Immunological Dysfunction in Tourette Syndrome and Related Disorders

Abstract
:1. Introduction
2. Association between Infection and TS and Related Disorders
2.1. GAS Infection
2.1.1. Sydenham’s Chorea (SC) and Pediatric Autoimmune Neuropsychiatric Disorders Associated with Streptococcal Infections (PANDAS)
2.1.2. GAS and TS
2.2. Other Pathogens
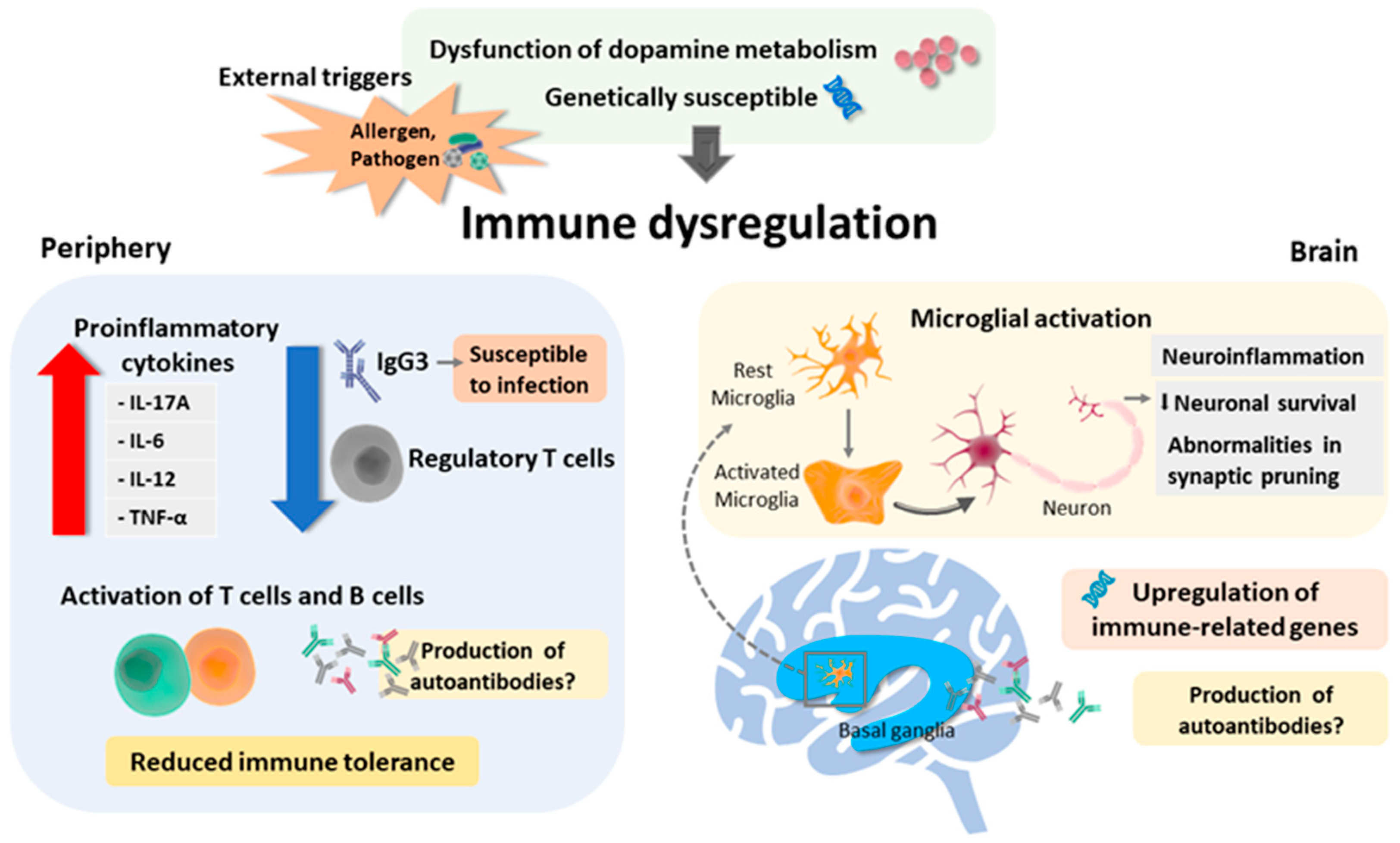
3. Immune Dysregulation in TS
3.1. Genetic Expression in TS
3.2. Aberrant Peripheral Immune Activities
3.2.1. Alternation in Immune Cell Subset and Immunophenotyping
3.2.2. Dysregulation of Effector Molecules and Immunoglobulin (Ig)
3.3. Aberrant Neuronal-Immune Activities
3.3.1. Alteration in Microglia
3.3.2. Antineuronal Antibodies
4. Animal Models for Immune Dysregulation in TS
4.1. Injection of Cytokines or Other Immune Mediators
4.2. Immunization with Specific Microorganism
4.3. Passive Sera Transfusion
4.4. Transgenic Animal Models
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de La Tourette, G. Étude sur une affection nerveuse caractérisée par l’incoordination motrice accompagnée d’écholalie et de copralalie. Arch. Neurol. 1885, 9, 19–42, 158–200. [Google Scholar]
- Dale, R.C. Tics and Tourette: A clinical, pathophysiological and etiological review. Curr. Opin. Pediatr. 2017, 29, 665–673. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5®); American Psychiatric Pub: Washington, DC, USA, 2013. [Google Scholar]
- Scharf, J.M.; Miller, L.L.; Gauvin, C.A.; Alabiso, J.; Mathews, C.A.; Ben-Shlomo, Y. Population prevalence of Tourette syndrome: A systematic review and meta-analysis. Mov. Disord. 2015, 30, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Greene, D.J.; Schlaggar, B.L.; Black, K.J. Neuroimaging in Tourette Syndrome: Research Highlights from 2014–2015. Curr. Dev. Disord. Rep. 2015, 2, 300–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganos, C.; Martino, D. Tics and tourette syndrome. Neurol. Clin. 2015, 33, 115–136. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Trescher, W.; Byler, D. Tourette syndrome and comorbid neuropsychiatric conditions. Curr. Dev. Disord. Rep. 2016, 3, 217–221. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.-T.; Huang, H.-L.; Wong, L.C.; Weng, W.-C.; Vasylenko, T.; Jong, Y.-J.; Lin, W.-S.; Ho, S.-Y. Tourette Syndrome as an Independent Risk Factor for Subsequent Sleep Disorders in Children: A Nationwide Population-Based Case–Control Study. Sleep 2017, 40, zsw072. [Google Scholar] [CrossRef]
- Roessner, V.; Schoenefeld, K.; Buse, J.; Bender, S.; Ehrlich, S.; Münchau, A. Pharmacological treatment of tic disorders and Tourette syndrome. Neuropharmacology 2013, 68, 143–149. [Google Scholar] [CrossRef]
- Müller-Vahl, K.R.; Grosskreutz, J.; Prell, T.; Kaufmann, J.; Bodammer, N.; Peschel, T. Tics are caused by alterations in prefrontal areas, thalamus and putamen, while changes in the cingulate gyrus reflect secondary compensatory mechanisms. BMC Neurosci. 2014, 15, 6. [Google Scholar] [CrossRef] [Green Version]
- Worbe, Y.; Marrakchi-Kacem, L.; Lecomte, S.; Valabregue, R.; Poupon, F.; Guevara, P.; Tucholka, A.; Mangin, J.F.; Vidailhet, M.; Lehericy, S.; et al. Altered structural connectivity of cortico-striato-pallido-thalamic networks in Gilles de la Tourette syndrome. Brain 2015, 138, 472–482. [Google Scholar] [CrossRef] [Green Version]
- Zapparoli, L.; Porta, M.; Paulesu, E. The anarchic brain in action: The contribution of task-based fMRI studies to the understanding of Gilles de la Tourette syndrome. Curr. Opin. Neurol. 2015, 28, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-J.; Wong, L.C.; Wang, H.-P.; Lee, W.-T. The multimodality neuroimage findings in individuals with Tourette syndrome. Pediatrics Neonatol. 2020, 61, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Singer, H.S.; Szymanski, S.; Giuliano, J.; Yokoi, F.; Dogan, A.S.; Brasic, J.R.; Zhou, Y.; Grace, A.A.; Wong, D.F. Elevated intrasynaptic dopamine release in Tourette’s syndrome measured by PET. Am. J. Psychiatry 2002, 159, 1329–1336. [Google Scholar] [CrossRef] [PubMed]
- Buse, J.; Schoenefeld, K.; Munchau, A.; Roessner, V. Neuromodulation in Tourette syndrome: Dopamine and beyond. Neurosci Biobehav. Rev. 2013, 37, 1069–1084. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.F.; Singer, H.S.; Brandt, J.; Shaya, E.; Chen, C.; Brown, J.; Kimball, A.; Gjedde, A.; Dannals, R.F.; Ravert, H.T. D2-like dopamine receptor density in Tourette syndrome measured by PET. J. Nucl. Med. 1997, 38, 1243–1247. [Google Scholar]
- Wong, D.F.; Brašić, J.R.; Singer, H.S.; Schretlen, D.J.; Kuwabara, H.; Zhou, Y.; Nandi, A.; Maris, M.A.; Alexander, M.; Ye, W. Mechanisms of dopaminergic and serotonergic neurotransmission in Tourette syndrome: Clues from an in vivo neurochemistry study with PET. Neuropsychopharmacology 2008, 33, 1239. [Google Scholar] [CrossRef] [PubMed]
- Landau, Y.E.; Steinberg, T.; Richmand, B.; Leckman, J.F.; Apter, A. Involvement of immunologic and biochemical mechanisms in the pathogenesis of Tourette’s syndrome. J. Neural. Transm. 2012, 119, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Martino, D.; Dale, R.C.; Gilbert, D.L.; Giovannoni, G.; Leckman, J.F. Immunopathogenic mechanisms in tourette syndrome: A critical review. Mov. Disord. 2009, 24, 1267–1279. [Google Scholar] [CrossRef] [Green Version]
- Kiessling, L.S.; Marcotte, A.C.; Culpepper, L. Antineuronal antibodies in movement disorders. Pediatrics 1993, 92, 39–43. [Google Scholar]
- Martino, D.; Chiarotti, F.; Buttiglione, M.; Cardona, F.; Creti, R.; Nardocci, N.; Orefici, G.; Veneselli, E.; Rizzo, R.; Italian Tourette Syndrome Study Group. The relationship between group A streptococcal infections and Tourette syndrome: A study on a large service-based cohort. Dev. Med. Child Neurol. 2011, 53, 951–957. [Google Scholar] [CrossRef]
- Swedo, S.E.; Leonard, H.L.; Garvey, M.; Mittleman, B.; Allen, A.J.; Perlmutter, S.; Lougee, L.; Dow, S.; Zamkoff, J.; Dubbert, B.K. Pediatric autoimmune neuropsychiatric disorders associated with streptococcal infections: Clinical description of the first 50 cases. Am. J. Psychiatry 1998, 155, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Swedo, S. From Research Subgroup to Clinical Syndrome: Modifying the PANDAS Criteria to Describe PANS (Pediatric Acute-onset Neuropsychiatric Syndrome). Pediatrics Ther. 2012, 2, 113. [Google Scholar] [CrossRef]
- Leckman, J.F.; Katsovich, L.; Kawikova, I.; Lin, H.; Zhang, H.; Krönig, H.; Morshed, S.; Parveen, S.; Grantz, H.; Lombroso, P.J. Increased serum levels of interleukin-12 and tumor necrosis factor-alpha in Tourette’s syndrome. Biol. Psychiatry 2005, 57, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, C.; Wurster, U.; Müller-Vahl, K.R. Oligoclonal bands in cerebrospinal fluid in patients with Tourette’s syndrome. Mov. Disord. 2011, 26, 343–346. [Google Scholar] [CrossRef]
- Pichichero, M.E. Group A streptococcal tonsillopharyngitis: Cost-effective diagnosis and treatment. Ann. Emerg. Med. 1995, 25, 390–403. [Google Scholar] [CrossRef]
- Shaikh, N.; Leonard, E.; Martin, J.M. Prevalence of streptococcal pharyngitis and streptococcal carriage in children: A meta-analysis. Pediatrics 2010, 126, e557–e564. [Google Scholar] [CrossRef]
- Swedo, S.E.; Rapoport, J.L.; Cheslow, D.L.; Leonard, H.L.; Ayoub, E.M.; Hosier, D.M.; Wald, E.R. High prevalence of obsessive-compulsive symptoms in patients with Sydenham’s chorea. Am. J. Psychiatry 1989, 146, 246–249. [Google Scholar] [CrossRef]
- Orlovska, S.; Vestergaard, C.H.; Bech, B.H.; Nordentoft, M.; Vestergaard, M.; Benros, M.E. Association of Streptococcal Throat Infection With Mental Disorders: Testing Key Aspects of the PANDAS Hypothesis in a Nationwide Study. JAMA Psychiatry 2017, 74, 740–746. [Google Scholar] [CrossRef]
- Kurlan, R.; Johnson, D.; Kaplan, E.L. Streptococcal infection and exacerbations of childhood tics and obsessive-compulsive symptoms: A prospective blinded cohort study. Pediatrics 2008, 121, 1188–1197. [Google Scholar] [CrossRef]
- Leckman, J.F.; King, R.A.; Gilbert, D.L.; Coffey, B.J.; Singer, H.S.; Dure, L.S.t.; Grantz, H.; Katsovich, L.; Lin, H.; Lombroso, P.J.; et al. Streptococcal upper respiratory tract infections and exacerbations of tic and obsessive-compulsive symptoms: A prospective longitudinal study. J. Am. Acad. Child Adolesc. Psychiatry 2011, 50, 108–118.e103. [Google Scholar] [CrossRef] [Green Version]
- Cox, C.J.; Zuccolo, A.J.; Edwards, E.V.; Mascaro-Blanco, A.; Alvarez, K.; Stoner, J.; Chang, K.; Cunningham, M.W. Antineuronal antibodies in a heterogeneous group of youth and young adults with tics and obsessive-compulsive disorder. J. Child Adolesc. Psychopharmacol. 2015, 25, 76–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, T.K.; Sajid, M.; Soto, O.; Shapira, N.; Edge, P.; Yang, M.; Lewis, M.H.; Goodman, W.K. Detecting pediatric autoimmune neuropsychiatric disorders associated with streptococcus in children with obsessive-compulsive disorder and tics. Biol. Psychiatry 2004, 55, 61–68. [Google Scholar] [CrossRef]
- Mell, L.K.; Davis, R.L.; Owens, D. Association between streptococcal infection and obsessive-compulsive disorder, Tourette’s syndrome, and tic disorder. Pediatrics 2005, 116, 56–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.C.; Lau, C.I.; Lin, C.C.; Chang, A.; Kao, C.H. Group A Streptococcal Infections Are Associated With Increased Risk of Pediatric Neuropsychiatric Disorders: A Taiwanese Population-Based Cohort Study. J. Clin. Psychiatry 2016, 77, e848–e854. [Google Scholar] [CrossRef]
- Singer, H.S.; Gause, C.; Morris, C.; Lopez, P. Serial immune markers do not correlate with clinical exacerbations in pediatric autoimmune neuropsychiatric disorders associated with streptococcal infections. Pediatrics 2008, 121, 1198–1205. [Google Scholar] [CrossRef] [PubMed]
- Mûœler, N.; Riedel, M.; Fõderreuther, S.; Blendinger, C.; Abele-Horn, M. Tourette’s Syndrome and Mycoplasma Pneumoniae Infection. Am. J. Psychiatry 2000, 157, 481-a. [Google Scholar] [CrossRef] [Green Version]
- Dehning, S.; Matz, J.; Riedel, M.; Kerle, I.A.; Müller, N. Symptom exacerbation in tourette syndrome due to bacterial reinfection. J. Clin. Psychiatry 2009, 70, 1606. [Google Scholar] [CrossRef]
- Müller, N.; Riedel, M.; Blendinger, C.; Oberle, K.; Jacobs, E.; Abele-Horn, M. Mycoplasma pneumoniae infection and Tourette’s syndrome. Psychiatry Res. 2004, 129, 119–125. [Google Scholar] [CrossRef]
- Tsai, C.S.; Yang, Y.H.; Huang, K.Y.; Lee, Y.; McIntyre, R.S.; Chen, V.C. Association of Tic Disorders and Enterovirus Infection: A Nationwide Population-Based Study. Medicine 2016, 95, e3347. [Google Scholar] [CrossRef]
- Lin, J.N.; Lin, C.L.; Yen, H.R.; Yang, C.H.; Lai, C.H.; Lin, H.H.; Kao, C.H. Increased Risk of Tics in Children Infected with Enterovirus: A Nationwide Population-Based Study. J. Dev. Behav. Pediatrics JDBP 2017, 38, 276–282. [Google Scholar] [CrossRef]
- Antonelli, F.; Borghi, V.; Galassi, G.; Nichelli, P.; Valzania, F. Can HIV infection cause a worsening of tics in Tourette patients? Neurol. Sci. 2011, 32, 191–192. [Google Scholar] [CrossRef] [PubMed]
- Riedel, M.; Straube, A.; Schwarz, M.J.; Wilske, B.; Müller, N. Lyme disease presenting as Tourette’s syndrome. Lancet 1998, 351, 418–419. [Google Scholar] [CrossRef]
- Chang, Y.T.; Li, Y.F.; Muo, C.H.; Chen, S.C.; Chin, Z.N.; Kuo, H.T.; Lin, H.C.; Sung, F.C.; Tsai, C.H.; Chou, I.C. Correlation of Tourette syndrome and allergic disease: Nationwide population-based case-control study. J. Dev. Behav. Pediatrics JDBP 2011, 32, 98–102. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Chen, X.; Ke, N.; Pi, L.; Liu, Q. Association between allergic conjunctivitis and provisional tic disorder in children. Int. Ophthalmol. 2020, 40, 247–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zykov, V.P.; Shcherbina, A.Y.; Novikova, E.B.; Shvabrina, T.V. Neuroimmune aspects of the pathogenesis of Tourette’s syndrome and experience in the use of immunoglobulins in children. Neurosci. Behav. Physiol. 2009, 39, 635–638. [Google Scholar] [CrossRef]
- Müller, N. Anti-inflammatory therapy with a COX-2 inhibitor in Tourette’s syndrome. Inflammopharmacology 2004, 12, 271–275. [Google Scholar] [CrossRef]
- Matarazzo, E.B. Tourette’s Syndrome Treated with ACTH and Prednisone: Report of Two Cases. J. Child Adolesc. Psychopharmacol. 1992, 2, 215–226. [Google Scholar] [CrossRef]
- Hoekstra, P.J.; Minderaa, R.B.; Kallenberg, C.G. Lack of effect of intravenous immunoglobulins on tics: A double-blind placebo-controlled study. J. Clin. Psychiatry 2004, 65, 537–542. [Google Scholar] [CrossRef]
- Willsey, A.J.; Fernandez, T.V.; Yu, D.; King, R.A.; Dietrich, A.; Xing, J.; Sanders, S.J.; Mandell, J.D.; Huang, A.Y.; Richer, P.; et al. De Novo Coding Variants Are Strongly Associated with Tourette Disorder. Neuron 2017, 94, 486–499.e9. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Mandell, J.D.; Kumar, Y.; Sun, N.; Morris, M.T.; Arbelaez, J.; Nasello, C.; Dong, S.; Duhn, C.; Zhao, X.; et al. De Novo Sequence and Copy Number Variants Are Strongly Associated with Tourette Disorder and Implicate Cell Polarity in Pathogenesis. Cell Rep. 2018, 24, 3441–3454.e12. [Google Scholar] [CrossRef] [Green Version]
- Ercan-Sencicek, A.G.; Stillman, A.A.; Ghosh, A.K.; Bilguvar, K.; O’Roak, B.J.; Mason, C.E.; Abbott, T.; Gupta, A.; King, R.A.; Pauls, D.L. L-histidine decarboxylase and Tourette’s syndrome. N. Engl. J. Med. 2010, 362, 1901–1908. [Google Scholar] [PubMed] [Green Version]
- Pittenger, C. Histidine Decarboxylase Knockout Mice as a Model of the Pathophysiology of Tourette Syndrome and Related Conditions. Handb. Exp. Pharmacol. 2017, 241, 189–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtsu, H. Histamine synthesis and lessons learned from histidine decarboxylase deficient mice. In Histamine in Inflammation; Springer: Berlin/Heidelberg, Germany, 2010; pp. 21–31. [Google Scholar]
- Tian, Y.; Gunther, J.R.; Liao, I.H.; Liu, D.; Ander, B.P.; Stamova, B.S.; Lit, L.; Jickling, G.C.; Xu, H.; Zhan, X.; et al. GABA- and acetylcholine-related gene expression in blood correlate with tic severity and microarray evidence for alternative splicing in Tourette syndrome: A pilot study. Brain Res. 2011, 1381, 228–236. [Google Scholar] [CrossRef]
- Gunther, J.; Tian, Y.; Stamova, B.; Lit, L.; Corbett, B.; Ander, B.; Zhan, X.; Jickling, G.; Bos-Veneman, N.; Liu, D.; et al. Catecholamine-related gene expression in blood correlates with tic severity in tourette syndrome. Psychiatry Res. 2012, 200, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, Y.; Kalanithi, P.S.; Grantz, H.; Schwartz, M.L.; Saper, C.; Leckman, J.F.; Vaccarino, F.M. Decreased number of parvalbumin and cholinergic interneurons in the striatum of individuals with Tourette syndrome. J. Comp. Neurol. 2010, 518, 277–291. [Google Scholar]
- Rane, M.J.; Gozal, D.; Butt, W.; Gozal, E.; Pierce, W.M.; Guo, S.Z.; Wu, R.; Goldbart, A.D.; Thongboonkerd, V.; McLeish, K.R. γ-amino butyric acid type B receptors stimulate neutrophil chemotaxis during ischemia-reperfusion. J. Immunol. 2005, 174, 7242–7249. [Google Scholar]
- Zhou, F.M.; Wilson, C.J.; Dani, J.A. Cholinergic interneuron characteristics and nicotinic properties in the striatum. J. Neurobiol. 2002, 53, 590–605. [Google Scholar]
- Kimura, R.; Ushiyama, N.; Fujii, T.; Kawashima, K. Nicotine-induced Ca2+ signaling and down-regulation of nicotinic acetylcholine receptor subunit expression in the CEM human leukemic T-cell line. Life Sci. 2003, 72, 2155–2158. [Google Scholar]
- Marazziti, D.; Consoli, G.; Masala, I.; Catena Dell’Osso, M.; Baroni, S. Latest advancements on serotonin and dopamine transporters in lymphocytes. Mini Rev. Med. Chem. 2010, 10, 32–40. [Google Scholar]
- Dominguez-Villar, M.; Hafler, D.A. Regulatory T cells in autoimmune disease. Nat. Immunol. 2018, 19, 665–673. [Google Scholar] [CrossRef]
- Kawikova, I.; Leckman, J.F.; Kronig, H.; Katsovich, L.; Bessen, D.E.; Ghebremichael, M.; Bothwell, A.L. Decreased numbers of regulatory T cells suggest impaired immune tolerance in children with tourette syndrome: A preliminary study. Biol. Psychiatry 2007, 61, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Maia, T.V.; Conceição, V.A. Dopaminergic Disturbances in Tourette Syndrome: An Integrative Account. Biol. Psychiatry 2018, 84, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Kipnis, J.; Cardon, M.; Avidan, H.; Lewitus, G.M.; Mordechay, S.; Rolls, A.; Shani, Y.; Schwartz, M. Dopamine, through the extracellular signal-regulated kinase pathway, downregulates CD4+CD25+ regulatory T-cell activity: Implications for neurodegeneration. J. Neurosci. 2004, 24, 6133–6143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arreola, R.; Alvarez-Herrera, S.; Pérez-Sánchez, G.; Becerril-Villanueva, E.; Cruz-Fuentes, C.; Flores-Gutierrez, E.O.; Garcés-Alvarez, M.E.; de la Cruz-Aguilera, D.L.; Medina-Rivero, E.; Hurtado-Alvarado, G.; et al. Immunomodulatory Effects Mediated by Dopamine. J. Immunol. Res. 2016, 2016, 3160486. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, M.; Termine, C.; Franciotta, D.; Castiglioni, E.; Pagani, A.; Lanzi, G.; Marino, F.; Lecchini, S.; Cosentino, M.; Balottin, U. Dopaminergic receptor D5 mRNA expression is increased in circulating lymphocytes of Tourette syndrome patients. J. Psychiatr. Res. 2008, 43, 24–29. [Google Scholar] [CrossRef]
- Piccirillo, C.A.; Shevach, E.M. Naturally-occurring CD4+CD25+ immunoregulatory T cells: Central players in the arena of peripheral tolerance. Semin. Immunol. 2004, 16, 81–88. [Google Scholar] [CrossRef]
- Möller, J.; Tackenberg, B.; Heinzel-Gutenbrunner, M.; Burmester, R.; Oertel, W.; Bandmann, O.; Müller-Vahl, K. Immunophenotyping in Tourette syndrome—A pilot study. Eur. J. Neurol. 2008, 15, 749–753. [Google Scholar] [CrossRef]
- Marzio, R.; Mauël, J.; Betz-Corradin, S. CD69 and regulation of the immune function. Immunopharmacol. Immunotoxicol. 1999, 21, 565–582. [Google Scholar] [CrossRef]
- Green, D.R.; Droin, N.; Pinkoski, M. Activation-induced cell death in T cells. Immunol. Rev. 2003, 193, 70–81. [Google Scholar] [CrossRef]
- Yeon, S.M.; Lee, J.H.; Kang, D.; Bae, H.; Lee, K.Y.; Jin, S.; Kim, J.R.; Jung, Y.W.; Park, T.W. A cytokine study of pediatric Tourette’s disorder without obsessive compulsive disorder. Psychiatry Res. 2017, 247, 90–96. [Google Scholar] [CrossRef]
- Matz, J.; Krause, D.L.; Dehning, S.; Riedel, M.; Gruber, R.; Schwarz, M.J.; Müller, N. Altered monocyte activation markers in Tourette’s syndrome: A case-control study. BMC Psychiatry 2012, 12, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bos-Veneman, N.G.; Bijzet, J.; Limburg, P.C.; Minderaa, R.B.; Kallenberg, C.G.; Hoekstra, P.J. Cytokines and soluble adhesion molecules in children and adolescents with a tic disorder. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2010, 34, 1390–1395. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, P.J.; Anderson, G.M.; Troost, P.W.; Kallenberg, C.G.; Minderaa, R.B. Plasma kynurenine and related measures in tic disorder patients. Eur. Child Adolesc. Psychiatry 2007, 16 (Suppl. S1), 71–77. [Google Scholar] [CrossRef]
- Bos-Veneman, N.G.P.; Olieman, R.; Tobiasova, Z.; Hoekstra, P.J.; Katsovich, L.; Bothwell, A.L.M.; Leckman, J.F.; Kawikova, I. Altered immunoglobulin profiles in children with Tourette syndrome. Brain Behav. Immun. 2011, 25, 532–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawikova, I.; Grady, B.P.X.; Tobiasova, Z.; Zhang, Y.; Vojdani, A.; Katsovich, L.; Richmand, B.J.; Park, T.W.; Bothwell, A.L.M.; Leckman, J.F. Children with Tourette’s syndrome may suffer immunoglobulin A dysgammaglobulinemia: Preliminary report. Biol. Psychiatry 2010, 67, 679–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lennington, J.B.; Coppola, G.; Kataoka-Sasaki, Y.; Fernandez, T.V.; Palejev, D.; Li, Y.; Huttner, A.; Pletikos, M.; Sestan, N.; Leckman, J.F. Transcriptome analysis of the human striatum in Tourette syndrome. Biol. Psychiatry 2016, 79, 372–382. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Williams, M.T.; Chugani, H.T. Evaluation of basal ganglia and thalamic inflammation in children with pediatric autoimmune neuropsychiatric disorders associated with streptococcal infection and tourette syndrome: A positron emission tomographic (PET) study using 11C-[R]-PK11195. J. Child Neurol. 2015, 30, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Ziv, Y.; Ron, N.; Butovsky, O.; Landa, G.; Sudai, E.; Greenberg, N.; Cohen, H.; Kipnis, J.; Schwartz, M. Immune cells contribute to the maintenance of neurogenesis and spatial learning abilities in adulthood. Nat. Neurosci. 2006, 9, 268–275. [Google Scholar] [CrossRef]
- Ueno, M.; Fujita, Y.; Tanaka, T.; Nakamura, Y.; Kikuta, J.; Ishii, M.; Yamashita, T. Layer V cortical neurons require microglial support for survival during postnatal development. Nat. Neurosci. 2013, 16, 543–551. [Google Scholar] [CrossRef]
- Frick, L.; Rapanelli, M.; Abbasi, E.; Ohtsu, H.; Pittenger, C. Histamine regulation of microglia: Gene-environment interaction in the regulation of central nervous system inflammation. Brain Behav. Immun. 2016, 57, 326–337. [Google Scholar] [CrossRef] [Green Version]
- Frick, L.; Pittenger, C. Microglial Dysregulation in OCD, Tourette Syndrome, and PANDAS. J. Immunol. Res. 2016, 2016, 8606057. [Google Scholar] [CrossRef] [PubMed]
- Kirvan, C.A.; Cox, C.J.; Swedo, S.E.; Cunningham, M.W. Tubulin is a neuronal target of autoantibodies in Sydenham’s chorea. J. Immunol. 2007, 178, 7412–7421. [Google Scholar] [CrossRef] [PubMed]
- Kirvan, C.A.; Swedo, S.E.; Heuser, J.S.; Cunningham, M.W. Mimicry and autoantibody-mediated neuronal cell signaling in Sydenham chorea. Nat. Med. 2003, 9, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Chain, J.L.; Alvarez, K.; Mascaro-Blanco, A.; Reim, S.; Bentley, R.; Hommer, R.; Grant, P.; Leckman, J.F.; Kawikova, I.; Williams, K.; et al. Autoantibody Biomarkers for Basal Ganglia Encephalitis in Sydenham Chorea and Pediatric Autoimmune Neuropsychiatric Disorder Associated With Streptococcal Infections. Front. Psychiatry 2020, 11, 564. [Google Scholar] [CrossRef] [PubMed]
- Dale, R.C.; Merheb, V.; Pillai, S.; Wang, D.; Cantrill, L.; Murphy, T.K.; Ben-Pazi, H.; Varadkar, S.; Aumann, T.D.; Horne, M.K.; et al. Antibodies to surface dopamine-2 receptor in autoimmune movement and psychiatric disorders. Brain 2012, 135, 3453–3468. [Google Scholar] [CrossRef] [PubMed]
- Ben-Pazi, H.; Stoner, J.A.; Cunningham, M.W. Dopamine receptor autoantibodies correlate with symptoms in Sydenham’s chorea. PLoS ONE 2013, 8, e73516. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, M.W. Rheumatic fever, autoimmunity, and molecular mimicry: The streptococcal connection. Int. Rev. Immunol. 2014, 33, 314–329. [Google Scholar] [CrossRef] [Green Version]
- Lisman, J.; Schulman, H.; Cline, H. The molecular basis of CaMKII function in synaptic and behavioural memory. Nat. Rev. Neurosci. 2002, 3, 175–190. [Google Scholar] [CrossRef]
- Lotan, D.; Benhar, I.; Alvarez, K.; Mascaro-Blanco, A.; Brimberg, L.; Frenkel, D.; Cunningham, M.W.; Joel, D. Behavioral and neural effects of intra-striatal infusion of anti-streptococcal antibodies in rats. Brain Behav. Immun. 2014, 38, 249–262. [Google Scholar] [CrossRef] [Green Version]
- Baglioni, V.; Coutinho, E.; Menassa, D.A.; Giannoccaro, M.P.; Jacobson, L.; Buttiglione, M.; Petruzzelli, O.; Cardona, F.; Vincent, A. Antibodies to neuronal surface proteins in Tourette Syndrome: Lack of evidence in a European paediatric cohort. Brain Behav. Immun. 2019, 81, 665–669. [Google Scholar] [CrossRef]
- Sühs, K.-W.; Skripuletz, T.; Pul, R.; Alvermann, S.; Schwenkenbecher, P.; Stangel, M.; Müller-Vahl, K. Gilles de la Tourette syndrome is not linked to contactin-associated protein receptor 2 antibodies. Mol. Brain 2015, 8, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgaertel, C.; Skripuletz, T.; Kronenberg, J.; Stangel, M.; Schwenkenbecher, P.; Sinke, C.; Müller-Vahl, K.R.; Sühs, K.-W. Immunity in Gilles de la Tourette-Syndrome: Results From a Cerebrospinal Fluid Study. Front. Neurol. 2019, 10, 732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pranzatelli, M.R.; Tate, E.D.; Allison, T.J. Case-control, exploratory study of cerebrospinal fluid chemokines/cytokines and lymphocyte subsets in childhood Tourette syndrome with positive streptococcal markers. Cytokine 2017, 96, 49–53. [Google Scholar] [CrossRef]
- Hornig, M.; Lipkin, W.I. Immune-mediated animal models of Tourette syndrome. Neurosci. Biobehav. Rev. 2013, 37, 1120–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalcman, S.; Green-Johnson, J.M.; Murray, L.; Nance, D.M.; Dyck, D.; Anisman, H.; Greenberg, A.H. Cytokine-specific central monoamine alterations induced by interleukin-1,-2 and-6. Brain Res. 1994, 643, 40–49. [Google Scholar] [CrossRef]
- Zalcman, S.; Murray, L.; Dyck, D.G.; Greenberg, A.H.; Nance, D.M. Interleukin-2 and-6 induce behavioral-activating effects in mice. Brain Res. 1998, 811, 111–121. [Google Scholar] [CrossRef]
- Ponzio, N.M.; Servatius, R.; Beck, K.; Marzouk, A.; Kreider, T. Cytokine levels during pregnancy influence immunological profiles and neurobehavioral patterns of the offspring. Annal. N. Y. Acad. Sci. 2007, 1107, 118–128. [Google Scholar] [CrossRef]
- Smith, S.E.; Li, J.; Garbett, K.; Mirnics, K.; Patterson, P.H. Maternal immune activation alters fetal brain development through interleukin-6. J. Neurosci. 2007, 27, 10695–10702. [Google Scholar] [CrossRef] [Green Version]
- Zalcman, S.S.; Patel, A.; Mohla, R.; Zhu, Y.; Siegel, A. Soluble cytokine receptors (sIL-2Rα, sIL-2Rβ) induce subunit-specific behavioral responses and accumulate in the cerebral cortex and basal forebrain. PLoS ONE 2012, 7, e36316. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.; Zhu, Y.; Kuzhikandathil, E.V.; Banks, W.A.; Siegel, A.; Zalcman, S.S. Soluble interleukin-6 receptor induces motor stereotypies and co-localizes with gp130 in regions linked to cortico-striato-thalamo-cortical circuits. PLoS ONE 2012, 7, e41623. [Google Scholar] [CrossRef]
- Hoffman, K.L.; Hornig, M.; Yaddanapudi, K.; Jabado, O.; Lipkin, W.I. A murine model for neuropsychiatric disorders associated with group A β-hemolytic streptococcal infection. J. Neurosci. 2004, 24, 1780–1791. [Google Scholar] [CrossRef] [PubMed]
- Yaddanapudi, K.; Hornig, M.; Serge, R.; De Miranda, J.; Baghban, A.; Villar, G.; Lipkin, W.I. Passive transfer of streptococcus-induced antibodies reproduces behavioral disturbances in a mouse model of pediatric autoimmune neuropsychiatric disorders associated with streptococcal infection. Mol. Psychiatry 2010, 15, 712–726. [Google Scholar] [CrossRef] [PubMed]
- Ajmone-Cat, M.A.; Spinello, C.; Valenti, D.; Franchi, F.; Macri, S.; Vacca, R.A.; Laviola, G. Brain-Immune Alterations and Mitochondrial Dysfunctions in a Mouse Model of Paediatric Autoimmune Disorder Associated with Streptococcus: Exacerbation by Chronic Psychosocial Stress. J. Clin. Med. 2019, 8, 1514. [Google Scholar] [CrossRef] [Green Version]
- Hallett, J.J.; Harling-Berg, C.J.; Knopf, P.M.; Stopa, E.G.; Kiessling, L.S. Anti-striatal antibodies in Tourette syndrome cause neuronal dysfunction. J. Neuroimmunol. 2000, 111, 195–202. [Google Scholar] [PubMed]
- Taylor, J.R.; Morshed, S.A.; Parveen, S.; Mercadante, M.T.; Scahill, L.; Peterson, B.S.; King, R.A.; Leckman, J.F.; Lombroso, P.J. An animal model of Tourette’s syndrome. Am. J. Psychiatry 2002, 159, 657–660. [Google Scholar]
- Loiselle, C.R.; Lee, O.; Moran, T.H.; Singer, H.S. Striatal microinfusion of Tourette syndrome and PANDAS sera: Failure to induce behavioral changes. Mov. Disord. 2004, 19, 390–396. [Google Scholar]
- Ben-Pazi, H.; Sadan, O.; Offen, D. Striatal microinjection of Sydenham chorea antibodies: Using a rat model to examine the dopamine hypothesis. J. Mol. Neurosci. 2012, 46, 162–166. [Google Scholar]
- Zhang, D.; Patel, A.; Zhu, Y.; Siegel, A.; Zalcman, S.S. Anti-streptococcus IgM antibodies induce repetitive stereotyped movements: Cell activation and co-localization with Fcα/μ receptors in the striatum and motor cortex. Brain Behav. Immun. 2012, 26, 521–533. [Google Scholar]
- Baldan, L.C.; Williams, K.A.; Gallezot, J.-D.; Pogorelov, V.; Rapanelli, M.; Crowley, M.; Anderson, G.M.; Loring, E.; Gorczyca, R.; Billingslea, E. Histidine decarboxylase deficiency causes tourette syndrome: Parallel findings in humans and mice. Neuron 2014, 81, 77–90. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Li, L.; Ohtsu, H.; Pittenger, C. Histidine decarboxylase knockout mice, a genetic model of Tourette syndrome, show repetitive grooming after induced fear. Neurosci. Lett. 2015, 595, 50–53. [Google Scholar] [CrossRef] [Green Version]
- Greer, J.M.; Capecchi, M.R. Hoxb8 is required for normal grooming behavior in mice. Neuron 2002, 33, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.-K.; Tvrdik, P.; Peden, E.; Cho, S.; Wu, S.; Spangrude, G.; Capecchi, M.R. Hematopoietic origin of pathological grooming in Hoxb8 mutant mice. Cell 2010, 141, 775–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gariup, M.; Gonzalez, A.; Lázaro, L.; Torres, F.; Serra-Pagès, C.; Morer, A. IL-8 and the innate immunity as biomarkers in acute child and adolescent psychopathology. Psychoneuroendocrinology 2015, 62, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Morer, A.; Chae, W.; Henegariu, O.; Bothwell, A.L.; Leckman, J.F.; Kawikova, I. Elevated expression of MCP-1, IL-2 and PTPR-N in basal ganglia of Tourette syndrome cases. Brain Behav. Immun. 2010, 24, 1069–1073. [Google Scholar] [CrossRef] [PubMed]
- Zalcman, S.S. Interleukin-2-induced increases in climbing behavior: Inhibition by dopamine D-1 and D-2 receptor antagonists. Brain Res. 2002, 944, 157–164. [Google Scholar] [CrossRef]
- Singer, H.S.; Mink, J.W.; Loiselle, C.R.; Burke, K.A.; Ruchkina, I.; Morshed, S.; Parveen, S.; Leckman, J.F.; Hallett, J.J.; Lombroso, P.J. Microinfusion of antineuronal antibodies into rodent striatum: Failure to differentiate between elevated and low titers. J. Neuroimmunol. 2005, 163, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.-B.; Shui, H.-A.; Chu, T.-H.; Chen, Y.-A.; Tsung, H.-C.; Shyu, J.-F. Hyperpolarisation-activated cyclic nucleotide channel 4 (HCN4) involvement in Tourette’s syndrome autoimmunity. J. Neuroimmunol. 2012, 250, 18–26. [Google Scholar] [CrossRef]
- Ohtsu, H.; Tanaka, S.; Terui, T.; Hori, Y.; Makabe-Kobayashi, Y.; Pejler, G.; Tchougounova, E.; Hellman, L.; Gertsenstein, M.; Hirasawa, N. Mice lacking histidine decarboxylase exhibit abnormal mast cells. FEBS Lett. 2001, 502, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, R.; Santos, T.; Gonçalves, J.; Baltazar, G.; Ferreira, L.; Agasse, F.; Bernardino, L. Histamine modulates microglia function. J. Neuroinflamm. 2012, 9, 90. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Reference | Method | Specimen | Main Findings |
|---|---|---|---|
| [87] | CBA | Serum | Elevated D2R IgG in 4/44 (9%) |
| [92] | CBA | Serum | In a pediatric TS cohort of 30 siblings at preclinical and onset, 158 at chronic phase, 2 had NMDAR Ab weak positive, no other specific Abs (NMDAR, CASPR2, LGI1, AMPAR, and GABAAR) were detected; found Ab reactive with brain tissue, mainly to the hippocampus, the basal ganglia or the cerebellum in 12% |
| [93] | CBA | Serum | In 51 adult TS, no specific Abs (CASPR2, LGI1, NMDAR, AMPA1, AMPA/2, or GABAB1/B2) were detected |
| [25] | IF | CSF | Presence of OCB in 38% (8 of 21) adult TS |
| [94] | IF, CBA | CSF | Presence of OCB in 20% (4 of 20) adult TS; did not detect Abs to any of the antigens on SY5Y neuronal stem-cells and astrocytes cultures |
| [95] | IF | CSF | In 5 pediatric TS, none were detected for OCB and other immunoglobulins |
| Cytokine Injection | |||
| Reference | Method | Animal Type | Main Finding |
| [97] | Treated with IL-1, IL-2, and IL-6 | Mouse | IL-1: multiple central monoamine alteration and increased plasma corticosterone level IL-2: increased NE utilization and enhanced DA activity IL-6: increased activity of 5-HT and DA |
| [98] | Treated with IL-2 and IL-6 | Mouse | Increased digging, rearing, grooming and more ambulatory or nonambulatory exploration |
| [99] | Prenatal exposure with IL-2 | Mouse | Increased self-grooming and eye-blinking |
| [100] | Prenatal exposure with IL-6 | Mouse | Deficits of prepulse inhibition test |
| [101] | Injection of soluble IL-2 receptors | Mouse | Increased ambulatory behaviors and stereotypies Increased neuronal activities over cortex and striatum |
| [102] | Injection of soluble IL-6 receptors | Mouse | Induced repetitive stereotypies |
| Immunization with Specific Microorganism | |||
| Reference | Method | Main Finding | |
| [103] | Immunized with GAS homogenate | Mouse | Increased immunity response was observed in deep cerebellar nucleus, thalamus, and global pallidus |
| [104] | Immunized with GAS homogenate | Mouse | Increased rearing, submission, and defensive-escape behavior and decrease environmental exploration |
| [105] | Environmental psychosocial stress for GAS-infected mouse | Mouse | Altered cytokine, including IL-1β, TNF-α, IL-10 Impaired mitochondrial function |
| Passive Sera Transfusion | |||
| Reference | Method | Main Finding | |
| [106] | Intrastriatal microinjection | Rat | Increased motor stereotypies and episodic vocalization |
| [107] | Intrastriatal microinjection | Rat | Increased tic-like behaviors |
| [108,109] | Intrastriatal microinjection | Rat | No difference in behavioral change or immunohistology staining between experimental and control groups |
| [110] | Intravenous injection | Mouse | Antistreptococcus IgG stimulated vertical activities and ambulation Antistreptococcus IgM induced Fos-like immunoactivities in CSTC structures |
| Transgenic Animal Model | |||
| Reference | Method | Main Finding | |
| [52,53,111,112] | Hdc-knockout mice | Mouse | Increased stereotypes elevated repetitive behaviors Dysregulated dopamine level and microglia in neuroinflammatory process |
| [113,114] | Hoxb8-knockout mice | Mouse | Excessive grooming behaviors and hair loss Abnormal behaviors could be treated with bone marrow transplantation from wild-type mice |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, C.-J.; Wong, L.-C.; Lee, W.-T. Immunological Dysfunction in Tourette Syndrome and Related Disorders. Int. J. Mol. Sci. 2021, 22, 853. https://doi.org/10.3390/ijms22020853
Hsu C-J, Wong L-C, Lee W-T. Immunological Dysfunction in Tourette Syndrome and Related Disorders. International Journal of Molecular Sciences. 2021; 22(2):853. https://doi.org/10.3390/ijms22020853
Chicago/Turabian StyleHsu, Chia-Jui, Lee-Chin Wong, and Wang-Tso Lee. 2021. "Immunological Dysfunction in Tourette Syndrome and Related Disorders" International Journal of Molecular Sciences 22, no. 2: 853. https://doi.org/10.3390/ijms22020853
APA StyleHsu, C. -J., Wong, L. -C., & Lee, W. -T. (2021). Immunological Dysfunction in Tourette Syndrome and Related Disorders. International Journal of Molecular Sciences, 22(2), 853. https://doi.org/10.3390/ijms22020853