ballantyne, C.M.; Hoogeveen, R.; Coresh, J.; Selvin, E. Chronic hyperglycemia and subclinical myocardial injury. J. Am. Coll. Cardiol. 2012, 59, 484–489. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Guan, X.; Zuo, X.; Wang, J.; Yin, W. The role of high mobility group box 1 (HMGB1) in the pathogenesis of kidney diseases. Acta Pharm. Sin. B 2016, 6, 183–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, D.P.; Dahllof, M.; Lundh, M.; Rasmussen, D.N.; Nielsen, M.D.; Billestrup, N.; Grunnet, L.G.; Mandrup-Poulsen, T. Histone deacetylase (HDAC) inhibition as a novel treatment for diabetes mellitus. Mol. Med. 2011, 17, 378–390. [Google Scholar] [CrossRef]
- Ng, F.; Tang, B.L. When is Sirt1 activity bad for dying neurons? Front. Cell. Neurosci. 2013, 7, 186. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Wang, J.; Zhao, A.; Li, J. SIRT1 activation inhibits hyperglycemia-induced apoptosis by reducing oxidative stress and mitochondrial dysfunction in human endothelial cells. Mol. Med. Rep. 2017, 16, 3331–3338. [Google Scholar] [CrossRef] [Green Version]
- Liyanagamage, D.; Martinus, R.D. Role of Mitochondrial Stress Protein HSP60 in Diabetes-Induced Neuroinflammation. Mediators Inflamm. 2020, 2020, 8073516. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.J.; Wang, S.; Wang, B.J.; Xiao, M.; Guo, Y.; Tang, Y.; Zhang, J.; Gu, J. Epigenetic Regulation Associated With Sirtuin 1 in Complications of Diabetes Mellitus. Front. Endocrinol. 2020, 11, 598012. [Google Scholar] [CrossRef]
- Hussain, S.; Khan, A.W.; Akhmedov, A.; Suades, R.; Costantino, S.; Paneni, F.; Caidahl, K.; Mohammed, S.A.; Hage, C.; Gkolfos, C.; et al. Hyperglycemia Induces Myocardial Dysfunction via Epigenetic Regulation of JunD. Circ. Res. 2020, 127, 1261–1273. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Chilian, W.M.; Dong, F. Epigenetic regulation in diabetes-associated oxidative stress and myocardial dysfunction. Int. J. Cardiol. 2018, 268, 193–194. [Google Scholar] [CrossRef] [PubMed]
- Davidson, M.M.; Nesti, C.; Palenzuela, L.; Walker, W.F.; Hernandez, E.; Protas, L.; Hirano, M.; Isaac, N.D. Novel cell lines derived from adult human ventricular cardiomyocytes. J. Mol. Cell Cardiol. 2005, 39, 133–147. [Google Scholar] [CrossRef]
- Chattopadhyay, M.; Mata, M.; Fink, D.J. Continuous delta-opioid receptor activation reduces neuronal voltage-gated sodium channel (NaV1.7) levels through activation of protein kinase C in painful diabetic neuropathy. J. Neurosci. 2008, 28, 6652–6658. [Google Scholar] [CrossRef]
- Thakur, V.; Alcoreza, N.; Delgado, M.; Joddar, B.; Chattopadhyay, M. Cardioprotective Effect of Glycyrrhizin on Myocardial Remodeling in Diabetic Rats. Biomolecules 2021, 11, 569. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thakur, V.; Alcoreza, N.; Cazares, J.; Chattopadhyay, M. Changes in Stress-Mediated Markers in a Human Cardiomyocyte Cell Line under Hyperglycemia. Int. J. Mol. Sci. 2021, 22, 10802. https://doi.org/10.3390/ijms221910802
Thakur V, Alcoreza N, Cazares J, Chattopadhyay M. Changes in Stress-Mediated Markers in a Human Cardiomyocyte Cell Line under Hyperglycemia. International Journal of Molecular Sciences. 2021; 22(19):10802. https://doi.org/10.3390/ijms221910802
Chicago/Turabian StyleThakur, Vikram, Narah Alcoreza, Jasmine Cazares, and Munmun Chattopadhyay. 2021. "Changes in Stress-Mediated Markers in a Human Cardiomyocyte Cell Line under Hyperglycemia" International Journal of Molecular Sciences 22, no. 19: 10802. https://doi.org/10.3390/ijms221910802
APA StyleThakur, V., Alcoreza, N., Cazares, J., & Chattopadhyay, M. (2021). Changes in Stress-Mediated Markers in a Human Cardiomyocyte Cell Line under Hyperglycemia. International Journal of Molecular Sciences, 22(19), 10802. https://doi.org/10.3390/ijms221910802