
Developmental Exposure to Low Concentrations of Methylmercury Causes Increase in Anxiety-Related Behaviour and Locomotor Impairments in Zebrafish



Abstract
:1. Introduction
2. Results
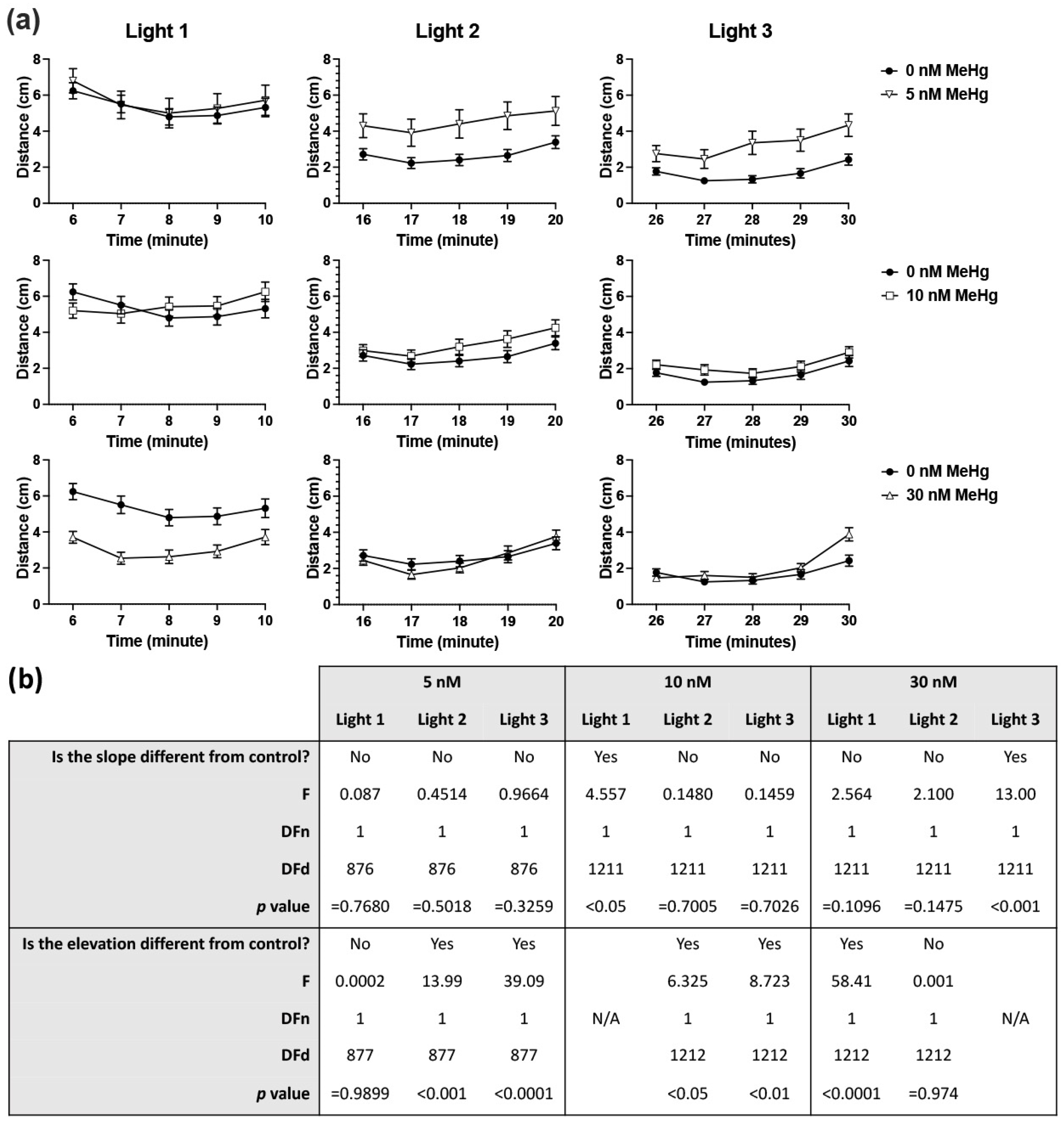
2.1. Larval Light/Dark Locomotor Activity Test
2.2. Adult Behavioural Testing
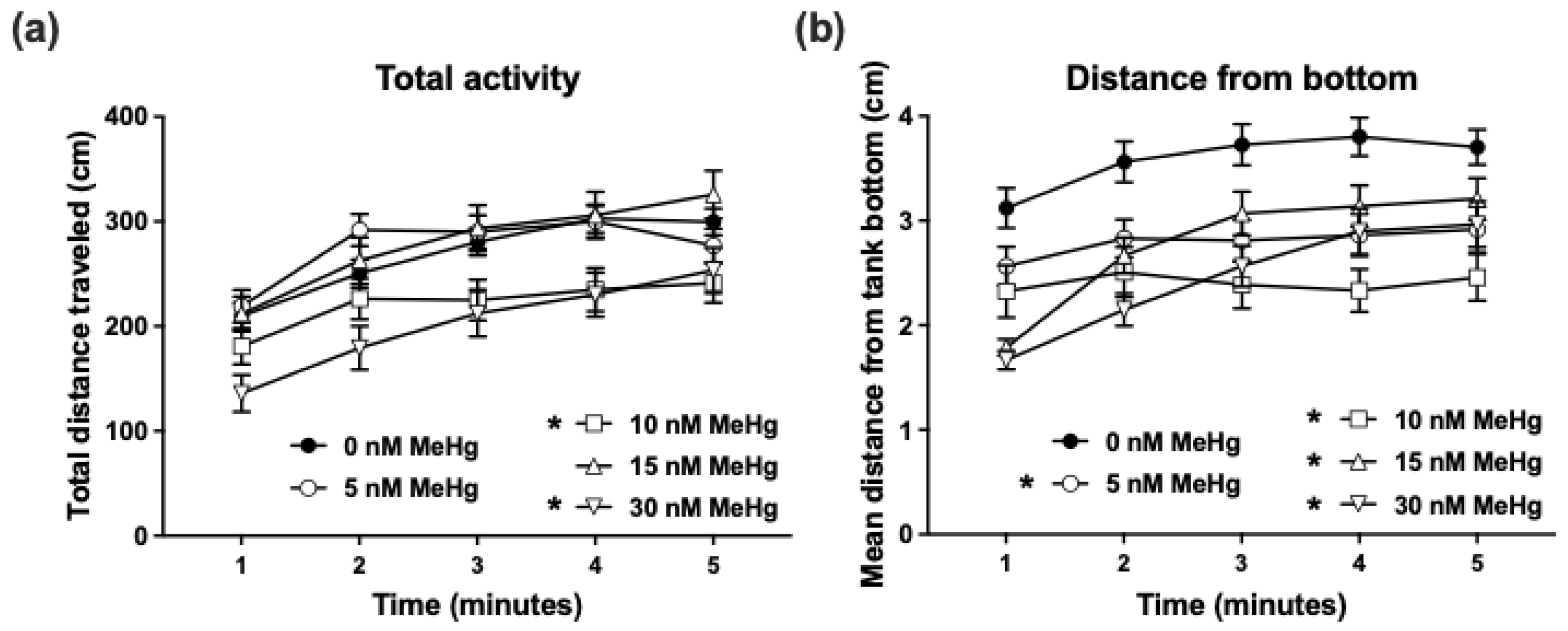
2.2.1. Adult Novel Tank Dive Test
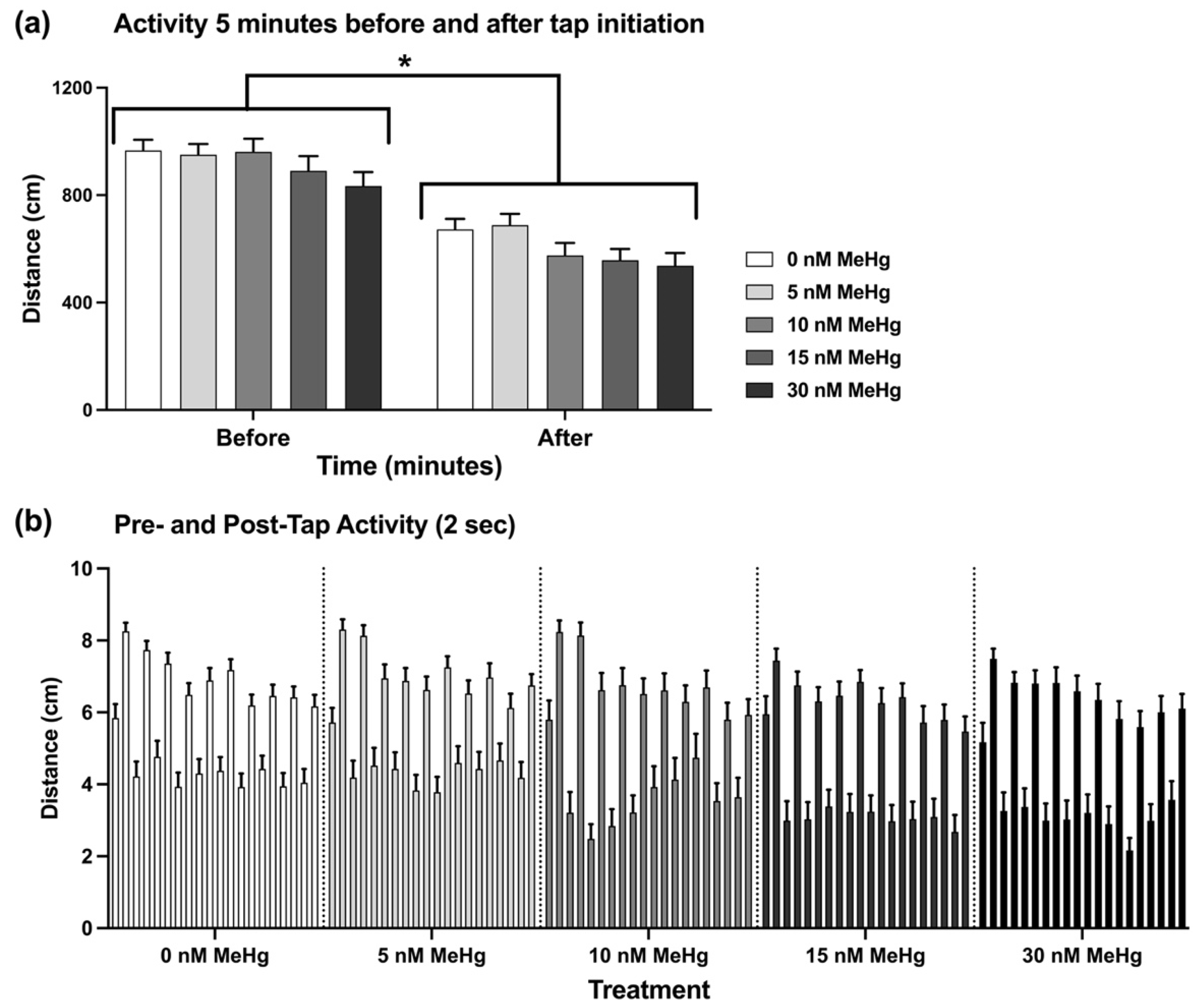
2.2.2. Adult Startle Tap Test
2.3. Larval and Adult Gene Expression
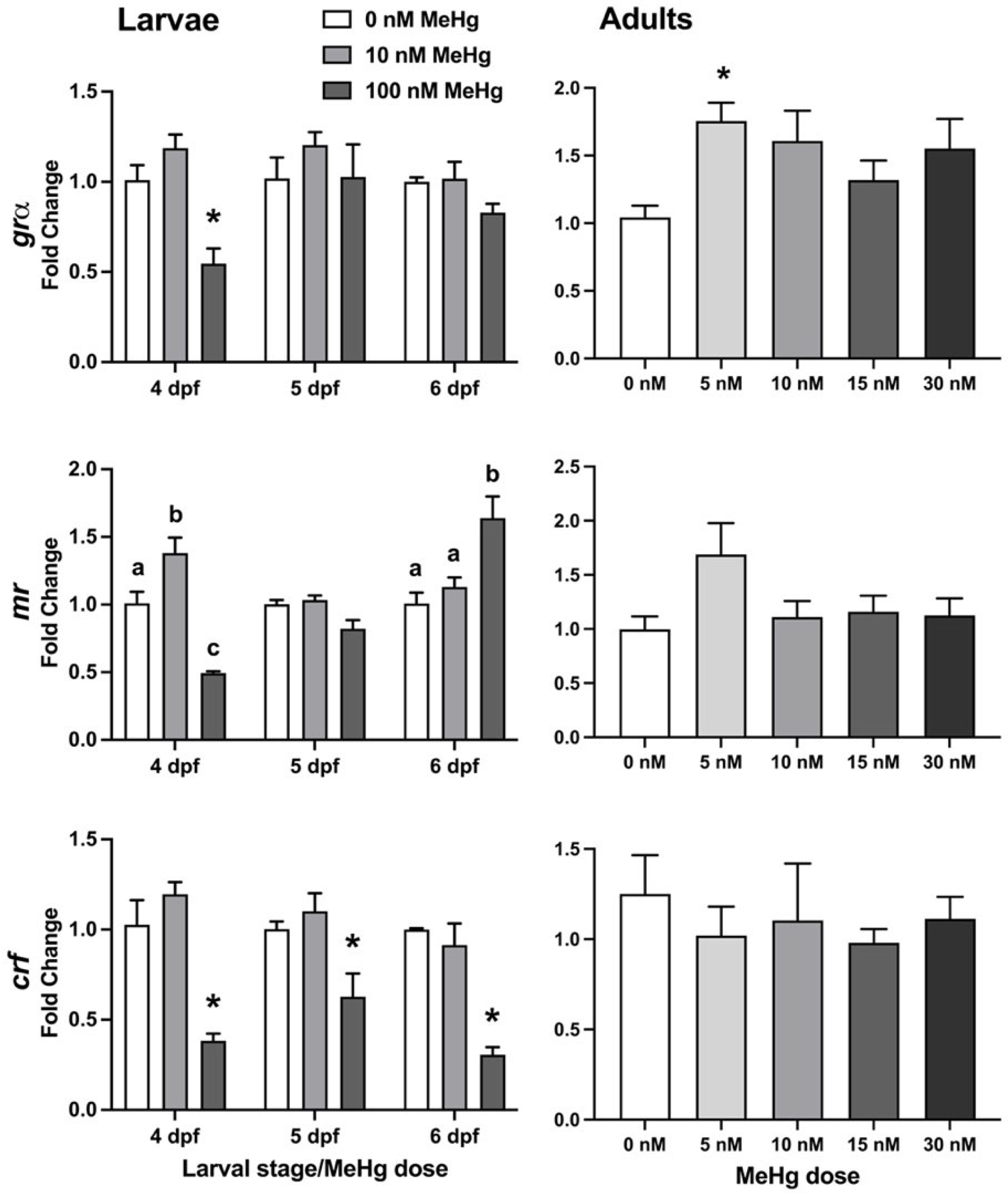
2.3.1. HPI-Axis Gene Expression
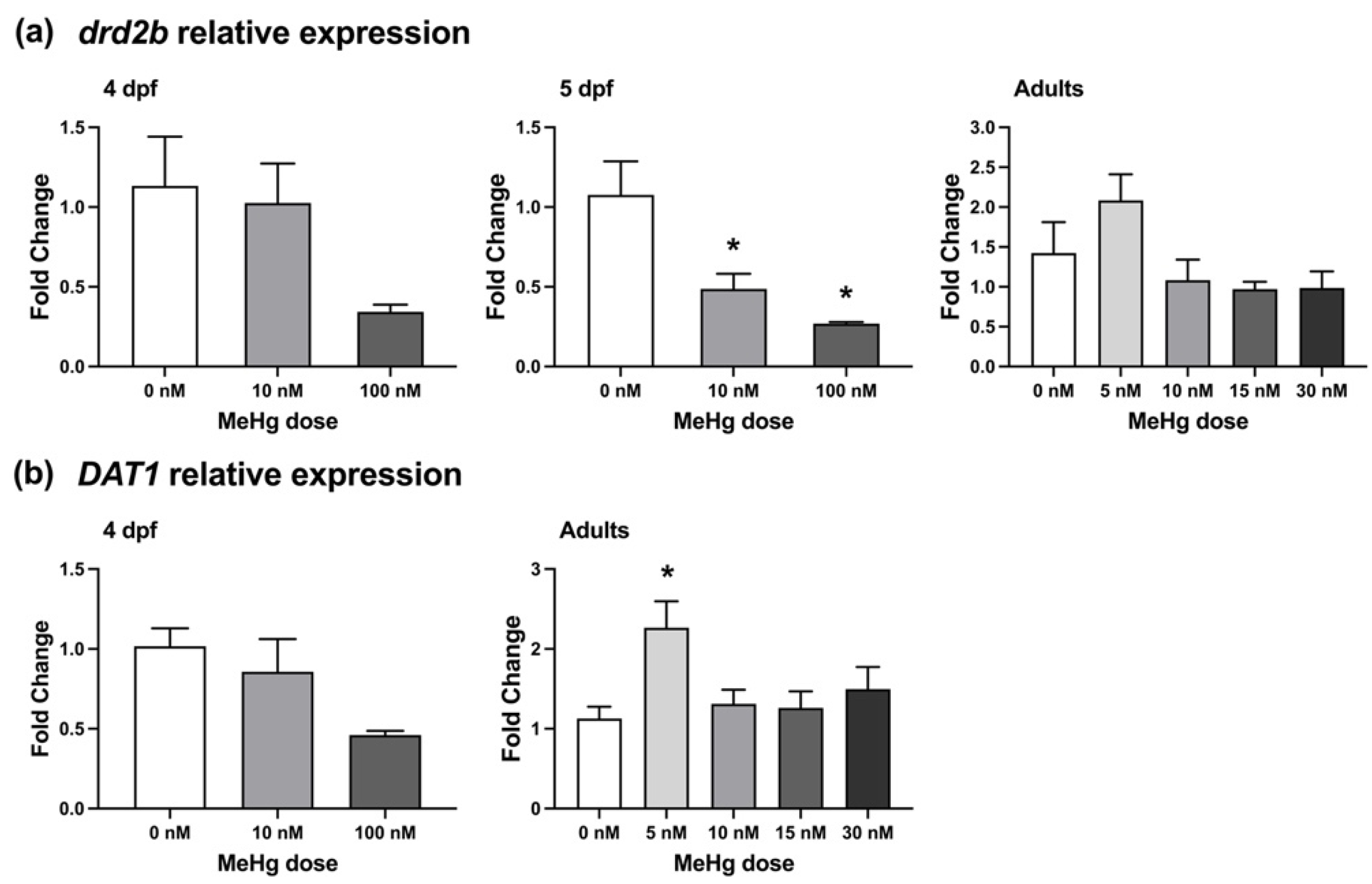
2.3.2. Dopamine-Related Gene Expression
3. Discussion
4. Materials and Methods
4.1. Fish Husbandry and Breeding
4.2. Developmental Exposure to Methylmercury
4.3. Larval Light/Dark Locomotor Activity Test
4.4. Adult Behavioural Test Battery
4.4.1. Novel Tank Dive Test
4.4.2. Adult Startle Tap Test
4.5. Whole Larvae and Adult Brain Sampling, RNA Extraction and Real-Time RT qPCR
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kampalath, R.A.; Jay, J.A. Sources of mercury exposure to children in low- and middle-income countries. J. Health Pollut. 2015, 5, 33–51. [Google Scholar] [CrossRef] [PubMed]
- Tatsuta, N.; Nakai, K.; Sakamoto, M.; Murata, K.; Satoh, H. Methylmercury exposure and developmental outcomes in Tohoku study of child development at 18 months of age. Toxics 2018, 6, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grandjean, P.; Weihe, P.; White, R.F.; Debes, F.; Araki, S.; Yokoyama, K.; Murata, K.; Sorensen, N.; Dahl, R.; Jorgensen, P.J. Cognitive deficit in 7-year-old children with prenatal exposure to methylmercury. Neurotoxicol. Teratol. 1997, 19, 417–428. [Google Scholar] [CrossRef]
- Ceccatelli, S.; Bose, R.; Edoff, K.; Onishchenko, N.; Spulber, S. Long-lasting neurotoxic effects of exposure to methylmercury during development. J. Intern. Med. 2013, 273, 490–497. [Google Scholar] [CrossRef]
- Grandjean, P.; Landrigan, P.J. Developmental neurotoxicity of industrial chemicals. Lancet 2006, 368, 2167–2178. [Google Scholar] [CrossRef]
- Karagas, M.R.; Choi, A.L.; Oken, E.; Horvat, M.; Schoeny, R.; Kamai, E.; Cowell, W.; Grandjean, P.; Korrick, S. Evidence on the human health effects of low-level methylmercury exposure. Environ. Health Perspect. 2012, 120, 799–806. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, M.C.; Burke, T.A.; Navas-Acien, A.; Breysse, P.N.; McGready, J.; Fox, M.A. Global methylmercury exposure from seafood consumption and risk of developmental neurotoxicity: A systematic review. Bull. World Health Organ. 2014, 92, 254–269F. [Google Scholar] [CrossRef]
- Grandjean, P.; Weihe, P.; White, R.F.; Debes, F. Cognitive performance of children prenatally exposed to “safe” levels of methylmercury. Environ. Res. 1998, 77, 165–172. [Google Scholar] [CrossRef]
- Debes, F.; Weihe, P.; Grandjean, P. Cognitive deficits at age 22 years associated with prenatal exposure to methylmercury. Cortex 2016, 74, 358–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kjellström, T.; Kennedy, P.; Wallis, S.; Stewart, A.; Friberg, L.; Lind, B.; Wutherspoon, T.; Mantell, C. Physical and Mental Development of Children with Prenatal Exposure to Mercury from Fish. Stage 2. Interviews and Psychological Tests at Age 6; National Swedish Environmental Protection Board: Stockholm, Sweden, 1989; ISBN 91-620-3642-4. [Google Scholar]
- Patel, N.B.; Xu, Y.; McCandless, L.C.; Chen, A.; Yolton, K.; Braun, J.; Jones, R.L.; Dietrich, K.N.; Lanphear, B.P. Very low-level prenatal mercury exposure and behaviors in children: The HOME Study. Environ. Health 2019, 18, 4. [Google Scholar] [CrossRef] [Green Version]
- Sagiv, S.K.; Thurston, S.W.; Bellinger, D.C.; Amarasiriwardena, C.; Korrick, S.A. Prenatal exposure to mercury and fish consumption during pregnancy and attention-deficit/hyperactivity disorder-related behavior in children. Arch. Pediat. Adol. Med. 2012, 166, 1123–1131. [Google Scholar] [CrossRef] [Green Version]
- Garrecht, M.; Austin, D.W. The plausibility of a role for mercury in the etiology of autism: A cellular perspective. Toxicol. Environ. Chem. 2011, 93, 1251–1273. [Google Scholar] [CrossRef] [Green Version]
- Geier, D.A.; King, P.G.; Sykes, L.K.; Geier, M.R. A comprehensive review of mercury provoked autism. Indian J. Med. Res. 2008, 128, 383–411. [Google Scholar] [PubMed]
- Gorini, F.; Muratori, F.; Morales, M.A. The role of heavy metal pollution in neurobehavioral disorders: A focus on autism. Rev. J. Autism Dev. Disord. 2014, 1, 354–372. [Google Scholar] [CrossRef] [Green Version]
- Grandjean, P.; Cordier, S.; Kjellström, T.; Weihe, P.; Budtz-Jorgensen, E. Dynamics of Mercury Pollution on Regional and Global Scales: Atmospheric Processes and Human Exposures around the World; Springer Science & Business Media: New York, NY, USA, 2005; p. 744. [Google Scholar]
- Onishchenko, N.; Karpova, N.; Sabri, F.; Castren, E.; Ceccatelli, S. Long-lasting depression-like behavior and epigenetic changes of BDNF gene expression induced by perinatal exposure to methylmercury. J. Neurochem. 2008, 106, 1378–1387. [Google Scholar] [CrossRef] [PubMed]
- Onishchenko, N.; Tamm, C.; Vahter, M.; Hokfelt, T.; Johnson, J.A.; Johnson, D.A.; Ceccatelli, S. Developmental exposure to methylmercury alters learning and induces depression-like behavior in male mice. Toxicol. Sci. 2007, 97, 428–437. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Fujimura, M.; Bo, D. Assessing pre/post-weaning neurobehavioral development for perinatal exposure to low doses of methylmercury. J. Environ. Sci. 2015, 38, 36–41. [Google Scholar] [CrossRef]
- Sakamoto, M.; Kakita, A.; Wakabayashi, K.; Takahashi, H.; Nakano, A.; Akagi, H. Evaluation of changes in methylmercury accumulation in the developing rat brain and its effects: A study with consecutive and moderate dose exposure throughout gestation and lactation periods. Brain Res. 2002, 949, 51–59. [Google Scholar] [CrossRef]
- Widholm, J.J.; Villareal, S.; Seegal, R.F.; Schantz, S.L. Spatial alternation deficits following developmental exposure to Aroclor 1254 and/or methylmercury in rats. Toxicol. Sci. 2004, 82, 577–589. [Google Scholar] [CrossRef] [Green Version]
- Doré, F.Y.; Goulet, S.; Gallagher, A.; Harvey, P.O.; Cantin, J.F.; D’Aigle, T.; Mirault, M.E. Neurobehavioral changes in mice treated with methylmercury at two different stages of fetal development. Neurotoxicol. Teratol. 2001, 23, 463–472. [Google Scholar] [CrossRef]
- Yoshida, M.; Shimizu, N.; Suzuki, M.; Watanabe, C.; Satoh, M.; Mori, K.; Yasutake, A. Emergence of delayed methylmercury toxicity after perinatal exposure in metallothionein-null and wild-type C57BL mice. Environ. Health Perspect. 2008, 116, 746–751. [Google Scholar] [CrossRef] [Green Version]
- Samson, J.C.; Goodridge, R.; Olobatuyi, F.; Weis, J.S. Delayed effects of embryonic exposure of zebrafish (Danio rerio) to methylmercury (MeHg). Aquat. Toxicol. 2001, 51, 369–376. [Google Scholar] [CrossRef]
- Selderslaghs, I.W.; Hooyberghs, J.; Blust, R.; Witters, H.E. Assessment of the developmental neurotoxicity of compounds by measuring locomotor activity in zebrafish embryos and larvae. Neurotoxicol. Teratol. 2013, 37, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.E.; Carvan, M.J., 3rd; Dellinger, J.A.; Ghorai, J.K.; White, D.B.; Williams, F.E.; Weber, D.N. Developmental selenomethionine and methylmercury exposures affect zebrafish learning. Neurotoxicol. Teratol. 2010, 32, 246–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Lamb, C.; Smith, M.; Schaefer, L.; Carvan, M.J.; Weber, D.N. Developmental methylmercury exposure affects avoidance learning outcomes in adult zebrafish. J. Toxicol. Environ. Health A 2012, 4, 85–91. [Google Scholar]
- Weber, D.N.; Connaughton, V.P.; Dellinger, J.A.; Klemer, D.; Udvadia, A.; Carvan, M.J. Selenomethionine reduces visual deficits due to developmental methylmercury exposures. Physiol. Behav. 2008, 93, 250–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aaseth, J.; Wallace, D.R.; Vejrup, K.; Alexander, J. Methylmercury and developmental neurotoxicity: A global concern. Curr. Opin. Toxicol. 2020, 19, 80–87. [Google Scholar] [CrossRef]
- Antunes dos Santos, A.; Appel Hort, M.; Culbreth, M.; Lopez-Granero, C.; Farina, M.; Rocha, J.B.; Aschner, M. Methylmercury and brain development: A review of recent literature. J. Trace Elem. Med. Biol. 2016, 38, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Farina, M.; Aschner, M. Glutathione antioxidant system and methylmercury-induced neurotoxicity: An intriguing interplay. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 129285. [Google Scholar] [CrossRef]
- Dare, E.; Fetissov, S.; Hokfelt, T.; Hall, H.; Ogren, S.O.; Ceccatelli, S. Effects of prenatal exposure to methylmercury on dopamine-mediated locomotor activity and dopamine D2 receptor binding. Naunyn Schmiedebergs Arch. Pharmacol. 2003, 367, 500–508. [Google Scholar] [CrossRef]
- Spulber, S.; Raciti, M.; Dulko-Smith, B.; Lupu, D.; Ruegg, J.; Nam, K.; Ceccatelli, S. Methylmercury interferes with glucocorticoid receptor: Potential role in the mediation of developmental neurotoxicity. Toxicol. Appl. Pharm. 2018, 354, 94–100. [Google Scholar] [CrossRef]
- Glazer, L.; Hawkey, A.B.; Wells, C.N.; Drastal, M.; Odamah, K.A.; Behl, M.; Levin, E.D. Developmental exposure to low concentrations of organophosphate flame retardants causes life-long behavioral alterations in zebrafish. Toxicol. Sci. 2018, 165, 487–498. [Google Scholar] [CrossRef] [Green Version]
- Glazer, L.; Wells, C.N.; Drastal, M.; Odamah, K.A.; Galat, R.E.; Behl, M.; Levin, E.D. Developmental exposure to low concentrations of two brominated flame retardants, BDE-47 and BDE-99, causes life-long behavioral alterations in zebrafish. Neurotoxicology 2018, 66, 221–232. [Google Scholar] [CrossRef]
- Gimenez-Llort, L.; Ahlbom, E.; Dare, E.; Vahter, M.; Ogren, S.; Ceccatelli, S. Prenatal exposure to methylmercury changes dopamine-modulated motor activity during early ontogeny: Age and gender-dependent effects. Environ. Toxicol. Pharmacol. 2001, 9, 61–70. [Google Scholar] [CrossRef]
- Zimmer, B.; Schildknecht, S.; Kuegler, P.B.; Tanavde, V.; Kadereit, S.; Leist, M. Sensitivity of dopaminergic neuron differentiation from stem cells to chronic low-dose methylmercury exposure. Toxicol. Sci. 2011, 121, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Best, C.; Kurrasch, D.M.; Vijayan, M.M. Maternal cortisol stimulates neurogenesis and affects larval behaviour in zebrafish. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Lin, J.; Zhu, Y.; Liu, X.; Zhang, Y.; Ji, Y.; Yang, X.; Zhang, Y.; Guo, N.; Li, Q. Anxiety-related behavioral responses of pentylenetetrazole-treated zebrafish larvae to light-dark transitions. Pharmacol. Biochem. Behav. 2016, 145, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Padilla, S.; Hunter, D.L.; Padnos, B.; Frady, S.; MacPhail, R.C. Assessing locomotor activity in larval zebrafish: Influence of extrinsic and intrinsic variables. Neurotoxicol. Teratol. 2011, 33, 624–630. [Google Scholar] [CrossRef]
- Noyes, P.D.; Haggard, D.E.; Gonnerman, G.D.; Tanguay, R.L. Advanced morphological-behavioral test platform reveals neurodevelopmental defects in embryonic zebrafish exposed to comprehensive suite of halogenated and organophosphate flame retardants. Toxicol. Sci. 2015, 145, 177–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarema, K.A.; Hunter, D.L.; Shaffer, R.M.; Behl, M.; Padilla, S. Acute and developmental behavioral effects of flame retardants and related chemicals in zebrafish. Neurotoxicol. Teratol. 2015, 52, 194–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.S.Y.; Noble, S.; Godoy, R.; Ekker, M.; Chan, H.M. Delayed effects of methylmercury on the mitochondria of dopaminergic neurons and developmental toxicity in zebrafish larvae (Danio rerio). Aquat. Toxicol. 2016, 175, 73–80. [Google Scholar] [CrossRef]
- Bencan, Z.; Sledge, D.; Levin, E.D. Buspirone, chlordiazepoxide and diazepam effects in a zebrafish model of anxiety. Pharmacol. Biochem. Behav. 2009, 94, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Cachat, J.; Stewart, A.; Grossman, L.; Gaikwad, S.; Kadri, F.; Chung, K.M.; Wu, N.; Wong, K.; Roy, S.; Suciu, C.; et al. Measuring behavioral and endocrine responses to novelty stress in adult zebrafish. Nat. Protoc. 2010, 5, 1786–1799. [Google Scholar] [CrossRef]
- Egan, R.J.; Bergner, C.L.; Hart, P.C.; Cachat, J.M.; Canavello, P.R.; Elegante, M.F.; Elkhayat, S.I.; Bartels, B.K.; Tien, A.K.; Tien, D.H.; et al. Understanding behavioral and physiological phenotypes of stress and anxiety in zebrafish. Behav. Brain Res. 2009, 205, 38–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maximino, C.; de Brito, T.M.; Batista, A.W.D.; Herculano, A.M.; Morato, S.; Gouveia, A. Measuring anxiety in zebrafish: A critical review. Behav. Brain Res. 2010, 214, 157–171. [Google Scholar] [CrossRef]
- Levin, E.D.; Bencan, Z.; Cerutti, D.T. Anxiolytic effects of nicotine in zebrafish. Physiol. Behav. 2007, 90, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Baiamonte, M.; Parker, M.O.; Vinson, G.P.; Brennan, C.H. Sustained effects of developmental exposure to ethanol on zebrafish anxiety-like behaviour. PLoS ONE 2016, 11, e0148425. [Google Scholar] [CrossRef]
- Nesan, D.; Vijayan, M.M. Role of glucocorticoid in developmental programming: Evidence from zebrafish. Gen. Comp. Endocrinol. 2013, 181, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Wendelaar Bonga, S.E. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef]
- Sireeni, J.; Bakker, N.; Jaikumar, G.; Obdam, D.; Slabbekoorn, H.; Tudorache, C.; Schaaf, M. Profound effects of glucocorticoid resistance on anxiety-related behavior in zebrafish adults but not in larvae. Gen. Comp. Endocrinol. 2020, 292, 113461. [Google Scholar] [CrossRef]
- Kelly, M.A.; Rubinstein, M.; Phillips, T.J.; Lessov, C.N.; Burkhart-Kasch, S.; Zhang, G.; Bunzow, J.R.; Fang, Y.; Gerhardt, G.A.; Grandy, D.K. Locomotor activity in D2 dopamine receptor-deficient mice is determined by gene dosage, genetic background, and developmental adaptations. J. Neurosci. 1998, 18, 3470–3479. [Google Scholar] [CrossRef] [Green Version]
- Cabib, S.; Puglisi-Allegra, S. The mesoaccumbens dopamine in coping with stress. Neurosci. Biobehav. Rev. 2012, 36, 79–89. [Google Scholar] [CrossRef]
- Thornqvist, P.O.; McCarrick, S.; Ericsson, M.; Roman, E.; Winberg, S. Bold zebrafish (Danio rerio) express higher levels of delta opioid and dopamine D2 receptors in the brain compared to shy fish. Behav. Brain Res. 2019, 359, 927–934. [Google Scholar] [CrossRef]
- Sullivan, R.M.; Dufresne, M.M. Mesocortical dopamine and HPA axis regulation: Role of laterality and early environment. Brain Res. 2006, 1076, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Reed, M.N.; Newland, M.C. Gestational methylmercury exposure selectively increases the sensitivity of operant behavior to cocaine. Behav. Neurosci. 2009, 123, 408–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Wang, C.; Gao, X.; Zhu, J.; Wang, L.; Cao, S.; Wu, Q.; Qiao, S.; Zhang, Z.; Li, L. Comparative effects of mercury chloride and methylmercury exposure on early neurodevelopment in zebrafish larvae. RSC Adv. 2019, 9, 10766–10775. [Google Scholar] [CrossRef] [Green Version]
- Bjorklund, O.; Kahlstrom, J.; Salmi, P.; Ogren, S.O.; Vahter, M.; Chen, J.F.; Fredholm, B.B.; Dare, E. The effects of methylmercury on motor activity are sex- and age-dependent, and modulated by genetic deletion of adenosine receptors and caffeine administration. Toxicology 2007, 241, 119–133. [Google Scholar] [CrossRef]
- Newland, M.C.; Reed, M.N.; Rasmussen, E. A hypothesis about how early developmental methylmercury exposure disrupts behavior in adulthood. Behav. Process 2015, 114, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farina, M.; Rocha, J.B.T.; Aschner, M. Mechanisms of methylmercury-induced neurotoxicity: Evidence from experimental studies. Life Sci. 2011, 89, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Wullimann, M.F. Secondary neurogenesis and telencephalic organization in zebrafish and mice: A brief review. Integr. Zool. 2009, 4, 123–133. [Google Scholar] [CrossRef]
- Nishimura, Y.; Murakami, S.; Ashikawa, Y.; Sasagawa, S.; Umemoto, N.; Shimada, Y.; Tanaka, T. Zebrafish as a systems toxicology model for developmental neurotoxicity testing. Congenit. Anom. 2015, 55, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Crosby, E.B.; Bailey, J.M.; Oliveri, A.N.; Levin, E.D. Neurobehavioral impairments caused by developmental imidacloprid exposure in zebrafish. Neurotoxicol. Teratol. 2015, 49, 81–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eddins, D.; Cerutti, D.; Williams, P.; Linney, E.; Levin, E.D. Zebrafish provide a sensitive model of persisting neurobehavioral effects of developmental chlorpyrifos exposure: Comparison with nicotine and pilocarpine effects and relationship to dopamine deficits. Neurotoxicol. Teratol. 2010, 32, 99–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposure | MeHg Concentrations | Number of Embryos Exposed | Survival at 6 dpf | Behaviour Testing | Comments |
|---|---|---|---|---|---|
| 1-behaviour | 0 nM | 50 embryos × 3 dishes = 150 per treatment | >95% | Larval only |
|
| 30 nM | >95% | ||||
| 100 nM | >95% | ||||
| 300 nM | All dead by 6 dpf | ||||
| 2-behaviour | 0 nM | 100 embryos × 3 dishes = 300 per treatment |
| Adult | 5 nM- euthanised at juvenile stage due to experimental design |
| 5 nM | >95% | ||||
| 15 nM | >95% | ||||
| 30 nM | >95% | ||||
| 3-behaviour | 0 nM | 100 embryos × 3 dishes = 300 per treatment | Low at 14 dpf in all groups | Larval only | Due to overall low survival at 14 dpf- all groups were euthanised at 3 weeks |
| 10 nM | |||||
| 30 nM | |||||
| 4-behaviour | 0 nM | 100 embryos × 3 dishes = 300 per treatment | >95% | Larval and adult | 30 nM- Due to low survival at juvenile stage, tanks 1 and 2 were pooled before adult testing |
| 5 nM | >95% | ||||
| 10 nM | >95% | ||||
| 30 nM | >95% | ||||
| 5-Larval RNA | 0 nM | 50 embryos × 3 dishes = 150 per treatment | >95% | Non |
|
| 10 nM | >95% | ||||
| 100 nM | >95% |
| Gene | Primer Sequences | Product Size (bp) |
|---|---|---|
| Glucocorticoid Receptor alpha (grα) | F 5’ ACTCCATGCACGACTTGGTG 3’ R 5’ GCATTTCGGGAAACTCCACG 3’ | 90 |
| Mineralocorticoid Receptor (mr) | F 5’ CTTCCAGGTTTCCGCAGTCTAC 3’ R 5’ GGAGGAGAGACACATCCAGGAAT 3’ | 74 |
| Corticotrophin Releasing Factor (crf) | F 5’ CGAGACATCCCAGTATCCAAAAAG 3’ R 5’ TCCAACAGACGCTGCGTTAA 3’ | 59 |
| Dopamine transporter (dat1) | F 5’ GCCTGGTTTTACGGAGTGGA 3’ R 5’ GGAGGATTGAAGGTGGCGAA 3’ | 152 |
| Dopamine receptor 2b (drd2b) | F 5’ GCACGGCCAGCATTCTTAATC 3’ R 5’ GAAAGCACCCAAACAACGGA 3’ | 136 |
| ß-actin | F 5’ CGAGCTGTCTTCCCATCCA 3’ R 5’ TCACCAACGTAGCTGTCTTTCTG 3’ | 86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glazer, L.; Brennan, C.H. Developmental Exposure to Low Concentrations of Methylmercury Causes Increase in Anxiety-Related Behaviour and Locomotor Impairments in Zebrafish. Int. J. Mol. Sci. 2021, 22, 10961. https://doi.org/10.3390/ijms222010961
Glazer L, Brennan CH. Developmental Exposure to Low Concentrations of Methylmercury Causes Increase in Anxiety-Related Behaviour and Locomotor Impairments in Zebrafish. International Journal of Molecular Sciences. 2021; 22(20):10961. https://doi.org/10.3390/ijms222010961
Chicago/Turabian StyleGlazer, Lilah, and Caroline H. Brennan. 2021. "Developmental Exposure to Low Concentrations of Methylmercury Causes Increase in Anxiety-Related Behaviour and Locomotor Impairments in Zebrafish" International Journal of Molecular Sciences 22, no. 20: 10961. https://doi.org/10.3390/ijms222010961
APA StyleGlazer, L., & Brennan, C. H. (2021). Developmental Exposure to Low Concentrations of Methylmercury Causes Increase in Anxiety-Related Behaviour and Locomotor Impairments in Zebrafish. International Journal of Molecular Sciences, 22(20), 10961. https://doi.org/10.3390/ijms222010961