
Polycyclic Aromatic Hydrocarbons Detected in Processed Meats Cause Genetic Changes in Colorectal Cancers

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. PAH Detection via GC-MS
2.2. Recovery of PAHs Using QuEChERS Extraction
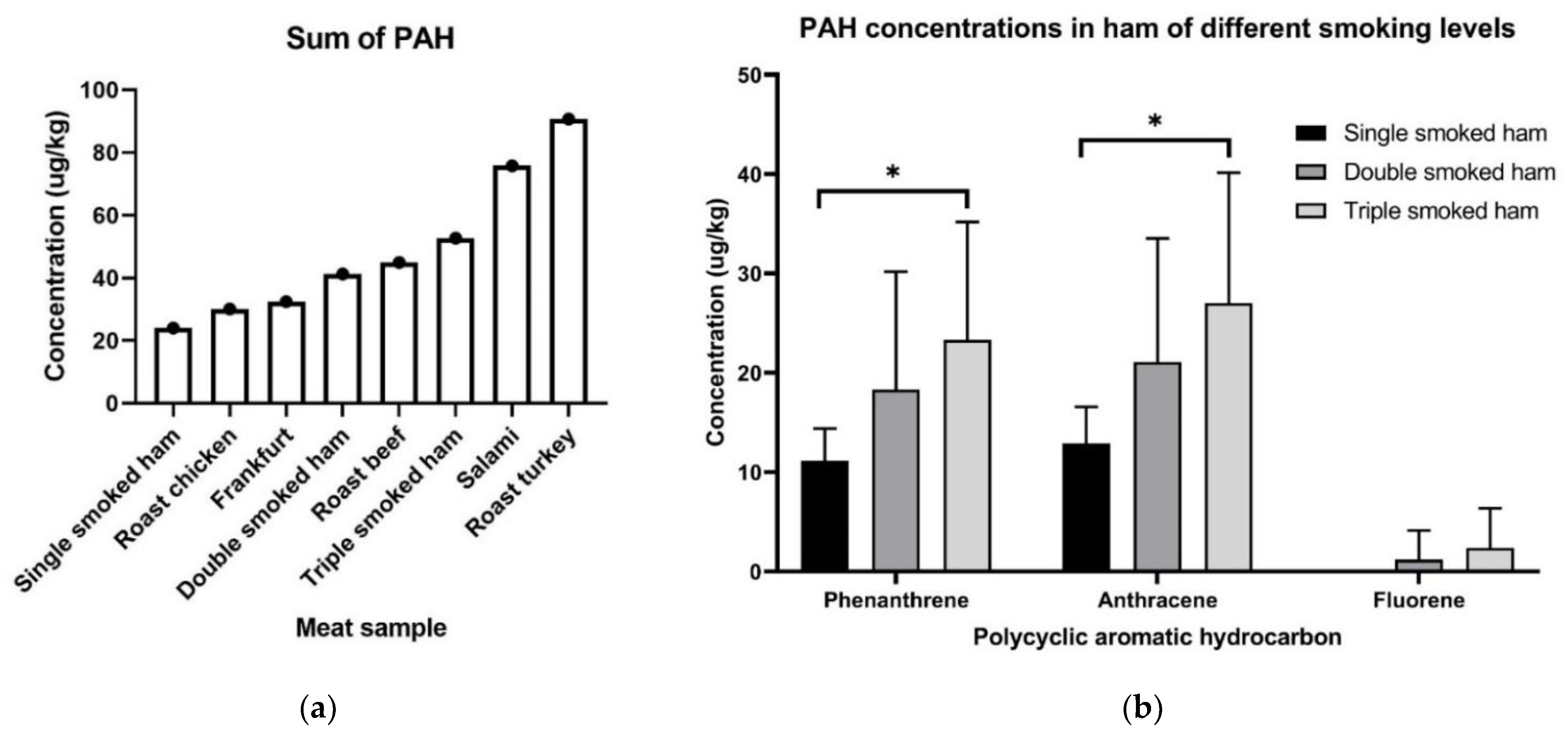
2.3. PAHs Are Found in Supermarket Meat Samples at Detectable Concentrations
2.4. Effect of Smoking in Ham on PAH
2.5. PAH Induced Genetic Changes In Vitro
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Meat Samples
4.3. Extraction and Clean-Up
4.4. GC-MS Equipment and Conditions
4.5. Recovery Study
4.6. Quantification of PAHs
4.7. Cell Culture
4.8. PAH Treatment
4.9. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (rtPCR)
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Prz. Gastroenterol. 2019, 14, 89–103. [Google Scholar] [CrossRef]
- Parkin, D.M.; Boyd, L.; Walker, L.C. 16. The fraction of cancer attributable to lifestyle and environmental factors in the UK in 2010. Br. J. Cancer 2011, 105, S77–S81. [Google Scholar] [CrossRef] [Green Version]
- Watson, A.J.M.; Collins, P.D. Colon cancer: A civilization disorder. Dig. Dis. 2011, 29, 222–228. [Google Scholar] [CrossRef]
- Hamidi, E.N.; Hajeb, P.; Selamat, J.; Razis, A.F.A. polycyclic aromatic hydrocarbons (pahs) and their bioaccessibility in meat: A tool for assessing human cancer risk. Asian Pac. J. Cancer Prev. 2016, 17, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Bouvard, V.; Loomis, D.; Guyton, K.Z.; Grosse, Y.; Ghissassi, F.E.; Benbrahim-Tallaa, L.; Guha, N.; Mattock, H.; Straif, K. International Agency for Research on Cancer Monograph working group carcinogenicity of consumption of red and processed meat. Lancet Oncol. 2015, 16, 1599–1600. [Google Scholar] [CrossRef] [Green Version]
- Zelinkova, Z.; Wenzl, T. The Occurrence of 16 EPA PAHs in Food—A Review. Polycycl. Aromat. Compd. 2015, 35, 248–284. [Google Scholar] [CrossRef] [Green Version]
- Duedahl-Olesen, L.; Ionas, A.C. Formation and mitigation of pahs in barbecued meat—A review. Crit. Rev. Food Sci. Nutr. 2021, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.H. Polycyclic Aromatic Hydrocarbons in the Diet. Mutat. Res. Genet. Toxicol. Environ. Mutagenes. 1999, 443, 139–147. [Google Scholar] [CrossRef]
- Australian Health Survey: Nutrition First Results-Foods and Nutrients, 2011–2012 Financial Year|Australian Bureau of Statistics. Available online: https://www.abs.gov.au/statistics/health/health-conditions-and-risks/australian-health-survey-nutrition-first-results-foods-and-nutrients/latest-release (accessed on 19 August 2021).
- Abdel-Shafy, H.I.; Mansour, M.S.M. A review on polycyclic aromatic hydrocarbons: Source, environmental impact, effect on human health and remediation. Egypt. J. Pet. 2016, 25, 107–123. [Google Scholar] [CrossRef] [Green Version]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Some Non-Heterocyclic Polycyclic aromatic hydrocarbons and some related exposures. IARC Monogr. Eval. Carcinog. Risks Hum. 2010, 92, 1–853. [Google Scholar]
- Moorthy, B.; Chu, C.; Carlin, D.J. Polycyclic aromatic hydrocarbons: From metabolism to lung cancer. Toxicol. Sci. 2015, 145, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lippi, G.; Mattiuzzi, C.; Cervellin, G. Meat consumption and cancer risk: A critical review of published meta-analyses. Crit. Rev. Oncol. Hematol. 2016, 97, 1–14. [Google Scholar] [CrossRef]
- Diggs, D.L.; Huderson, A.C.; Harris, K.L.; Myers, J.N.; Banks, L.D.; Rekhadevi, P.V.; Niaz, M.S.; Ramesh, A. Polycyclic aromatic hydrocarbons and digestive tract cancers—A perspective. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2011, 29, 324–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korsh, J.; Shen, A.; Aliano, K.; Davenport, T. Polycyclic aromatic hydrocarbons and breast cancer: A review of the literature. Breast Care 2015, 10, 316–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reszka, E.; Wasowicz, W.; Gromadzinska, J. Genetic polymorphism of xenobiotic metabolising enzymes, diet and cancer susceptibility. Br. J. Nutr. 2006, 96, 609–619. [Google Scholar]
- Coles, B.F.; Kadlubar, F.F. Human Alpha Class Glutathione S-Transferases: Genetic Polymorphism, Expression, and Susceptibility to Disease. Methods Enzymol. 2005, 401, 9–42. [Google Scholar]
- Chung, F.F.-L.; Mai, C.W.; Ng, P.Y.; Leong, C.-O. Cytochrome P450 2W1 (CYP2W1) in colorectal cancers. Curr. Cancer Drug Targets 2016, 16, 71–78. [Google Scholar] [CrossRef]
- Sinha, R.; Peters, U.; Cross, A.J.; Kulldorff, M.; Weissfeld, J.L.; Pinsky, P.F.; Rothman, N.; Hayes, R.B. Meat, Meat cooking methods and preservation, and risk for colorectal adenoma. Cancer Res. 2005, 65, 8034–8041. [Google Scholar] [CrossRef] [Green Version]
- Sinha, R.; Kulldorff, M.; Gunter, M.J.; Strickland, P.; Rothman, N. Dietary Benzo[a]Pyrene intake and risk of colorectal adenoma. Cancer Epidemiol. Biomark. Prev. 2005, 14, 2030–2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, P.E.; Lazarus, P.; Lesko, S.M.; Cross, A.J.; Sinha, R.; Laio, J.; Zhu, J.; Harper, G.; Muscat, J.E.; Hartman, T.J. Meat-related compounds and colorectal cancer risk by anatomical subsite. Nutr. Cancer 2013, 65, 202–226. [Google Scholar] [CrossRef]
- Wohak, L.E.; Krais, A.M.; Kucab, J.E.; Stertmann, J.; Øvrebø, S.; Seidel, A.; Phillips, D.H.; Arlt, V.M. Carcinogenic polycyclic aromatic hydrocarbons induce cyp1a1 in human cells via a P53-dependent mechanism. Arch. Toxicol. 2016, 90, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Alexandrov, K.; Rojas, M.; Kadlubar, F.F.; Lang, N.P.; Bartsch, H. Evidence of anti-benzo[a]pyrene diolepoxide-dna adduct formation in human colon mucosa. Carcinogenesis 1996, 17, 2081–2083. [Google Scholar] [CrossRef] [PubMed]
- Armaghany, T.; Wilson, J.D.; Chu, Q.; Mills, G. Genetic alterations in colorectal cancer. Gastrointest. Cancer Res. 2012, 5, 19–27. [Google Scholar]
- Al-Thaiban, H.; Al-Tamimi, N.; Helaleh, M. Development of QuEChERS Method for the Determination of Polycyclic Aromatic Hydrocarbons in Smoked Meat Products Using GC-MS from Qatar. Available online: https://www.hindawi.com/journals/jamc/2018/9206237/ (accessed on 23 February 2021).
- 2002/657/EC: Commission Decision of 12 August 2002 Implementing Council Directive 96/23/EC Concerning the Performance of Analytical Methods and the Interpretation of Results (Text with EEA Relevance) (Notified under Document Number C(2002) 3044). Available online: https://op.europa.eu/en/publication-detail/-/publication/ed928116-a955-4a84-b10a-cf7a82bad858/language-en (accessed on 10 September 2021).
- García-Falcón, M.S.; Simal-Gándara, J. Polycyclic aromatic hydrocarbons in smoke from different woods and their transfer during traditional smoking into chorizo sausages with collagen and tripe casings. Food Addit. Contam. 2005, 22, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Jakszyn, P.; Agudo, A.; Ibáñez, R.; García-Closas, R.; Pera, G.; Amiano, P.; González, C.A. Development of a food database of nitrosamines, heterocyclic amines, and polycyclic aromatic hydrocarbons. J. Nutr. 2004, 134, 2011–2014. [Google Scholar] [CrossRef]
- Farhadian, A.; Jinap, S.; Abas, F.; Sakar, Z.I. Determination of polycyclic aromatic hydrocarbons in grilled meat. Food Control 2010, 21, 606–610. [Google Scholar] [CrossRef]
- Babaoglu, A.S.; Karakaya, M.; Öz, F. Formation of polycyclic aromatic hydrocarbons in beef and lamb kokorec: Effects of different animal fats. Int. J. Food Prop. 2017, 20, 1960–1970. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-G.; Kim, S.-Y.; Moon, J.-S.; Kim, S.-H.; Kang, D.-H.; Yoon, H.-J. Effects of grilling procedures on levels of polycyclic aromatic hydrocarbons in grilled meats. Food Chem. 2016, 199, 632–638. [Google Scholar] [CrossRef]
- Zuo, J.; Brewer, D.S.; Arlt, V.M.; Cooper, C.S.; Phillips, D.H. Benzo Pyrene-Induced DNA adducts and gene expression profiles in target and non-target organs for carcinogenesis in mice. BMC Genom. 2014, 15, 880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Center for Biotechnology Information (US). The P53 Tumor Suppressor Protein; National Center for Biotechnology Information (US): Bethesda, MI, USA, 1998.
- Liu, Z.; Muehlbauer, K.-R.; Schmeiser, H.H.; Hergenhahn, M.; Belharazem, D.; Hollstein, M.C. P53 mutations in benzo(a)pyrene-exposed human p53 knock-in murine fibroblasts correlate with p53 mutations in human lung tumors. Cancer Res. 2005, 65, 2583–2587. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-Y.; Lee, S.-M.; Ye, S.-K.; Yoon, S.-H.; Chung, M.-H.; Choi, J. Benzo[a]Pyrene-induced dna damage and p53 modulation in human hepatoma HepG2 cells for the identification of potential biomarkers for pah monitoring and risk assessment. Toxicol. Lett. 2006, 167, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Li, Y.; Sun, Y.; Long, J.; Kong, Y.; Yang, S.; Wang, Y. A Common Carcinogen Benzo[a]Pyrene Causes P53 overexpression in mouse cervix via dna damage. Mutat. Res. Genet. Toxicol. Environ. Mutagenes. 2011, 724, 69–75. [Google Scholar] [CrossRef] [PubMed]
- KRAS KRAS Proto-Oncogene, GTPase [Homo Sapiens (Human)]-Gene-NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/3845 (accessed on 14 July 2021).
- Jerina, D.M.; Sayer, J.M.; Thakker, D.R.; Yagi, H.; Levin, W.; Wood, A.W.; Conney, A.H. Carcinogenicity of polycyclic aromatic hydrocarbons: The bay-region theory. In Carcinogenesis: Fundamental Mechanisms and Environmental Effects; Pullman, B., Ts’o, P.O.P., Gelboin, H., Eds.; Springer: Dordrecht, The Netherlands, 1980; pp. 1–12. [Google Scholar]
- Gamage, S.M.K.; Islam, F.; Cheng, T.; Aktar, S.; Lu, C.T.; Ranaweera, C.D.; Lee, K.T.W.; Dissabandara, L.; Gopalan, V.; Lam, A.K. HFE variants in colorectal cancer and their clinicopathological correlations. Hum. Pathol. 2021, 117, 9–30. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Polycyclic Aromatic Hydrocarbon | IARC Group |
|---|---|
| Acenaphthylene | N/A |
| Anthracene | 3 |
| Benz[a]anthracene | 2B |
| Benzo[a]pyrene | 1 |
| Benzo[b]fluoranthene | 2B |
| Benzo[ghi]perylene | 3 |
| Benzo[k]fluoranthene | 2B |
| Chrysene | 2B |
| Dibenz[a,h]anthracene | 2A |
| Fluorene | 3 |
| Indeno[1,2,3-cd]pyrene | 2B |
| Phenanthrene | 3 |
| Pyrene | 3 |
| Standards | Mass (g) | Concentration Range (ppb) | Curve Equation | Regression Coefficient |
|---|---|---|---|---|
| Acenaphthylene | 152.00 > 150.10 | 10–200 | 0.9977 | |
| Fluorene | 165.00 > 163.10 | 10–200 | 0.9971 | |
| Phenanthrene | 178.00 > 152.10 | 10–200 | 0.9948 | |
| Anthracene | 178.00 > 176.10 | 10–200 | 0.9967 | |
| Pyrene | 202.00 > 200.10 | 10–200 | 0.9933 | |
| Benzo[a]anthracene | 228.00 > 226.10 | 10–200 | 0.9977 | |
| Chrysene | 228.00 > 226.00 | 10–200 | 0.9960 | |
| Benzo[b]fluoranthene | 252.00 > 250.00 | 10–200 | 0.9961 | |
| Benzo[k]fluoranthene | 252.00 > 250.10 | 10–200 | 0.9963 | |
| Benzo[a]pyrene | 252.00 > 250.10 | 10–200 | 0.9945 | |
| Indeno[1,2,3-cd]pyrene | 276.00 > 274.00 | 10–200 | 0.9932 | |
| Dibenz[a,h]anthracene | 278.00 > 276.10 | 10–200 | 0.9961 | |
| Benzo[ghi]perylene | 276.00 > 274.00 | 10–200 | 0.9951 |
| PAH | Spiked Concentration ppb | Mean Concentration Detected ppb | Unspiked Blank Concentration ppb | Mean Recovery % |
|---|---|---|---|---|
| Benzo[b]fluoranthene | 1000 | 943.5 | 0 | 94.4 |
| Dibenz[a,h]anthracene | 1000 | 744.5 | 0 | 74.5 |
| Sample Name | Meat | Preparation Method | Number of Replicates (Technical; Biological) |
|---|---|---|---|
| Roast turkey | Turkey | Roasted | 2; 3 |
| Roast beef | Beef | Roasted | 2; 3 |
| Roast chicken | Chicken | Roasted | 2; 3 |
| Frankfurt | Pork | Smoked meat trimmings | 2; 3 |
| Single smoked ham | Pork | Single smoked | 2; 3 |
| Double smoked ham | Pork | Double smoked | 2; 3 |
| Triple smoked ham | Pork | Triple smoked | 2; 3 |
| Salami | Pork | Fermented and matured with smoke | 2; 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, T.; Chaousis, S.; Kodagoda Gamage, S.M.; Lam, A.K.-y.; Gopalan, V. Polycyclic Aromatic Hydrocarbons Detected in Processed Meats Cause Genetic Changes in Colorectal Cancers. Int. J. Mol. Sci. 2021, 22, 10959. https://doi.org/10.3390/ijms222010959
Cheng T, Chaousis S, Kodagoda Gamage SM, Lam AK-y, Gopalan V. Polycyclic Aromatic Hydrocarbons Detected in Processed Meats Cause Genetic Changes in Colorectal Cancers. International Journal of Molecular Sciences. 2021; 22(20):10959. https://doi.org/10.3390/ijms222010959
Chicago/Turabian StyleCheng, Tracie, Stephanie Chaousis, Sujani Madhurika Kodagoda Gamage, Alfred King-yin Lam, and Vinod Gopalan. 2021. "Polycyclic Aromatic Hydrocarbons Detected in Processed Meats Cause Genetic Changes in Colorectal Cancers" International Journal of Molecular Sciences 22, no. 20: 10959. https://doi.org/10.3390/ijms222010959
APA StyleCheng, T., Chaousis, S., Kodagoda Gamage, S. M., Lam, A. K. -y., & Gopalan, V. (2021). Polycyclic Aromatic Hydrocarbons Detected in Processed Meats Cause Genetic Changes in Colorectal Cancers. International Journal of Molecular Sciences, 22(20), 10959. https://doi.org/10.3390/ijms222010959