Gliadin Sequestration as a Novel Therapy for Celiac Disease: A Prospective Application for Polyphenols



Abstract
:
1. Introduction
2. Gluten Proteins
3. Celiac Disease Pathogenesis
4. Current Treatment Strategies Undergoing Clinical Trial
4.1. Pathophysiology-Targeted Therapies
4.1.1. Sensitization
4.1.2. Gut Barrier Function Enhancement
4.1.3. Immunosuppression
4.1.4. Disruption of Antigen Presentation
4.2. Gliadin-Focused Therapies
4.2.1. Enzymatic Hydrolysis
4.2.2. Sequestration
5. Polyphenols as Therapeutics for Chronic Inflammatory Diseases
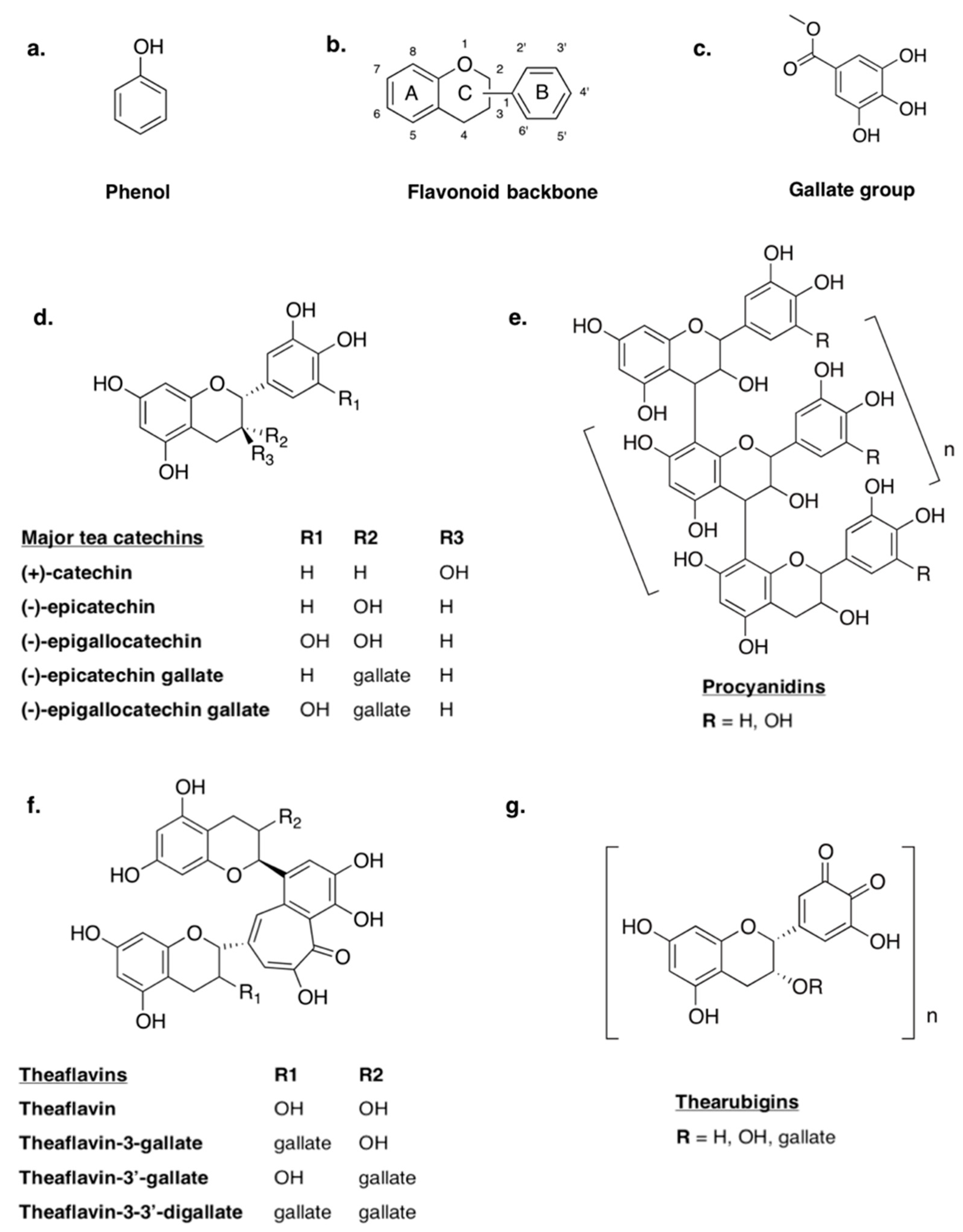
5.1. Protein–Polyphenol Interactions
5.2. Anti-Inflammatory Properties of Polyphenols
5.3. Anti-Nutritional Properties of Polyphenols
5.4. Therapeutic Applications of Polyphenols
5.4.1. Polyphenols as Therapeutic Agents for IBD
5.4.2. Polyphenols as Mediators of Allergic Responses
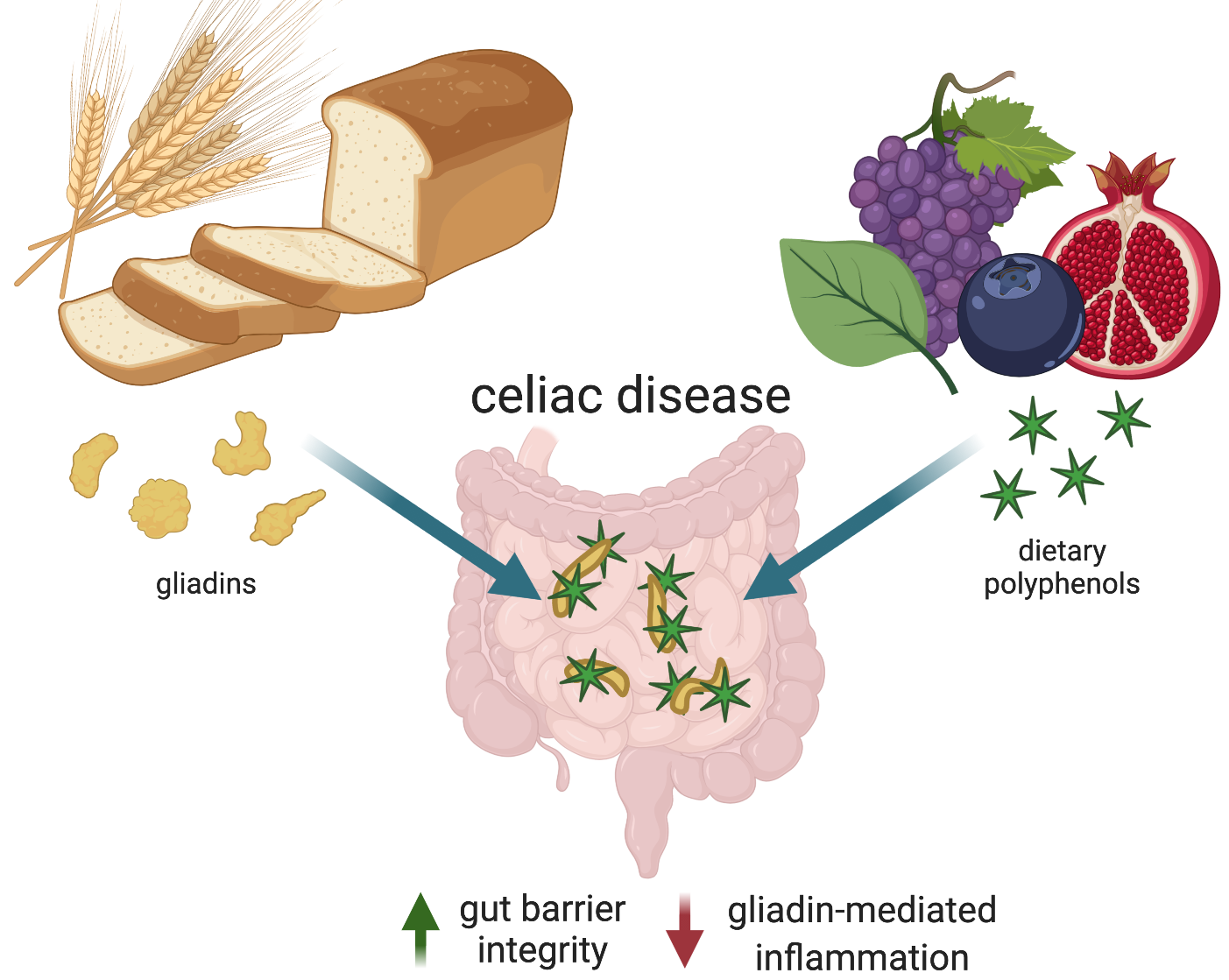
6. Polyphenols as a Prospective Therapy for Celiac Disease
6.1. Gliadin–Polpyhenol Interactions
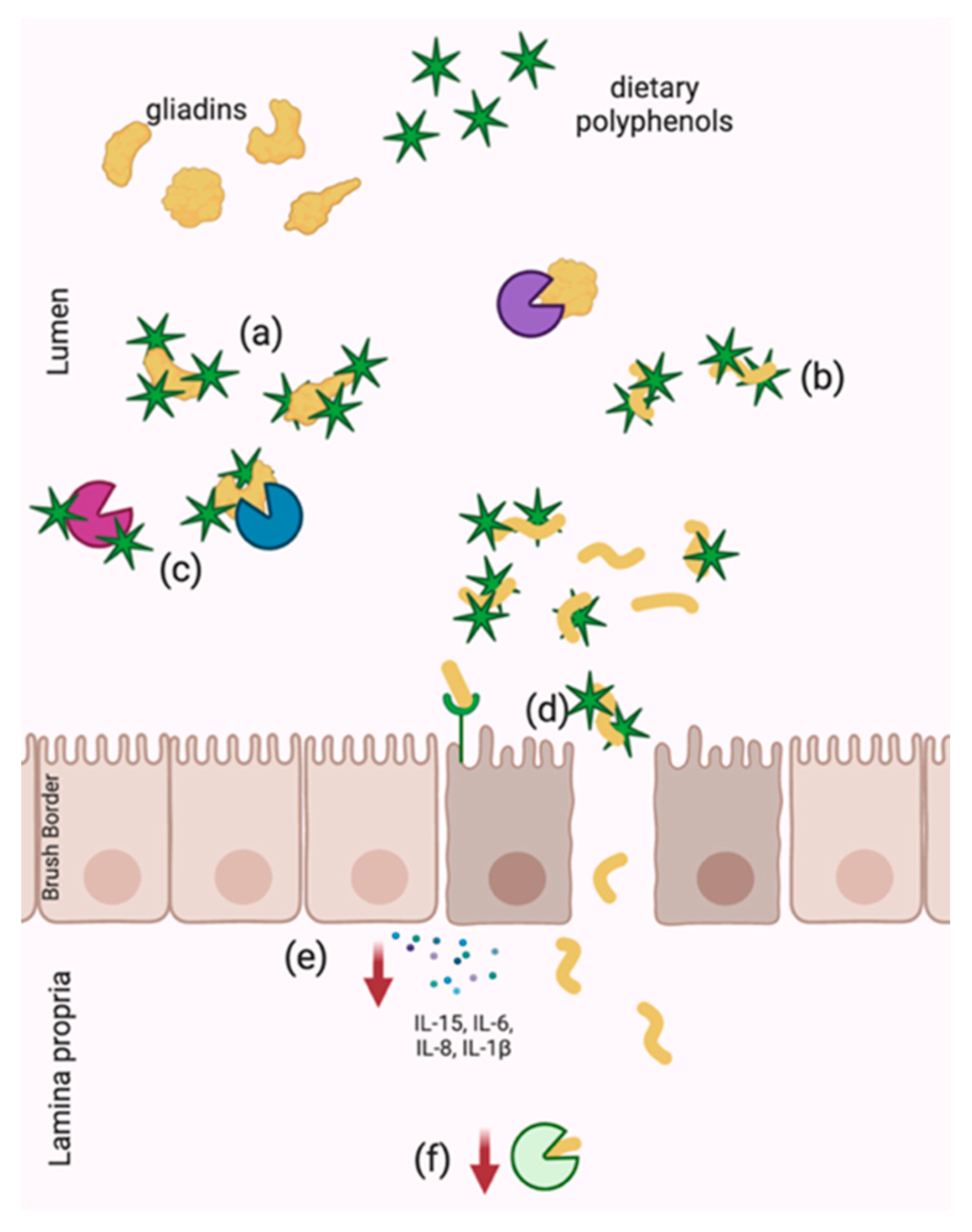
6.1.1. Physical Sequestration of Gliadins
6.1.2. Structural Change
6.2. Inhibition of Gliadin Digestion
6.3. Protective Effects of Polyphenols Towards Celiac Disease In Vitro
6.3.1. Initiation of Intestinal Permeability
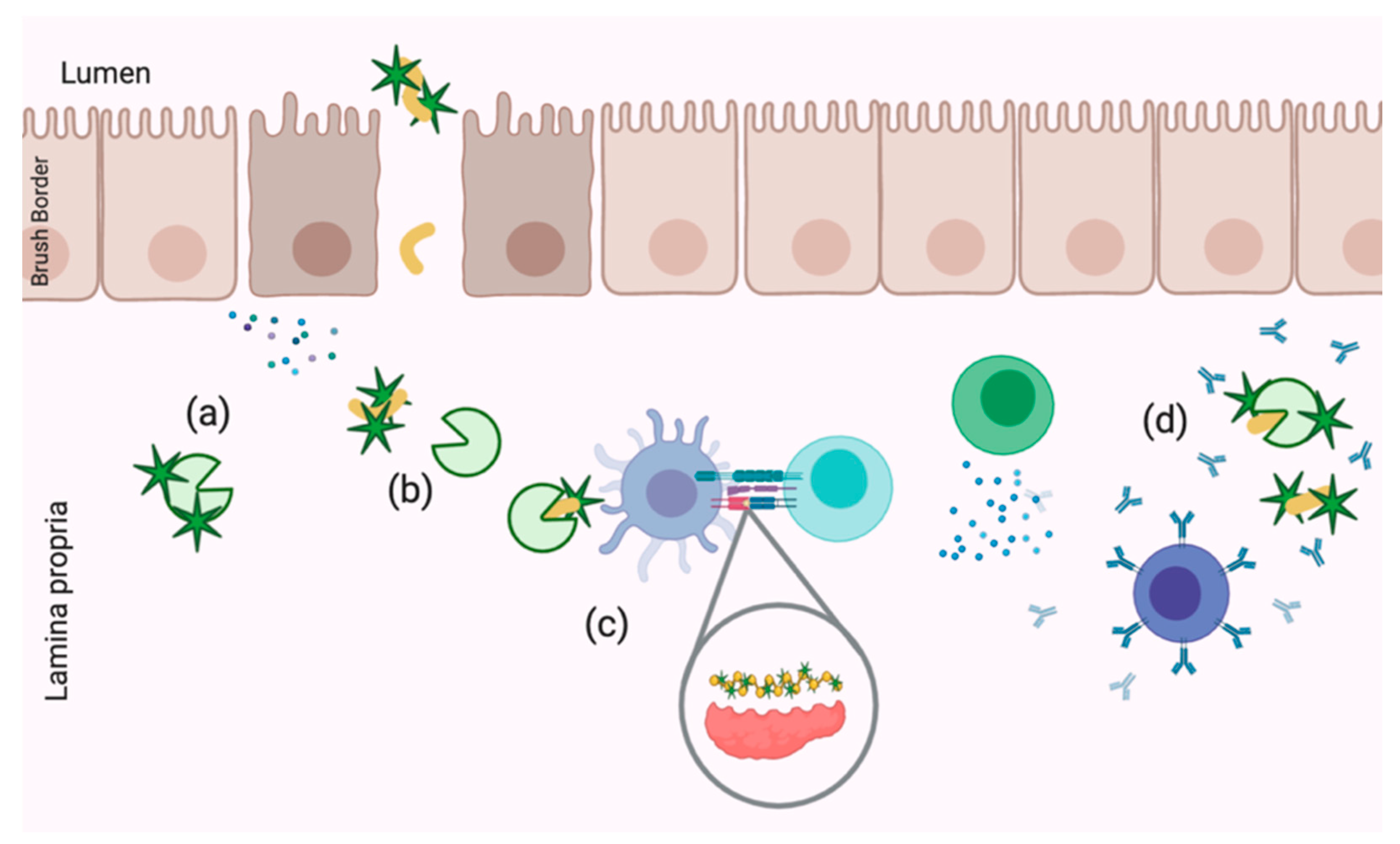
6.3.2. Transport
6.3.3. Amelioration of Gliadin-Mediated Inflammation
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| IBD | Inflammatory bowel disease |
| NMR | Nuclear magnetic resonance |
| EGCG | Epigallocatechin gallate |
| PPII | Polyproline II helix |
| TG2 | Tissue transglutaminase |
| IL | Interleukin |
| Ig | Immunoglobulin |
References
- Arentz-Hansen, H.; Körner, R.; Molberg, Ø.; Quarsten, H.; Vader, W.; Kooy, Y.M.; Lundin, K.E.; Koning, F.; Roepstorff, P.; Sollid, L.M.; et al. The intestinal T cell response to alpha-gliadin in adult celiac disease is focused on a single deamidated glutamine targeted by tissue transglutaminase. J. Exp. Med. 2000, 191, 603–612. [Google Scholar] [PubMed]
- Rubio-Tapia, A.; Ludvigsson, J.F.; Brantner, T.L.; Murray, J.A.; Everhart, J.E. The Prevalence of Celiac Disease in the United States. Am. J. Gastroenterol. 2012, 107, 1538–1544. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A.; Berti, I.; Gerarduzzi, T.; Not, T.; Colletti, R.B.; Drago, S.; Elitsur, Y.; Green, P.H.; Guandalini, S.; Hill, I.D.; et al. Prevalence of Celiac Disease in At-Risk and Not-At-Risk Groups in the United States. Arch. Intern. Med. 2003, 163, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Lionetti, E.; Catassi, C. The Role of Environmental Factors in the Development of Celiac Disease: What Is New? Diseases 2015, 3, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Ng, D.; Zivin, J.; Green, P. Economic burden of a gluten-free diet. J. Hum. Nutr. Diet. 2007, 20, 423–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szaflarska-Poplawska, A. Non-dietary methods in the treatment of celiac disease. Prz. Gastroenterol. 2015, 10, 12–17. [Google Scholar] [CrossRef]
- Sollid, L.M.; Khosla, C. Novel Therapies for Coeliac Disease. J. Intern. Med. 2012, 269, 604–613. [Google Scholar] [CrossRef] [Green Version]
- Shan, L.; Qiao, S.W.; Arentz-Hansen, H.; Molberg, Ø.; Gray, G.M.; Sollid, L.M.; Khosla, C. Identification and Analysis of Multivalent Proteolytically Resistant Peptides from Gluten: Implications for Celiac Sprue. J. Proteome Res. 2006, 4, 1732–1741. [Google Scholar]
- Pinier, M.; Verdu, E.F.; Nasser-Eddine, M.; David, C.S.; Vézina, A.; Rivard, N.; Leroux, J.C. Polymeric binders suppress gliadin-induced toxicity in the intestinal epithelium. Gastroenterology 2009, 136, 288–298. [Google Scholar] [CrossRef] [Green Version]
- Pinier, M.; Fuhrmann, G.; Galipeau, H.J.; Rivard, N.; Murray, J.A.; David, C.S.; Drasarova, H.; Tuckova, L.; Leroux, J.C.; Verdu, E.F. The copolymer P(HEMA-co-SS) binds gluten and reduces immune response in gluten-sensitized mice and human tissues. Gastroenterology 2012, 142, 316–325. [Google Scholar] [CrossRef] [Green Version]
- Van Buiten, C.B. Physicochemical Modification of Gliadin by Dietary Polyphenols and the Potential Implications for Celiac Disease. Ph.D. Thesis, Pennsylvania State University, University Park, PA, USA, 2017. [Google Scholar]
- Van Buiten, C.B.; Lambert, J.D.; Elias, R.J. Green Tea Polyphenols Mitigate Gliadin-Mediated Inflammation and Permeability in Vitro. Mol. Nutr. Food Res. 2018, 62, e1700879. [Google Scholar] [CrossRef] [PubMed]
- Van Buiten, C.B.; Yennawar, N.H.; Pacheco, C.N.; Hatzakis, E.; Elias, R.J. Physicochemical interactions with (-)-epigallocatechin-3-gallate drive structural modification of celiac-associated peptide α2-gliadin (57-89) at physiological conditions. Food Funct. 2019, 10, 2997–3007. [Google Scholar] [CrossRef] [PubMed]
- Salunkhe, D.; Jadhav, S.; Kadam, S.; Chavan, J.; Lus, B. Chemical, biochemical and biological significance of polyphenols in cereals and legumes. Crit. Rev. Food Sci. Nutr. 1983, 17, 277–305. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.; Wong, T.; Wei, C.; Huang, Y.; Lin, Y. Tannins and Human Health: A Review. Crit. Rev. Food Sci. Nutr. 1998, 38, 421–464. [Google Scholar] [CrossRef] [PubMed]
- Zuercher, A.; Holvoet, S.; Weiss, M.; Mercenier, A. Polyphenol-enriched apple extract attenuates food allergy in mice. Clin. Exp. Allergy 2010, 40, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Plundrich, N.J.; Kulis, M.; White, B.L.; Grace, M.H.; Guo, R.; Burks, A.W.; Davis, J.P.; Lila, M.A. Novel strategy to create hypoallergenic peanut protein-polyphenol edible matrices for oral immunotherapy. J. Agric. Food Chem. 2014, 62, 7010–7021. [Google Scholar] [CrossRef] [PubMed]
- Vesic, J.; Stambolic, I.; Apostolovic, D.; Milcic, M.; Stanic-Vucinic, D.; Cirkovic Velickovic, T. Complexes of green tea polyphenol, epigalocatechin-3-gallate, and 2S albumins of peanut. Food Chem. 2015, 185, 309–317. [Google Scholar] [CrossRef]
- Chung, S.Y.; Champagne, E.T. Reducing the allergenic capacity of peanut extracts and liquid peanut butter by phenolic compounds. Food Chem. 2009, 115, 1345–1349. [Google Scholar] [CrossRef]
- Parrot, I.; Huang, P.C.; Khosla, C. Circular dichroism and nuclear magnetic resonance spectroscopic analysis of immunogenic gluten peptides and their analogs. J. Biol. Chem. 2002, 277, 45572–45578. [Google Scholar] [CrossRef] [Green Version]
- Shan, L.; Molberg, Ø.; Parrot, I.; Hausch, F.; Filiz, F.; Gray, G.M.; Sollid, L.M.; Khosla, C. Structural basis for gluten intolerance in celiac sprue. Science 2002, 297, 2275–2279. [Google Scholar] [CrossRef] [Green Version]
- Shewry, P.R.; Halford, N.G. Cereal seed storage proteins: Structures, properties and role in grain utilization. J. Exp. Bot. 2002, 53, 947–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shewry, P.R.; Halford, N.G.; Belton, P.S.; Tatham, A.S. The structure and properties of gluten: An elastic protein from wheat grain. Philos. Trans. R. Soc. Lond. 2002, 357, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P.R.; Tatham, A.S. The prolamin storage proteins of cereal seeds: Structure and evolution. Biochem. J. 1990, 267, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinsella, J.E. Food Proteins; Fox, P., Condon, J.J., Eds.; Elsevier Applied Science: London, UK, 1982. [Google Scholar]
- Ortolan, F.; Steel, C.J. Protein Characteristics that Affect the Quality of Vital Wheat Gluten to be Used in Baking: A Review. Compr. Rev. Food Sci. Food Saf. 2017. [Google Scholar] [CrossRef] [Green Version]
- Howdle, P.D. Gliadin, glutenin or both? The search for the Holy Grail in coeliac disease. Eur. J. Gastroenterol. Hepatol. 2006, 18, 703–706. [Google Scholar] [CrossRef]
- Wieser, H. Relation between gliadin structure and coeliac toxicity. Acta Paediatr. Suppl. 1996, 412, 3–9. [Google Scholar] [CrossRef]
- Berg, J.M.; Tymoczko, J.L. Section 23.1, Proteins Are Degraded to Amino Acids. In Biochemistry; W. H. Freeman: New York, NY, USA, 2002. [Google Scholar]
- Hausch, F.; Shan, L.; Santiago, N.A.; Gray, G.M.; Khosla, C. Intestinal digestive resistance of immunodominant gliadin peptides. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 283, G996–G1003. [Google Scholar] [CrossRef] [Green Version]
- Iacomino, G.; Fierro, O.; D’Auria, S.; Picariello, G.; Ferranti, P.; Liguori, C.; Addeo, F.; Mamone, G. Structural analysis and Caco-2 cell permeability of the celiac-toxic A-gliadin peptide 31-55. J. Agric. Food Chem. 2013, 61, 1088–1096. [Google Scholar] [CrossRef]
- Lammers, K.M.; Lu, R.; Brownley, J.; Lu, B.; Gerard, C.; Rallabhandi, P.; Shea-Donohue, T.; Tamiz, A.; Alkan, S. Gliadin Induces an Increase in Intestinal Permeability and Zonulin Release by Binding to the Chemokine Receptor CXCR3. Gastroenterology 2008, 135, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Meresse, B.; Ripoche, J.; Heyman, M.; Cerf-Bensussan, N. Celiac disease: From oral tolerance to intestinal inflammation, autoimmunity and lymphomagenesis. Mucosal Immunol. 2009, 2, 8–23. [Google Scholar] [CrossRef]
- Fasano, A. Zonulin and Its Regulation of Intestinal Barrier Function: The Biological Door to Inflammation, Autoimmunity, and Cancer. Physiol. Rev. 2011, 91, 151–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matysiak-Budnik, T.; Moura, I.C.; Arcos-Fajardo, M.; Lebreton, C.; Menard, S.; Candalh, C.; Ben-Khalifa, K.; Dugave, C.; Tamouza, H.; Van Niel, G.; et al. Secretory IgA mediates retrotranscytosis of intact gliadin peptides via the transferrin receptor in celiac disease. J. Exp. Med. 2008, 205, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Schumann, M.; Richter, J.; Wedell, I.; Moos, V.; Schneider, T.; Daum, S.; Zeitz, M.; Fromm, M.; Schulake, J. Mechanisms of epithelial translocation of the α2-gliadin-33mer in celiac sprue. Gut 2008, 57, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A. Novel therapeutic/integrative approaches for celiac disease and dermatitis herpetiformis. Clin. Dev. Immunol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Waldmann, T.A. Targeting the interleukin-15/interleukin-15 receptor system in inflammatory autoimmune diseases. Arthritis Res. Ther. 2004, 6, 174–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abadie, V.; Jabri, B. IL-15: A central regulator of celiac disease immunopathology. Immunol. Rev. 2014, 260, 221–234. [Google Scholar] [CrossRef] [Green Version]
- Ciccocioppo, R.; Di Sabatino, A.; Corazza, G.R. The immune recognition of gluten in coeliac disease. Clin. Exp. Immunol. 2005, 140, 408–416. [Google Scholar] [CrossRef]
- Benahmed, M.; Meresse, B.; Arnulf, B.; Barbe, U.; Mention, J.J.; Verkarre, V.; Allez, M.; Cellier, C.; Hermine, O.; Cerf-Bensussan, N. Inhibition of TGF-beta Signaling by IL-15: A New Role for IL-15 in the Loss of Immune Homeostasis in Celiac Disease. Gastroenterology 2007, 132, 994–1008. [Google Scholar] [CrossRef]
- Thomas, K.; Sapone, A.; Fasano, A.; Vogel, S. Gliadin Stimulation of Murine Macrophage Inflammatory Gene Expression and Intestinal Permeability Are MyD88-Dependent: Role of the Innate Immune Response in Celiac Disease. J. Immunol. 2006, 176, 2512–2521. [Google Scholar] [CrossRef] [Green Version]
- Visser, J.; Rozing, J.; Sapone, A.; Lammers, K.; Fasano, A. Tight Junctions, Intestinal Permeability and Autoimmunity Celiac Disease and Type 1 Diabetes Paradigms. Ann. N. Y. Acad. Sci. 2010, 1165, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Neunlist, M.; Van Landeghem, L.; Mahé, M.M.; Derkinderen, P.; des Varannes, S.B.; Rolli-Derkinderen, M. The digestive neuronal-glial-epithelial unit: A new actor in gut health and disease. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Sander, G.R.; Cummins, A.G.; Powell, B.C. Rapid disruption of intestinal barrier function by gliadin involves altered expression of apical junctional proteins. FEBS Lett. 2005, 579, 4851–4855. [Google Scholar] [CrossRef] [PubMed]
- Clemente, M.; De Virgiliis, S.; Kang, J.; Macatagney, R.; Musu, M.; Di Pierro, M.; Drago, S.; Congia, M.; Fasano, A. Early effects of gliadin on enterocyte intracellular signalling involved in intestinal barrier function. Gut 2003, 52, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Bruce, S.; Bjarnson, I.; Peters, T. Human jejunal transglutaminase: Demonstration of activity, enzyme kinetics and substrate specificity with special relation to gliadin and coeliac disease. Clin. Sci. 1985, 68, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Skovbjerg, H.; Anthonsen, D.; Knudsen, E.; Sjöström, H. Deamidation of gliadin peptides in lamina propria: Implications for celiac disease. Dig. Dis. Sci. 2008, 53, 2917–2924. [Google Scholar] [CrossRef] [PubMed]
- Iversen, R.; Amundsen, S.F.; Kleppa, L.; du Pré, M.F.; Stamnaes, J.; Sollid, L.M. Evidence That Pathogenic Transglutaminase 2 in Celiac Disease Derives From Enterocytes. Gastroenterology 2020, 159, 788–790. [Google Scholar] [CrossRef]
- Du Pré, M.F.; Du Pré, M.F.; Blazevski, J.; Dewan, A.E.; Dewan, A.E.; Stamnaes, J.; Stamnaes, J.; Stamnaes, J.; Kanduri, C.; Kanduri, C.; et al. B cell tolerance and antibody production to the celiac disease autoantigen transglutaminase 2. J. Exp. Med. 2020, 217, e20190860. [Google Scholar] [CrossRef] [PubMed]
- Di Sabatino, A.; Vanoli, A.; Giuffrida, P.; Luinetti, O.; Solcia, E.; Roberto, G. The function of tissue transglutaminase in celiac disease. Autoimmun. Rev. 2012, 11, 746–753. [Google Scholar] [CrossRef]
- Qiao, S.W.; Bergseng, E.; Molberg, Ø.; Xia, J.; Fleckenstein, B.; Khosla, C.; Sollid, L.M. Antigen presentation to celiac lesion-derived T cells of a 33-mer gliadin peptide naturally formed by gastrointestinal digestion. J. Immunol. 2004, 173, 1757–1762. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.H.; Jardetzky, T.S.; Gorga, J.C.; Stern, L.J.; Urban, R.G.; Strominger, J.L.; Wiley, D.C. Three-dimensional structure of the human class II histocompatibility antigen HLA-DR1. Nature 1993, 364, 33–39. [Google Scholar] [CrossRef]
- Stern, L.J.; Brown, J.H.; Jardetzky, T.S.; Gorga, J.C.; Urban, R.G.; Strominger, J.L.; Wiley, D.C. Crystal structure of the human class II MHC protein HLA-DR1 complexed with an influenza virus peptide. Nature 1994, 368, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Qiao, S.W.; Sollid, L.M.; Blumberg, R.S. Antigen presentation in celiac disease. Curr. Opin. Immunol. 2009, 21, 111–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzarella, G. Effector and suppressor T cells in celiac disease. World J. Gastroenterol. 2015, 21, 7349–7356. [Google Scholar] [CrossRef] [PubMed]
- Gianfrani, C.; Auricchio, S.; Troncone, R. Adaptive and innate immune responses in celiac disease. Immunol. Lett. 2005, 99, 141–145. [Google Scholar] [CrossRef]
- Schuppan, D.; Junker, Y.; Barisani, D. Celiac disease: From pathogenesis to novel therapies. Gastroenterology 2009, 137, 1912–1933. [Google Scholar] [CrossRef]
- du Pré, M.F.; Sollid, L.M. T-cell and B-cell immunity in celiac disease. Best Pract. Res. Clin. Gastroenterol. 2015, 29, 413–423. [Google Scholar] [CrossRef]
- Catassi, C.; Fabiani, E.; Iacono, G.; D’Agate, C.; Francavilla, R.; Biagi, F.; Volta, U.; Accomando, S.; Picarelli, A.; De Vitis, I.; et al. A prospective, double-blind, placebo-controlled trial to establish a safe gluten threshold for patients with celiac disease. Am. J. Clin. Nutr. 2007, 85, 160–166. [Google Scholar] [CrossRef]
- Barker, J.; Liu, E. Celiac Disease: Pathophysiology, Clinical Manifestations and Associated Autoimmune Conditions. Adv. Pediatr. 2008, 55, 349–365. [Google Scholar] [CrossRef] [Green Version]
- Fasano, A.; Catassi, C. Current Approaches to Diagnosis and Treatment of Celiac Disease: An Evolving Spectrum. Gastroenterology 2001, 120, 636–651. [Google Scholar] [CrossRef]
- Fasano, A.; Catassi, C. Celiac Disease. N. Engl. J. Med. 2007, 367, 2419–2426. [Google Scholar] [CrossRef]
- Dickson, B.; Streutker, C.; Chetty, R. Coeliac disease: An update for pathologists. J. Clin. Pathol. 2006, 59, 1008–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clevers, H. The Intestinal Crypt, A Prototype Stem Cell Compartment. Cell 2013, 154, 274–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, S.T.; Greenson, J.K. The Clinical Significance of Duodenal Lymphocytosis With Normal Villus Architecture. Arch. Pathol. Lab. Med. 2013, 137, 1216–1219. [Google Scholar] [CrossRef] [PubMed]
- Dicke, W.; Weijers, H.; van de Kamer, J. Coeliac Disease: II. The Presence in Wheat of a Factor Having a Deleterious Effect in Cases of Coeliac Disease. Acta Paediatr. 1953, 42, 34–42. [Google Scholar] [CrossRef]
- Anderson, C.; Frazer, A.; French, J.; Gerrard, J.; Sammons, H.; Smellie, J. Coeliac Disease: Gastrointestinal Studies and the Effect of Dietary. Lancet 1952, 259, 836–842. [Google Scholar] [CrossRef]
- van de Kamer, J.; Weijers, H.; Dicke, W. Coeliac Disease: IV. An Investigation into the Injurious Constituents of Wheat in Connection with their Action on Patients with Celiac Disease. Acta Paediatr. 1953, 42, 223–231. [Google Scholar] [CrossRef]
- Murray, J.A.; Watson, T.; Clearman, B.; Mitros, F. Effect of a gluten-free diet on gastrointestinal symptoms in celiac disease. Am. J. Clin. Nutr. 2004, 79, 669–673. [Google Scholar] [CrossRef] [Green Version]
- Galli, G.; Esposito, G.; Lahner, E.; Pilozzi, E.; Corleto, V.; Di Giulio, E.; Aloe Spiriti, M.; Annibale, B. Histological recovery and gluten-free diet adherence: A prospective 1-year follow-up study of adult patients with coeliac disease. Aliment. Pharmacol. Ther. 2014, 40, 639–647. [Google Scholar] [CrossRef] [Green Version]
- Saturni, L.; Ferretti, G.; Bacchetti, T. The Gluten-Free Diet: Safety and Nutritional Quality. Nutrients 2010, 2, 16–34. [Google Scholar] [CrossRef] [Green Version]
- Stein, J.; Schuppan, D. Coeliac Disease—New Pathophysiological Findings and their Implications for Therapy. Viszeralmedizin 2014, 30, 156–165. [Google Scholar] [CrossRef] [Green Version]
- Goel, G.; Andrews, J.M.; Krause, R.; Fogel, R.; Barish, C.F.; Daveson, A.; Epstein, R.; King, T.; Kinney, T.; Krishnarajah, J.; et al. Nexvax2(R), a Peptide-Based Antigen-Specific Immunotherapy, Administered Intra-Dermally Three-Times Over 15-Days attenuates Responsiveness to Immuno-Dominant Gluten Peptides in HLA-DQ2.5+ People with Celiac Disease (CeD). Gastroenterol. Abstr. 2016, 150, S304. [Google Scholar] [CrossRef]
- Truitt, K.E.; Daveson, A.J.M.; Ee, H.C.; Goel, G.; MacDougall, J.; Neff, K.; Anderson, R.P. Randomised clinical trial: A placebo-controlled study of subcutaneous or intradermal NEXVAX2, an investigational immunomodulatory peptide therapy for coeliac disease. Aliment. Pharmacol. Ther. 2019, 50, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Daveson, A.J.M.; Popp, A.; Taavela, J.; Goldstein, K.E.; Isola, J.; Truitt, K.E.; Mäki, M.; Anderson, R.P.; Adams, A.; Andrews, J.; et al. Baseline quantitative histology in therapeutics trials reveals villus atrophy in most patients with coeliac disease who appear well controlled on gluten-free diet. GastroHep 2020, 2, 22–30. [Google Scholar] [CrossRef] [Green Version]
- ImmusanT Discontinues Phase 2 Clinical Trials for Nexvax2(R). Available online: https://www.coeliac.org.au/news-stories/immusant-discontinues-phase-2-clinical-trials-for-nexvax2/ (accessed on 27 November 2020).
- Lapara, J.M.; Olivares, M.; Gallina, O.; Sanz, Y. Bifidobacterium longum CECT 7347 Modulates Immune Responses in a Gliadin-Induced Enteropathy Animal Model. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Sarno, M.; Lania, G.; Cuomo, M.; Nigro, F.; Passannanti, F.; Budelli, A.; Fasano, F.; Troncone, R.; Auricchio, S.; Barone, M.V.; et al. Lactobacillus paracasei CBA L74 interferes with gliadin peptides entrance in Caco-2 cells. Int. J. Food Sci. Nutr. 2014, 65, 953–959. [Google Scholar] [CrossRef]
- Lindfors, K.; Blomqvist, T.; Juuti-Uusitalo, K.; Stenman, S.; Venäläinen, M.M.; Kaukinen, K. Live probiotic Bifidobacterium lactis bacteria inhibit the toxic effects induced by wheat gliadin in epithelial cell culture. Clin. Exp. Immunol. 2008, 152, 552–558. [Google Scholar] [CrossRef]
- Khaleghi, S.; Ju, J.M.; Lamba, A.; Murray, J.A. The potential utility of tight junction regulation in celiac disease: Focus on larazotide acetate. Therap. Adv. Gastroenterol. 2016, 9, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishnan, S.; Pandey, N.; Tamiz, A.P.; Vere, J.; Carrasco, R.; Somerville, R.; Tripathi, A.; Ginski, M.; Paterson, B.M.; Alkan, S.S. Mechanism of action of ZOT-derived peptide AT-1002, a tight junction regulator and absorption enhancer. Int. J. Pharm. 2009, 365, 121–130. [Google Scholar] [CrossRef]
- Leffler, D.A.; Kelly, C.P.; Green, P.H.R.; Fedorak, R.N.; DiMarino, A.; Perrow, W.; Rasmussen, H.; Wang, C.; Bercik, P.; Bachir, N.M.; et al. Larazotide acetate for persistent symptoms of celiac disease despite a gluten-free diet: A randomized controlled trial. Gastroenterology 2015, 148, 1311–1319. [Google Scholar] [CrossRef] [Green Version]
- Leffler, D.A.; Kelly, C.; Abdallah, H.; Colatrella, A.; Harris, L.; Leon, F.; Arterburn, L.; Paterson, B.; Lan, Z.; Murray, J. A randomized, double-blind study of larazotide acetate to prevent the activation of celiac disease during gluten challenge. Am. J. Gastroenterol. 2012, 107, 1554–1562. [Google Scholar] [CrossRef] [Green Version]
- De Palma, G.; Cinova, J.; Stepankova, R.; Tuckova, L.; Sanz, Y. Pivotal Advance: Bifidobacteria and Gram-negative bacteria differentially influence immune responses in the proinflammatory milieu of celiac disease. J. Leukoc. Biol. 2010, 87, 765–778. [Google Scholar] [CrossRef] [PubMed]
- de Sousa Moares, L.F.; Grzeskowiak, L.M.; de Sales Teixeira, T.F.; do Carmo Gouveia Peluzio, M. Intestinal Microbiota and Probiotics in Celiac Disease. Clin. Microbiol. Rev. 2014, 27, 482–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smecuol, E.; Hwang, H.; Sugai, E.; Corso, L.; Cherñavsky, A.; Bellavite, F.; González, A.; Vodánovich, F.; Moreno, M.; Vázquez, H.; et al. Exploratory, randomized, double-blind, placebo-controlled study on the effects of Bifidobacterium infantis natren life start strain super strain in active celiac disease. J. Clin. Gastroenterol. 2013, 47, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Croese, J.; Giacomin, P.; Navarro, S.; Clouston, A.; McCann, L.; Dougall, A.; Ferreira, I.; Susianto, A.; O’Rourke, P.; Howlett, M.; et al. Experimental hookworm infection and gluten microchallenge promote tolerance in celiac disease. J. Allergy Clin. Immunol. 2014, 135, 508–516. [Google Scholar] [CrossRef]
- Yokoyama, S.; Watanabe, N.; Sato, N.; Perera, P.Y.; Filkoski, L.; Tanaka, T.; Miyasaka, M.; Waldmann, T.A.; Hiroi, T.; Perera, L.P. Antibody-mediated blockade of IL-15 reverses the autoimmune intestinal damage in transgenic mice that overexpress IL-15 in enterocytes. PNAS 2009, 106, 15849–15854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosli, M.H.; Feagan, B.G. The emerging role of vedolizumab in the treatment of ulcerative colitis. Clin. Investig. 2012, 2, 1201–1212. [Google Scholar] [CrossRef]
- Soler, D.; Chapman, T.; Yang, L.; Wyant, T.; Egan, R.; Fedyk, E.R. The Binding Specificity and Selective Antagonism of Vedolizumab, an Anti-alpha4beta7 Integrin Therapeutic Antibody in Development for Inflammatory Bowel Diseases. J. Pharmacol. Exp. Ther. 2009, 330, 864–875. [Google Scholar] [CrossRef] [Green Version]
- Walters, M.J.; Wang, Y.; Lai, N.; Baumgart, T.; Zhao, B.N.; Dairaghi, D.J.; Bekker, P.; Ertl, L.S.; Penfold, M.E.T.; Jaen, J.C.; et al. Characterization of CCX282-B, an Orally Bioavailable Antagonist of the CCR9 Chemokine Receptor, for Treatment of Inflammatory Bowel Disease. J. Pharmacol. Exp. Ther. 2010, 335, 61–69. [Google Scholar] [CrossRef]
- Keshav, S.; Vanasek, T.; Niv, Y.; Petryka, R.; Howaldt, S.; Bafutto, M.; Racz, I.; Hetzel, D.; Nielsen, O.; Severine, V.; et al. A Randomized Controlled Trial of the Efficacy and Safety of CCX282-B, an Orally-Administered Blocker of Chemokine Receptor CCR9, for Patients with Crohn’s Disease. PLoS ONE 2013, 8, e60094. [Google Scholar] [CrossRef]
- Hoffmann, K.; Alminger, M.; Andlid, T.; Chen, T.; Olsson, O.; Sandberg, A.S. Blocking Peptides Decrease Tissue Transglutaminase Processing of Gliadin in Vitro. J. Agric. Food Chem. 2009, 57, 10150–10155. [Google Scholar] [CrossRef]
- Kapoerchan, V.V.; Wiesner, M.; Overhand, M.; van der Marel, G.A.; Koning, F.; Overkleeft, H.S. Design of azidoproline containing gluten peptides to suppress CD4 + T-cell responses associated with Celiac disease. Bioorg. Med. Chem. 2008, 16, 2053–2062. [Google Scholar] [CrossRef] [PubMed]
- Kapoerchan, V.V.; Wiesner, M.; Hillaert, U.; Drijfhout, J.W.; Overhand, M.; Alard, P.; van der Marel, G.A.; Overkleeft, H.S.; Koning, F. Design, synthesis and evaluation of high-affinity binders for the celiac disease associated HLA-DQ2 molecule. Mol. Immunol. 2010, 47, 1091–1097. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Siegel, M.; Bergseng, E.; Sollid, L.M.; Khosla, C. Inhibition of HLA-DQ2-Mediated Antigen Presentation by Analogues of a High Affinity 33-Residue Peptide from alpha2-Gliadin. J. Am. Chem. Soc. 2006, 128, 1859–1867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiuri, L.; Ciacci, C.; Ricciardelli, I.; Vacca, L.; Raia, V.; Rispo, A.; Griffin, M.; Issekutz, T.; Quaratino, S.; Londei, M. Unexpected Role of Surface Transglutaminase Type II in Celiac Disease. Gastroenterology 2005, 129, 1400–1413. [Google Scholar] [CrossRef] [PubMed]
- Ventura, M.A.E.; Sajko, K.; Hils, M.; Pasternack, R.; Greinwald, R.; Tewes, B.; Schuppan, D. The Oral Transglutaminase 2 (TG2) Inhibitor Zed1227 Blocks TG2 Activity in a Mouse Model of Intestinal Inflammation. Gastroenterology 2018, 154, S490. [Google Scholar] [CrossRef]
- Reversibly Acting Transglutaminase 2 Inhibitors: Drug Candidates for the Treatment of Fibrosis. Available online: https://zedira.com/News/Reversibly-acting-transglutaminase-2-inhibitors-drug-candidates-for-the-treatment-of-fibrosis_127 (accessed on 30 November 2020).
- Press Release: Dr. Falk Pharma and Zedira Announce Completion of Phase 1b Clinical Trial of ZED1227 for the Treatment of Celiac Disease and Move on to Proof of Concept Study. Available online: https://zedira.com/News/Press-release-Dr-Falk-Pharma-and-Zedira-announce-completion-of-phase-1b-clinical-trial-of-ZED1227-for-the-treatment-of-celiac-disease-and-move-on-to-proof-of-concept-study_97 (accessed on 30 November 2020).
- Rizzello, C.G.; De Angelis, M.; Di Cagno, R.; Camarca, A.; Silano, M.; Losito, I.; De Vincenzi, M.; De Bari, M.D.; Palmisano, F.; Maurano, F.; et al. Highly Efficient Gluten Degradation by Lactobacilli and Fungal Proteases During Food Processing: New Perspectives for Celiac Disease. Appl. Environ. Microbiol. 2007, 73, 4499–4507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engström, N.; Sandberg, A.S.; Scheers, N. Sourdough Fermentation of Wheat Flour does not Prevent the Interaction of Transglutaminase 2 with α2-Gliadin or Gluten. Nutrients 2015, 7, 2134–2144. [Google Scholar] [CrossRef] [Green Version]
- Tack, G.J.; van de Water, J.M.; Bruins, M.J.; Kooy-Winkelaar, E.M.; van Bergen, J.; Bonnet, P.; Vreugdenhil, A.C.; Korponay-Szabo, I.; Edens, L.; von Blomberg, B.M.E.; et al. Consumption of gluten with gluten-degrading enzyme by celiac patients: A pilot-study. World J. Gastroenterol. 2013, 19, 5837–5847. [Google Scholar] [CrossRef] [Green Version]
- Siegel, M.; Garber, M.E.; Spencer, A.G.; Botwick, W.; Kumar, P.; Williams, R.N.; Adelman, D.C. Safety, Tolerability, and Activity of ALV003: Results from Two Phase 1 Single, Escalating-Dose Clinical Trials. Dig. Dis. Sci. 2012, 57, 440–450. [Google Scholar] [CrossRef]
- Sample, D.A.; Sunwoo, H.H.; Huynh, H.Q.; Rylance, H.L.; Robert, C.L.; Xu, B.W.; Kang, S.H.; Gujral, N.; Dieleman, L.A. AGY, a Novel Egg Yolk-Derived Anti-gliadin Antibody, Is Safe for Patients with Celiac Disease. Dig. Dis. Sci. 2017, 62, 1277–1285. [Google Scholar] [CrossRef]
- Gujral, N.; Löbenberg, R.; Suresh, M.; Sunwoo, H. In-vitro and in-vivo binding activity of chicken egg yolk immunoglobulin y (IgY) against gliadin in food matrix. J. Agric. Food Chem. 2012, 60, 3166–3172. [Google Scholar] [CrossRef] [PubMed]
- McCarville, J.L.; Nisemblat, Y.; Galipeau, H.J.; Jury, J.; Tabakman, R.; Cohen, A.; Naftali, E.; Neiman, B.; Halbfinger, E.; Murray, J.A.; et al. BL-7010 Demonstrates Specific Binding to Gliadin and Reduces Gluten-Associated Pathology in a Chronic Mouse Model of Gliadin Sensitivity. PLoS ONE 2014, 9, e109972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, L.; Pinier, M.; Leroux, J.C.; Subirade, M. Interaction of alpha-gliadin with polyanions: Design considerations for sequestrants used in supportive treatment of celiac disease. Biopolymers 2010, 93, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Pinier, M.; Leroux, J.C.; Subirade, M. Interaction of alpha-gliadin with poly(HEMA-co-SS): Structural characterization and biological implication. Biopolymers 2009, 91, 169–178. [Google Scholar] [CrossRef]
- Lattanzio, V.; Lattanzio, V.M.; Cardinali, A. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. In Phytochemistry: Advances in Research; Imperato, F., Ed.; Research Signpost: Kerala, India, 2006; pp. 23–67. ISBN 8130800349. [Google Scholar]
- Simsrod, O.; Moe, S.T. Biopolymer Chemistry; Postmyr, L., Ed.; Tapir Academic Press: Trondheim, Norway, 2008. [Google Scholar]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Green, R.J.; Murphy, A.S.; Schulz, B.; Watkins, B.A.; Ferruzzi, M.G. Common tea formulations modulate in vitro digestive recovery of green tea catechins. Mol. Nutr. Food Res. 2007, 51, 1152–1162. [Google Scholar] [CrossRef]
- Li, Y.; Jongberg, S.; Andersen, M.L.; Davies, M.J.; Lund, M.N. Quinone-induced protein modifications: Kinetic preference for reaction of 1,2-benzoquinones with thiol groups in proteins. Free Radic. Biol. Med. 2016, 97, 148–157. [Google Scholar] [CrossRef]
- Bordenave, N.; Hamaker, B.R.; Ferruzzi, M.G. Nature and consequences of non-covalent interactions between flavonoids and macronutrients in foods. Food Funct. 2014, 5, 18–34. [Google Scholar] [CrossRef]
- Hagerman, A.E.; Butler, L.G. The specificity of proanthocyanidin-protein interactions. J. Biol. Chem. 1981, 256, 4494–4497. [Google Scholar] [CrossRef]
- Jöbstl, E.; O’Connell, J.; Fairclough, J.P.; Williamson, M.P. Molecular model for astringency produced by polyphenol/protein interactions. Biomacromolecules 2004, 5, 942–949. [Google Scholar] [CrossRef]
- Charlton, A.J.; Baxter, N.J.; Khan, M.L.; Moir, A.J.; Haslam, E.; Davies, A.P.; Williamson, M.P. Polyphenol/peptide binding and precipitation. J. Agric. Food Chem. 2002, 50, 1593–1601. [Google Scholar] [CrossRef] [PubMed]
- Pietta, P.G. Flavonoids as Antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Elias, R.J. Investigating the hydrogen peroxide quenching capacity of proteins in polyphenol-rich foods. J. Agric. Food Chem. 2011, 59, 8915–8922. [Google Scholar] [CrossRef] [PubMed]
- Nanjo, F.; Mori, M.; Goto, K.; Hara, Y. Radical Scavenging Activity of Tea Catechins and Their Related Compounds. Biosci. Biotechnol. Biochem. 1999, 63, 1621–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balentine, D.A.; Wiseman, S.A.; Bouwens, L.C. The Chemistry of Tea Flavonoids. Crit. Rev. Food Sci. Nutr. 1997, 37, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Higdon, J.V.; Frei, B. Tea catechins and polyphenols: Health effects, metabolism, and antioxidant functions. Crit. Rev. Food Sci. Nutr. 2003, 43, 89–143. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.J.; Chun, K.S.; Cha, H.H.; Han, S.S.; Keum, Y.S.; Park, K.K.; Lee, S.S. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: Down-regulation of COX-2 and iNOS through suppression of NF-kB activation. Fundam. Mol. Mech. Mutagen. 2001, 480, 243–268. [Google Scholar] [CrossRef]
- Chan, M.M.; Fong, D.; Ho, C.T.; Huang, H.I. Inhibition of Inducible Nitric Oxide Synthase Gene Expression and Enzyme Activity by Epigallocatechin Gallate, a Natural Product from Green Tea. Biochem. Pharmacol. 1997, 54, 1281–1286. [Google Scholar] [CrossRef]
- Lin, Y.L.; Lin, J.K. (-)-Epigallocatechin-3-gallate Blocks the Induction of Nitric Oxide Synthase by Down-Regulating Lipopolysaccharide- Induced Activity of Transcription Factor Nuclear Factor-kB. Mol. Pharmacol. 1997, 52, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.; Smith, T.J.; Ho, C.T.; August, D.A.; Yang, C.S. Effects of purified green and black tea polyphenols on cyclooxygenase- and lipoxygenase-dependent metabolism of arachidonic acid in human colon mucosa and colon tumor tissues. Biochem. Pharmacol. 2001, 62, 1175–1183. [Google Scholar] [CrossRef]
- Zheng, Y.; Toborek, M.; Hennig, B. Epigallocatechin gallate–mediated protection against tumor necrosis factor-α–induced monocyte chemoattractant protein–1 expression is heme oxygenase–1 dependent. Metabolism 2010, 59, 1528–1535. [Google Scholar] [CrossRef] [PubMed]
- Sumi, T.; Shirakami, Y.; Shimizu, M.; Kochi, T.; Ohno, T.; Kubota, M.; Shiraki, M.; Tsurumi, H.; Tanaka, T.; Moriwaki, H. (-)-Epigallocatechin-3-gallate suppresses hepatic preneoplastic lesions developed in a novel rat model of non-alcoholic steatohepatitis. Springerplus 2013, 2, 690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, J.M.; Jialal, I.; Devaraj, S. Effects of epigallocatechin gallate on regulatory T cell number and function in obese v. lean volunteers. Br. J. Nutr. 2010, 103, 1771–1777. [Google Scholar] [CrossRef] [Green Version]
- Makkar, H.; Singh, B.; Dawra, R. Effect of tannin-rich leaves of oak (Quercus incana) on various microbial enzyme activities of the bovine rumen. Br. J. Nutr. 1988, 60, 287–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horigome, T.; Kumar, R.; Okamoto, K. Effects of condensed tannins prepared from leaves of fodder plants on digestive enzymes in vitro and in the intestine of rats. Br. J. Nutr. 1988, 60, 275–285. [Google Scholar] [CrossRef]
- Koo, S.I.; Noh, S.K. Green Tea as Inhibitor of the Intestinal Absorption of Lipids: Potential Mechanism for its Lipid-Lowering Effect. J. Nutr. Biochem. 2007, 18, 179–183. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Noh, S.K.; Koo, S.I. Epigallocatechin gallate and caffeine differentially inhibit the intestinal absorption of cholesterol and fat in ovariectomized rats. J. Nutr. 2006, 136, 2791–2796. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; He, W.; Yao, L.; Zhang, H.; Liu, Z.; Wang, W.; Ye, Y.; Cao, J. Characterization of binding interactions of (-)-epigallocatechin-3-gallate from green tea and lipase. J. Agric. Food Chem. 2013, 61, 8829–8835. [Google Scholar] [CrossRef]
- Hussein, L.; Abbas, H. Nitrogen balance studies among boys fed combinations of faba beans and wheat differing in polyphenolic contents. Nutr. Rep. Int. 1985, 31, 67–81. [Google Scholar]
- Naz, S.; Siddiqi, R.; Dew, T.P.; Williamson, G. Epigallocatechin-3-gallate inhibits lactase but is alleviated by salivary proline-rich proteins. J. Agric. Food Chem. 2011, 59, 2734–2738. [Google Scholar] [CrossRef]
- He, Q.; Lv, Y.; Yao, K. Food Chemistry Effects of tea polyphenols on the activities of alpha-amylase, pepsin, trypsin and lipase. Food Chem. 2006, 101, 1178–1182. [Google Scholar] [CrossRef]
- Tantoush, Z.; Apostolovic, D.; Kravic, B.; Prodic, I.; Mihajlovic, L.; Stanic-Vucinic, D.; Cirkovic Velickovic, T. Green tea catechins of food supplements facilitate pepsin digestion of major food allergens, but hampers their digestion if oxidized by phenol oxidase. J. Funct. Foods 2012, 4, 650–660. [Google Scholar] [CrossRef]
- Martinez-Gonzalez, A.; Diaz-Sanchez, A.; de la Rosa, L.; Vargas-Requena, C.; Bustos-Jaimes, I.; Alvarez-Parrilla, E. Polyphenolic Compounds and Digestive Enzymes: In Vitro Non-Covalent Interactions. Molecules 2017, 22, 669. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.Z.; Li, Y.Y. Inflammatory bowel disease: Pathogenesis. World Gastroenterol. 2014, 20, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Vezza, T.; Rodríguez-Nogales, A.; Algieri, F.; Utrilla, M.P.; Rodriguez-Cabezas, M.E.; Galvez, J. Flavonoids in Inflammatory Bowel Disease: A Review. Nutrients 2016, 8, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, C.N. Treatment of IBD: Where We Are and Where We Are Going. Am. J. Gastroenterol. 2015, 110, 114–126. [Google Scholar] [CrossRef]
- Brückner, M.; Westphal, S.; Domschke, W.; Kucharzik, T.; Lügering, A. Green tea polyphenol epigallocatechin-3-gallate shows therapeutic antioxidative effects in a murine model of colitis. J. Crohn’s Colitis 2012, 6, 226–235. [Google Scholar] [CrossRef] [Green Version]
- Oz, H.S.; Chen, T.; De Villiers, W.J.S. Green tea polyphenols and sulfasalazine have parallel anti-inflammatory properties in colitis models. Front. Immunol. 2013, 4, 132. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; De Villiers, W.J.; McClain, C.J.; Varilek, G.W. Green Tea Polyphenols Block Endotoxin-Induced Tumor Necrosis Factor- Production and Lethality in a Murine Model. J. Nutr. 1998, 128, 2334–2340. [Google Scholar] [CrossRef]
- Sergent, T.; Piront, N.; Meurice, J.; Toussaint, O.; Schneider, Y.J. Anti-inflammatory effects of dietary phenolic compounds in an in vitro model of inflamed human intestinal epithelium. Chem. Biol. Interact. 2010, 188, 659–667. [Google Scholar] [CrossRef]
- Al-Sadi, R.; Ye, D.; Boivin, M.; Guo, S.; Hashimi, M.; Ereifej, L.; Ma, T.Y. Interleukin-6 Modulation of Intestinal Epithelial Tight Junction Permeability Is Mediated by JNK Pathway Activation of Claudin-2 Gene. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claud, E.C.; Savidge, T.; Walker, W.A. Modulation of Human Intestinal Epithelial Cell IL-8 Secretion by Human Milk Factors. Pediatr. Res. 2003, 53, 419–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Zamora-Ros, R.; Chan, S.; Cross, A.J.; Ward, H.; Jakszyn, P.; Luben, R.; Opstelten, J.L.; Oldenburg, B.; Hallmans, G.; et al. Dietary Polyphenols in the Aetiology of Crohn’s Disease and Ulcerative Colitis-A Multicenter European Prospective Cohort Study (EPIC). Inflamm. Bowel Dis. 2017, 23, 2072–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Venancio, V.P.; Fang, C.; Dupont, A.W.; Talcott, S.T.; Mertens-Talcott, S.U. Mango (Mangifera indica L.) polyphenols reduce IL-8, GRO, and GM-SCF plasma levels and increase Lactobacillus species in a pilot study in patients with inflammatory bowel disease. Nutr. Res. 2020, 75, 85–94. [Google Scholar] [CrossRef]
- Roth, S.; Spalinger, M.R.; Gottier, C.; Biedermann, L.; Zeitz, J.; Lang, S.; Weber, A.; Rogler, G.; Scharl, M. Bilberry-derived anthocyanins modulate cytokine expression in the intestine of patients with ulcerative colitis. PLoS ONE 2016, 11, e0154817. [Google Scholar] [CrossRef] [Green Version]
- Katayama, S.; Kukita, T.; Ishikawa, E.; Nakashima, S.; Masuda, S.; Kanda, T.; Akiyama, H.; Teshima, R.; Nakamura, S. Apple polyphenols suppress antigen presentation of ovalbumin by THP-1-derived dendritic cells. Food Chem. 2013, 138, 757–761. [Google Scholar] [CrossRef]
- Chung, S.Y.; Reed, S. Removing peanut allergens by tannic acid. Food Chem. 2012, 134, 1468–1473. [Google Scholar] [CrossRef]
- Plundrich, N.J.; White, B.L.; Dean, L.L.; Davis, J.P.; Foegeding, E.A.; Ann, M. Stability and immunogenicity of hypoallergenic peanut protein–polyphenol complexes during in vitro pepsin digestion. Food Funct. 2015, 6, 2145–2154. [Google Scholar] [CrossRef]
- Cattaneo, A.; Ballabio, C.; Bertelli, A.; Fiocchi, A.; Galli, C.; Isoardi, P.; Terracciano, L.; Restani, P. Evaluation of Residual Immunoreactivity in Red and White Wines Clarified with Gluten or Gluten Derivatives. Int. J. Tissue React. 2003, 25, 57–64. [Google Scholar]
- Restani, P.; Beretta, B.; Ballabio, C.; Galli, C.; Bertelli, A. Evaluation by SDS-PAGE and Immunoblotting of Residual Antigenicity in Gluten-Treated Wine: A Preliminary Study. Int. J. Tissue React. 2002, 24, 45–51. [Google Scholar]
- Tozzi, S.; Zanna, N.; Taddei, P. Study on the interaction between gliadins and a coumarin as molecular model system of the gliadins–anthocyanidins complexes. Food Chem. 2013, 141, 3586–3597. [Google Scholar] [CrossRef] [PubMed]
- Taddei, P.; Zanna, N.; Tozzi, S. Raman characterization of the interactions between gliadins and anthocyanins. J. Raman Spectrosc. 2013, 44. [Google Scholar] [CrossRef]
- Mazzaracchio, P.; Tozzi, S.; Boga, C.; Forlani, L.; Pifferi, P.G.; Barbiroli, G. Interaction between gliadins and anthocyan derivatives. Food Chem. 2011, 129, 1100–1107. [Google Scholar] [CrossRef] [PubMed]
- Mazzaracchio, P.; Kindt, M.; Pifferi, P.G.; Tozzi, S.; Barbiroli, G. Adsorption behaviour of some anthocyanins by wheat gluten and its fractions in acidic conditions. Int. J. Food Sci. Technol. 2012, 47, 390–398. [Google Scholar] [CrossRef]
- Dias, R.; Perez-Gregorio, R.; Mateus, N.; Freitas, V. De The interaction between tannins and gliadin derived peptides in a celiac disease perspective. RSC Adv. 2015, 5, 32151–32158. [Google Scholar] [CrossRef]
- Girard, A.L.; Bean, S.R.; Tilley, M.; Adrianos, S.L.; Awika, J.M. Interaction mechanisms of condensed tannins (proanthocyanidins) with wheat gluten proteins. Food Chem. 2018, 245, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Pérot, M.; Lupi, R.; Guyot, S.; Delayre-Orthez, C.; Gadonna-Widehem, P.; Thébaudin, J.Y.; Bodinier, M.; Larré, C. Polyphenol Interactions Mitigate the Immunogenicity and Allergenicity of Gliadins. J. Agric. Food Chem. 2017, 65, 6442–6451. [Google Scholar] [CrossRef] [PubMed]
- Plundrich, N.J.; Cook, B.T.; Maleki, S.J.; Fourches, D.; Lila, M.A. Binding of peanut allergen Ara h 2 with Vaccinium fruit polyphenols. Food Chem. 2019, 284, 287–295. [Google Scholar] [CrossRef]
- Ognjenović, J.; Stojadinović, M.; Milčić, M.; Apostolović, D.; Vesić, J.; Stambolić, I.; Atanasković-Marković, M.; Simonović, M.; Velickovic, T.C. Interactions of epigallo-catechin 3-gallate and ovalbumin, the major allergen of egg white. Food Chem. 2014, 164, 36–43. [Google Scholar] [CrossRef]
- Dias, R.; Brás, N.F.; Pérez-Gregorio, M.; Fernandes, I.; Mateus, N.; Freitas, V. A multi-spectroscopic study on the interaction of food polyphenols with a bioactive gluten peptide: From chemistry to biological implications. Food Chem. 2019, 299, 125051. [Google Scholar] [CrossRef]
- Dias, R.; Brás, N.F.; Fernandes, I.; Pérez-Gregorio, M.; Mateus, N.; Freitas, V. Molecular insights on the interaction and preventive potential of epigallocatechin-3-gallate in Celiac Disease. Int. J. Biol. Macromol. 2018, 112, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- van de Wal, Y.; Kooy, Y.M.; Drijfhout, J.W.; Amons, R.; Koning, F. Peptide binding characteristics of the coeliac disease-associated DQ(aII*0501, 1*0201) molecule. Immunogenetics 1996, 44, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Vader, L.W.; de Ru, A.; van der Wal, Y.; Kooy, Y.M.; Benckhuijsen, W.; Mearin, M.L.; Drijfhout, J.W.; van Veelen, P.; Koning, F. Specificity of tissue transglutaminase explains cereal toxicity in celiac disease. J. Exp. Med. 2002, 195, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Sambuy, Y.; De Angelis, I.; Ranaldi, G.; Scarino, M.; Stammati, A.; Zucco, F. The Caco-2 cell line as a model of the intestinal barrier: Influence of cell and culture-related factors on Caco-2 cell functional characteristics. Cell Biol. Toxicol. 2005, 21, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Giovannini, C.; Sanchez, M.; Straface, E.; Scazzocchio, B.; Silano, M.; De Vincenzi, M. Induction of apoptosis in Caco-2 cells by wheat gliadin peptides. Toxicology 2000, 145, 63–71. [Google Scholar] [CrossRef]
- Capozzi, A.; Vincentini, O.; Gizzi, P.; Porzia, A.; Longo, A.; Felli, C.; Mattei, V.; Mainiero, F.; Silano, M.; Sorice, M.; et al. Modulatory Effect of Gliadin Peptide 10-mer on Epithelial Intestinal CACO-2 Cell Inflammatory Response. PLoS ONE 2013, 8, e66561. [Google Scholar] [CrossRef] [Green Version]
- Kramer, K.; Yeboah-Awudzi, M.; Magazine, N.; King, J.M.; Xu, Z.; Losso, J.N. Procyanidin B2 rich cocoa extracts inhibit inflammation in Caco-2 cell model of in vitro celiac disease by down-regulating interferon-gamma- or gliadin peptide 31-43-induced transglutaminase-2 and interleukin-15. J. Funct. Foods 2019, 57, 112–120. [Google Scholar] [CrossRef]
- Bitzer, Z.T.; Elias, R.J.; Vijay-Kumar, M.; Lambert, J.D. (-)-Epigallocatechin-3-gallate decreases colonic inflammation and permeability in a mouse model of colitis, but reduces macronutrient digestion and exacerbates weight loss. Mol. Nutr. Food Res. 2016, 60, 2267–2274. [Google Scholar] [CrossRef]
- González-Quilen, C.; Grau-Bové, C.; Jorba-Martín, R.; Caro-Tarragó, A.; Pinent, M.; Ardévol, A.; Beltrán-Debón, R.; Terra, X.; Blay, M.T. Protective properties of grape-seed proanthocyanidins in human ex vivo acute colonic dysfunction induced by dextran sodium sulfate. Eur. J. Nutr. 2020. [Google Scholar] [CrossRef]
- Daum, S.; Bauer, U.; Foss, H.D.; Schuppan, D.; Stein, H.; Riecken, E.O.; Ullrich, R. Increased expression of mRNA for matrix metalloproteinases-1 and -3 and tissue inhibitor of metalloproteinases-1 in intestinal biopsy specimens from patients with coeliac disease. Gut 1999, 44, 17–25. [Google Scholar] [CrossRef]
- Manavalan, J.S.; Hernandez, L.; Shah, J.G.; Konikkara, J.; Naiyer, A.J.; Lee, A.R.; Ciaccio, E.; Minaya, M.T.; Green, P.; Bhagat, G. Serum cytokine elevations in celiac disease: Association with disease presentation. Hum. Immunol. 2010, 71, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Cinova, J.; Palová-Jelinková, L.; Smythies, L.E.; Černá, M.; Pecharová, B.; Dvorák, M.; Fruhauf, P.; Tlaskalová-Hogenová, H.; Smith, P.D.; Tucková, L. Gliadin Peptides Activate Blood Monocytes from Patients with Celiac Disease. J. Clin. Immunol. 2007, 27, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Jimenez, N.; Castellanos-Rubio, A.; Plaza-Izurieta, L.; Irastorza, I.; Elcoroaristizabal, X.; Jauregi-Miguel, A.; Lopez-Euba, T.; Tutau, C.; de Pancorbo, M.M.; Vitoria, J.C.; et al. Coregulation and modulation of NFκB-related genes in celiac disease: Uncovered aspects of gut mucosal inflammation. Hum. Mol. Genet. 2014, 23, 1298–1310. [Google Scholar] [CrossRef] [PubMed]
- Vincentini, O.; Maialetti, F.; Gonnelli, E.; Silano, M. Gliadin-dependent cytokine production in a bidimensional cellular model of celiac intestinal mucosa. Clin. Exp. Med. 2015, 15, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Lammers, K.M.; Khandelwal, S.; Chaudhry, F.; Kryszak, D.; Puppa, E.L.; Casolaro, V.; Fasano, A. Identification of a novel immunomodulatory gliadin peptide that causes interleukin-8 release in a chemokine receptor CXCR3-dependent manner only in patients with coeliac disease. Immunology 2011, 132, 432–440. [Google Scholar] [CrossRef]
- Molberg, Ø.; McAdam, S.N.; Sollid, L.M. Role of Tissue Transglutaminase in Celiac Disease. J. Pediatr. Gastroenterol. Nutr. 2000, 30, 232–240. [Google Scholar] [CrossRef]
- Kagnoff, M.F. Celiac disease: Pathogenesis of a model immunogenetic disease. J. Clin. Investig. 2007, 117, 41–49. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Intervention | Outcomes | Identifier and Phase |
|---|---|---|
| Probiotic | ||
| Lactobacillus plantarum, Lactobacillus casei, Bifidobacterium breve, Bifidobacterium animalis | Gastrointestinal symptom improvement | NCT01699191 |
| Pentabiocel (probiotic cocktail) | 1°: Changes in celiac symptom index; 2°: Changes in serology, BMI | NCT03857360 |
| Lactobacilli culture | Anti-TG2 antibodies | NCT03176095 1 |
| BL NCC 2705 | Safety and tolerability | NCT03775499 |
| L. paracasei, L. plantarum | 1°: Serum antibodies for TG2, GADA, IA-2, IAA, ZnT8a, TPOA; 2°: Concentration of gluten peptides in urine | NCT04014660 |
| Bifidobacterium infantis | 1°: Gastrointestinal symptom improvement; 2°: Changes in celiac symptom index, fecal microbiome diversity, changes in gluten immunogenic peptides, serology, anthropometric measurements | NCT03271138 1 Phase 2 |
| 1°: Decreased intestinal permeability; 2°: Changes in cytokine profile | NCT01257620 1 | |
| Prebiotic | ||
| Inulin-type fructans | 1°: Decreased intestinal permeability; 2°: Adverse events, short-chain fatty acids, molecular characteristics of fecal microbiota | NCT03064997 1 |
| Antibiotic | ||
| Rifaximin | 1°: Gastrointestinal symptom improvement; 2°: Decreased small intestine bacterial overgrowth | NCT01137955 1 |
| Parabiotic | ||
| Necator americanus infection | 1°: Histopathology (villous height:crypt depth); 2°: IEL counts over the course of treatment, gastrointestinal symptom improvement, anti-TG2 antibodies | NCT02754609 1 Phase 1b |
| 1°: Histopathology (villous height:crypt depth); 2°: IEL count, Marsh score, anti-TG2 antibodies | NCT01661933 1 Phase 1/2 | |
| 1°: Marsh score; 2°: PBMC gluten recognition, T cell proliferation, cytokine profiles | NCT00671138 1 Phase 2 | |
| Anti-Inflammatory | ||
| PTG-100 | 1°: Histopathology (villous height:crypt depth); 2°: Changes in anti-TG2, antiDGP antibodies, CD3-positive IEL density, changes in celiac symptom index | NCT04524221 Phase 1 |
| Anti-IL-15 Antibody | ||
| Hu-Mik-β-1 | Safety in celiac disease patients | NCT01893775 1 Phase 1 |
| CALY-002 | Safety and tolerability | NCT04593251 Phase 1 |
| PRV-015 | 1°: Celiac disease patient-reported outcomes; 2°: IEL density, safety and tolerability, pharmacokinetic analysis, anti-PRV-015 antibodies | NCT04424927 Phase 2 |
| AMG 714 | 1°: Reduction in IELs; 2°: Improvement of histopathology, gastrointestinal symptom improvement | NCT02633020 1 Phase 2a |
| TM-β1 | 1°: Attenuation of mucosal injury; 2°: Decreased IEL infiltration, attenuation of anti-gliadin and anti-TG2 serum antibodies, gastrointestinal symptom improvement | NCT02637141 1 Phase 2a |
| Immunomodulators | ||
| Cathepsin S inhibitor (RG 7625) | 1°: Decreased number of responses to gluten challenge; 2°: Number of patients with adverse events, attenuation of anti-TG2 and anti-gliadin antibodies, improved lactulose/mannitol ratio, decreased circulating white blood cells, concentrations of CD74 B cells | NCT02679014 1 Phase 1 |
| Vedolizumab | Histopathology | NCT02929316 2 Phase 2 |
| Nexvax2 | Safety and tolerability | NCT00879749 1 Phase 1 |
| 1°: Safety and tolerability, intervention bioavailability; 2°: Pharmacodynamic measures, pharmacokinetic analysis | NCT03543540 1 Phase 1 | |
| 1°: Safety in celiac disease patients; 2°: Gastrointestinal symptom improvement, plasma cytokine levels | NCT02528799 1 Phase 1 | |
| 1°: Celiac disease patient-reported outcomes; 2°: Pharmacodynamic measures of immune activation, gastrointestinal symptoms, safety and tolerability | NCT03644069 Phase 2 | |
| Vercirnon (CCX282-B) | 1°: Histopathology (villous height:crypt depth); 2°: Small intestinal inflammation, gluten-induced celiac-type serology, gastrointestinal symptom improvement | NCT00540657 1 Phase 2 |
| Short-course steroids (Prednisolone) | 1°: Gastrointestinal symptom improvement, histopathology; 2°: Maintenance of histopathology | NCT01045837 1 Phase 2/3 |
| KAN-101 | 1°: Safety and tolerability; 2°: Pharmacokinetic analysis | NCT04248855 Phase 1 |
| TIMP-GLIA | 1°: safety and tolerability, clinically significant change in physical examination, vital signs; 2°: Pharmacokinetic analysis | NCT03486990 1 Phase 1 |
| 1°: Changes in baseline plasma IFN-ɣ; 2°: Changes in gliadin-specific T cell proliferation, cytokine secretion, changes from baseline in T cells, histopathology (villous height:crypt depth), changes in number of IELs, plasma concentration of intervention | NCT03738475 1 Phase 2 | |
| TAK-101 | 1°: Changes in baseline plasma IFN-ɣ; 2°: Safety and tolerability, changes in celiac symptom index, changes from baseline IL-2, pharmacodynamic measures | NCT04530123 Phase 2 |
| Zonulin Antagonist | ||
| Larazotide acetate | 1°: Safety and tolerance, pharmacokinetic analysis, intestinal permeability, zonulin levels; 2°: Gastrointestinal symptoms Pharmacodynamic measures | NCT00386165 1 Phase 1 |
| 1°: Safety and tolerability; 2°: Intestinal permeability, celiac disease signs and symptoms | NCT00362856 1 Phase 2 | |
| 1°: Intestinal permeability; 2°: Composite celiac disease activity index, adverse events | NCT004929601 Phase 2 | |
| 1°: Response to gluten; 2°: Attenuation of anti-TG2 antibodies | NCT00889473 1 Phase 2 | |
| 1°: Histopathology (villous height:crypt depth); 2°: Safety and tolerability | NCT006204511 Phase 2 | |
| Gastrointestinal symptom improvement | NCT01396213 1 Phase 2b | |
| Celiac disease patient-reported outcomes | NCT03569007 Phase 3 | |
| TG2 Inhibitor | ||
| GSK3915393 | 1°: Safety and tolerability, clinically significant change in vital signs, physical examination; 2°: Pharmacokinetic analysis | NCT04604795 Phase 1 |
| Dietary Replacement | ||
| Triticum monococcum | 1°: Histopathology; 2°: Attenuation of anti-TG2 and anti-endomysial antibodies, gastrointestinal symptom improvement | NCT02220166 1 Phase 2 |
| Microbial transglutaminase-treated wheat flour | 1°: Attenuation of anti-TG2, anti-endomysium antibodies, histopathology (villous height:crypt depth, IEL count, Marsh–Oberhuber score); 2°: Gastrointestinal symptom improvement | NCT02472119 Phase 2 |
| Gluten-friendly bread | 1°: Changes in serology; 2°: Fecal microbiome diversity, gastrointestinal symptoms, lactulose/mannitol excretion ratio | NCT03168490 3 |
| Small bowel mucosal density, anti-TG2 IgA EMA and AGA antibody levels | NCT03137862 1 | |
| Protalsafe | 1°: Lactulose/mannitol excretion ratio, serum zonulin 2°: Fecal microbiome profile, gastrointestinal symptoms, quality of life, changes from baseline in serum cytokines | NCT03483805 1 |
| Enzyme Supplement | ||
| Aspergillus niger prolyl endoprotease | 1°: Histopathology/Marsh score, attenuation of anti-TG2, anti-endomysial and anti-gliadin antibodies; 2°: Presence and activity of gluten-reactive T cells, immunophenotype of lymphocytes, clinical symptoms after gluten challenge | NCT00810654 1 Phase 1/2 |
| Pancreatic enzyme | 1°: Gastrointestinal symptom improvement; 2°: Fecal elastase measurement | NCT02475369 2 Phase 4 |
| Proprietary food-grade enzyme blend | 1°: Decreased anti-TG2 antibodies; 2°: Decreased anti-endomysial antibodies, decreased celiac-related antibodies in blood, change in symptoms, including skin rash, histopathology improvement | NCT00962182 1 Phase 1/2 |
| ALV003 protease | 1°: Clinical symptoms, celiac-specific serology; 2°: Immunotoxic epitope profile of digests | NCT00859391 1 Phase 1 |
| Safety and tolerability | NCT00626184 1 Phase 1 | |
| Safety and tolerability | NCT00669825 1 Phase 1 | |
| 1°: Histopathology, tolerability; 2°: IEL count/phenotype, serological markers | NCT01255696 1 Phase 2 | |
| 1°: Histopathology (villous height:crypt depth); 2°: IEL density, safety and tolerability, celiac-specific serology | NCT01917630 Phase 2 | |
| 1°: Histopathology, tolerability; 2°: IEL count/phenotype, serological markers | NCT00959114 1 Phase 2a | |
| PvP001, PvP002, PvP003 | 1°: Safety and tolerability; 2°: Pharmacokinetic analysis | NCT03701555 Phase 1 |
| Latiglutenase | 1°: Histopathology; 2°: Gastrointestinal symptom improvement | NCT03585478 Phase 2 |
| 1°: Gastrointestinal symptom improvement, gluten degradation after a study meal; 2°: Health-related quality of life, gluten degradation after a study meal, clinically significant change in vital signs and physical examination | NCT04243551 Phase 2 | |
| Synthetic Sequestrant | ||
| BL-7010 | 1°: Safety and tolerability; 2°: Plasma levels of BL-7010 | NCT01990885 1 Phase 1/2 |
| Anti-Gliadin Antibody Supplement | ||
| AGY | 1°: Safety and tolerability; 2°: Gastrointestinal symptoms | NCT01765647 1 Phase 1 |
| Changes in celiac-related symptoms | NCT03707730 Phase 2 | |
| Protective Effects | Gliadin Source | Polyphenol Classification | Reference |
|---|---|---|---|
| Improved barrier integrity; reduced secretion of IL-6, IL-8 | Pepsin-trypsin-digested gliadin | Green tea extract (653 mg g−1 catechins inc. 413 mg g−1 EGCG) | Van Buiten, Lambert and Elias (2018) |
| Reduced paracellular transport of gliadins | p58–89 | EGCG | Dias, Brás, Fernandes, Pérez-Gregorio, Mateus and Freitas (2018) |
| Procyanidin B3, procyanidin C2 | Dias, Brás, Pérez-Gregorio, Fernandes, Mateus and Freitas (2019) | ||
| Reduced TG2 and COX-2 expression, reduced secretion of IL-15, IL-1β, IL-6, IL-8 | p31–43 | Cocoa extract (21.39 mg g−1 procyanidin B2) | Kramer, Yeboah-Awudzi, Magazine, King and Xu (2020) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Buiten, C.B.; Elias, R.J. Gliadin Sequestration as a Novel Therapy for Celiac Disease: A Prospective Application for Polyphenols. Int. J. Mol. Sci. 2021, 22, 595. https://doi.org/10.3390/ijms22020595
Van Buiten CB, Elias RJ. Gliadin Sequestration as a Novel Therapy for Celiac Disease: A Prospective Application for Polyphenols. International Journal of Molecular Sciences. 2021; 22(2):595. https://doi.org/10.3390/ijms22020595
Chicago/Turabian StyleVan Buiten, Charlene B., and Ryan J. Elias. 2021. "Gliadin Sequestration as a Novel Therapy for Celiac Disease: A Prospective Application for Polyphenols" International Journal of Molecular Sciences 22, no. 2: 595. https://doi.org/10.3390/ijms22020595
APA StyleVan Buiten, C. B., & Elias, R. J. (2021). Gliadin Sequestration as a Novel Therapy for Celiac Disease: A Prospective Application for Polyphenols. International Journal of Molecular Sciences, 22(2), 595. https://doi.org/10.3390/ijms22020595