
Purinergic Receptor Blockade with Suramin Increases Survival of Postnatal Neural Progenitor Cells In Vitro

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
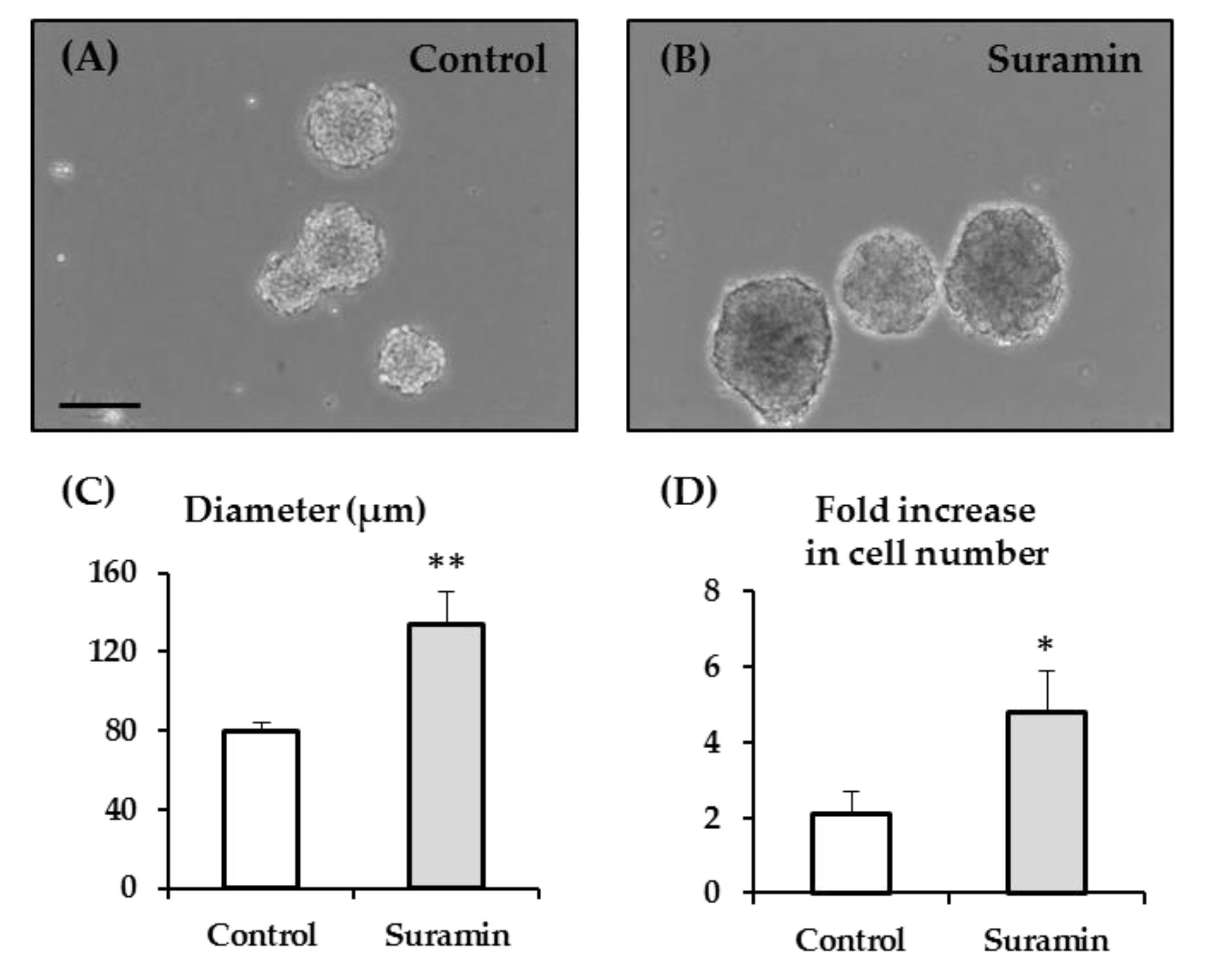
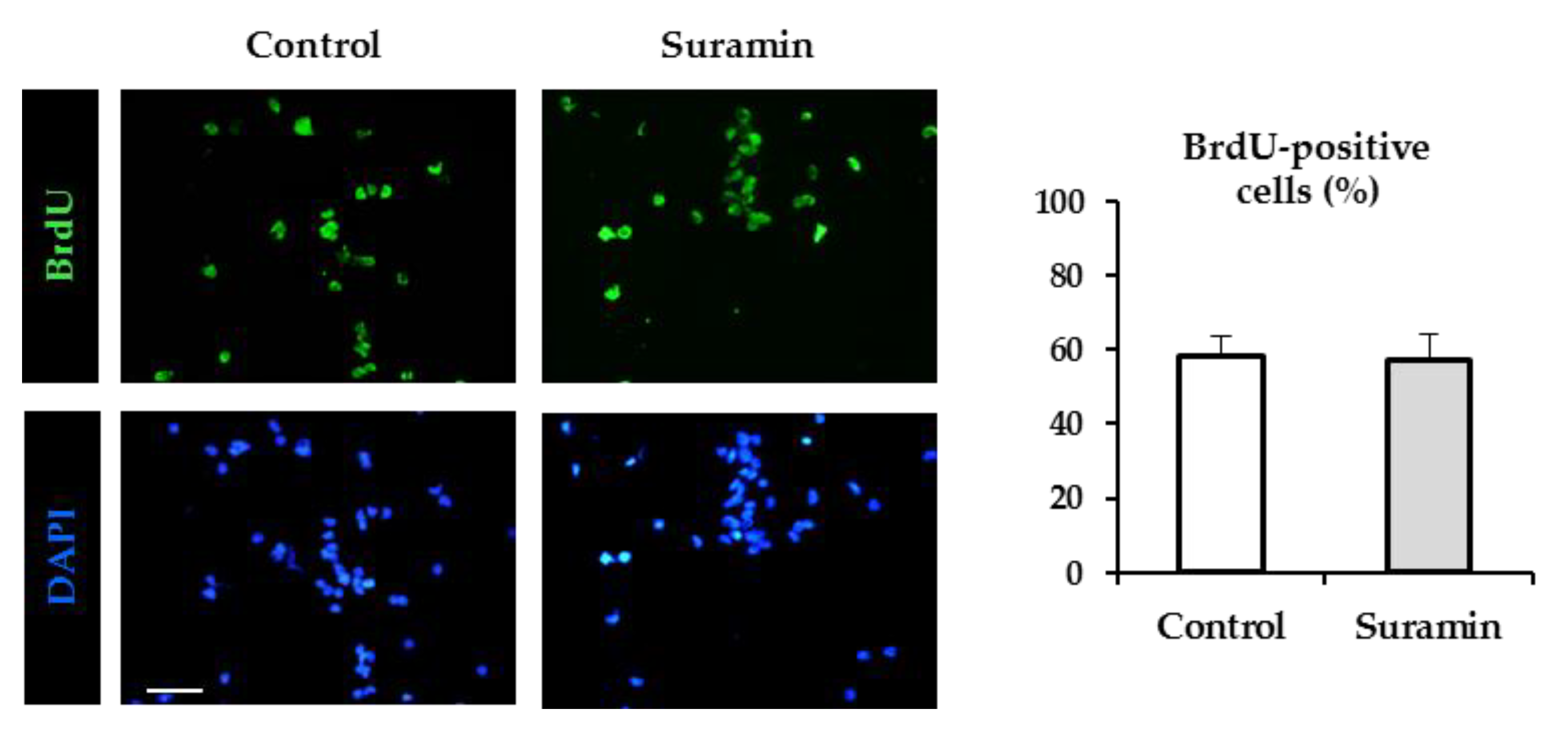
2.1. Suramin Treatment of NPC Cultures Induces an Increase in Neurosphere Size and in Cell Number Which is Not Attributable to a Higher NPC Proliferation Rate or to a Higher Incidence of Neurosphere Fusion
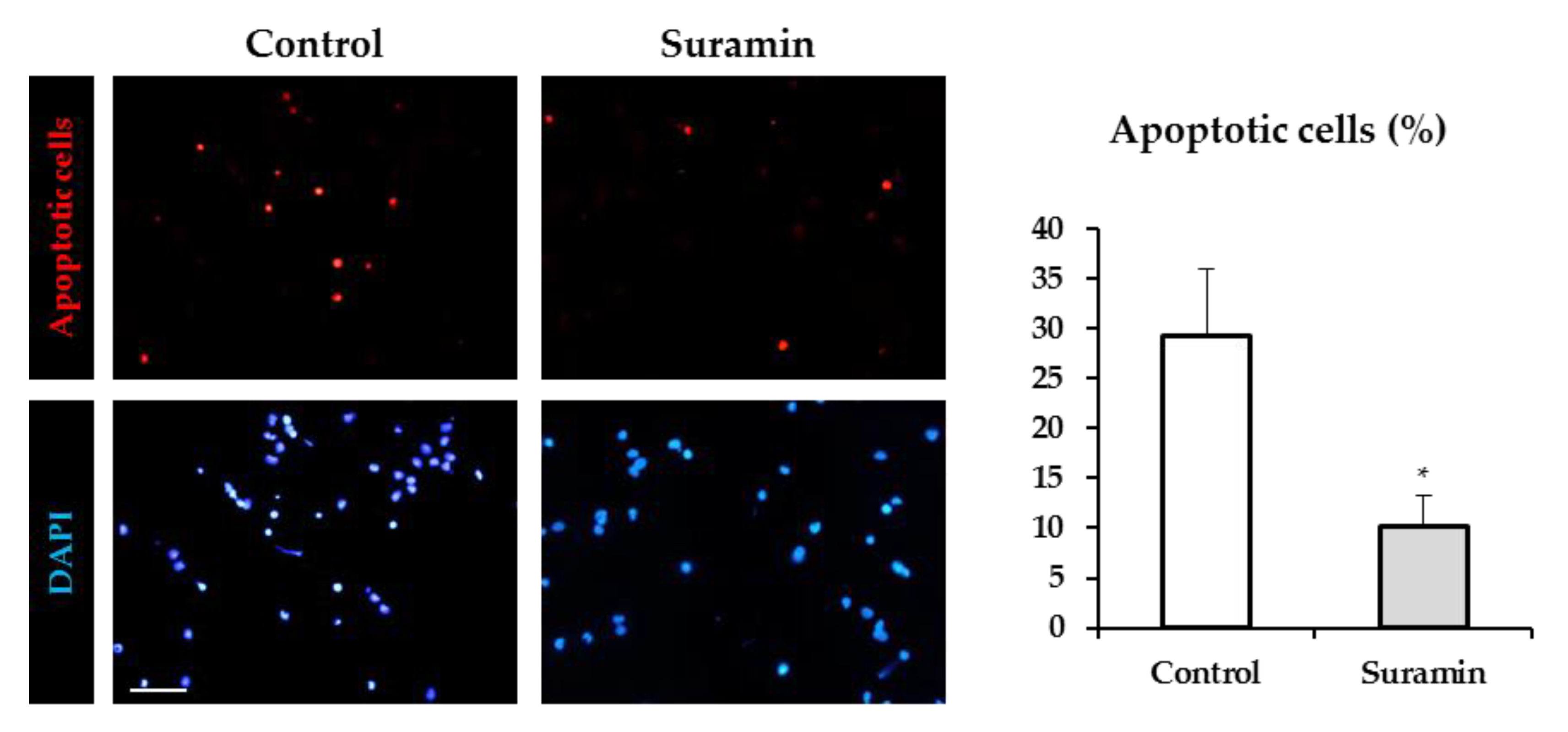
2.2. Suramin Treatment Reduces Apoptosis in NPCs
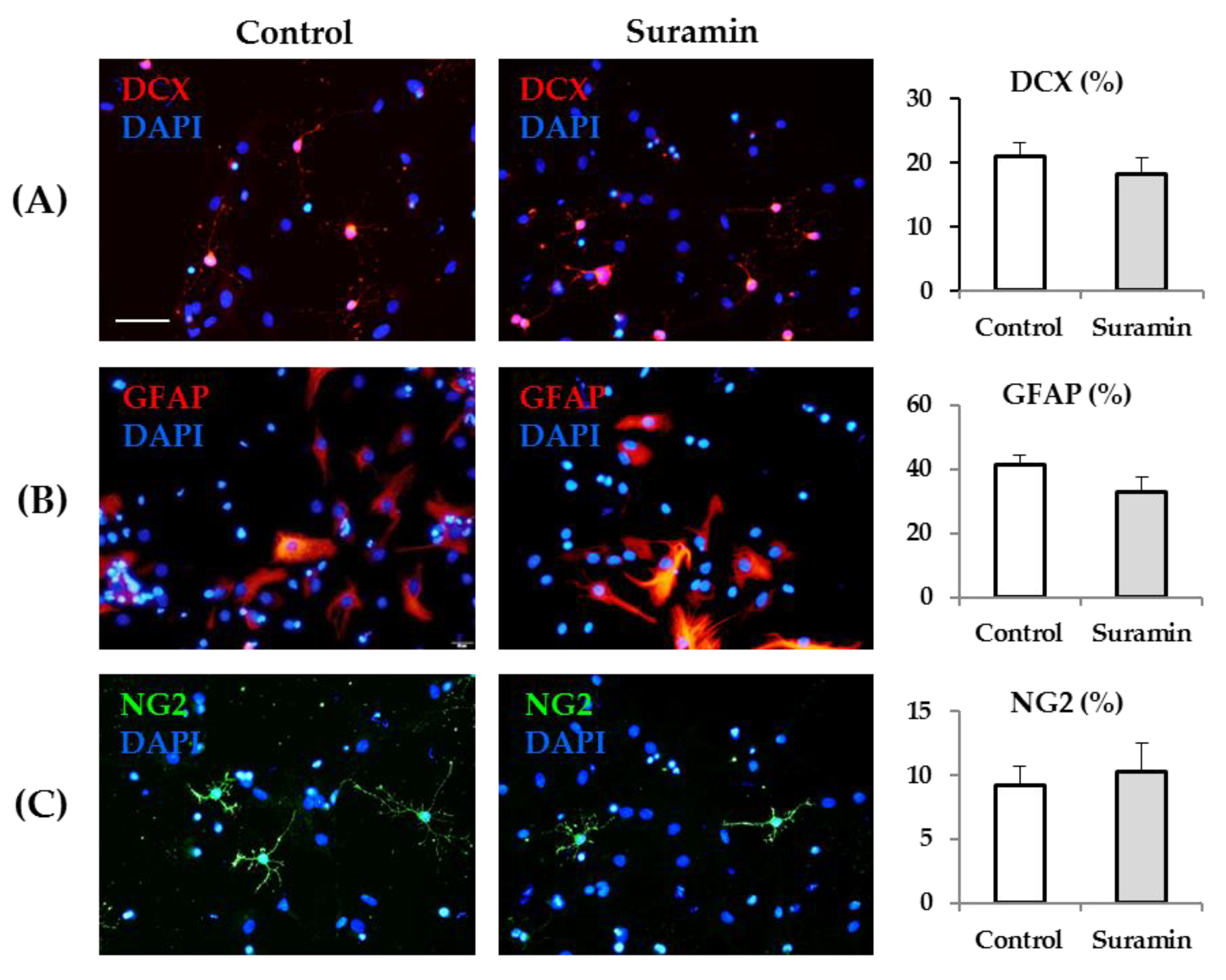
2.3. Neural Lineage Potential is Not Affected in Neurospheres after Suramin Treatment
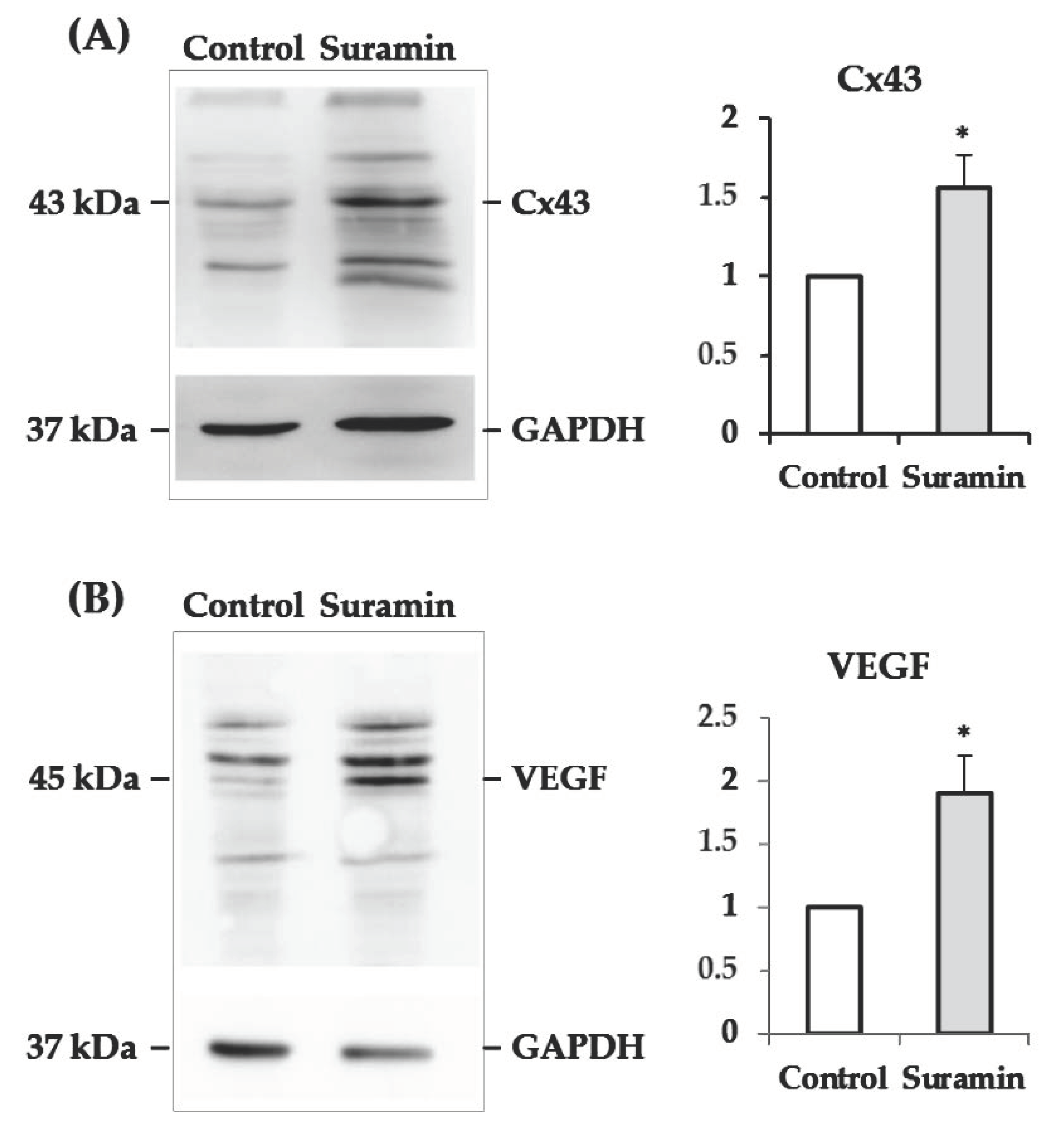
2.4. The Levels of Connexin43 and Vascular Endothelial Growth Factor Increase in Neurospheres after Suramin Treatment
3. Discussion
4. Materials and Methods
4.1. Neurosphere Culture
4.2. Analysis of Neurosphere Diameter, Bromodeoxyuridine (BrdU) Incorporation and Apoptosis
4.3. Time-Lapse Microscopy
4.4. Analysis of Differentiation of Neurosphere-Derived Cells
4.5. Epifluorescence Microscopy
4.6. Western Blot
4.7. Statistics
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Gage, F.H. Mammalian Neural Stem Cells. Science 2000, 287, 1433–1438. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Buylla, A.; Seri, B.; Doetsch, F. Identification of neural stem cells in the adult vertebrate brain. Brain Res. Bull. 2002, 57, 751–758. [Google Scholar] [CrossRef]
- Anderson, D.J. Stem cells and pattern formation in the nervous system: The possible versus the actual. Neuron 2001, 30, 19–35. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, B.A.; Weiss, S. Generation of neurons and astrocytes from isolated cells of the adult mammalian central nervous system. Science 1992, 255, 1707–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenter, M.K.; Cui, X.; Hu, Z.; Jackson, J.; Sherman, S.; Seiger, Å.; Wahlberg, L.U. In Vitro Expansion of a Multipotent Population of Human Neural Progenitor Cells. Exp. Neurol. 1999, 158, 265–278. [Google Scholar] [CrossRef]
- Vescovi, A.L.; Reynolds, B.A.; Fraser, D.D.; Weiss, S. bFGF regulates the proliferative fate of unipotent (neuronal) and bipotent (neuronal/astroglial) EGF-generated CNS progenitor cells. Neuron 1993, 11, 951–966. [Google Scholar] [CrossRef]
- Richardson, R.M.; Broaddus, W.C.; Holloway, K.L.; Fillmore, H.L. Grafts of adult subependymal zone neuronal progenitor cells rescue hemiparkinsonian behavioral decline. Brain Res. 2005, 1032, 11–22. [Google Scholar] [CrossRef]
- Ziv, Y.; Avidan, H.; Pluchino, S.; Martino, G.; Schwartz, M. Synergy between immune cells and adult neural stem progenitor cells promotes functional recovery from spinal cord injury. Proc. Natl. Acad. Sci. USA 2006, 103, 13174–13179. [Google Scholar] [CrossRef] [Green Version]
- Hicks, A.U.; Hewlett, K.; Windle, V.; Chernenko, G.; Ploughman, M.; Jolkkonen, J.; Weiss, S.; Corbett, D. Enriched environment enhances transplanted subventricular zone stem cell migration and functional recovery after stroke. Neuroscience 2007, 136, 31–40. [Google Scholar] [CrossRef]
- Cusimano, M.; Biziato, D.; Brambilla, E.; Donegà, M.; Alfaro-Cervello, C.; Snider, S.; Salani, G.; Pucci, F.; Comi, G.; Garcia-Verdugo, J.M.; et al. Transplanted neural stem/precursor cells instruct phagocytes and reduce secondary tissue damage in the injured spinal cord. Brain 2012, 135, 447–460. [Google Scholar] [CrossRef] [Green Version]
- Miltiadous, P.; Kouroupi, G.; Stamatakis, A.; Koutsoudaki, P.N.; Matsas, R.; Stylianopoulou, F. Subventricular zone-derived neural stem cell grafts protect against hippocampal degeneration and restore cognitive function in the mouse following intrahippocampal kainic acid administration. Stem Cells Transl. Med. 2013, 2, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Morado-Díaz, C.J.; Matarredona, E.R.; Morcuende, S.; Talaverón, R.; Davis-López de Carrizosa, M.A.; de la Cruz, R.R.; Pastor, A.M. Neural progenitor cell implants in the lesioned medial longitudinal fascicle of adult cats regulate synaptic composition and firing properties of abducens internuclear neurons. J. Neurosci. 2014, 34, 7007–7017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnstock, G.; Ulrich, H. Purinergic signaling in embryonic and stem cell development. Cell. Mol. Life Sci. 2011, 68, 1369–1394. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, H.; Abbracchio, M.P.; Burnstock, G. Extrinsic Purinergic Regulation of Neural Stem/Progenitor Cells: Implications for CNS Development and Repair. Stem Cell Rev. Rep. 2012, 8, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Weissman, T.A.; Riquelme, P.A.; Ivic, L.; Flint, A.C.; Kriegstein, A.R. Calcium waves propagate through radial glial cells and modulate proliferation in the developing neocortex. Neuron 2004, 43, 647–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köles, L.; Leichsenring, A.; Rubini, P.; Illes, P. P2 Receptor Signaling in Neurons and Glial Cells of the Central Nervous System. Adv. Pharmacol. 2011, 61, 441–493. [Google Scholar] [PubMed]
- Burnstock, G. Short-and long-term (Trophic) purinergic signalling. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150422. [Google Scholar] [CrossRef] [Green Version]
- Neary, J.T.; Zimmermann, H. Trophic functions of nucleotides in the central nervous system. Trends Neurosci. 2009, 32, 189–198. [Google Scholar] [CrossRef]
- Burnstock, G. Physiology and pathophysiology of purinergic neurotransmission. Physiol. Rev. 2007, 87, 659–797. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, H.; Zebisch, M.; Sträter, N. Cellular function and molecular structure of ecto-nucleotidases. Purinergic Signal. 2012, 8, 437–502. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.H.C.; Takano, T.; Arcuino, G.; Wang, X.; Hu, F.; Darzynkiewicz, Z.; Nunes, M.; Goldman, S.A.; Nedergaard, M. Purinergic signaling regulates neural progenitor cell expansion and neurogenesis. Dev. Biol. 2007, 302, 356–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gampe, K.; Stefani, J.; Hammer, K.; Brendel, P.; Pötzsch, A.; Enikolopov, G.; Enjyoji, K.; Acker-Palmer, A.; Robson, S.C.; Zimmermann, H. NTPDase2 and purinergic signaling control progenitor cell proliferation in neurogenic niches of the adult mouse brain. Stem Cells 2015, 33, 253–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langer, D.; Ikehara, Y.; Takebayashi, H.; Hawkes, R.; Zimmermann, H. The ectonucleotidases alkaline phosphatase and nucleoside triphosphate diphosphohydrolase 2 are associated with subsets of progenitor cell populations in the mouse embryonic, postnatal and adult neurogenic zones. Neuroscience 2007, 150, 863–879. [Google Scholar] [CrossRef] [PubMed]
- Stafford, M.R.; Bartlett, P.F.; Adams, D.J. Purinergic receptor activation inhibits mitogen-stimulated proliferation in primary neurospheres from the adult mouse subventricular zone. Mol. Cell. Neurosci. 2007, 35, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Benito-Muñoz, M.; Matute, C.; Cavaliere, F. Adenosine A1 receptor inhibits postnatal neurogenesis and sustains astrogliogenesis from the subventricular zone. Glia 2016, 64, 1465–1478. [Google Scholar] [CrossRef] [PubMed]
- Messemer, N.; Kunert, C.; Grohmann, M.; Sobottka, H.; Nieber, K.; Zimmermann, H.; Franke, H.; Nörenberg, W.; Straub, I.; Schaefer, M.; et al. P2X7 receptors at adult neural progenitor cells of the mouse subventricular zone. Neuropharmacology 2013, 73, 122–137. [Google Scholar] [CrossRef]
- Suyama, S.; Sunabori, T.; Kanki, H.; Sawamoto, K.; Gachet, C.; Koizumi, S.; Okano, H. Purinergic signaling promotes proliferation of adult mouse subventricular zone cells. J. Neurosci. 2012, 32, 9238–9247. [Google Scholar] [CrossRef] [Green Version]
- Leeson, H.C.; Kasherman, M.A.; Chan-Ling, T.; Lovelace, M.D.; Brownlie, J.C.; Toppinen, K.M.; Gu, B.J.; Weible, M.W. P2X7 receptors regulate phagocytosis and proliferation in adult hippocampal and SVZ neural progenitor cells: Implications for inflammation in neurogenesis. Stem Cells 2018, 36, 1764–1777. [Google Scholar] [CrossRef] [Green Version]
- Lacar, B.; Herman, P.; Platel, J.-C.; Kubera, C.; Hyder, F.; Bordey, A. Neural progenitor cells regulate capillary blood flow in the postnatal subventricular zone. J. Neurosci. 2012, 32, 16435–16448. [Google Scholar] [CrossRef]
- Yoon, M.J.; Lee, H.J.; Lee, Y.S.; Kim, J.H.; Park, J.K.; Chang, W.K.; Shin, H.C.; Kim, D.K. Extracellular ATP is involved in the induction of apoptosis in murine hematopoietic cells. Biol. Pharm. Bull. 2007, 30, 671–676. [Google Scholar] [CrossRef] [Green Version]
- Tchernookova, B.K.; Heer, C.; Young, M.; Swygart, D.; Kaufman, R.; Gongwer, M.; Shepherd, L.; Caringal, H.; Jacoby, J.; Kreitzer, M.A.; et al. Activation of retinal glial (Müller) cells by extracellular ATP induces pronounced increases in extracellular H+ flux. PLoS ONE 2018, 13, e0190893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tritsch, N.X.; Yi, E.; Gale, J.E.; Glowatzki, E.; Bergles, D.E. The origin of spontaneous activity in the developing auditory system. Nature 2007, 450, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purine and purinergic receptors. Brain Neurosci. Adv. 2018, 2, 239821281881749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravella, A.; Ringstedt, T.; Brion, J.; Pandolfo, M.; Herlenius, E. Adult neural precursor cells form connexin-dependent networks that improve their survival. Neuroreport 2015, 26, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Talaveron, R.; Matarredona, E.R.; de la Cruz, R.R.; Pastor, A.M. Neural progenitor cell implants modulate vascular endothelial growth factor and brain-derived neurotrophic factor expression in rat axotomized neurons. PLoS ONE 2013, 8, e54519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, K.; Mao, X.O.; Batteur, S.P.; McEachron, E.; Leahy, A.; Greenberg, D.A. Caspase-3 and the regulation of hypoxic neuronal death by vascular endothelial growth factor. Neuroscience 2001, 108, 351–358. [Google Scholar] [CrossRef]
- Volm, M.; Mattern, J.; Koomägi, R. Inverse correlation between apoptotic (Fas ligand, caspase-3) and angiogenic factors (VEGF, microvessel density) in squamous cell lung carcinomas. Anticancer Res. 1999, 19, 1669–1671. [Google Scholar]
- Talaverón, R.; Fernández, P.; Escamilla, R.; Pastor, A.M.; Matarredona, E.R.; Sáez, J.C. Neural progenitor cells isolated from the subventricular zone present hemichannel activity and form functional gap junctions with glial cells. Front. Cell Neurosci. 2015, 9, 411. [Google Scholar] [CrossRef] [Green Version]
- Delarasse, C.; Gonnord, P.; Galante, M.; Auger, R.; Daniel, H.; Motta, I.; Kanellopoulos, J.M. Neural progenitor cell death is induced by extracellular ATP via ligation of P2X7 receptor. J. Neurochem. 2009, 109, 846–857. [Google Scholar] [CrossRef] [PubMed]
- Suh, H.; Deng, W.; Gage, F.H. Signaling in adult neurogenesis. Annu. Rev. Cell Dev. Biol. 2009, 25, 253–275. [Google Scholar] [CrossRef]
- Boccazzi, M.; Rolando, C.; Abbracchio, M.P.; Buffo, A.; Ceruti, S. Purines regulate adult brain subventricular zone cell functions: Contribution of reactive astrocytes. Glia 2014, 62, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Braun, N.; Shukla, V.; Füllgrabe, M.; Schomerus, C.; Korf, H.-W.; Gachet, C.; Ikehara, Y.; Sévigny, J.; Robson, S.C.; et al. Extracellular nucleotide signaling in adult neural stem cells: Synergism with growth factor-mediated cellular proliferation. Development 2006, 133, 675–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, A.; Tang, H.; Cai, J.; Zhu, M.; Zhang, X.; Rao, M.; Mattson, M.P. Gap junctional communication is required to maintain mouse cortical neural progenitor cells in a proliferative state. Dev. Biol. 2004, 272, 203–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duval, N.; Gomès, D.; Calaora, V.; Calabrese, A.; Meda, P.; Bruzzone, R. Cell coupling and Cx43 expression in embryonic mouse neural progenitor cells. J. Cell Sci. 2002, 115, 3241–3251. [Google Scholar] [PubMed]
- Jäderstad, J.; Jäderstad, L.M.; Li, J.; Chintawar, S.; Salto, C.; Pandolfo, M.; Ourednik, V.; Teng, Y.D.; Sidman, R.L.; Arenas, E.; et al. Communication via gap junctions underlies early functional and beneficial interactions between grafted neural stem cells and the host. Proc. Natl. Acad. Sci. USA 2010, 107, 5184–5189. [Google Scholar] [CrossRef] [Green Version]
- Talaverón, R.; Matarredona, E.R.; de la Cruz, R.R.; Macías, D.; Gálvez, V.; Pastor, A.M. Implanted neural progenitor cells regulate glial reaction to brain injury and establish gap junctions with host glial cells. Glia 2014, 62, 623–638. [Google Scholar] [CrossRef]
- Ruiz de Almodovar, C.; Lambrechts, D.; Mazzone, M.; Carmeliet, P. Role and therapeutic potential of VEGF in the nervous system. Physiol. Rev. 2009, 89, 607–648. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrera, A.; Morcuende, S.; Talaverón, R.; Benítez-Temiño, B.; Pastor, A.M.; Matarredona, E.R. Purinergic Receptor Blockade with Suramin Increases Survival of Postnatal Neural Progenitor Cells In Vitro. Int. J. Mol. Sci. 2021, 22, 713. https://doi.org/10.3390/ijms22020713
Herrera A, Morcuende S, Talaverón R, Benítez-Temiño B, Pastor AM, Matarredona ER. Purinergic Receptor Blockade with Suramin Increases Survival of Postnatal Neural Progenitor Cells In Vitro. International Journal of Molecular Sciences. 2021; 22(2):713. https://doi.org/10.3390/ijms22020713
Chicago/Turabian StyleHerrera, Alejandro, Sara Morcuende, Rocío Talaverón, Beatriz Benítez-Temiño, Angel M. Pastor, and Esperanza R. Matarredona. 2021. "Purinergic Receptor Blockade with Suramin Increases Survival of Postnatal Neural Progenitor Cells In Vitro" International Journal of Molecular Sciences 22, no. 2: 713. https://doi.org/10.3390/ijms22020713
APA StyleHerrera, A., Morcuende, S., Talaverón, R., Benítez-Temiño, B., Pastor, A. M., & Matarredona, E. R. (2021). Purinergic Receptor Blockade with Suramin Increases Survival of Postnatal Neural Progenitor Cells In Vitro. International Journal of Molecular Sciences, 22(2), 713. https://doi.org/10.3390/ijms22020713