Candidate Effectors from Botryosphaeria dothidea Suppress Plant Immunity and Contribute to Virulence

 and
and 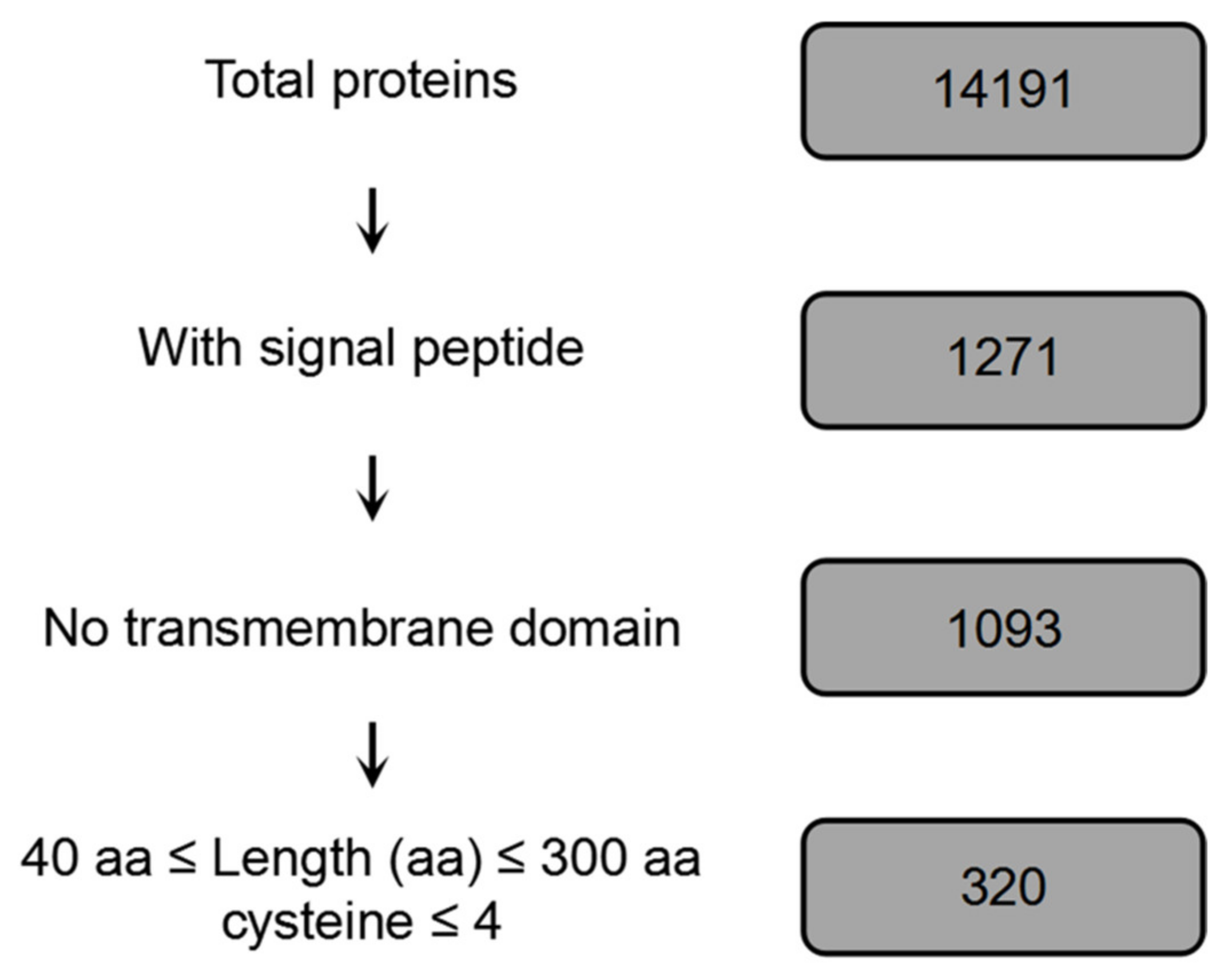{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Bioinformatic Prediction of the B. dothidea Effectome
2.2. Suppression of BT-PCD by a Majority of B. dothidea Candidate Effectors
2.3. Functional Validation of Predicted SPs of Candidate Effectors
2.4. Candidate Effector Suppression of PCD Triggered by Different Elicitors
3. Discussion
4. Materials and Methods
4.1. Fungi and Plants
4.2. Bioinformatic Analysis and Candidate Effector Prediction
4.3. Construction of A. tumefaciens Binary PVX Vectors
4.4. Cell Death Induction/Suppression Assay in N. benthamiana
4.5. Western Blotting
4.6. Validation of the SP Secretory Activities of Candidate Effectors
4.7. Generation of Gene Deletion Transformants
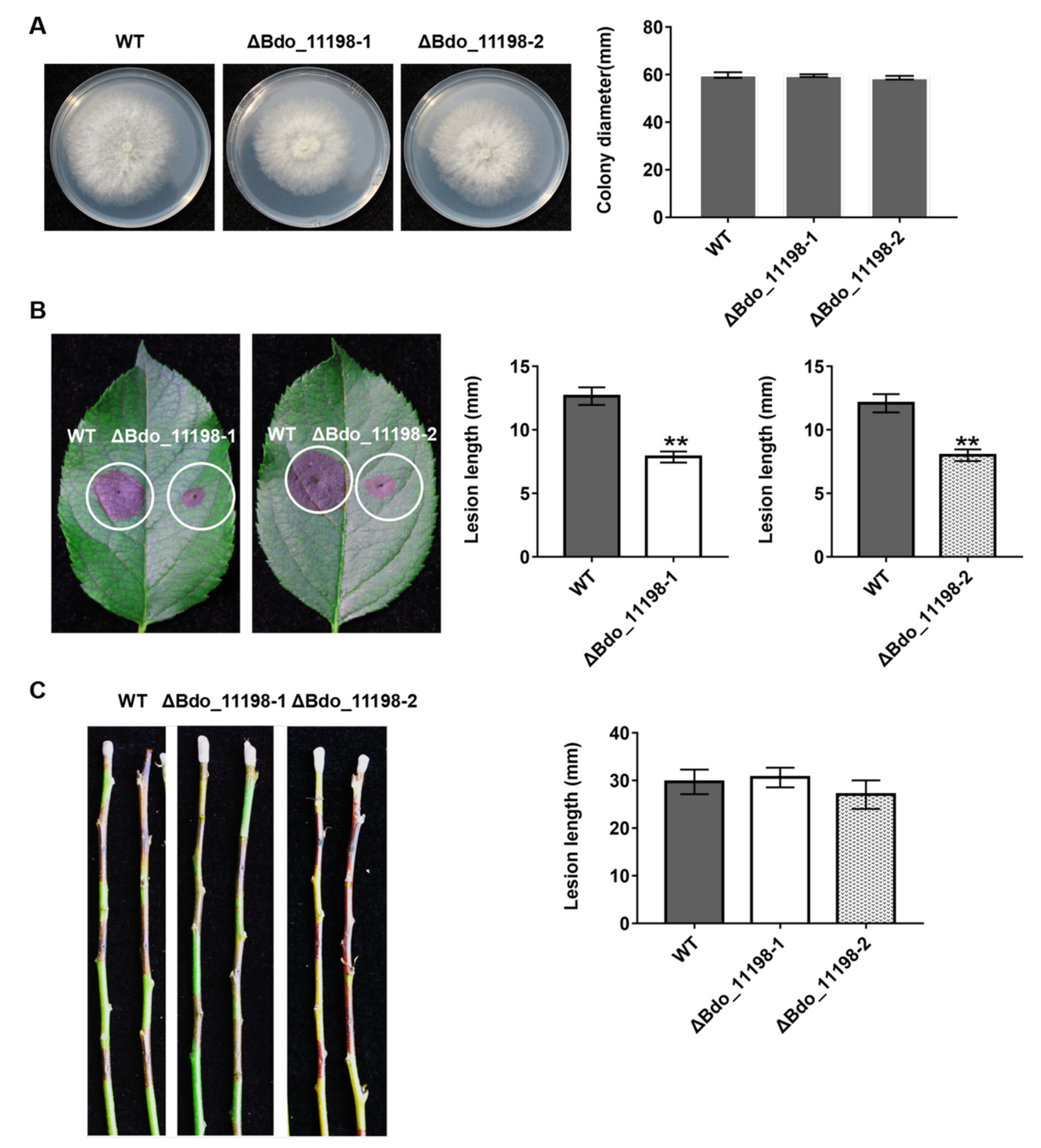
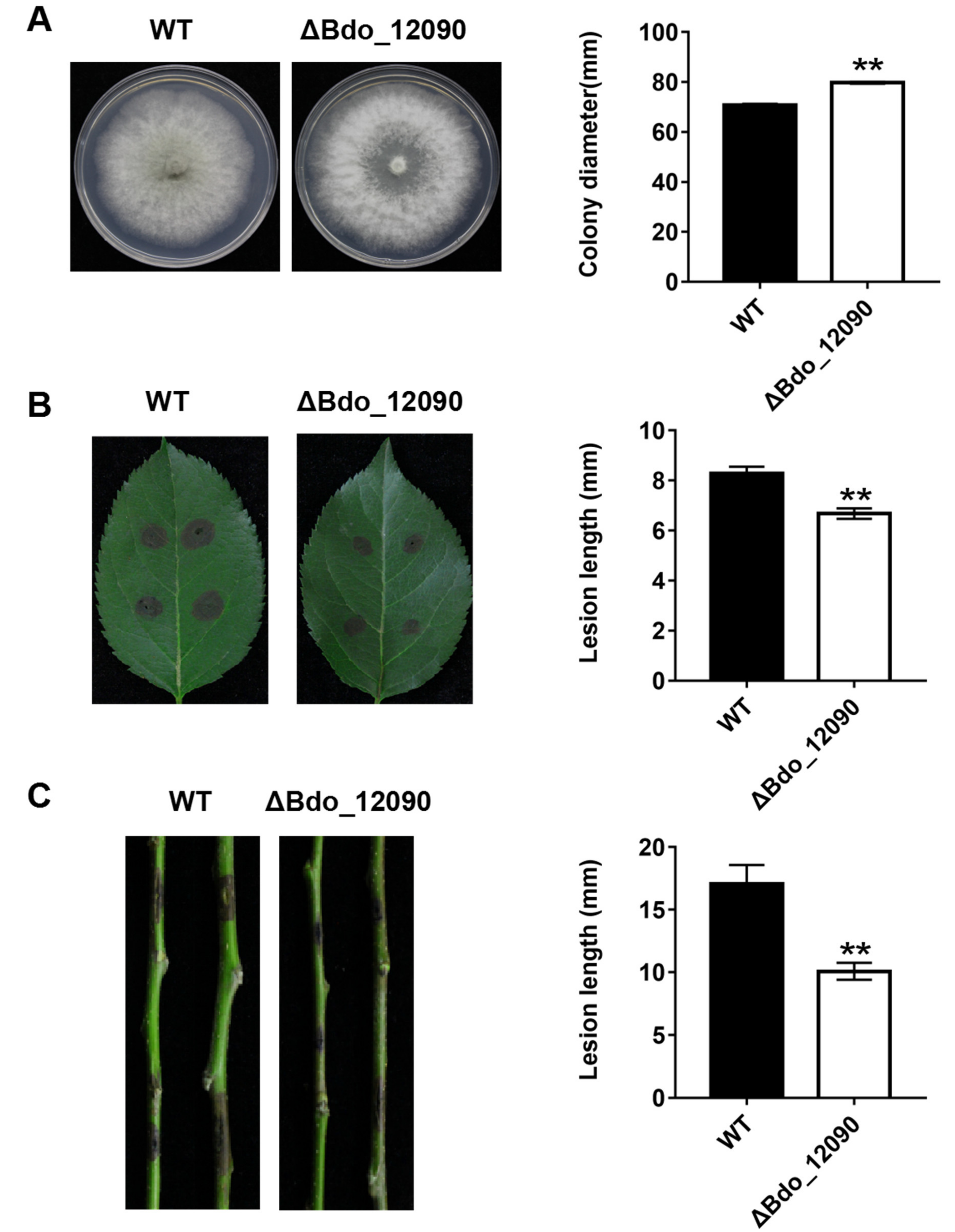
4.8. Morphology and Pathogenicity Assay
4.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marsberg, A.; Kemler, M.; Jami, F.; Nagel, J.H.; Postma-Smidt, A.; Naidoo, S.; Wingfield, M.J.; Crous, P.W.; Spatafora, J.W.; Hesse, C.N.; et al. Botryosphaeria dothidea: A latent pathogen of global importance to woody plant health. Mol. Plant Pathol. 2017, 18, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Michailides, T.J. Pathogenicity, distribution, sources of inoculum, and infection courts of Botryosphaeria dothidea on pistachio. Phytopathology 1991, 81, 566–573. [Google Scholar] [CrossRef]
- Van Niekerk, J.M.; Fourie, P.H.; Halleen, F.; Crous, P.W. Botryosphaeria spp. as grapevine trunk disease pathogens. Phytopathol. Mediterr. 2006, 45, S43–S54. [Google Scholar]
- Tang, W.; Ding, Z.; Zhou, Z.Q.; Wang, Y.Z.; Guo, L.Y. Phylogenetic and pathogenic analyses show that the causal agent of apple ring rot in China is Botryosphaeria Dothidea. Plant Dis. 2012, 96, 486–496. [Google Scholar] [CrossRef] [Green Version]
- Nazerian, E.; Mirabolfathy, M.; Ashnaei, S.P.; Beiki, F. Characterization of Botryosphaeria dothidea as new pathogen of kiwifruit in Iran. J. Plant Prot. Res. 2019, 59. [Google Scholar] [CrossRef]
- Guo, L.; Li, J.; Li, B.; Zhang, X.; Zhou, Z.; Li, G.; Wang, Y.; Li, X.; Huang, L.; Sun, G.; et al. Investigations on the occurrence and chemical control of Botryosphaeria canker of apple in China. Plant Prot. 2009, 35, 120–123. (In Chinese) [Google Scholar]
- Ren, J.; Wang, S.; Hu, T.; Wang, Y.; Cao, K. Evaluation of apple yield loss caused by apple ring rot. J. Henan Agric. Sci. 2014, 43, 90–92. [Google Scholar]
- Wang, B.; Liang, X.; Gleason, M.L.; Zhang, R.; Sun, G. Comparative genomics of Botryosphaeria dothidea and B. kuwatsukai, causal agents of apple ring rot, reveals both species expansion of pathogenicity-related genes and variations in virulence gene content during speciation. IMA Fungus 2018, 9, 243–257. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Zhao, W.S.; Chen, Z.; Xing, Q.K.; Hang, Q.G.; Chethana, K.W.T.; Xue, M.F.; Xu, J.P.; Phillips, A.J.L.; Wang, Y.; et al. Comparative genome and transcriptome analyses reveal adaptations to opportunistic infections in woody plant degrading pathogens of Botryosphaeriaceae. DNA Res. 2018, 25, 87–102. [Google Scholar] [CrossRef] [Green Version]
- Dong, B.-Z.; Guo, L. An efficient gene disruption method for the woody plant pathogen Botryosphaeria dothidea. BMC Biotechnol. 2020, 20, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nat. Cell Biol. 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boller, T.; Felix, G. A Renaissance of elicitors: Perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef] [PubMed]
- Macho, A.P.; Zipfel, C. Plant PRRs and the activation of innate immune signaling. Mol. Cell 2014, 54, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couto, D.; Zipfel, C. Regulation of pattern recognition receptor signalling in plants. Nat. Rev. Immunol. 2016, 16, 537–552. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C.; Oldroyd, G.E.D. Plant signalling in symbiosis and immunity. Nat. Cell Biol. 2017, 543, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Howden, A.; Huitema, E. Effector-triggered post-translational modifications and their role in suppression of plant immunity. Front. Plant Sci. 2012, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, D.; Zhou, J. Phytopathogen effectors subverting host immunity: Different foes, similar battleground. Cell Host Microbe 2012, 12, 484–495. [Google Scholar] [CrossRef] [Green Version]
- Boller, T.; He, S.Y. Innate Immunity in Plants: An arms race between pattern recognition receptors in plants and effectors in microbial pathogens. Science 2009, 324, 742–744. [Google Scholar] [CrossRef] [Green Version]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Hogenhout, S.A.; Van Der Hoorn, R.A.L.; Terauchi, R.; Kamoun, S. Emerging concepts in effector biology of plant-associated organisms. Mol. Plant Microbe Interact. 2009, 22, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Han, C.; Ferreira, A.O.; Yu, X.; Ye, W.; Tripathy, S.; Kale, S.D.; Gu, B.; Sheng, Y.; Sui, Y.; et al. Transcriptional programming and functional interactions within the Phytophthora sojae RXLR effector repertoire. Plant Cell 2011, 23, 2064–2086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, B.; Wang, H.; Yang, B.; Jiang, W.; Jing, M.; Li, H.; Xia, Y.; Xu, Y.; Hu, Q.; Wang, F.; et al. Phytophthora sojae effector PsAvh240 inhibits host aspartic protease secretion to promote infection. Mol. Plant 2019, 12, 552–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Wang, Y.; Guo, B.; Jing, M.; Zhou, H.; Li, Y.; Wang, H.; Huang, J.; Wang, Y.; Ye, W. The Phytophthora sojae RXLR effector Avh238 destabilizes soybean Type2 GmACSs to suppress ethylene biosynthesis and promote infection. New Phytol. 2019, 224, 1697. [Google Scholar]
- Lin, Y.; Hu, Q.; Zhou, J.; Yin, W.; Yao, D.; Shao, Y.; Zhao, Y.; Guo, B.; Xia, Y.; Chen, Q.; et al. Phytophthora sojae effector Avr1d functions as E2 competitor and inhibits ubiquitination activity of GmPUB13 to facilitate infection. BioRxiv 2020. [Google Scholar] [CrossRef]
- Xiang, J.; Li, X.; Wu, J.; Yin, L.; Zhang, Y.; Lu, J. Studying the mechanism of Plasmopara viticola RxLR effectors on suppressing plant immunity. Front. Microbiol. 2016, 7, 709. [Google Scholar] [CrossRef]
- Lan, X.; Liu, Y.; Song, S.; Yin, L.; Xiang, J.; Qu, J.; Lu, J. Plasmopara viticola effector PvRXLR131 suppresses plant immunity by targeting plant receptor-like kinase inhibitor BKI1. Mol. Plant Pathol. 2019, 20, 765–783. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Songkumarn, P.; Wang, G.-L.; Venu, R.C.; Gowda, M.; Bellizzi, M.; Hu, J.; Liu, W.; Ebbole, D.; Meyers, B.; et al. Identification and characterization of in planta; expressed secreted effector proteins from Magnaporthe oryzae that induce cell death in rice. Mol. Plant Microbe Interact. 2013, 26, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Li, Y.; Zhao, M.; Jing, M.; Liu, X.; Liu, M.; Guo, X.; Zhang, X.; Chen, Y.; Liu, Y.; et al. Global genome and transcriptome analyses of Magnaporthe oryzae epidemic isolate 98-06 uncover novel effectors and pathogenicity-related genes, revealing gene gain and lose dynamics in genome evolution. PLoS Pathog. 2015, 11, e1004801. [Google Scholar] [CrossRef] [Green Version]
- Fang, A.; Han, Y.; Zhang, N.; Zhang, M.; Liu, L.; Li, S.; Lu, F.; Sun, W. Identification and characterization of plant cell death-inducing secreted proteins from Ustilaginoidea virens. Mol. Plant Microbe Interact. 2016, 29, 405–416. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Yang, J.; Fang, A.; Wang, J.; Li, D.; Li, Y.; Wang, S.; Cui, F.; Yu, J.; Liu, Y.; et al. The essential effector SCRE1 in Ustilaginoidea virens suppresses rice immunity via a small peptide region. Mol. Plant Pathol. 2020, 21, 445–459. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Xie, S.; Zhao, Y.; Meng, X.; Song, L.; Feng, H.; Huang, L. Hce2 domain-containing effectors contribute to the full virulence of Valsa mali in a redundant manner. Mol. Plant Pathol. 2019, 20, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, Y.; Jian, H.; Yang, D.; Dai, Y.; Pan, L.; Shi, F.; Yang, S.; Liu, Q. Large-scale identification and characterization of Heterodera Avenae putative effectors suppressing or inducing cell death in Nicotiana benthamiana. Front. Plant Sci. 2018, 8, 2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Pan, L.; Chen, Y.; Yang, D.; Liu, Q.; Jian, H. Heterodera avenae GLAND5 effector interacts with pyruvate dehydrogenase subunit of plant to promote nematode parasitism. Front. Microbiol. 2019, 10, 1241. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z. Study of the Infection Process of Botryosphaeria Dothidea on Apple Shoots and Identification of a Mycovirus Infect Botryosphaeria Dothidea. Ph.D. Thesis, China Agricultural University, Beijing, China, 2018. [Google Scholar]
- Jacobs, K.A.; Collins-Racie, L.A.; Colbert, M.; Duckett, M.; Golden-Fleet, M.; Kelleher, K.; Kriz, R.; LaVallie, E.R.; Merberg, D.; Spaulding, V.; et al. A genetic selection for isolating cDNAs encoding secreted proteins. Gene 1997, 198, 289–296. [Google Scholar] [CrossRef]
- Cheng, Y.; Wu, K.; Yao, J.; Li, S.; Wang, X.; Huang, L.; Kang, Z. PSTha5a23, a candidate effector from the obligate biotrophic pathogen Puccinia striiformis f. sp. tritici, is involved in plant defense suppression and rust pathogenicity. Environ. Microbiol. 2017, 19, 1717–1729. [Google Scholar] [CrossRef]
- Cheng, B.; Yu, X.; Ma, Z.; Dong, S.; Dou, D.; Wang, Y.; Zheng, X. Phytophthora sojae effector Avh331 suppresses the plant defence response by disturbing the MAPK signalling pathway. Physiol. Mol. Plant Pathol. 2012, 77, 1–9. [Google Scholar] [CrossRef]
- Kamoun, S.; Van West, P.; Vleeshouwers, V.G.; De Groot, K.E.; Govers, F. Resistance of Nicotiana benthamiana to Phytophthora infestans is mediated by the recognition of the elicitor protein INF1. Plant Cell 1998, 10, 1413–1425. [Google Scholar]
- Kamoun, S.; van West, P.; de Jong, A.J.; de Groot, K.E.; Vleeshouwers, V.G.; Govers, F. A gene encoding a protein elicitor of Phytophthora infestans is down-regulated during infection of potato. Mol. Plant Microbe Interact. 1997, 10, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Kamoun, S. Groovy times: Filamentous pathogen effectors revealed. Curr. Opin. Plant Biol. 2007, 10, 358–365. [Google Scholar] [CrossRef]
- Göhre, V.; Robatzek, S. Breaking the barriers: Microbial effector molecules subvert plant immunity. Annu. Rev. Phytopathol. 2008, 46, 189–215. [Google Scholar] [CrossRef] [Green Version]
- Stergiopoulos, I.; de Wit, P.J. Fungal effector proteins. Annu. Rev. Phytopathol. 2009, 47, 233–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraldo, M.C.; Valent, B. Filamentous plant pathogen effectors in action. Nat. Rev. Genet. 2013, 11, 800–814. [Google Scholar] [CrossRef] [PubMed]
- Rovenich, H.; Boshoven, J.C.; Thomma, B.P.H.J. Filamentous pathogen effector functions: Of pathogens, hosts and microbiomes. Curr. Opin. Plant Biol. 2014, 20, 96–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Presti, L.L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal effectors and plant susceptibility. Annu. Rev. Plant. Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yin, Z.; Fan, Y.; Xu, M.; Kang, Z.; Huang, L. Candidate effector proteins of the necrotrophic apple canker pathogen Valsa mali can suppress BAX-induced PCD. Front. Plant Sci. 2015, 6, 579. [Google Scholar] [CrossRef] [Green Version]
- Qi, T.; Guo, J.; Liu, P.; He, F.; Wan, C.; Islam, A.; Tyler, B.M.; Kang, Z.; Guo, J. Stripe rust effector PstGSRE1 disrupts nuclear localization of ROS-promoting transcription factor TaLOL2 to defeat ROS-induced defense in wheat. Mol. Plant. 2019, 12, 1624–1638. [Google Scholar] [CrossRef]
- Xu, Q.; Tang, C.; Wang, X.; Sun, S.; Zhao, J.; Kang, Z.; Wang, X. An effector protein of the wheat stripe rust fungus targets chloroplasts and suppresses chloroplast function. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Huai, B.; Lu, Y.; Cai, K.; Guo, J.; Zhu, X.; Kang, Z.; Guo, J. A stripe rust effector Pst18363 targets and stabilises TaNUDX23 that promotes stripe rust disease. New Phytol. 2019, 225, 880–895. [Google Scholar] [CrossRef]
- Qi, M.; Mei, Y.; Grayczyk, J.P.; Darben, L.M.; Rieker, M.E.G.; Seitz, J.M.; Voegele, R.T.; Whitham, S.A.; Link, T.I. Candidate effectors from Uromyces appendiculatus, the causal agent of rust on common bean, can be discriminated based on suppression of immune responses. Front. Plant Sci. 2019, 10, 1182. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.X.; Zhang, C.J.; Weng, S.H.; Guo, L.Y.; Zhu, X.Q. Candidate effectors of Botryosphaeria dothidea inhibit Bax induced PCD in Nicotiana benthamiana and promote the infection of Phytophthora nicotianae. Acta Phytopathol. Sin. 2019, 49, 254–261. (In Chinese) [Google Scholar]
- Sonah, H.; Deshmukh, R.K.; Bélanger, R.R. Computational prediction of effector proteins in fungi: Opportunities and challenges. Front. Plant Sci. 2016, 7, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, L.; Hamilton, A.J.; Voinnet, O.; Thomas, C.L.; Maule, A.J.; Baulcombe, D.C. RNA–DNA interactions and DNA methylation in post-transcriptional gene silencing. Plant Cell 1999, 11, 2291–2301. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.-J.; Wang, S.-X.; Liang, Y.-N.; Wen, S.-H.; Dong, B.-Z.; Ding, Z.; Guo, L.-Y.; Zhu, X.-Q. Candidate Effectors from Botryosphaeria dothidea Suppress Plant Immunity and Contribute to Virulence. Int. J. Mol. Sci. 2021, 22, 552. https://doi.org/10.3390/ijms22020552
Zhang C-J, Wang S-X, Liang Y-N, Wen S-H, Dong B-Z, Ding Z, Guo L-Y, Zhu X-Q. Candidate Effectors from Botryosphaeria dothidea Suppress Plant Immunity and Contribute to Virulence. International Journal of Molecular Sciences. 2021; 22(2):552. https://doi.org/10.3390/ijms22020552
Chicago/Turabian StyleZhang, Chuan-Jie, Shi-Xing Wang, Yan-Na Liang, Sheng-Hui Wen, Bao-Zhu Dong, Zheng Ding, Li-Yun Guo, and Xiao-Qiong Zhu. 2021. "Candidate Effectors from Botryosphaeria dothidea Suppress Plant Immunity and Contribute to Virulence" International Journal of Molecular Sciences 22, no. 2: 552. https://doi.org/10.3390/ijms22020552
APA StyleZhang, C. -J., Wang, S. -X., Liang, Y. -N., Wen, S. -H., Dong, B. -Z., Ding, Z., Guo, L. -Y., & Zhu, X. -Q. (2021). Candidate Effectors from Botryosphaeria dothidea Suppress Plant Immunity and Contribute to Virulence. International Journal of Molecular Sciences, 22(2), 552. https://doi.org/10.3390/ijms22020552