
Protein Cargo of Salivary Small Extracellular Vesicles as Potential Functional Signature of Oral Squamous Cell Carcinoma

 ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
2. Results
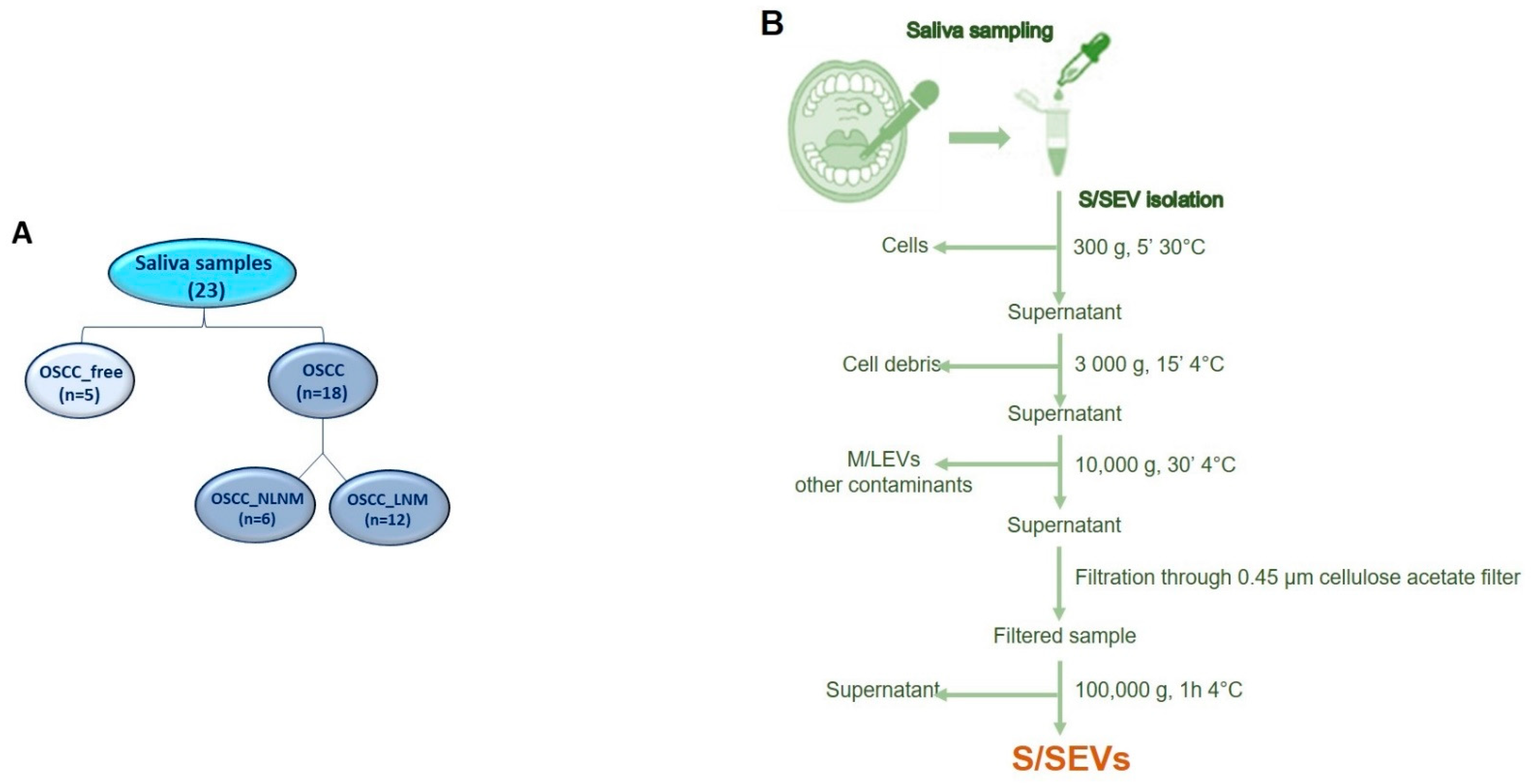
2.1. Enrolled Subjects and Sample Collections
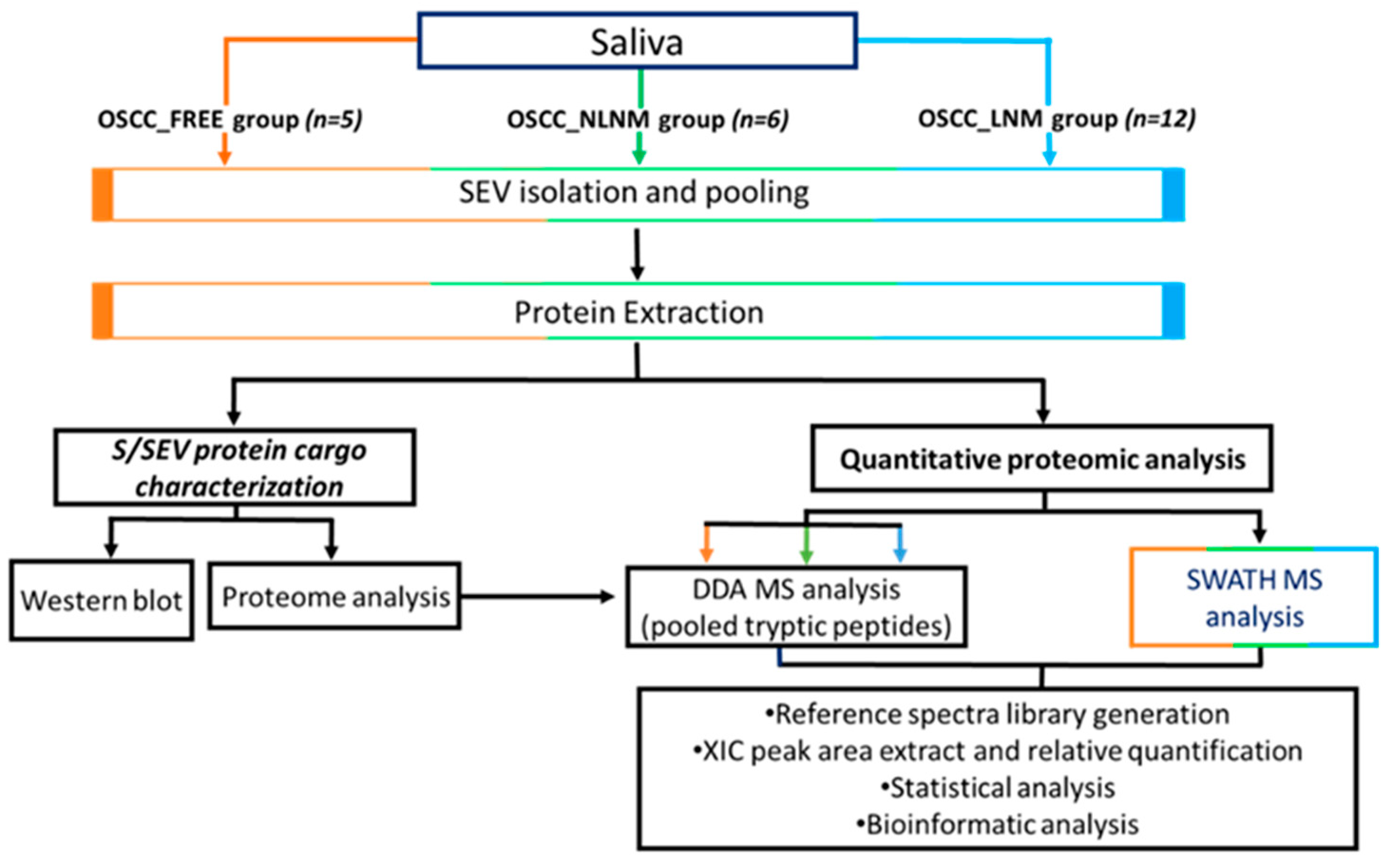
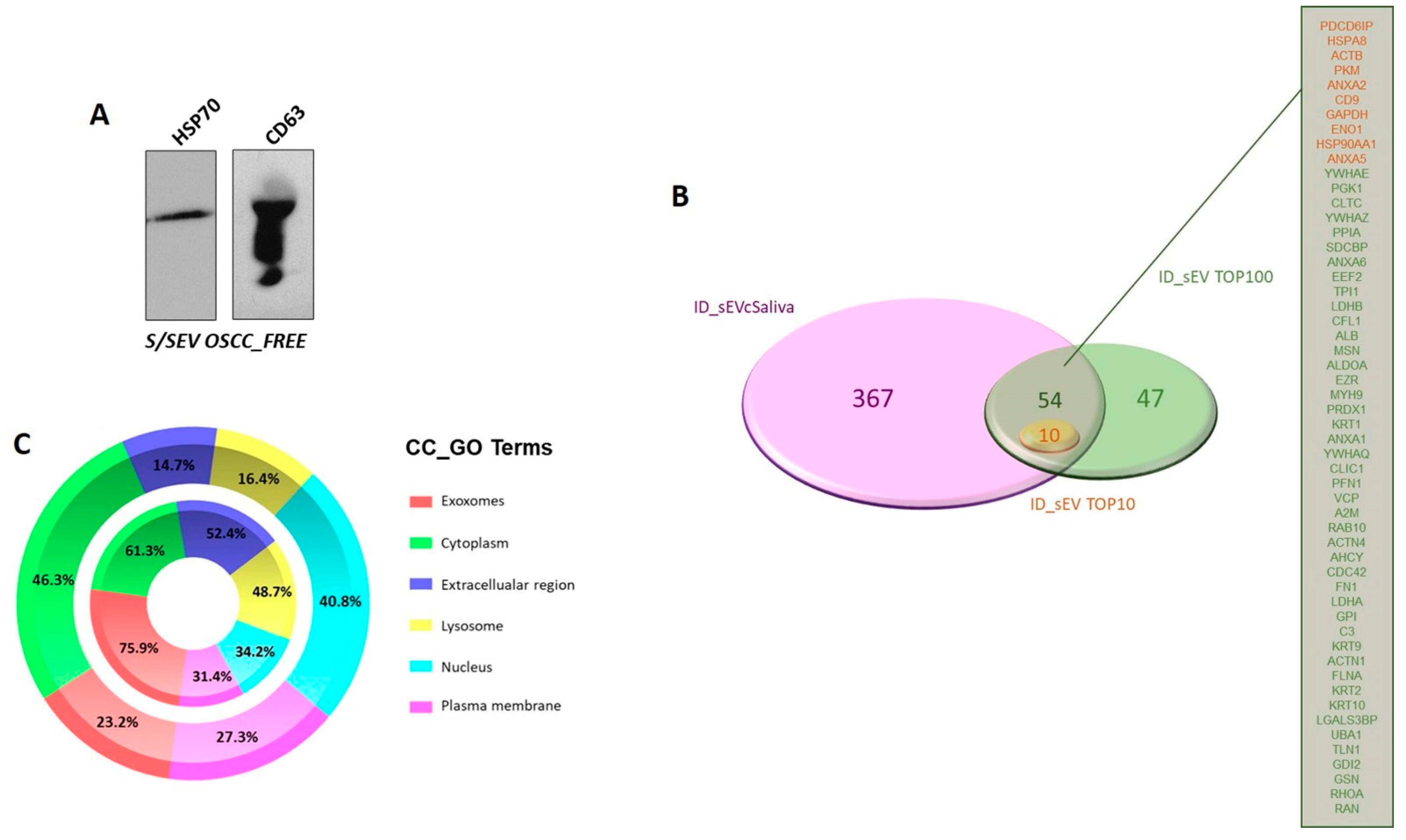
2.2. S/SEV Isolation and Protein Cargo Characterization
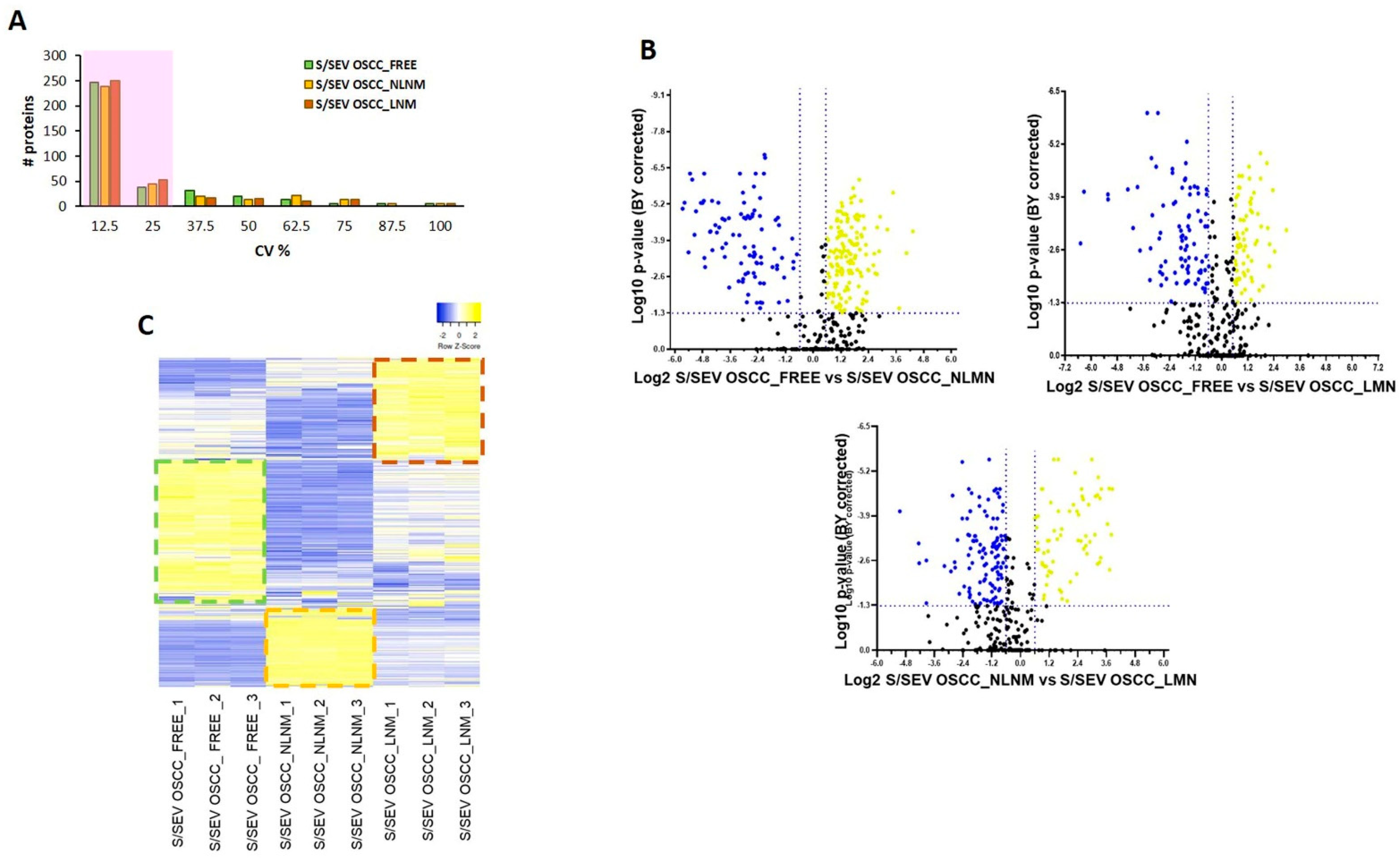
2.3. Protein Profile Characterization of S/SEVs
3. Discussion
4. Materials and Methods
4.1. Subject Enrolment and Saliva Collection
4.2. Saliva SEV Isolation
4.3. Western Blot
4.4. Proteomic Analyses: In-Solution Protein Digestion and SWATH-MS Analysis
4.5. Bioinformatic and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA. Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Rivera, C. Essentials of oral cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 11884–11894. [Google Scholar]
- Coxon, D.; Campbell, C.; Walter, F.M.; Scott, S.E.; Neal, R.D.; Vedsted, P.; Emery, J.; Rubin, G.; Hamilton, W.; Weller, D. The Aarhus statement on cancer diagnostic research: Turning recommendations into new survey instruments. BMC Health Serv. Res. 2018, 18, 677. [Google Scholar] [CrossRef]
- Fuller, C.; Camilon, R.; Nguyen, S.; Jennings, J.; Day, T.; Gillespie, M.B. Adjunctive diagnostic techniques for oral lesions of unknown malignant potential: Systematic review with meta-analysis. Head Neck 2014, 37, 755–762. [Google Scholar] [CrossRef]
- Mascitti, M.; Orsini, G.; Tosco, V.; Monterubbianesi, R.; Balercia, A.; Putignano, A.; Procaccini, M.; Santarelli, A. An overview on current non-invasive diagnostic devices in oral oncology. Front. Physiol. 2018, 9, 1510. [Google Scholar] [CrossRef] [Green Version]
- Babayan, A.; Pantel, K. Advances in liquid biopsy approaches for early detection and monitoring of cancer. Genome Med. 2018, 10, 1–3. [Google Scholar] [CrossRef]
- Cristaldi, M.; Mauceri, R.; Di Fede, O.; Giuliana, G.; Campisi, G.; Panzarella, V. Salivary biomarkers for oral squamous cell carcinoma diagnosis and follow-up: Current status and perspectives. Front. Physiol. 2019, 10, 1476. [Google Scholar] [CrossRef]
- Gissi, D.B.; Morandi, L.; Colella, G.; De Luca, R.; Ssd, G.C.; Mauceri, R.; Romeo, U.; Tenore, G.; Mignogna, M.D.; Adamo, D.; et al. Clinical validation of 13-gene DNA methylation analysis in oral brushing samples for detection of oral carcinoma: Italian multicenter study. Head Neck 2021, 43, 1563–1573. [Google Scholar] [CrossRef]
- Schmidt, H.; Kulasinghe, A.; Perry, C.; Nelson, C.; Punyadeera, C. A liquid biopsy for head and neck cancers. Expert Rev. Mol. Diagn. 2016, 16, 165–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lousada-Fernandez, F.; Rapado-Gonzalez, O.; Lopez-Cedrun, J.-L.; Lopez-Lopez, R.; Muinelo-Romay, L.; Suarez-Cunqueiro, M.M. Liquid biopsy in oral cancer. Int. J. Mol. Sci. 2018, 19, 1704. [Google Scholar] [CrossRef] [Green Version]
- de Miguel Perez, D.; Rodriguez Martinez, A.; Ortigosa Palomo, A.; Delgado Urena, M.; Garcia Puche, J.L.; Robles Remacho, A.; Exposito Hernandez, J.; Lorente Acosta, J.A.; Ortega Sanchez, F.G.; Serrano, M.J. Extracellular vesicle-miRNAs as liquid biopsy biomarkers for disease identification and prognosis in metastatic colorectal cancer patients. Sci. Rep. 2020, 10, 3974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitts, C.A.; Ji, N.; Li, Y.; Tan, C. Exploiting Exosomes in Cancer Liquid Biopsies and Drug Delivery. Adv. Healthc. Mater. 2019, 8, 18012688. [Google Scholar] [CrossRef] [PubMed]
- Giannopoulou, L.; Zavridou, M.; Kasimir-Bauer, S.; Lianidou, E.S. Liquid biopsy in ovarian cancer: The potential of circulating miRNAs and exosomes. Transl. Res. 2019, 205, 77–91. [Google Scholar] [CrossRef]
- Zheng, X.; Li, X.; Wang, X. Extracellular vesicle-based liquid biopsy holds great promise for the management of ovarian cancer. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188395. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raimondo, S.; Pucci, M.; Alessandro, R.; Fontana, S. Extracellular vesicles and tumor-immune escape: Biological functions and clinical perspectives. Int. J. Mol. Sci. 2020, 21, 2286. [Google Scholar] [CrossRef] [Green Version]
- Wee, I.; Syn, N.; Sethi, G.; Goh, B.C.; Wang, L. Role of tumor-derived exosomes in cancer metastasis. Biochim. Biophys. Acta Rev. Cancer 2019, 1871, 12–19. [Google Scholar] [CrossRef]
- Whiteside, T.L. Tumor-derived exosomes and their role in cancer progression. Adv. Clin. Chem. 2016, 74, 103–141. [Google Scholar] [PubMed] [Green Version]
- Conigliaro, A.; Fontana, S.; Raimondo, S.; Alessandro, R. Exosomes: Nanocarriers of biological messages. Adv. Exp. Med. Biol. 2017, 998, 23–43. [Google Scholar] [PubMed]
- Czystowska-Kuzmicz, M.; Whiteside, T.L. The potential role of tumor-derived exosomes in diagnosis, prognosis, and response to therapy in cancer. Expert Opin. Biol. Ther. 2021, 21, 241–258. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zhong, J.; Zhong, B.; Huang, J.; Jiang, L.; Jiang, Y.; Yuan, J.; Sun, J.; Dai, L.; Yang, C.; et al. Exosomes as potential sources of biomarkers in colorectal cancer. Cancer Lett. 2020, 476, 13–22. [Google Scholar] [CrossRef]
- Lee, J.M.; Garon, E.; Wong, D.T. Salivary diagnostics. Orthod. Craniofac. Res. 2009, 12, 206–211. [Google Scholar] [CrossRef]
- Malamud, D. Saliva as a diagnostic fluid. Dent. Clin. North. Am. 2011, 55, 159–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshizawa, J.M.; Schafer, C.A.; Schafer, J.J.; Farrell, J.J.; Paster, B.J.; Wong, D.T.W. Salivary biomarkers: Toward future clinical and diagnostic utilities. Clin. Microbiol. Rev. 2013, 26, 781–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonaka, T.; Wong, D.T.W. Saliva-exosomics in cancer: Molecular characterization of cancer-derived exosomes in saliva. Enzymes 2017, 42, 125–151. [Google Scholar]
- Corfield, A.P. Mucins: A biologically relevant glycan barrier in mucosal protection. Biochim. Biophys. Acta 2015, 1850, 236–252. [Google Scholar] [CrossRef]
- Gibbins, H.L.; Proctor, G.B.; Yakubov, G.E.; Wilson, S.; Carpenter, G.H. SIgA binding to mucosal surfaces is mediated by mucin-mucin interactions. PLoS ONE 2015, 10, 0119677. [Google Scholar] [CrossRef] [Green Version]
- Hannig, C.; Hannig, M.; Kensche, A.; Carpenter, G. The mucosal pellicle – An underestimated factor in oral physiology. Arch. Oral Biol. 2017, 80, 144–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Rosa, G.R.M.; Gattuso, G.; Pedullà, E.; Rapisarda, E.; Nicolosi, D.; Salmeri, M. Association of oral dysbiosis with oral cancer development (Review). Oncol. Lett. 2020, 19, 3045–3058. [Google Scholar] [CrossRef] [Green Version]
- Sardaro, N.; Della Vella, F.; Incalza, M.A.; di Stasio, D.; Lucchese, A.; Contaldo, M.; Laudadio, C.; Petruzzi, M. Oxidative stress and oral mucosal diseases: An overview. In Vivo 2019, 33, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.-S.; Tang, Y.-L.; Pang, X.; Zheng, M.; Liang, X.-H. The maintenance of an oral epithelial barrier. Life Sci. 2019, 227, 129–136. [Google Scholar] [CrossRef]
- Shalapour, S.; Karin, M. Immunity, inflammation, and cancer: An eternal fight between good and evil. J. Clin. Investig. 2015, 125, 3347–3355. [Google Scholar] [CrossRef] [Green Version]
- Afshar-Kharghan, V. The role of the complement system in cancer. J. Clin. Investig. 2017, 127, 780–789. [Google Scholar] [CrossRef] [Green Version]
- Sahu, S.; Routray, S. Assessing the analytical efficacy of TEX in diagnosing oral cancer using a systematic review approach. J. Oral Pathol. Med. 2021, 50, 123–128. [Google Scholar] [CrossRef]
- Gai, C.; Camussi, F.; Broccoletti, R.; Gambino, A.; Cabras, M.; Molinaro, L.; Carossa, S.; Camussi, G.; Arduino, P.G. Salivary extracellular vesicle-associated miRNAs as potential biomarkers in oral squamous cell carcinoma. BMC Cancer 2018, 18, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chiabotto, G.; Gai, C.; Deregibus, M.C.; Camussi, G. Salivary Extracellular Vesicle-Associated exRNA as Cancer Biomarker. Cancers 2019, 11, 891. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, M.; Zargartalebi, H.; Salahandish, R.; Aburashed, R.; Wey Yong, K.; Sanati-Nezhad, A. Emerging technologies and commercial products in exosome-based cancer diagnosis and prognosis. Biosens. Bioelectron. 2021, 183, 113176. [Google Scholar] [CrossRef] [PubMed]
- Ghantasala, S.; Gollapalli, K.; Epari, S.; Moiyadi, A.; Srivastava, S. Glioma tumor proteomics: Clinically useful protein biomarkers and future perspectives. Expert Rev. Proteom. 2020, 17, 221–232. [Google Scholar] [CrossRef]
- Ludwig, C.; Gillet, L.; Rosenberger, G.; Amon, S.; Collins, B.C.; Aebersold, R. Data-independent acquisition-based SWATH-MS for quantitative proteomics: A tutorial. Mol. Syst. Biol. 2018, 14, 8126. [Google Scholar] [CrossRef]
- Yu, L.; Yang, Z.; Liu, Y.; Liu, F.; Shang, W.; Shao, W.; Wang, Y.; Xu, M.; Wang, Y.-N.; Fu, Y.; et al. Identification of SPRR3 as a novel diagnostic/prognostic biomarker for oral squamous cell carcinoma via RNA sequencing and bioinformatic analyses. PeerJ 2020, 8, 9393. [Google Scholar] [CrossRef]
- Leiva, O.; Newcomb, R.; Connors, J.M.; Al-Samkari, H. Cancer and thrombosis: New insights to an old problem. J. Med. Vasc. 2020, 45, 6S8–6S16. [Google Scholar]
- Sherman, M.H. Lipid carriers in cancer: Context matters. Cancer Res. 2021, 81, 4186–4187. [Google Scholar] [CrossRef]
- Bång-Rudenstam, A.; Cerezo-Magaña, M.; Belting, M. Pro-metastatic functions of lipoproteins and extracellular vesicles in the acidic tumor microenvironment. Cancer Metastasis Rev. 2019, 38, 79–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jukic, A.; Bakiri, L.; Wagner, E.F.; Tilg, H.; Adolph, T.E. Calprotectin: From biomarker to biological function. Gut 2021, 70, 1978–1988. [Google Scholar] [CrossRef]
- Wiesner, J.; Vilcinskas, A. Antimicrobial peptides: The ancient arm of the human immune system. Virulence 2010, 1, 440–464. [Google Scholar] [CrossRef]
- Dziarski, R.; Gupta, D. Review: Mammalian peptidoglycan recognition proteins (PGRPs) in innate immunity. Innate Immun. 2010, 16, 168–174. [Google Scholar] [CrossRef]
- Rosa, L.; Lepanto, M.S.; Cutone, A.; Ianiro, G.; Pernarella, S.; Sangermano, R.; Musci, G.; Ottolenghi, L.; Valenti, P. Lactoferrin and oral pathologies: A therapeutic treatment. Biochem. Cell Biol. 2021, 99, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, S.; Roushandeh, A.M.; Ahmadzadeh, E.; Jahanian-Najafabadi, A.; Roudkenar, M.H. Implication and role of neutrophil gelatinase-associated lipocalin in cancer: Lipocalin-2 as a potential novel emerging comprehensive therapeutic target for a variety of cancer types. Mol. Bio.l Rep. 2020, 47, 2327–2346. [Google Scholar] [CrossRef]
- Chattopadhyay, I.; Verma, M.; Panda, M. Role of oral microbiome signatures in diagnosis and prognosis of oral cancer. Technol. Cancer Res. Treat. 2019, 18, 1533033819867354. [Google Scholar] [CrossRef] [Green Version]
- Irfan, M.; Delgado, R.Z.R.; Frias-Lopez, J. The oral microbiome and cancer. Front. Immunol. 2020, 11, 591088. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Tang, Q.; Yu, S.; Xie, M.; Xie, Y.; Chen, G.; Chen, L. Role of the oral microbiota in cancer evolution and progression. Cancer Med. 2020, 9, 6306–6321. [Google Scholar] [CrossRef] [PubMed]
- Şenel, S. An overview of physical, microbiological and immune barriers of oral mucosa. Int. J. Mol. Sci. 2021, 22, 7821. [Google Scholar] [CrossRef] [PubMed]
- Argyris, P.; Slama, Z.; Ross, K.; Khammanivong, A.; Herzberg, M. Calprotectin and the initiation and progression of head and neck cancer. J. Dent. Res. 2018, 97, 674–682. [Google Scholar] [CrossRef]
- Raffat, M.A.; Hadi, N.I.; Hosein, M.; Mirza, S.; Ikram, S.; Akram, Z. S100 proteins in oral squamous cell carcinoma. Clin. Chim. Acta 2018, 480, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Raimondi, L.; De Luca, A.; Amodio, N.; Manno, M.; Raccosta, S.; Taverna, S.; Bellavia, D.; Naselli, F.; Fontana, S.; Schillaci, O.; et al. Involvement of multiple myeloma cell-derived exosomes in osteoclast differentiation. Oncotarget 2015, 6, 13772–13789. [Google Scholar] [CrossRef] [Green Version]
- Schillaci, O.; Fontana, S.; Monteleone, F.; Taverna, S.; Di Bella, M.A.; Di Vizio, D.; Alessandro, R. Exosomes from metastatic cancer cells transfer amoeboid phenotype to non-metastatic cells and increase endothelial permeability: Their emerging role in tumor heterogeneity. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Pathan, M.; Keerthikumar, S.; Chisanga, D.; Alessandro, R.; Ang, C.-S.; Askenase, P.; Batagov, A.O.; Martin, A.B.; Camussi, G.; Clayton, A.; et al. A novel community driven software for functional enrichment analysis of extracellular vesicles data. J. Extracell. Vesicles 2017, 6, 1321455. [Google Scholar] [CrossRef] [Green Version]
- Monteleone, F.; Taverna, S.; Alessandro, R.; Fontana, S. SWATH-MS based quantitative proteomics analysis reveals that curcumin alters the metabolic enzyme profile of CML cells by affecting the activity of miR-22/IPO7/HIF-1alpha axis. J. Exp. Clin. Cancer Res. 2018, 37, 170. [Google Scholar] [CrossRef] [Green Version]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
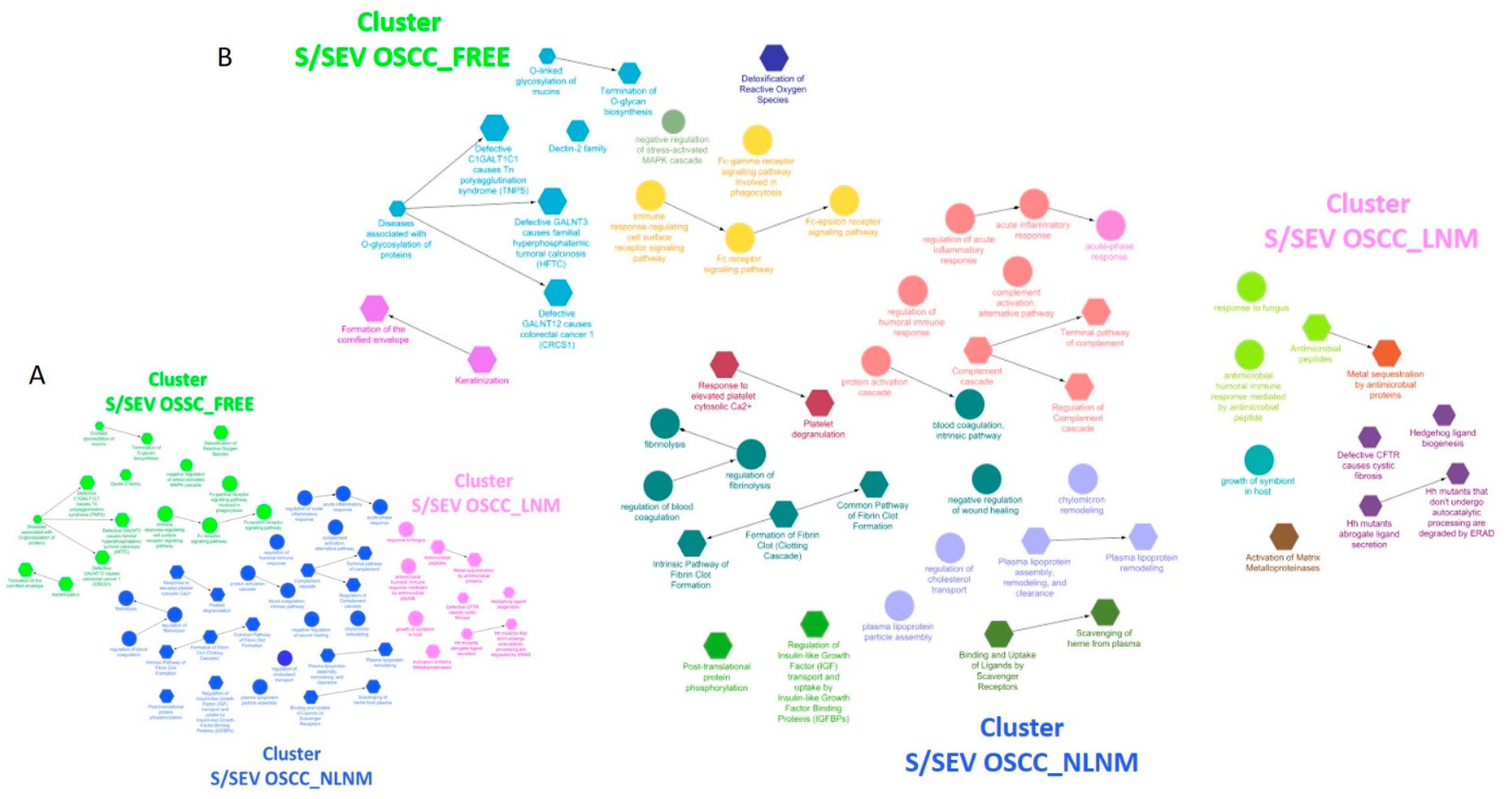{kind=link}
{kind=link}
| Anamnestic Data | OSCC_FREE Group (n = 5) | OSCC_NLNM Group (n = 6)) | OSCC_LNM Group (n = 12) | |
|---|---|---|---|---|
| Mean Age | 61.4 (±11.2) | 68.2 (±7.8) | 67.4 (±9.6) | |
| Gender | ||||
| Male | 3 (60%) | 1 (16.7%) | 8 (66.7%) | |
| Female | 2 (40%) | 5 (83.3%) | 4 (33.3%) | |
| Smoking Habit | ||||
| Non-smokers | 4 (80%) | 4 (66.7%) | 5 (41.7%) | |
| Smokers | 1 (20%) | 2 (33.3%) | 7 (58.3%) | |
| Alcohol Consumption | ||||
| Non-drinkers | 5 (100%) | 5 (83.3%) | 10 (83.3%) | |
| Former drinkers | 0 | 1 (16.7%) | 2 (16.7%) | |
| Group | Age | Sex | OSCC Site | Grading | TNM | Stage |
|---|---|---|---|---|---|---|
| OSCC_NLNM | F | 69 | Anterior 2/3 of tongue | G2-G3 | T1N0M0 | I |
| F | 58 | Anterior 2/3 of tongue | G2-G3 | T2N0M0 | II | |
| F | 70 | Anterior 2/3 of tongue | G3 | T2N0M0 | II | |
| F | 67 | Anterior 2/3 of tongue | G3 | T1N0M0 | I | |
| F | 83 | Gum | G2-G3 | T2N0M0 | II | |
| M | 62 | Anterior 2/3 of tongue | G2 | T2N0M0 | II | |
| OSCC_LNM | F | 78 | Gum | G2 | T2N1M0 | III |
| M | 63 | Retromolar area | G2-G3 | T4aN2bM0 | IVA | |
| M | 77 | Buccal mucosa | G2 | T3N2M0 | IVA | |
| F | 74 | Anterior 2/3 of tongue | G2 | T2N2aM0 | IVA | |
| M | 73 | Retromolar area | G2-G3 | T3N1M0 | III | |
| F | 66 | Gum | G2-G3 | T4aN2aM0 | IVA | |
| M | 48 | Retromolar area | G2 | T3N2cM0 | IVA | |
| F | 82 | Buccal mucosa | G2-G3 | T3N1M0 | III | |
| M | 54 | Anterior 2/3 of tongue | G3 | T2N1M0 | III | |
| M | 67 | Anterior floor of mouth | G3 | T4aN2M1 | IVC | |
| M | 66 | Anterior 2/3 of tongue | G3 | T2N2cM0 | IVA | |
| M | 61 | Retromolar area | G3 | T3N2bM1 | IVC |
| S/SEV OSCC_FREE | S/SEV OSCC_NLNM | S/SEV OSCC_LNM | |
|---|---|---|---|
| Number of identified proteins(DDA protein library) | 421 | ||
| Number of proteins quantified in SWATH-MS analysis | 365 | ||
| Number (and percentage) of quantified protein with CV ≤ 25% among technical replicates | 284 (78%) | 284 (78%) | 303 (83%) |
| S/SEV OSCC_FREE vs. S/SEV OSCC_NLNM | S/SEV OSCC_FREE vs. S/SEV OSCC_LNM | S/SEV OSCC_NLNM vs. S/SEV OSCC_LNM | |
|---|---|---|---|
| Number of modulated proteins | 235 | 157 | 189 |
| Number of up-regulated proteins | 144 | 68 | 70 |
| Number of down-regulated proteins | 91 | 89 | 119 |
| PROTEINS UP-REPRESENTED in S/SEV OSCC_FREE vs. S/SEV OSCC_NLNM and S/SEV OSCC_LNM | ||||
|---|---|---|---|---|
| S/SEV OSCC_FREE vs. S/SEV OSCC_NLNM | S/SEV OSCC_FREE vs. S/SEV OSCC_LNM | |||
| Gene Name | FC | BY p-Value | FC | BY p-Value |
| A2ML1 | 4.219 | 4.76 × 10−6 | 4.225 | 1.85 × 10−5 |
| ARHGDIB | 20.046 | 5.94 × 10−5 | 2.205 | 7.94 × 10−4 |
| B4GALT1 | 2.344 | 5.92 × 10−5 | 2.381 | 2.26 × 10−5 |
| BPIFB2 | 4.057 | 2.66 × 10−4 | 1.774 | 1.29 × 10−2 |
| CD59 | 2.406 | 7.49 × 10−6 | 1.713 | 1.32 × 10−3 |
| CEACAM1 | 1.539 | 1.01 × 10−2 | 2.306 | 3.65 × 10−3 |
| CSTB | 3.717 | 1.79 × 10−4 | 1.599 | 2.85 × 10−3 |
| DPP4 | 4.911 | 4.00 × 10−4 | 2.517 | 1.85 × 10−4 |
| DSG3 | 3.780 | 1.40 × 10−3 | 1.576 | 5.78 × 10−3 |
| ERO1A | 3.741 | 1.02 × 10−4 | 4.327 | 7.30 × 10−4 |
| EZR | 4.487 | 1.66 × 10−2 | 2.570 | 1.21 × 10−2 |
| FABP5 | 2.839 | 7.34 × 10−4 | 2.038 | 1.39 × 10−3 |
| FCGBP | 3.124 | 3.82 × 10−6 | 1.782 | 7.47 × 10−5 |
| GDI2 | 4.233 | 7.87 × 10−5 | 2.101 | 2.39 × 10−2 |
| GLRX | 3.345 | 8.31 × 10−4 | 1.514 | 1.55 × 10−2 |
| GSTP1 | 3.986 | 8.28 × 10−7 | 2.429 | 4.43 × 10−5 |
| IGHA1 | 1.682 | 3.01 × 10−4 | 1.600 | 1.35 × 10−2 |
| IGHA2 | 3.181 | 4.30 × 10−5 | 1.630 | 9.69 × 10−4 |
| IGHV1-2 | 3.206 | 1.63 × 10−4 | 1.645 | 2.01 × 10−3 |
| IGHV1-8 | 3.395 | 1.81 × 10−4 | 1.700 | 2.16 × 10−3 |
| IGHV3-15 | 2.833 | 2.06 × 10−5 | 3.462 | 4.43 × 10−5 |
| IGHV3-23 | 3.340 | 1.67 × 10−6 | 3.433 | 1.06 × 10−5 |
| IGHV3-7 | 2.175 | 8.91 × 10−5 | 2.790 | 9.93 × 10−5 |
| IGHV3-72 | 2.703 | 2.01 × 10−2 | 3.301 | 2.48 × 10−2 |
| IGHV3-9 | 2.832 | 3.01 × 10−4 | 2.500 | 2.67 × 10−4 |
| IGHV4-31 | 3.132 | 1.40 × 10−4 | 2.467 | 1.67 × 10−4 |
| IGHV5-51 | 3.084 | 1.49 × 10−3 | 2.591 | 4.67 × 10−3 |
| IGKC | 1.645 | 1.03 × 10−2 | 1.596 | 1.77 × 10−2 |
| IGKV1-13 | 16.563 | 3.57 × 10−4 | 7.833 | 8.21 × 10−4 |
| IGKV2-24 | 2.669 | 8.37 × 10−5 | 2.641 | 2.99 × 10−4 |
| IGKV2D-28 | 3.104 | 3.95 × 10−4 | 2.124 | 2.03 × 10−3 |
| IGKV3-7 | 1.867 | 3.35 × 10−4 | 1.808 | 4.58 × 10−4 |
| IGKV4-1 | 2.346 | 5.56 × 10−6 | 1.919 | 3.84 × 10−5 |
| IGLV1-47 | 2.801 | 2.63 × 10−5 | 1.718 | 1.28 × 10−2 |
| IL36A | 2.991 | 3.74 × 10−4 | 2.180 | 3.70 × 10−4 |
| KLK1 | 2.371 | 3.57 × 10−4 | 1.789 | 1.84 × 10−3 |
| KLK11 | 2.869 | 7.32 × 10−3 | 1.883 | 2.90 × 10−2 |
| KRT1 | 6.860 | 4.30 × 10−5 | 3.514 | 6.68 × 10−4 |
| KRT10 | 3.864 | 2.32 × 10−5 | 3.157 | 3.29 × 10−4 |
| KRT9 | 9.278 | 5.09 × 10−5 | 5.088 | 8.53 × 10−5 |
| LEG1 | 7.471 | 1.78 × 10−5 | 2.021 | 4.70 × 10−4 |
| MIF | 2.618 | 1.41 × 10−5 | 1.737 | 3.83 × 10−5 |
| MUC16 | 2.237 | 1.64 × 10−3 | 3.081 | 1.51 × 10−3 |
| MUC5B | 2.996 | 1.29 × 10−4 | 1.652 | 1.21 × 10−3 |
| MUC7 | 4.333 | 9.18 × 10−5 | 1.968 | 9.74 × 10−4 |
| PAM | 2.954 | 4.11 × 10−3 | 1.650 | 4.89 × 10−2 |
| PDCD6IP | 3.097 | 6.34 × 10−4 | 2.913 | 1.80 × 10−3 |
| PFN1 | 4.645 | 4.67 × 10−5 | 1.539 | 4.63 × 10−3 |
| PIGR | 3.157 | 5.56 × 10−6 | 1.712 | 7.35 × 10−5 |
| PRDX1 | 2.745 | 1.50 × 10−5 | 1.682 | 1.12 × 10−4 |
| PRDX6 | 2.902 | 1.52 × 10−3 | 1.995 | 5.45 × 10−3 |
| PROM1 | 3.190 | 7.64 × 10−4 | 1.613 | 8.08 × 10−3 |
| RAP1A | 2.686 | 4.76 × 10−4 | 1.848 | 2.03 × 10−3 |
| SERPINB13 | 4.951 | 9.62 × 10−3 | 4.588 | 1.82 × 10−3 |
| SERPINB3 | 3.054 | 8.31 × 10−4 | 1.618 | 8.09 × 10−3 |
| SERPINB5 | 3.617 | 2.09 × 10−5 | 2.574 | 1.36 × 10−4 |
| SPRR3 | 5.290 | 1.49 × 10−3 | 2.303 | 8.08 × 10−3 |
| TFF3 | 3.929 | 5.57 × 10−3 | 2.641 | 1.49 × 10−2 |
| TXN | 4.129 | 1.76 × 10−3 | 2.003 | 9.17 × 10−3 |
| YWHAZ | 3.510 | 1.75 × 10−5 | 1.592 | 1.28 × 10−4 |
| ZG16B | 11.069 | 2.42 × 10−6 | 1.548 | 2.01 × 10−4 |
| PROTEINS UP-REPRESENTED in S/SEV OSCC_NLNM vs. S/SEV OSCC_FREE and S/SEV OSCC_LNM | ||||
|---|---|---|---|---|
| S/SEV OSCC_NLNM vs. S/SEV OSCC_FREE | S/SEV OSCC_NLNM vs. S/SEV OSCC_LNM | |||
| Gene Name | FC | BY p-Value | FC | BY p-Value |
| A1BG | 2.798 | 2.10 × 10−2 | 1.592 | 2.75 × 10−4 |
| A2M | 6.270 | 1.74 × 10−5 | 2.210 | 6.61 × 10−4 |
| AHSG | 6.747 | 4.30 × 10−4 | 1.876 | 3.21 × 10−3 |
| ALB | 4.256 | 1.36 × 10−7 | 2.626 | 2.88 × 10−6 |
| AMBP | 14.291 | 4.88 × 10−4 | 2.456 | 5.62 × 10−3 |
| APCS | 16.885 | 4.30 × 10−5 | 5.393 | 1.17 × 10−4 |
| APOA1 | 8.000 | 5.56 × 10−6 | 1.668 | 1.28 × 10−4 |
| APOA2 | 7.601 | 2.06 × 10−5 | 1.646 | 7.63 × 10−4 |
| APOA4 | 12.525 | 6.10 × 10−3 | 3.344 | 3.29 × 10−2 |
| APOB | 4.968 | 1.66 × 10−5 | 4.149 | 9.25 × 10−5 |
| APOC1 | 33.965 | 1.13 × 10−5 | 4.655 | 5.97 × 10−5 |
| APOC3 | 5.447 | 2.16 × 10−2 | 5.776 | 6.82 × 10−4 |
| APOE | 5.997 | 2.06 × 10−4 | 2.907 | 6.02 × 10−4 |
| APOH | 5.406 | 8.31 × 10−4 | 7.510 | 7.17 × 10−4 |
| C1R | 8.320 | 3.84 × 10−3 | 9.948 | 2.81 × 10−3 |
| C1S | 21.978 | 5.91 × 10−5 | 6.646 | 4.85 × 10−4 |
| C3 | 8.758 | 1.43 × 10−5 | 2.998 | 9.25 × 10−5 |
| C4A | 7.062 | 1.10 × 10−2 | 13.261 | 4.62 × 10−3 |
| C4B | 13.800 | 1.63 × 10−4 | 14.008 | 4.44 × 10−4 |
| C4BPA | 47.980 | 5.59 × 10−06 | 13.112 | 2.01 × 10−5 |
| C4BPB | 22.217 | 6.19 × 10−5 | 6.869 | 1.60 × 10−3 |
| C5 | 8.230 | 3.04 × 10−3 | 1.962 | 3.29 × 10−2 |
| C6 | 35.474 | 7.87 × 10−5 | 3.882 | 3.77 × 10−2 |
| C7 | 7.835 | 2.23 × 10−4 | 2.299 | 5.19 × 10−3 |
| C8A | 20.753 | 2.79 × 10−4 | 2.112 | 3.70 × 10−3 |
| C9 | 7.681 | 2.04 × 10−3 | 2.666 | 3.39 × 10−4 |
| CD5L | 6.049 | 4.71 × 10−4 | 1.975 | 1.52 × 10−3 |
| CFB | 4.768 | 8.31 × 10−4 | 6.263 | 1.15 × 10−3 |
| CFH | 4.943 | 4.27 × 10−3 | 3.210 | 4.59 × 10−4 |
| CLU | 1.810 | 7.34 × 10−4 | 2.291 | 6.55 × 10−4 |
| ECM1 | 1.876 | 1.08 × 10−4 | 1.685 | 1.20 × 10−3 |
| F2 | 20.900 | 4.76 × 10−6 | 9.344 | 3.13 × 10−5 |
| FGA | 50.711 | 9.10 × 10−6 | 14.394 | 2.08 × 10−5 |
| FGB | 38.017 | 8.28 × 10−7 | 7.917 | 2.88 × 10−6 |
| FGG | 29.326 | 5.65 × 10−6 | 10.431 | 2.08 × 10−5 |
| FN1 | 14.280 | 2.32 × 10−5 | 5.879 | 2.08 × 10−5 |
| GC | 5.916 | 3.35 × 10−4 | 2.567 | 2.69 × 10−3 |
| GLUL | 2.553 | 5.29 × 10−4 | 5.317 | 6.49 × 10−4 |
| HABP2 | 18.099 | 6.53 × 10−5 | 12.473 | 2.17 × 10−4 |
| HBA1 | 4.942 | 3.79 × 10−6 | 1.524 | 1.50 × 10−4 |
| HBB | 4.413 | 5.07 × 10−7 | 1.850 | 2.08 × 10−5 |
| HBD | 2.964 | 1.33 × 10−3 | 2.189 | 6.93 × 10−3 |
| HP | 9.067 | 5.07 × 10−7 | 2.988 | 2.88 × 10−6 |
| HPR | 5.949 | 4.76 × 10−4 | 1.875 | 2.92 × 10−3 |
| HPX | 4.703 | 5.01 × 10−5 | 2.413 | 5.15 × 10−5 |
| HRG | 17.313 | 5.56 × 10−6 | 6.654 | 9.27 × 10−4 |
| IGHG1 | 5.954 | 1.47 × 10−6 | 1.899 | 2.08 × 10−5 |
| IGHG2 | 9.230 | 8.15 × 10−5 | 3.302 | 3.05 × 10−4 |
| IGHG3 | 15.347 | 2.31 × 10−4 | 7.226 | 5.95 × 10−5 |
| ITIH1 | 6.718 | 2.00 × 10−5 | 2.244 | 6.62 × 10−4 |
| ITIH2 | 18.967 | 3.54 × 10−5 | 5.154 | 9.25 × 10−5 |
| ITIH4 | 8.286 | 2.42 × 10−6 | 4.882 | 7.04 × 10−6 |
| KLK13 | 1.811 | 1.25 × 10−3 | 1.578 | 1.33 × 10−4 |
| KLKB1 | 26.394 | 5.30 × 10−4 | 9.351 | 3.13 × 10−3 |
| KNG1 | 6.160 | 1.43 × 10−5 | 5.107 | 6.15 × 10−3 |
| LPA | 26.553 | 5.07 × 10−7 | 5.994 | 3.38 × 10−5 |
| ORM1 | 27.811 | 5.65 × 10−6 | 9.577 | 9.54 × 10−6 |
| PLG | 26.842 | 4.76 × 10−6 | 10.061 | 3.91 × 10−5 |
| PROS1 | 11.100 | 7.87 × 10−5 | 1.581 | 1.22 × 10−3 |
| SERPINA3 | 15.356 | 2.09 × 10−5 | 2.224 | 1.33 × 10−3 |
| SERPINA4 | 4.895 | 2.10 × 10−2 | 8.646 | 4.44 × 10−4 |
| SERPINC1 | 12.622 | 2.00 × 10−5 | 8.810 | 3.38 × 10−5 |
| SERPIND1 | 3.400 | 1.14 × 10−3 | 2.006 | 3.67 × 10−3 |
| SERPINF2 | 25.834 | 1.14 × 10−3 | 10.267 | 2.06 × 10−3 |
| TF | 5.764 | 2.53 × 10−3 | 1.991 | 2.57 × 10−2 |
| VTN | 40.677 | 5.07 × 10−7 | 6.165 | 4.22 × 10−5 |
| PROTEINS UP-REPRESENTED in S/SEV OSCC_NLNM vs. S/SEV OSCC_FREE and S/SEV OSCC_LNM | ||||
|---|---|---|---|---|
| S/SEV OSCC_NLNM vs. S/SEV OSCC_FREE | S/SEV OSCC_NLNM vs. S/SEV OSCC_LNM | |||
| Gene Name | FC | BY p-Value | FC | BY p-Value |
| ACTN1 | 1.641 | 3.62 × 10−3 | 2.970 | 1.25 × 10−3 |
| ACTR3 | 1.570 | 6.66 × 10−4 | 3.768 | 4.85 × 10−4 |
| ANXA3 | 1.951 | 8.08 × 10−3 | 2.258 | 4.60 × 10−3 |
| AZU1 | 13.474 | 2.60 × 10−3 | 2.253 | 1.34 × 10−2 |
| CA6 | 1.869 | 4.86 × 10−3 | 3.846 | 6.77 × 10−4 |
| CST5 | 1.509 | 6.38 × 10−3 | 2.083 | 1.01 × 10−3 |
| ELANE | 14.603 | 7.16 × 10−5 | 2.648 | 1.25 × 10−4 |
| FCN1 | 2.756 | 1.09 × 10−4 | 4.392 | 4.63 × 10−4 |
| FTL | 3.868 | 6.66 × 10−4 | 3.202 | 4.39 × 10−2 |
| GAPDH | 2.107 | 1.33 × 10−2 | 2.718 | 6.49 × 10−3 |
| H2BC21 | 9.628 | 2.30 × 10−3 | 2.076 | 4.03 × 10−3 |
| KLK10 | 1.695 | 2.27 × 10−2 | 1.944 | 5.40 × 10−3 |
| KLK14 | 2.870 | 3.90 × 10−4 | 6.661 | 4.03 × 10−3 |
| LCN2 | 1.614 | 1.09 × 10−4 | 2.041 | 9.99 × 10−4 |
| LTA4H | 1.553 | 1.66 × 10−2 | 5.109 | 1.42 × 10−2 |
| LTF | 9.363 | 1.39 × 10−5 | 1.659 | 5.97 × 10−5 |
| MMP9 | 1.698 | 2.74 × 10−2 | 2.376 | 3.90 × 10−2 |
| MPO | 10.720 | 1.08 × 10−6 | 2.478 | 2.88 × 10−6 |
| MYL6 | 2.264 | 4.11 × 10−3 | 1.654 | 2.91 × 10−3 |
| NUCB2 | 3.670 | 2.25 × 10−3 | 19.150 | 7.94 × 10−4 |
| PGLYRP1 | 1.659 | 3.29 × 10−4 | 2.814 | 9.25 × 10−5 |
| PI3 | 19.846 | 8.17 × 10−5 | 2.694 | 3.12 × 10−3 |
| PRB1 | 89.610 | 1.74 × 10−3 | 18.859 | 3.00 × 10−3 |
| PRR27 | 4.359 | 1.67 × 10−4 | 5.401 | 1.50 × 10−4 |
| PRTN3 | 2.127 | 2.78 × 10−4 | 4.468 | 2.08 × 10−5 |
| PSMA5 | 3.158 | 1.67 × 10−2 | 3.295 | 2.32 × 10−2 |
| PSMB2 | 5.007 | 4.67 × 10−2 | 1.730 | 3.89 × 10−2 |
| RETN | 2.731 | 7.47 × 10−5 | 1.676 | 9.25 × 10−5 |
| RNASE3 | 16.775 | 7.30 × 10−4 | 4.570 | 4.44 × 10−4 |
| S100A12 | 37.287 | 1.44 × 10−4 | 4.506 | 1.74 × 10−3 |
| S100A9 | 3.173 | 4.43 × 10−5 | 4.128 | 2.74 × 10−5 |
| SCGB3A1 | 2.108 | 1.03 × 10−4 | 3.282 | 6.47 × 10−5 |
| SMR3B | 3.503 | 6.35 × 10−3 | 15.324 | 2.49 × 10−3 |
| TIMP1 | 2.154 | 7.47 × 10−5 | 5.408 | 3.40 × 10−6 |
| TKT | 2.904 | 1.46 × 10−2 | 7.511 | 5.17 × 10−3 |
| TMEM198 | 80.329 | 9.30 × 10−5 | 33.058 | 9.25 × 10−5 |
| VCP | 2.052 | 8.87 × 10−3 | 1.922 | 8.65 × 10−3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fontana, S.; Mauceri, R.; Novara, M.E.; Alessandro, R.; Campisi, G. Protein Cargo of Salivary Small Extracellular Vesicles as Potential Functional Signature of Oral Squamous Cell Carcinoma. Int. J. Mol. Sci. 2021, 22, 11160. https://doi.org/10.3390/ijms222011160
Fontana S, Mauceri R, Novara ME, Alessandro R, Campisi G. Protein Cargo of Salivary Small Extracellular Vesicles as Potential Functional Signature of Oral Squamous Cell Carcinoma. International Journal of Molecular Sciences. 2021; 22(20):11160. https://doi.org/10.3390/ijms222011160
Chicago/Turabian StyleFontana, Simona, Rodolfo Mauceri, Maria Eugenia Novara, Riccardo Alessandro, and Giuseppina Campisi. 2021. "Protein Cargo of Salivary Small Extracellular Vesicles as Potential Functional Signature of Oral Squamous Cell Carcinoma" International Journal of Molecular Sciences 22, no. 20: 11160. https://doi.org/10.3390/ijms222011160
APA StyleFontana, S., Mauceri, R., Novara, M. E., Alessandro, R., & Campisi, G. (2021). Protein Cargo of Salivary Small Extracellular Vesicles as Potential Functional Signature of Oral Squamous Cell Carcinoma. International Journal of Molecular Sciences, 22(20), 11160. https://doi.org/10.3390/ijms222011160