Purification and Biochemical Characterization of Sucrose synthase from the Stem of Nettle (Urtica dioica L.)




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
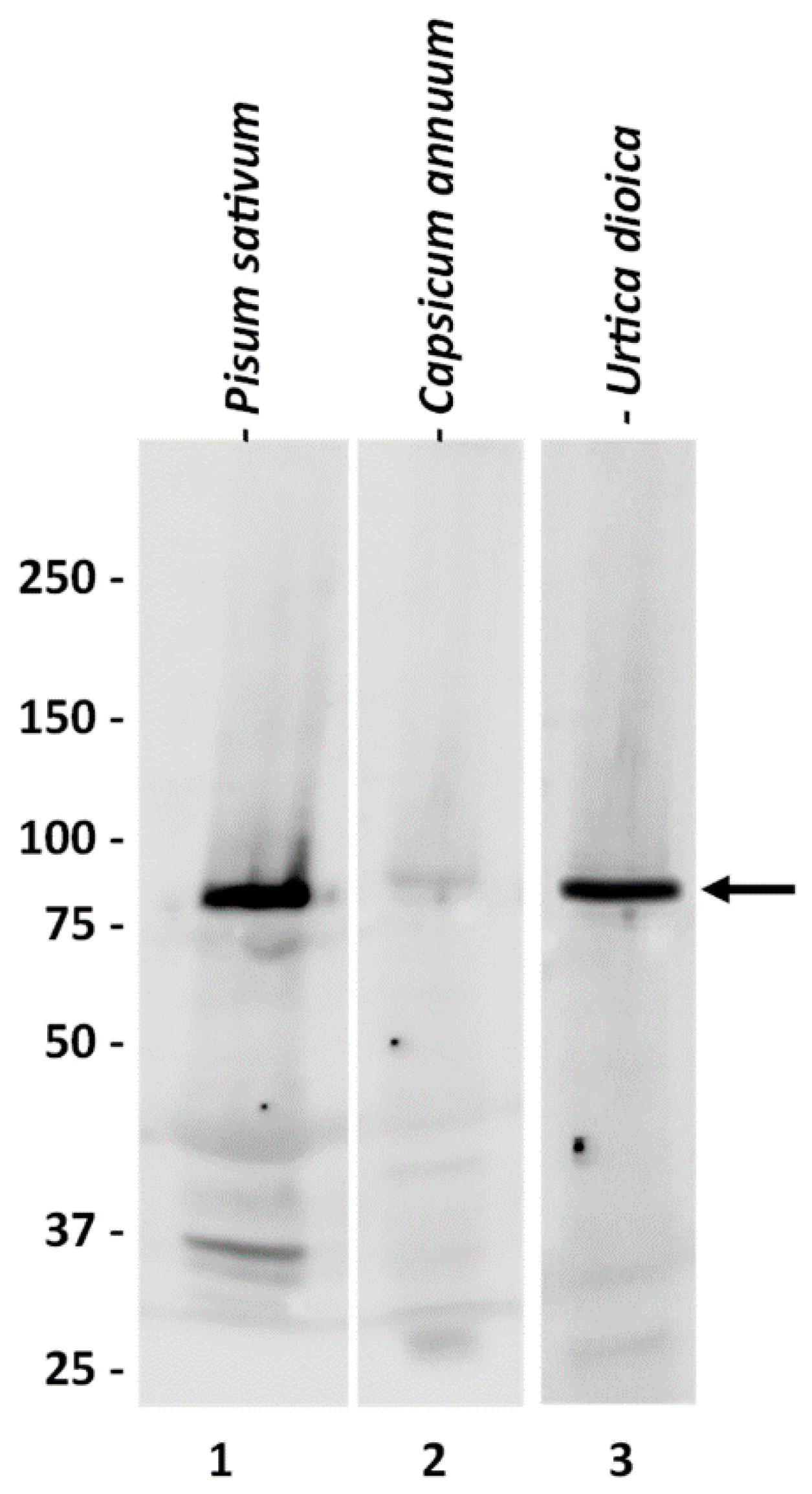
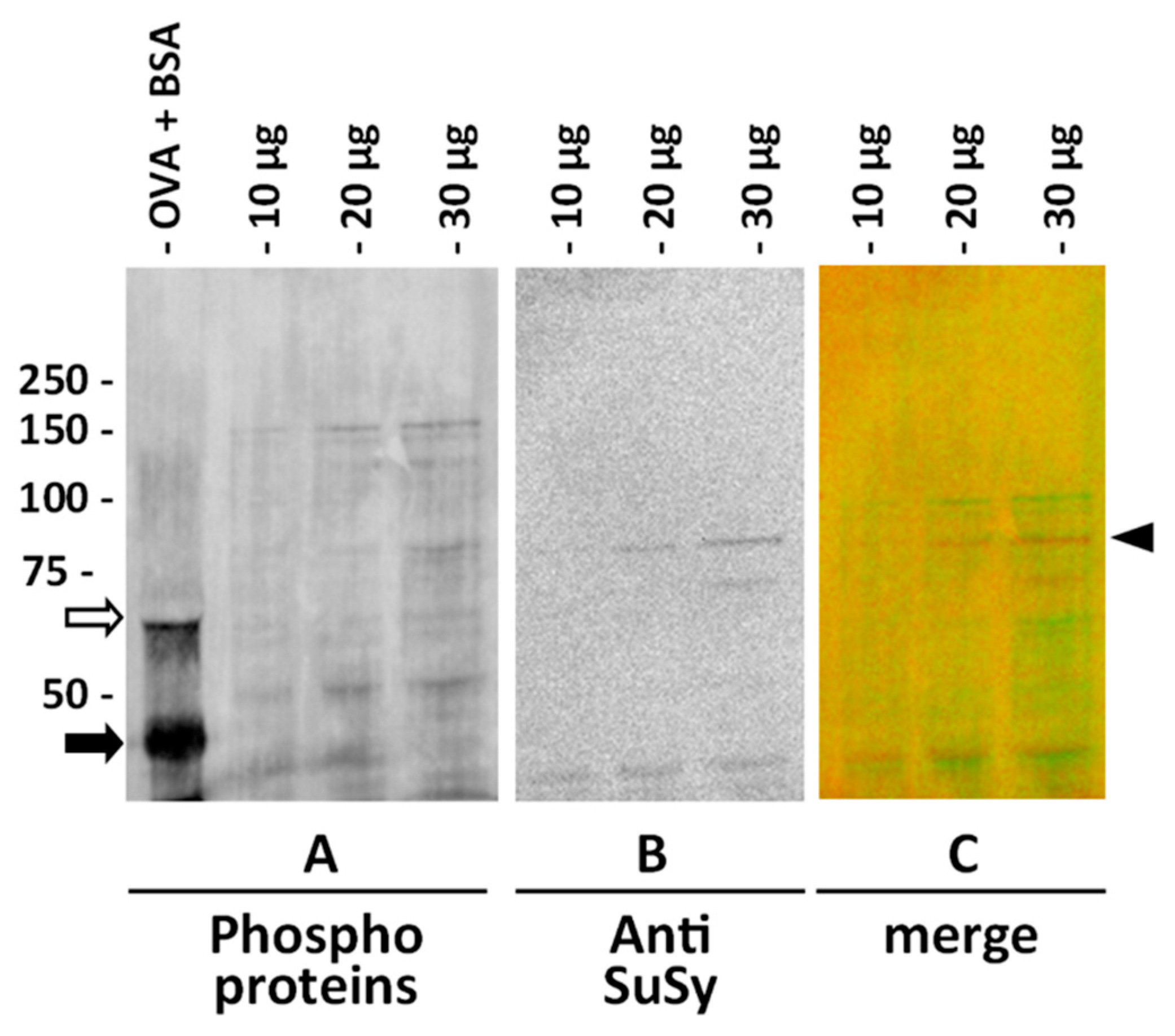
2.1. Characterization of the Anti-SuSy Antibody
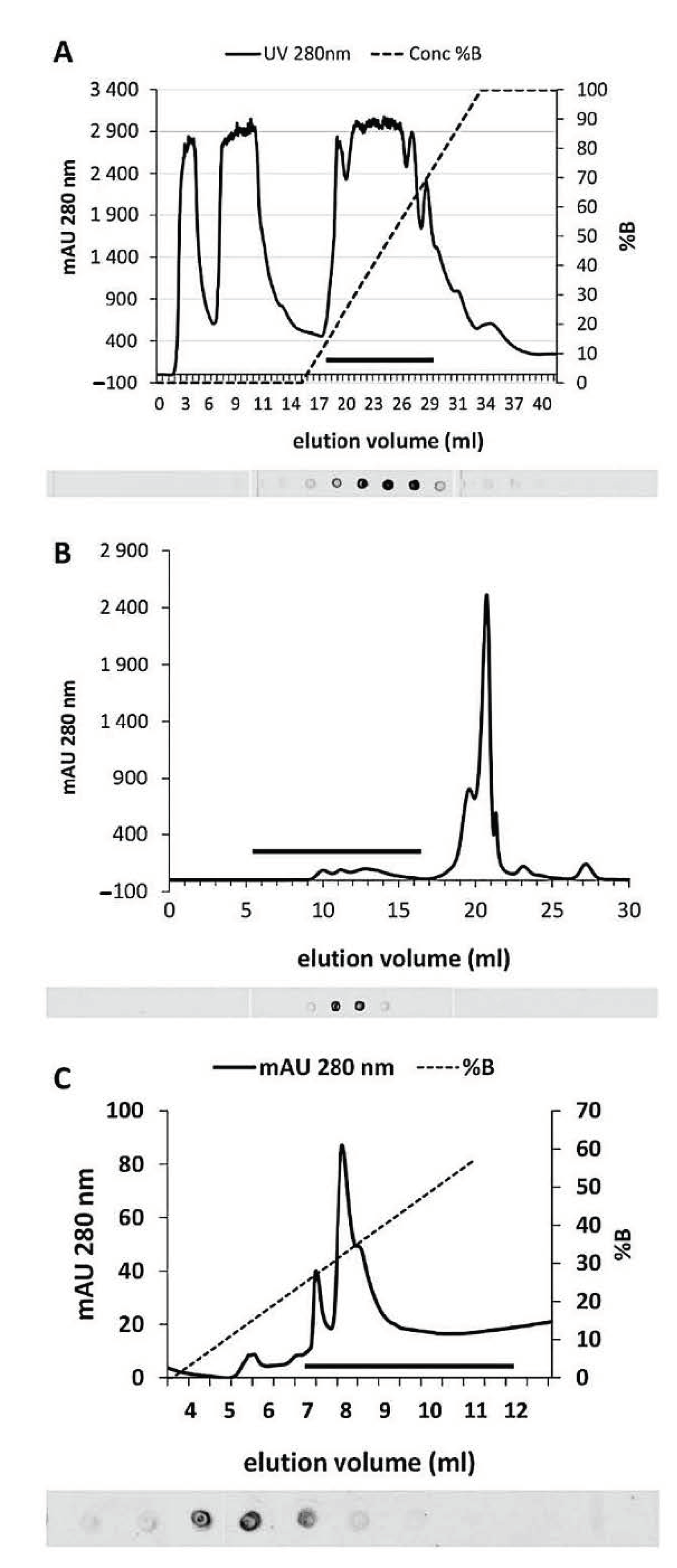
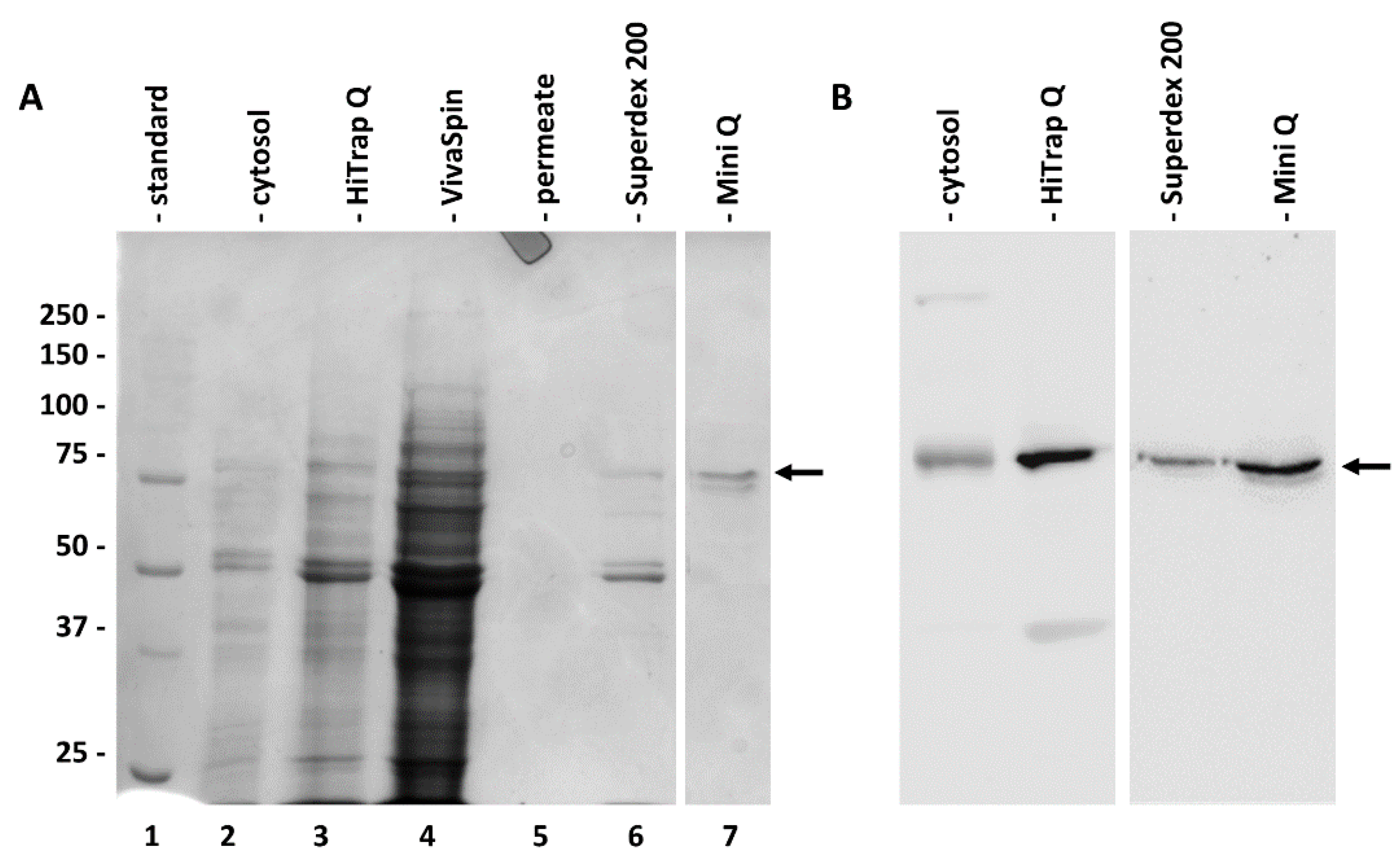
2.2. Purification of Cytosolic SuSy from Nettle Stems
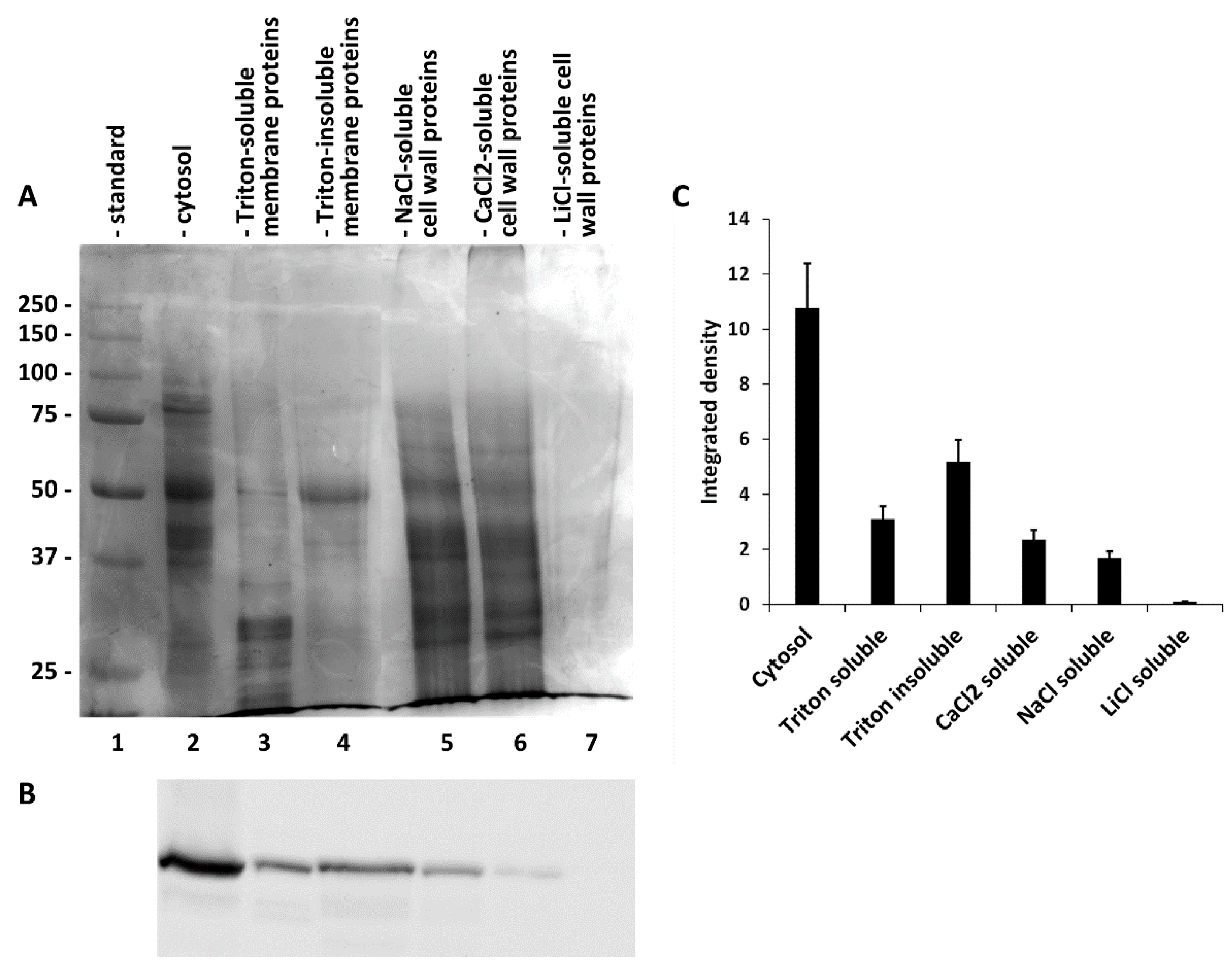
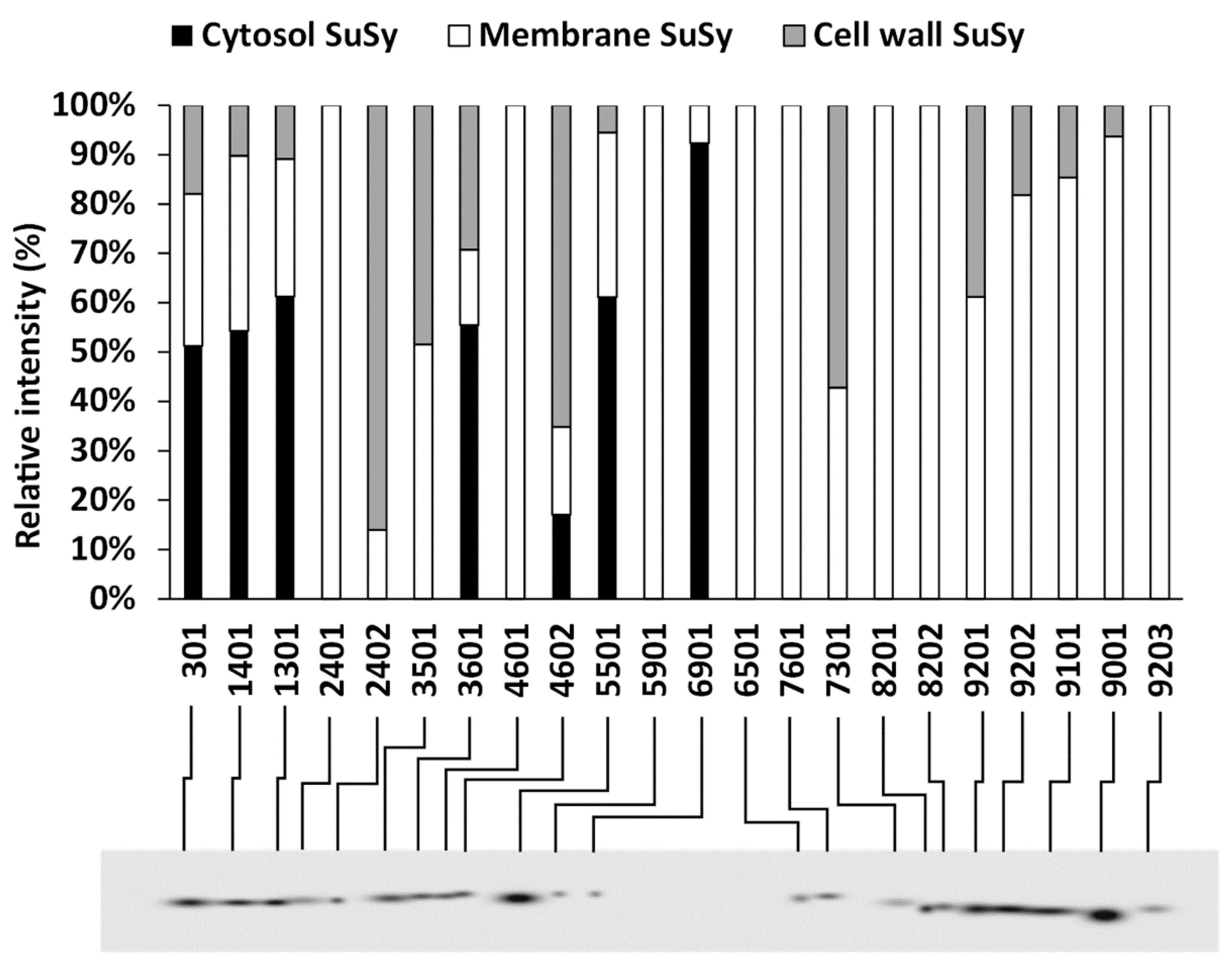
2.3. Sequential Extraction of SuSy from the Cytosol, Membranes and Cell Wall
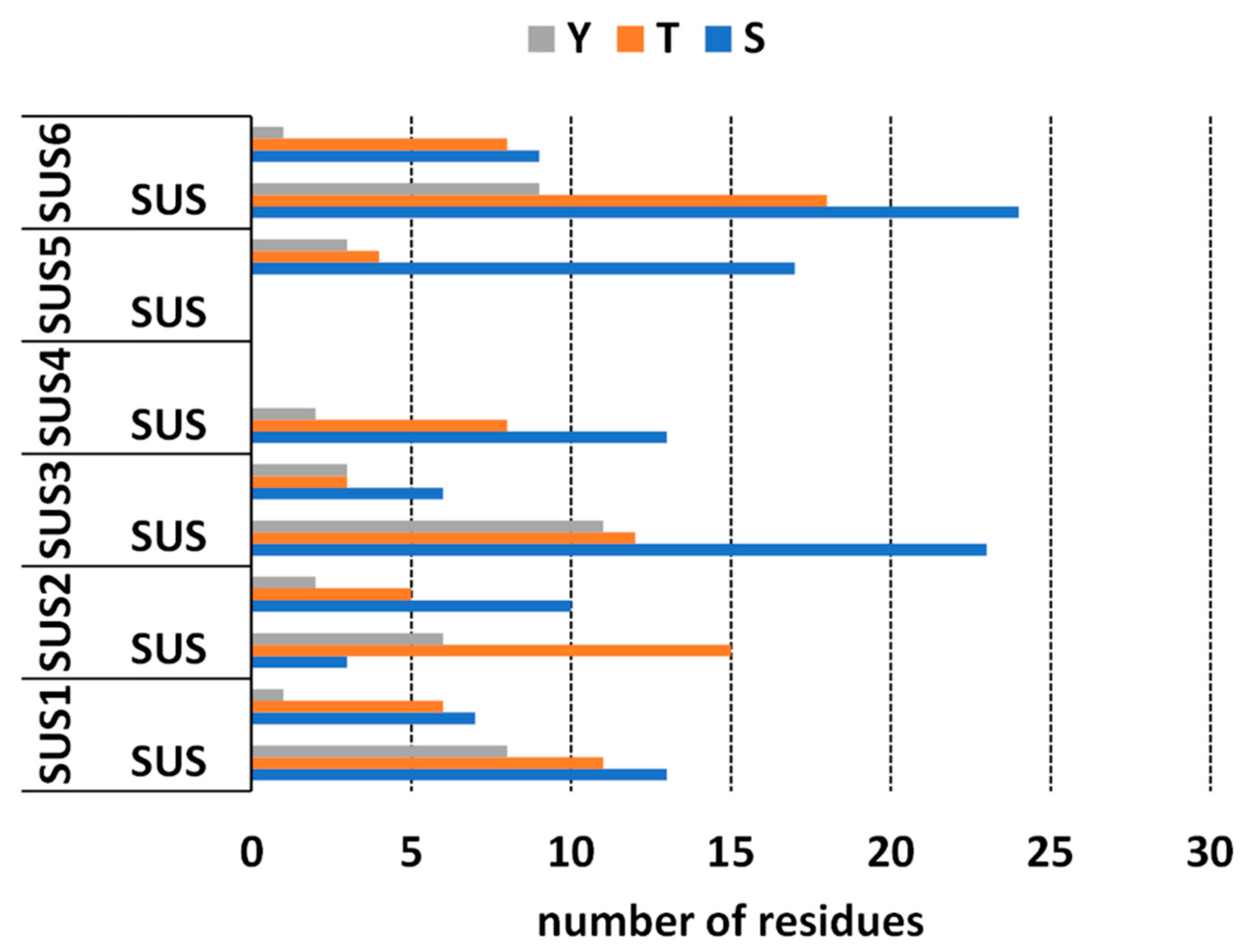
2.4. Bioinformatics Analysis of Putative Phosphorylation Sites of SuSy
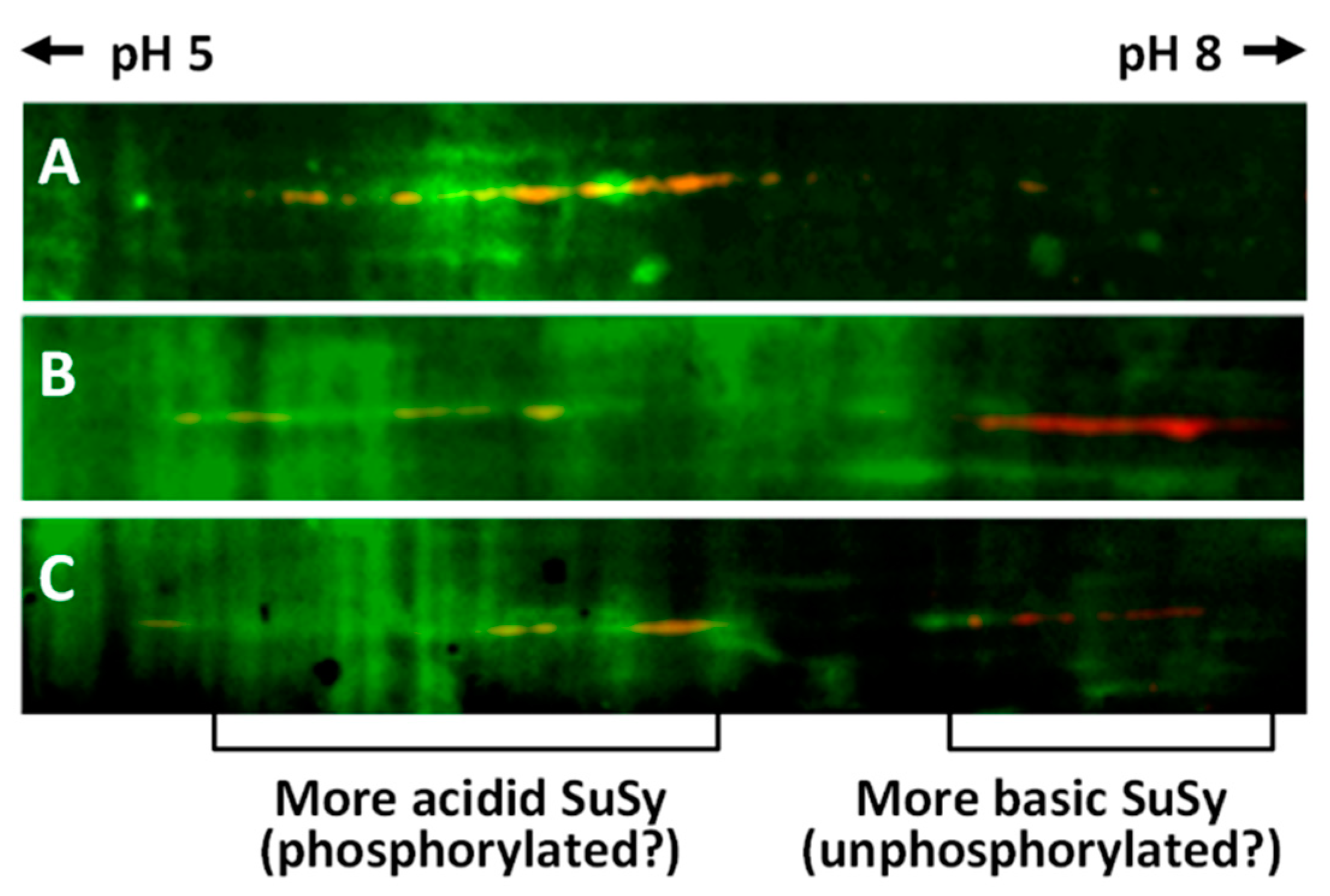
2.5. Analysis of SuSy Phosphorylation after Two-Dimensional Electrophoresis
2.6. Two-Dimensional Characterization of SuSy Isoforms
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. General Protein Extraction from Plant Species
4.3. Sequential Protein Extraction from Nettle Stem
4.4. Preparation of Soluble (S), Membrane (M) and Cell Wall (CW) Proteins
4.5. Purification of SuSy from Nettle Stem
4.6. Spot Test
4.7. 1-D and 2-D Electrophoresis and Immunoblotting
4.8. Assay of Phosphoproteins on Membranes
4.9. In Silico Prediction of Phosphorylation Sites
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Stein, O.; Granot, D. An overview of Sucrose synthases in plants. Front. Plant Sci. 2019, 10, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmölzer, K.; Gutmann, A.; Diricks, M.; Desmet, T.; Nidetzky, B. Sucrose synthase: A unique glycosyltransferase for biocatalytic glycosylation process development. Biotechnol. Adv. 2016, 34, 88–111. [Google Scholar] [CrossRef] [PubMed]
- Baud, S.; Vaultier, M.N.; Rochat, C. Structure and expression profile of the Sucrose synthase multigene family in Arabidopsis. J. Exp. Bot. 2004, 55, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.L.; Su, J.C. Quaternary structure of Sucrose synthetase from banana fruits. J. Chin. Biochem. Soc 1980, 9, 100–101. [Google Scholar]
- Sytykiewicz, H.; Czerniewicz, P.; Leszczyñski, B. Molecular characteristics of Sucrose synthase isolated from bird cherry leaves. Herba Pol. 2008, 54, 3. [Google Scholar]
- Verma, E.; Sharma, B.; Singal, H.R.; Munjal, R. Purification of Sucrose synthase from thermotolerant wheat grains and its characterization. J. Environ. Biol. 2018, 39, 459–466. [Google Scholar] [CrossRef]
- Fujii, S.; Hayashi, T.; Mizuno, K. Sucrose synthase is an integral component of the cellulose synthesis machinery. Plant Cell Physiol. 2010, 51, 294–301. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Anderson, S.; Zhang, Y.; Garavito, R.M. The structure of Sucrose synthase-1 from Arabidopsis thaliana and its functional implications. J. Biol. Chem. 2011, 286, 36108–36118. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, M.; Beevers, H. Subcellular distribution of gluconeogenetic enzymes in germinating castor bean endosperm. Plant Physiol. 1979, 64, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Amor, Y.; Haigler, C.H.; Johnson, S.; Wainscott, M.; Delmer, D.P. A membrane-associated form of Sucrose synthase and its potential role in synthesis of cellulose and callose in plants. Proc. Natl. Acad. Sci. USA 1995, 92, 9353–9357. [Google Scholar] [CrossRef] [Green Version]
- Persia, D.; Cai, G.; Del Casino, C.; Faleri, C.; Willemse, M.T.M.; Cresti, M. Sucrose synthase is associated with the cell wall of tobacco pollen tubes. Plant Physiol. 2008, 147, 1603–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, S.J.; Chourey, P.S. Evidence for plasma membrane-associated forms of Sucrose synthase in maize. Mol. Gen. Genet. 1996, 252, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Haigler, C.H.; Ivanova-Datcheva, M.; Hogan, P.S.; Salnikov, V.V.; Hwang, S.; Martin, K.; Delmer, D.P. Carbon partitioning to cellulose synthesis. Plant Mol. Biol. 2001, 47, 29–51. [Google Scholar] [CrossRef] [PubMed]
- Winter, H.; Huber, J.L.; Huber, S.C. Membrane association of Sucrose synthase: Changes during the graviresponse and possible control by protein phosphorylation. FEBS Lett. 1997, 420, 151–155. [Google Scholar] [CrossRef] [Green Version]
- Komina, O.; Zhou, Y.; Sarath, G.; Chollet, R. In vivo and in vitro phosphorylation of membrane and soluble forms of soybean nodule Sucrose synthase. Plant Physiol. 2002, 129, 1664–1673. [Google Scholar] [CrossRef] [Green Version]
- Konishi, T.; Ohmiya, Y.; Hayashi, T. Evidence that sucrose loaded into the phloem of a poplar leaf is used directly by Sucrose synthase associated with various beta-glucan synthases in the stem. Plant Physiol. 2004, 134, 1146–1152. [Google Scholar] [CrossRef] [Green Version]
- Brill, E.; van Thournout, M.; White, R.G.; Llewellyn, D.; Campbell, P.M.; Engelen, S.; Ruan, Y.L.; Arioli, T.; Furbank, R.T. A novel isoform of Sucrose synthase is targeted to the cell wall during secondary cell wall synthesis in cotton fiber. Plant Physiol. 2011, 157, 40–54. [Google Scholar] [CrossRef] [Green Version]
- Abdullah, M.; Cao, Y.; Cheng, X.; Meng, D.; Chen, Y.; Shakoor, A.; Gao, J.; Cai, Y. The Sucrose synthase gene family in Chinese pear (Pyrus bretschneideri Rehd.): Structure, expression, and evolution. Molecules 2018, 23, 1144. [Google Scholar] [CrossRef] [Green Version]
- Chourey, P.S.; Nelson, O.E. The enzymatic deficiency conditioned by the shrunken-1 mutations in maize. Biochem. Genet. 1976, 14, 1041–1055. [Google Scholar] [CrossRef]
- Tang, G.Q.; Sturm, A. Antisense repression of Sucrose synthase in carrot (Daucus carota L.) affects growth rather than sucrose partitioning. Plant Mol.Biol. 1999, 41, 465–479. [Google Scholar] [CrossRef]
- Zrenner, R.; Salanoubat, M.; Willmitzer, L.; Sonnewald, U. Evidence of the crucial role of Sucrose synthase for sink strength using transgenic potato plants (Solanum tuberosum L.). Plant J. 1995, 7, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Barratt, D.H.P.; Derbyshire, P.; Findlay, K.; Pike, M.; Wellner, N.; Lunn, J.; Feil, R.; Simpson, C.; Maule, A.J.; Smith, A.M. Normal growth of Arabidopsis requires cytosolic invertase but not Sucrose synthase. Proc. Natl. Acad. Sci. USA 2009, 106, 13124–13129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salnikov, V.V.; Grimson, M.J.; Seagull, R.W.; Haigler, C.H. Localization of Sucrose synthase and callose in freeze-substituted secondary-wall-stage cotton fibers. Protoplasma 2003, 221, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Delmer, D.P. Cellulose biosynthesis: Exciting times for a difficult field of study. Annu. Rev. Plant Physiol. 1999, 50, 245–276. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.L.; Llewellyn, D.J.; Furbank, R.T. Suppression of Sucrose synthase gene expression represses cotton fiber cell initiation, elongation, and seed development. Plant Cell 2003, 15, 952–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, Y. Rapid cell expansion and Cellulose synthesis regulated by plasmodesmata and sugar: Insights from the single-celled cotton fibre. Funct. Plant Biol. 2007, 34, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hong, Z.; Zhang, Z.; Olson, J.M.; Verma, D.P. A novel UDP-glucose transferase is part of the callose synthase complex and interacts with phragmoplastin at the forming cell plate. Plant Cell 2001, 13, 769–780. [Google Scholar]
- Debnath, S. Great potential of stinging nettle for sustainable textile and fashion. In Handbook of Sustainable Luxury Textiles and Fashion. Environmental Footprints and Eco-Design of Products and Processes; Gardetti, M., Muthu, S., Eds.; Springer: Singapore, 2015; pp. 43–57. ISBN 9789812876331. [Google Scholar]
- Guerriero, G.; Behr, M.; Backes, A.; Faleri, C.; Hausman, J.F.; Lutts, S.; Cai, G. Bast fibre formation: Insights from Next-Generation Sequencing. Procedia Eng. 2017, 200, 229–235. [Google Scholar] [CrossRef]
- Guerriero, G.; Sergeant, K.; Hausman, J.F. Integrated-omics: A powerful approach to understanding the heterogeneous lignification of fibre crops. Int. J. Mol. Sci. 2013, 14, 10958–10978. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Backes, A.; Legay, S.; Berni, R.; Faleri, C.; Gatti, E.; Hausman, J.F.; Cai, G.; Guerriero, G. Cell wall composition and transcriptomics in stem tissues of stinging nettle (Urtica dioica L.): Spotlight on a neglected fibre crop. Plant Direct 2019, 3, e00151. [Google Scholar] [CrossRef] [Green Version]
- Backes, A.; Behr, M.; Xu, X.; Gatti, E.; Legay, S.; Predieri, S.; Hausman, J.F.; Deyholos, M.K.; Cai, G.; Guerriero, G. Sucrose synthase gene expression analysis in the fibre nettle (Urtica dioica L.) cultivar “clone 13”. Ind. Crops Prod. 2018, 123, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Blom, N.; Gammeltoft, S.; Brunak, S. Sequence and structure-based prediction of eukaryotic protein phosphorylation sites. J. Mol. Biol. 1999, 294, 1351–1362. [Google Scholar] [CrossRef] [PubMed]
- Blom, N.; Sicheritz-Pontén, T.; Gupta, R.; Gammeltoft, S.; Brunak, S. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence. Proteomics 2004, 4, 1633–1649. [Google Scholar] [CrossRef] [PubMed]
- Zuk, M.; Richter, D.; Matuła, J.; Szopa, J. Linseed, the multipurpose plant. Ind. Crops Prod. 2015, 75, 165–177. [Google Scholar] [CrossRef]
- Andre, C.M.; Hausman, J.-F.; Guerriero, G. Cannabis sativa: The plant of the thousand and one molecules. Front. Plant Sci. 2016, 7, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jan, K.N.; Zarafshan, K.; Singh, S. Stinging nettle (Urtica dioica L.): A reservoir of nutrition and bioactive components with great functional potential. J. Food Meas. Charact. 2017, 11, 423–433. [Google Scholar] [CrossRef]
- Guerriero, G.; Berni, R.; Muñoz-Sanchez, J.A.; Apone, F.; Abdel-Salam, E.M.; Qahtan, A.A.; Alatar, A.A.; Cantini, C.; Cai, G.; Hausman, J.F.; et al. Production of plant secondary metabolites: Examples, tips and suggestions for biotechnologists. Genes 2018, 9, 309. [Google Scholar] [CrossRef] [Green Version]
- Schöttner, M.; Ganßer, D.; Spiteller, G. Lignans from the roots of Urtica dioica and their metabolites bind to human sex hormone binding globulin (SHBG). Planta Med. 1997, 63, 529–532. [Google Scholar] [CrossRef]
- Xu, X.; Guignard, C.; Renaut, J.; Hausman, J.-F.; Gatti, E.; Predieri, S.; Guerriero, G. Insights into lignan composition and biosynthesis in stinging nettle (Urtica dioica L.). Molecules 2019, 24, 3863. [Google Scholar] [CrossRef] [Green Version]
- Ullah, R.; Hussain, I.; Ahmad, S. Diocanol; one new phenol derivative isolated and characterized from Urtica dioica. Arab. J. Chem. 2017, 10, S1284–S1286. [Google Scholar] [CrossRef] [Green Version]
- Agus Suryawan, I.G.P.; Suardana, N.P.G.; Suprapta Winaya, I.N.; Budiarsa Suyasa, I.W.; Tirta Nindhia, T.G. Study of stinging nettle (Urtica dioica L.) fibers reinforced green composite materials: A review. IOP Conf. Ser. Mater. Sci. Eng. 2017, 201, 012001. [Google Scholar] [CrossRef] [Green Version]
- Duncan, K.A.; Hardin, S.C.; Huber, S.C. The three maize Sucrose synthase isoforms differ in distribution, localization, and phosphorylation. Plant Cell Physiol. 2006, 47, 959–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouhier, H.; Usuda, H. Spatial and temporal distribution of Sucrose synthase in the radish hypocotyl in relation to thickening growth. Plant Cell Physiol. 2001, 42, 583–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleczkowski, L.A.; Kunz, S.; Wilczynska, M. Mechanisms of UDP-glucose synthesis in plants. Crit. Rev. Plant Sci. 2010, 29, 191–203. [Google Scholar] [CrossRef]
- Nilsson, R.; Bernfur, K.; Gustavsson, N.; Bygdell, J.; Wingsle, G.; Larsson, C. Proteomics of plasma membranes from poplar trees reveals tissue distribution of transporters, receptors, and proteins in cell wall formation. Mol. Cell. Proteom. 2010, 9, 368. [Google Scholar] [CrossRef] [Green Version]
- Pozueta-Romero, J.; Pozueta-Romero, D.; Gonzalez, P.; Etxeberria, E. Activity of membrane-associated Sucrose synthase is regulated by its phosphorylation status in cultured cells of sycamore (Acer pseudoplatanus). Physiol. Plant. 2004, 122, 275–280. [Google Scholar] [CrossRef]
- Wei, Z.; Qu, Z.; Zhang, L.; Zhao, S.; Bi, Z.; Ji, X.; Wang, X.; Wei, H. Overexpression of poplar xylem Sucrose synthase in tobacco leads to a thickened cell wall and increased height. PLoS ONE 2015, 10, e0120669. [Google Scholar] [CrossRef] [Green Version]
- Rende, U.; Wang, W.; Gandla, M.L.; Jonsson, L.J.; Niittyla, T. Cytosolic invertase contributes to the supply of substrate for cellulose biosynthesis in developing wood. New Phytol. 2017, 214, 796–807. [Google Scholar] [CrossRef]
- Slewinski, T.L. Diverse functional roles of monosaccharide transporters and their homologs in vascular plants: A physiological perspective. Mol. Plant 2011, 4, 641–662. [Google Scholar] [CrossRef]
- Bessueille, L.; Sindt, N.; Guichardant, M.; Djerbi, S.; Teeri, T.T.; Bulone, V. Plasma membrane microdomains from hybrid aspen cells are involved in cell wall polysaccharide Biosynthesis. Biochem. J. 2009, 420, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Briolay, A.; Bouzenzana, J.; Guichardant, M.; Deshayes, C.; Sindt, N.; Bessueille, L.; Bulone, V. Cell wall polysaccharide synthases are located in detergent-resistant membrane microdomains in Oomycetes. Appl. Environ. Microbiol. 2009, 75, 1938–1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrotta, L.; Faleri, C.; Cresti, M.; Cai, G. Heat stress affects the cytoskeleton and the delivery of Sucrose synthase in tobacco pollen tubes. Planta 2016, 243, 43–63. [Google Scholar] [CrossRef] [PubMed]
- Schafer, W.E.; Rohwer, J.M.; Botha, F.C. Partial purification and characterisation of Sucrose synthase in sugarcane. J. Plant Physiol. 2005, 162, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.Y.; Wang, A.Y. Purification and characterization of Sucrose synthase isozymes from etiolated rice seedlings. Biochem 1998, 46, 107–113. [Google Scholar] [CrossRef]
- Sebkova, V.; Unger, C.; Hardegger, M.; Sturm, A. Biochemical, physiological, and molecular characterization of Sucrose synthase from Daucus carota. Plant Physiol. 1995, 108, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Schulze, W.X. Proteomics approaches to understand protein phosphorylation in pathway modulation. Curr. Opin. Plant Biol. 2010, 13, 279–286. [Google Scholar] [CrossRef]
- van Bentem, S.D.L.F.; Anrather, D.; Roitinger, E.; Djamei, A.; Hufnagl, T.; Barta, A.; Csaszar, E.; Dohnal, I.; Lecourieux, D.; Hirt, H. Phosphoproteomics reveals extensive in vivo phosphorylation of Arabidopsis proteins involved in RNA metabolism. Nucleic Acids Res. 2006, 34, 3267–3278. [Google Scholar] [CrossRef]
- Nühse, T.S.; Bottrill, A.R.; Jones, A.M.E.; Peck, S.C. Quantitative phosphoproteomic analysis of plasma membrane proteins reveals regulatory mechanisms of plant innate immune responses. Plant J. 2007, 51, 931–940. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhou, H.; Berke, L.; Heck, A.J.R.; Mohammed, S.; Scheres, B.; Menke, F.L.H. Quantitative phosphoproteomics after auxin-stimulated lateral root induction identifies an SNX1 protein phosphorylation site required for growth. Mol. Cell. Proteomics 2013, 12, 1158–1169. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Sklodowski, K.; Encke, B.; Schulze, W.X. A kinase-phosphatase signaling module with BSK8 and BSL2 involved in regulation of Sucrose-phosphate synthase. J. Proteome Res. 2014, 13, 3397–3409. [Google Scholar] [CrossRef]
- Hardin, S.C.; Winter, H.; Huber, S.C. Phosphorylation of the amino terminus of maize Sucrose synthase in relation to membrane association and enzyme activity. Plant Physiol. 2004, 134, 1427–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, K. Sucrose metabolism: Regulatory mechanisms and pivotal roles in sugar sensing and plant development. Curr. Opin. Plant Biol. 2004, 7, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Hardin, S.C.; Tang, G.Q.; Scholz, A.; Holtgraewe, D.; Winter, H.; Huber, S.C. Phosphorylation of Sucrose synthase at serine 170: Occurrence and possible role as a signal for proteolysis. Plant J. 2003, 35, 588–603. [Google Scholar] [CrossRef]
- Nakai, T.; Konishi, T.; Zhang, X.Q.; Chollet, R.; Tonouchi, N.; Tsuchida, T.; Yoshinaga, F.; Mori, H.; Sakai, F.; Hayashi, T. An increase in apparent affinity for sucrose of mung bean Sucrose synthase is caused by in vitro phosphorylation or directed mutagenesis of Ser11. Plant Cell Physiol. 1998, 39, 1337–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, K.; Albrecht, S.; Schaller, A. Targeted analysis of protein phosphorylation by 2D electrophoresis. In Methods in Molecular Biology; Humana Press: New York, NY, USA, 2015; pp. 167–176. [Google Scholar]
- Duncan, K.A.; Huber, S.C. Sucrose synthase oligomerization and F-actin association are regulated by sucrose concentration and phosphorylation. Plant Cell Physiol. 2007, 48, 1612–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anguenot, R.; Nguyen-Quoc, B.; Yelle, S.; Michaud, D. Protein phosphorylation and membrane association of Sucrose synthase in developing tomato fruit. Plant Physiol. Biochem. 2006, 44, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Subbaiah, C.C.; Sachs, M.M. Altered patterns of Sucrose synthase phosphorylation and localization precede callose induction and root tip death in anoxic maize seedlings. Plant Physiol. 2001, 125, 585–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, H.; Niikura, M.; Narumi, A.; Aoki, H.; Sasaki, T.; Shimada, H. Phosphorylation of rice Sucrose synthase isoforms promotes the activity of sucrose degradation. Plant Biotechnol. 2017, 34, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Tanase, K.; Shiratake, K.; Mori, H.; Yamaki, S. Changes in the phosphorylation state of Sucrose synthase during development of Japanese pear fruit. Physiol. Plant 2002, 114, 21–26. [Google Scholar] [CrossRef]
- Gorshkova, T.A.; Sal’nikov, V.V.; Chemikosova, S.B.; Ageeva, M.V.; Pavlencheva, N.V.; Van Dam, J.E.G. The snap point: A transition point in Linum usitatissimum bast fiber development. Ind. Crop. Prod. 2003, 18, 213–221. [Google Scholar] [CrossRef]
- Chabi, M.; Goulas, E.; Leclercq, C.C.; De Waele, I.; Rihouey, C.; Cenci, U.; Day, A.; Blervacq, A.-S.; Neutelings, G.; Duponchel, L. A cell wall proteome and targeted cell wall analyses provide novel information on hemicellulose metabolism in flax. Mol. Cell. Proteomics 2017, 16, 1634–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulas, E.; Le Dily, F.; Teissedre, L.; Corbel, G.; Robin, C.; Ourry, A. Vegetative storage proteins in white clover (Trifolium repens L.): Quantitative and qualitative features. Ann. Bot. 2001, 88, 789–795. [Google Scholar]
- Feiz, L.; Irshad, M.; Pont-Lezica, R.F.; Canut, H.; Jamet, E. Evaluation of cell wall preparations for proteomics: A new procedure for purifying cell walls from Arabidopsis hypocotyls. Plant Methods 2006, 2, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mareri, L.; Guerriero, G.; Hausman, J.-F.; Cai, G. Purification and Biochemical Characterization of Sucrose synthase from the Stem of Nettle (Urtica dioica L.). Int. J. Mol. Sci. 2021, 22, 851. https://doi.org/10.3390/ijms22020851
Mareri L, Guerriero G, Hausman J-F, Cai G. Purification and Biochemical Characterization of Sucrose synthase from the Stem of Nettle (Urtica dioica L.). International Journal of Molecular Sciences. 2021; 22(2):851. https://doi.org/10.3390/ijms22020851
Chicago/Turabian StyleMareri, Lavinia, Gea Guerriero, Jean-Francois Hausman, and Giampiero Cai. 2021. "Purification and Biochemical Characterization of Sucrose synthase from the Stem of Nettle (Urtica dioica L.)" International Journal of Molecular Sciences 22, no. 2: 851. https://doi.org/10.3390/ijms22020851
APA StyleMareri, L., Guerriero, G., Hausman, J. -F., & Cai, G. (2021). Purification and Biochemical Characterization of Sucrose synthase from the Stem of Nettle (Urtica dioica L.). International Journal of Molecular Sciences, 22(2), 851. https://doi.org/10.3390/ijms22020851