
Telomeric-Like Repeats Flanked by Sequences Retrotranscribed from the Telomerase RNA Inserted at DNA Double-Strand Break Sites during Vertebrate Genome Evolution

 , ,
, ,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
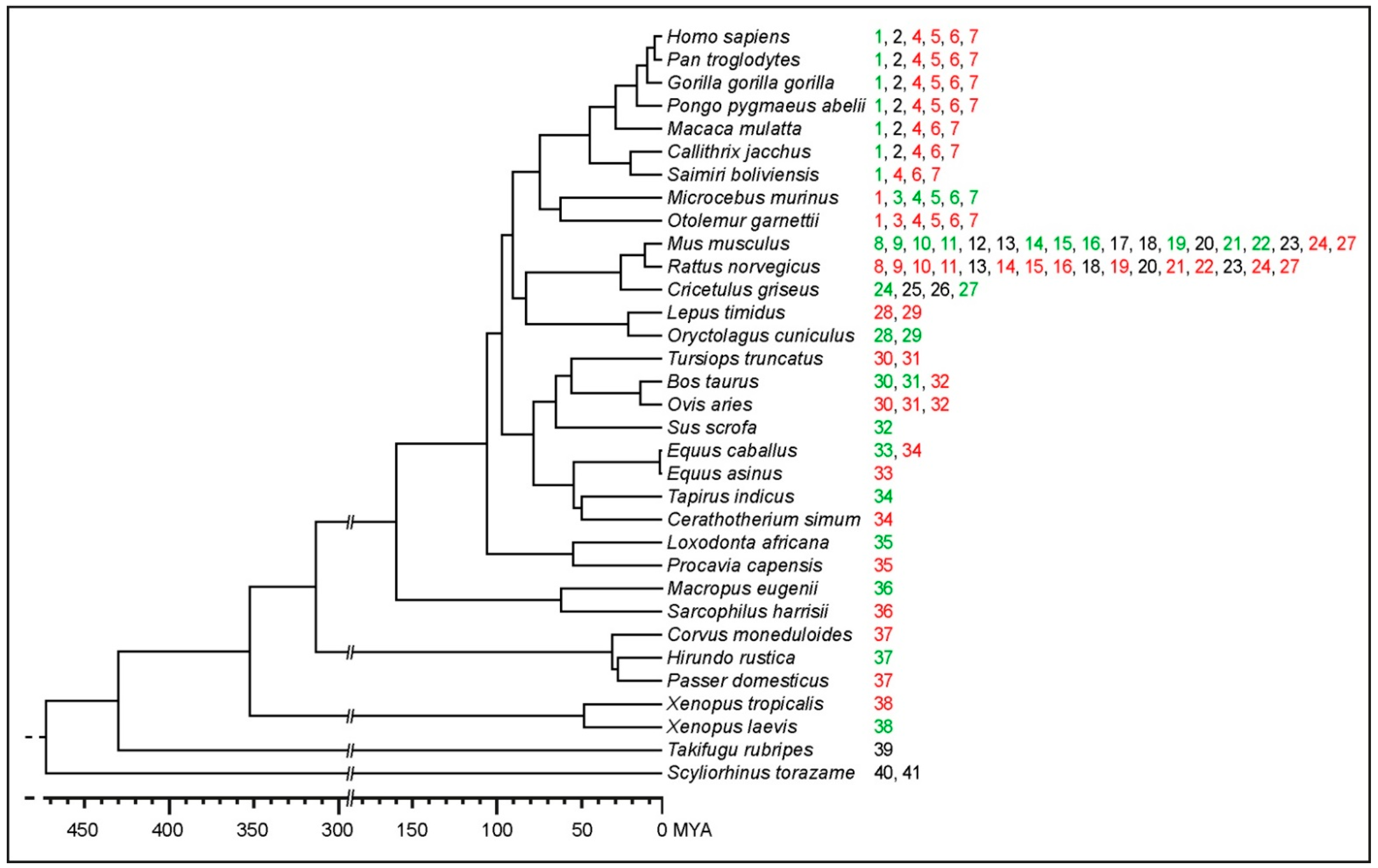
2.1. The Search for TERC-ITS Loci in Vertebrates
2.2. Evolutionary Conservation Analysis
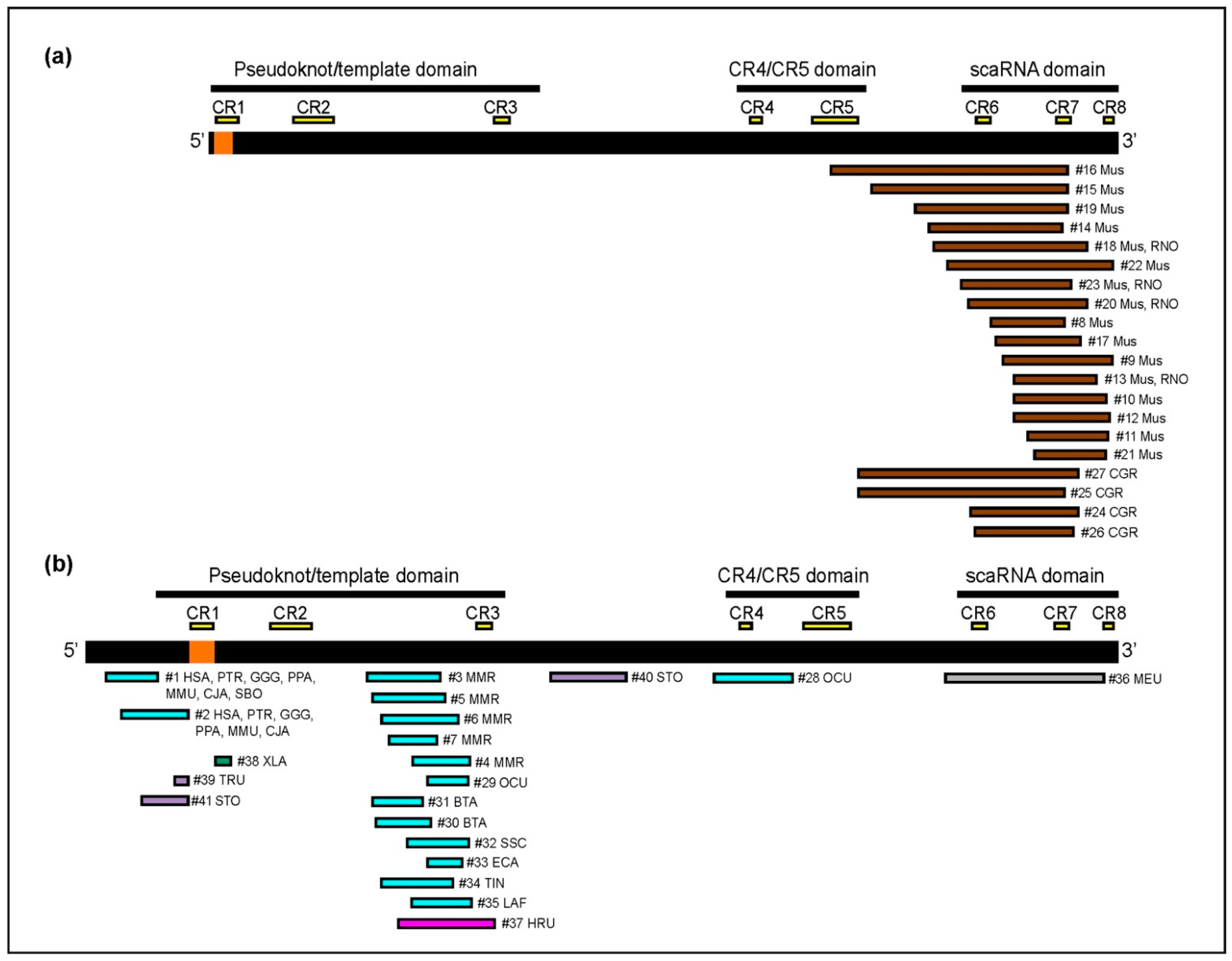
2.3. Insertion Site Characterization
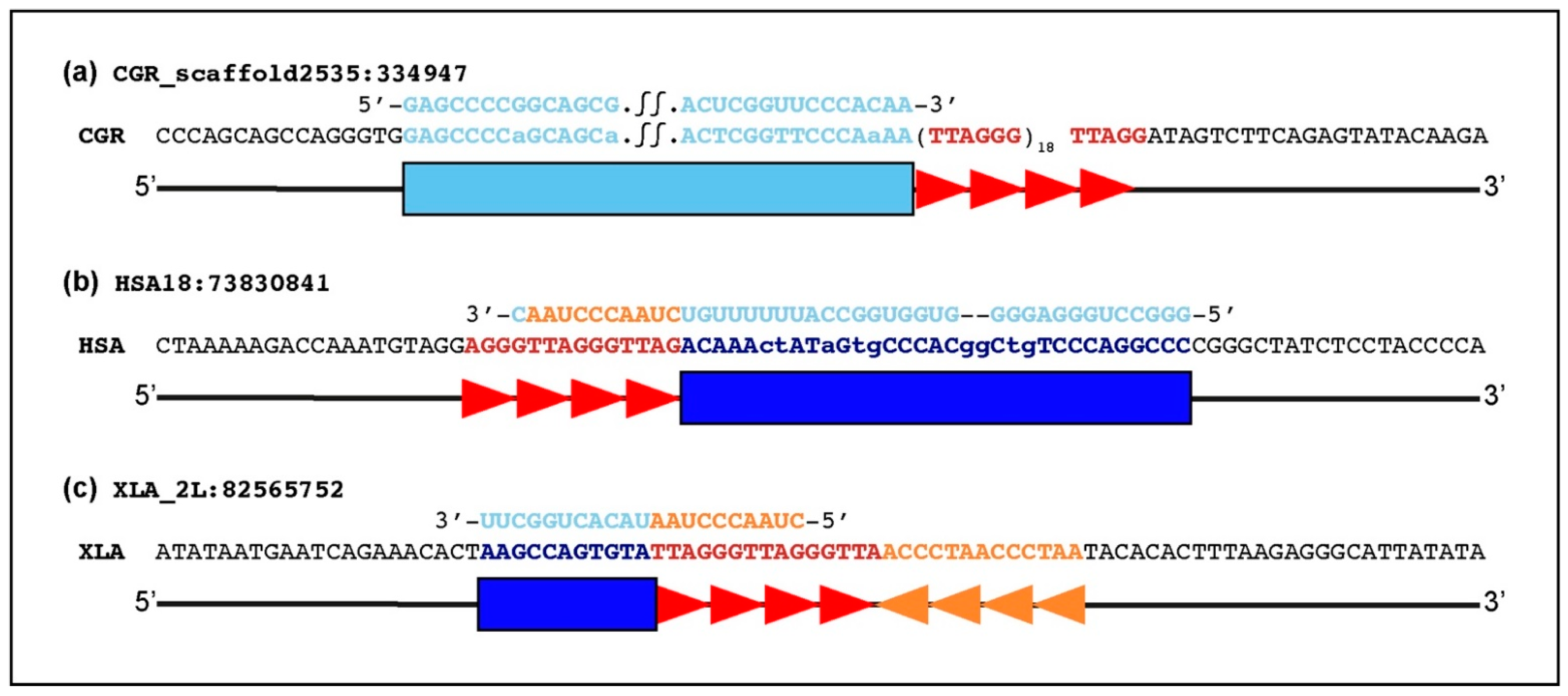
2.4. Sequence Organization of TERC-ITS Loci
3. Discussion
3.1. TERC-ITSs in Vertebrates
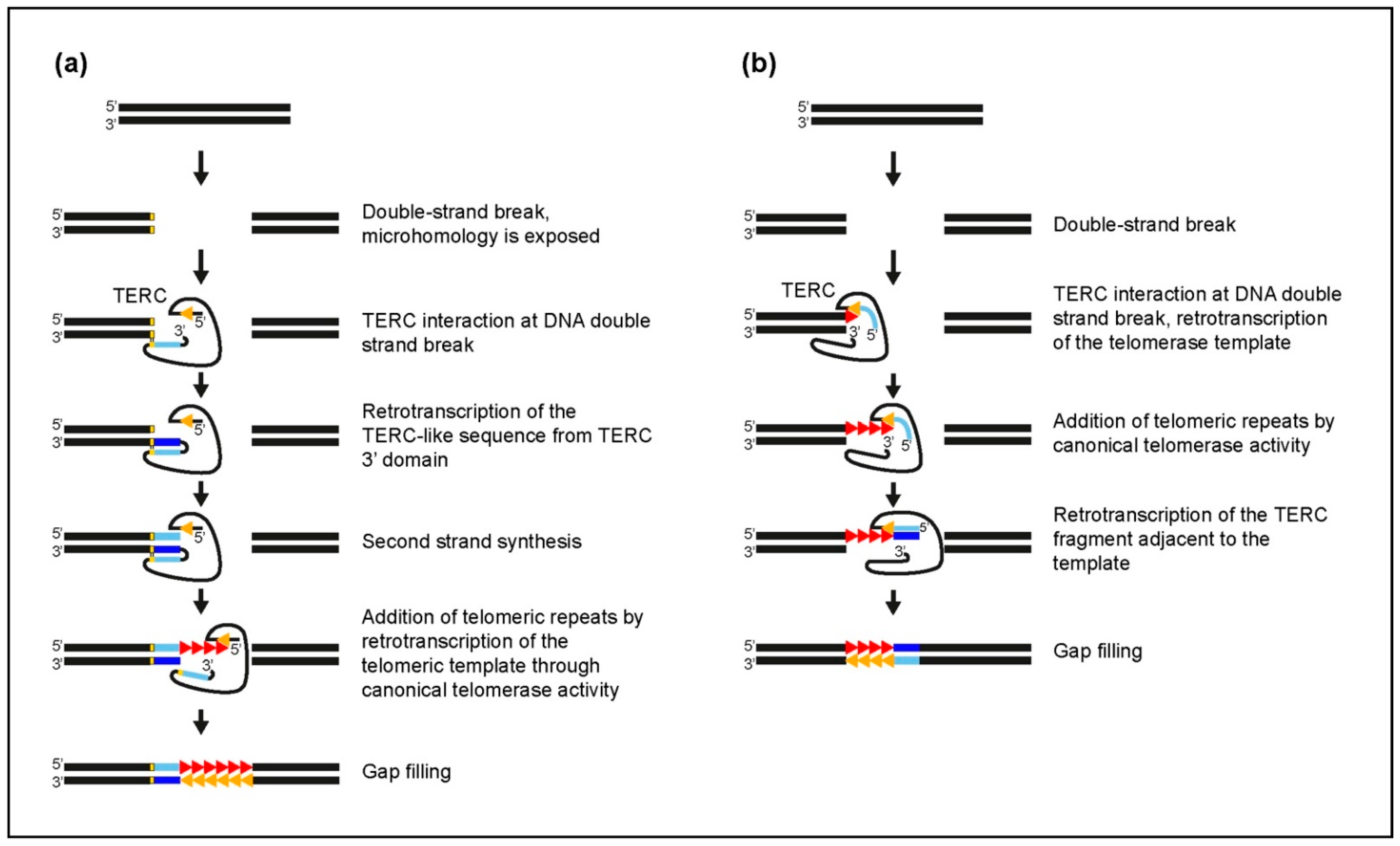
3.2. Insertion Mechanisms of TERC-ITSs
4. Materials and Methods
4.1. Identification of TERC Genes in 30 Vertebrate Species
4.2. Search for TERC-ITS Loci in Vertebrate Genomes
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- De Lange, T. Shelterin-Mediated Telomere Protection. Annu. Rev. Genet. 2018, 52, 223–247. [Google Scholar] [CrossRef] [PubMed]
- Azzalin, C.M.; Reichenbach, P.; Khoriauli, L.; Giulotto, E.; Lingner, J. Telomeric Repeat Containing RNA and RNA Surveillance Factors at Mammalian Chromosome Ends. Science 2007, 318, 798–801. [Google Scholar] [CrossRef] [PubMed]
- Bettin, N.; Oss Pegorar, C.; Cusanelli, E. The Emerging Roles of TERRA in Telomere Maintenance and Genome Stability. Cells 2019, 8, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitelli, V.; Falvo, P.; G Nergadze, S.; Santagostino, M.; Khoriauli, L.; Pellanda, P.; Bertino, G.; Occhini, A.; Benazzo, M.; Morbini, P.; et al. Telomeric Repeat-Containing RNAs (TERRA) Decrease in Squamous Cell Carcinoma of the Head and Neck Is Associated with Worsened Clinical Outcome. Int. J. Mol. Sci. 2018, 19, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Liu, D.; Yang, H.; Yu, S.; He, H. Long Noncoding RNAs in Head and Neck Squamous Cell Carcinoma: Biological Functions and Mechanisms. Mol. Biol. Rep. 2020, 47, 8075–8090. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.L.; Blasco, M.A.; Greider, C.W. Secondary Structure of Vertebrate Telomerase RNA. Cell 2000, 100, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Mosig, A.; Qi, X.; Li, Y.; Stadler, P.F.; Chen, J.J.-L. Structure and Function of the Smallest Vertebrate Telomerase RNA from Teleost Fish. J. Biol. Chem. 2008, 283, 2049–2059. [Google Scholar] [CrossRef] [Green Version]
- Podlevsky, J.D.; Chen, J.J.-L. Evolutionary Perspectives of Telomerase RNA Structure and Function. RNA Biol. 2016, 13, 720–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logeswaran, D.; Li, Y.; Podlevsky, J.D.; Chen, J.J.-L. Monophyletic Origin and Divergent Evolution of Animal Telomerase RNA. Mol. Biol. Evol. 2021, 38, 215–228. [Google Scholar] [CrossRef]
- Greider, C.W.; Blackburn, E.H. A Telomeric Sequence in the RNA of Tetrahymena Telomerase Required for Telomere Repeat Synthesis. Nature 1989, 337, 331–337. [Google Scholar] [CrossRef]
- Qi, X.; Li, Y.; Honda, S.; Hoffmann, S.; Marz, M.; Mosig, A.; Podlevsky, J.D.; Stadler, P.F.; Selker, E.U.; Chen, J.J.-L. The Common Ancestral Core of Vertebrate and Fungal Telomerase RNAs. Nucleic Acids Res. 2013, 41, 450–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunisova, S.; Elboher, E.; Nosek, J.; Gorkovoy, V.; Brown, Y.; Lucier, J.-F.; Laterreur, N.; Wellinger, R.J.; Tzfati, Y.; Tomaska, L. Identification and Comparative Analysis of Telomerase RNAs from Candida Species Reveal Conservation of Functional Elements. RNA 2009, 15, 546–559. [Google Scholar] [CrossRef] [Green Version]
- Tesmer, V.M.; Ford, L.P.; Holt, S.E.; Frank, B.C.; Yi, X.; Aisner, D.L.; Ouellette, M.; Shay, J.W.; Wright, W.E. Two Inactive Fragments of the Integral RNA Cooperate to Assemble Active Telomerase with the Human Protein Catalytic Subunit (HTERT) in Vitro. Mol. Cell Biol. 1999, 19, 6207–6216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachand, F.; Autexier, C. Functional Regions of Human Telomerase Reverse Transcriptase and Human Telomerase RNA Required for Telomerase Activity and RNA-Protein Interactions. Mol. Cell Biol. 2001, 21, 1888–1897. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.-L.; Opperman, K.K.; Greider, C.W. A Critical Stem-Loop Structure in the CR4-CR5 Domain of Mammalian Telomerase RNA. Nucleic Acids Res. 2002, 30, 592–597. [Google Scholar] [CrossRef]
- Jády, B.E.; Bertrand, E.; Kiss, T. Human Telomerase RNA and Box H/ACA ScaRNAs Share a Common Cajal Body-Specific Localization Signal. J. Cell Biol. 2004, 164, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Ly, H.; Hussain, A.; Abraham, M.; Pearl, S.; Tzfati, Y.; Parslow, T.G.; Blackburn, E.H. A Universal Telomerase RNA Core Structure Includes Structured Motifs Required for Binding the Telomerase Reverse Transcriptase Protein. Proc. Natl. Acad. Sci. USA 2004, 101, 14713–14718. [Google Scholar] [CrossRef] [Green Version]
- Brown, Y.; Abraham, M.; Pearl, S.; Kabaha, M.M.; Elboher, E.; Tzfati, Y. A Critical Three-Way Junction Is Conserved in Budding Yeast and Vertebrate Telomerase RNAs. Nucleic Acids Res. 2007, 35, 6280–6289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greider, C.W. Telomerase Is Processive. Mol. Cell Biol. 1991, 11, 4572–4580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyne, J.; Baker, R.J.; Hobart, H.H.; Hsu, T.C.; Ryder, O.A.; Ward, O.G.; Wiley, J.E.; Wurster-Hill, D.H.; Yates, T.L.; Moyzis, R.K. Distribution of Non-Telomeric Sites of the (TTAGGG)n Telomeric Sequence in Vertebrate Chromosomes. Chromosoma 1990, 99, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Herrera, A.; Nergadze, S.G.; Santagostino, M.; Giulotto, E. Telomeric Repeats Far from the Ends: Mechanisms of Origin and Role in Evolution. Cytogenet. Genome Res. 2008, 122, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Faravelli, M.; Moralli, D.; Bertoni, L.; Attolini, C.; Chernova, O.; Raimondi, E.; Giulotto, E. Two Extended Arrays of a Satellite DNA Sequence at the Centromere and at the Short-Arm Telomere of Chinese Hamster Chromosome 5. Cytogenet. Cell Genet. 1998, 83, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Faravelli, M.; Azzalin, C.M.; Bertoni, L.; Chernova, O.; Attolini, C.; Mondello, C.; Giulotto, E. Molecular Organization of Internal Telomeric Sequences in Chinese Hamster Chromosomes. Gene 2002, 283, 11–16. [Google Scholar] [CrossRef]
- Nanda, I.; Schrama, D.; Feichtinger, W.; Haaf, T.; Schartl, M.; Schmid, M. Distribution of Telomeric (TTAGGG)(n) Sequences in Avian Chromosomes. Chromosoma 2002, 111, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Ventura, K.; Silva, M.J.J.; Fagundes, V.; Christoff, A.U.; Yonenaga-Yassuda, Y. Non-Telomeric Sites as Evidence of Chromosomal Rearrangement and Repetitive (TTAGGG)n Arrays in Heterochromatic and Euchromatic Regions in Four Species of Akodon (Rodentia, Muridae). Cytogenet. Genome Res. 2006, 115, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Kratochvíl, L.; Altmanová, M.; Johnson Pokorná, M. Interstitial Telomeric Motifs in Squamate Reptiles: When the Exceptions Outnumber the Rule. PLoS ONE 2015, 10, e0134985. [Google Scholar] [CrossRef] [Green Version]
- Zattera, M.L.; Lima, L.; Duarte, I.; de Sousa, D.Y.; Araújo, O.G.D.S.; Gazoni, T.; Mott, T.; Recco-Pimentel, S.M.; Bruschi, D.P. Chromosome Spreading of the (TTAGGG)n Repeats in the Pipa Carvalhoi Miranda-Ribeiro, 1937 (Pipidae, Anura) Karyotype. Comp. Cytogenet. 2019, 13, 297–309. [Google Scholar] [CrossRef] [PubMed]
- IJdo, J.W.; Baldini, A.; Ward, D.C.; Reeders, S.T.; Wells, R.A. Origin of Human Chromosome 2: An Ancestral Telomere-Telomere Fusion. Proc. Natl. Acad. Sci. USA 1991, 88, 9051–9055. [Google Scholar] [CrossRef] [Green Version]
- Azzalin, C.M.; Nergadze, S.G.; Giulotto, E. Human Intrachromosomal Telomeric-like Repeats: Sequence Organization and Mechanisms of Origin. Chromosoma 2001, 110, 75–82. [Google Scholar] [CrossRef]
- Azzalin, C.M.; Mucciolo, E.; Bertoni, L.; Giulotto, E. Fluorescence in Situ Hybridization with a Synthetic (T2AG3)n Polynucleotide Detects Several Intrachromosomal Telomere-like Repeats on Human Chromosomes. Cytogenet. Cell Genet. 1997, 78, 112–115. [Google Scholar] [CrossRef]
- Ruiz-Herrera, A.; García, F.; Azzalin, C.; Giulotto, E.; Egozcue, J.; Ponsà, M.; Garcia, M. Distribution of Intrachromosomal Telomeric Sequences (ITS) on Macaca Fascicularis (Primates) Chromosomes and Their Implication for Chromosome Evolution. Hum. Genet. 2002, 110, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Nergadze, S.G.; Rocchi, M.; Azzalin, C.M.; Mondello, C.; Giulotto, E. Insertion of Telomeric Repeats at Intrachromosomal Break Sites during Primate Evolution. Genome Res. 2004, 14, 1704–1710. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Herrera, A.; García, F.; Giulotto, E.; Attolini, C.; Egozcue, J.; Ponsà, M.; Garcia, M. Evolutionary Breakpoints Are Co-Localized with Fragile Sites and Intrachromosomal Telomeric Sequences in Primates. Cytogenet. Genome Res. 2005, 108, 234–247. [Google Scholar] [CrossRef]
- Nergadze, S.G.; Santagostino, M.A.; Salzano, A.; Mondello, C.; Giulotto, E. Contribution of Telomerase RNA Retrotranscription to DNA Double-Strand Break Repair during Mammalian Genome Evolution. Genome Biol. 2007, 8, R260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santagostino, M.; Piras, F.M.; Cappelletti, E.; Del Giudice, S.; Semino, O.; Nergadze, S.G.; Giulotto, E. Insertion of Telomeric Repeats in the Human and Horse Genomes: An Evolutionary Perspective. Int. J. Mol. Sci. 2020, 21, 2838. [Google Scholar] [CrossRef] [Green Version]
- Rebuzzini, P.; Khoriauli, L.; Azzalin, C.M.; Magnani, E.; Mondello, C.; Giulotto, E. New Mammalian Cellular Systems to Study Mutations Introduced at the Break Site by Non-Homologous End-Joining. DNA Repair. (Amst.) 2005, 4, 546–555. [Google Scholar] [CrossRef]
- Onozawa, M.; Zhang, Z.; Kim, Y.J.; Goldberg, L.; Varga, T.; Bergsagel, P.L.; Kuehl, W.M.; Aplan, P.D. Repair of DNA Double-Strand Breaks by Templated Nucleotide Sequence Insertions Derived from Distant Regions of the Genome. Proc. Natl. Acad. Sci. USA 2014, 111, 7729–7734. [Google Scholar] [CrossRef] [Green Version]
- NCBI Gene. Available online: https://www.ncbi.nlm.nih.gov/gene (accessed on 1 April 2021).
- Podlevsky, J.D.; Bley, C.J.; Omana, R.V.; Qi, X.; Chen, J.J.-L. The Telomerase Database. Nucleic Acids Res. 2008, 36, D339–D343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telomerase Database. Available online: http://telomerase.asu.edu (accessed on 1 April 2021).
- Smith, J.; Baldeyron, C.; De Oliveira, I.; Sala-Trepat, M.; Papadopoulo, D. The Influence of DNA Double-Strand Break Structure on End-Joining in Human Cells. Nucleic Acids Res. 2001, 29, 4783–4792. [Google Scholar] [CrossRef] [Green Version]
- UCSC Table Browser. Available online: https://genome.ucsc.edu/cgi-bin/hgTables (accessed on 5 October 2021).
- Smit, A.; Hubley, R.; Green, P. RepeatMasker Open-4.0. Available online: http://www.repeatmasker.org (accessed on 1 April 2021).
- Nergadze, S.G.; Lupotto, M.; Pellanda, P.; Santagostino, M.; Vitelli, V.; Giulotto, E. Mitochondrial DNA Insertions in the Nuclear Horse Genome. Anim. Genet. 2010, 41 (Suppl. S2), 176–185. [Google Scholar] [CrossRef]
- Zemojtel, T.; Penzkofer, T.; Schultz, J.; Dandekar, T.; Badge, R.; Vingron, M. Exonization of Active Mouse L1s: A Driver of Transcriptome Evolution? BMC Genomics 2007, 8, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platt, R.N.; Vandewege, M.W.; Ray, D.A. Mammalian Transposable Elements and Their Impacts on Genome Evolution. Chromosome Res. 2018, 26, 25–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.-L.; Greider, C.W. Template Boundary Definition in Mammalian Telomerase. Genes Dev. 2003, 17, 2747–2752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouet, P.; Smih, F.; Jasin, M. Introduction of Double-Strand Breaks into the Genome of Mouse Cells by Expression of a Rare-Cutting Endonuclease. Mol. Cell. Biol. 1994, 14, 8096–8106. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.H.Y.; Pannunzio, N.R.; Adachi, N.; Lieber, M.R. Non-Homologous DNA End Joining and Alternative Pathways to Double-Strand Break Repair. Nat. Rev. Mol. Cell Biol. 2017, 18, 495–506. [Google Scholar] [CrossRef]
- Hsu, H.L.; Gilley, D.; Blackburn, E.H.; Chen, D.J. Ku Is Associated with the Telomere in Mammals. Proc. Natl. Acad. Sci. USA 1999, 96, 12454–12458. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, P.S.; Stavropoulos, D.J.; Meyn, M.S. Human Telomeric Protein TRF2 Associates with Genomic Double-Strand Breaks as an Early Response to DNA Damage. Nat. Genet. 2005, 37, 193–197. [Google Scholar] [CrossRef]
- Ting, N.S.Y.; Yu, Y.; Pohorelic, B.; Lees-Miller, S.P.; Beattie, T.L. Human Ku70/80 Interacts Directly with HTR, the RNA Component of Human Telomerase. Nucleic Acids Res. 2005, 33, 2090–2098. [Google Scholar] [CrossRef] [Green Version]
- Simonet, T.; Zaragosi, L.-E.; Philippe, C.; Lebrigand, K.; Schouteden, C.; Augereau, A.; Bauwens, S.; Ye, J.; Santagostino, M.; Giulotto, E.; et al. The Human TTAGGG Repeat Factors 1 and 2 Bind to a Subset of Interstitial Telomeric Sequences and Satellite Repeats. Cell Res. 2011, 21, 1028–1038. [Google Scholar] [CrossRef] [Green Version]
- Imran, S.A.M.; Yazid, M.D.; Cui, W.; Lokanathan, Y. The Intra- and Extra-Telomeric Role of TRF2 in the DNA Damage Response. Int. J. Mol. Sci. 2021, 22, 9900. [Google Scholar] [CrossRef]
- Stellwagen, A.E.; Haimberger, Z.W.; Veatch, J.R.; Gottschling, D.E. Ku Interacts with Telomerase RNA to Promote Telomere Addition at Native and Broken Chromosome Ends. Genes Dev. 2003, 17, 2384–2395. [Google Scholar] [CrossRef] [Green Version]
- Flint, J.; Craddock, C.F.; Villegas, A.; Bentley, D.P.; Williams, H.J.; Galanello, R.; Cao, A.; Wood, W.G.; Ayyub, H.; Higgs, D.R. Healing of Broken Human Chromosomes by the Addition of Telomeric Repeats. Am. J. Hum. Genet. 1994, 55, 505–512. [Google Scholar] [PubMed]
- Sprung, C.N.; Reynolds, G.E.; Jasin, M.; Murnane, J.P. Chromosome Healing in Mouse Embryonic Stem Cells. Proc. Natl. Acad. Sci. USA 1999, 96, 6781–6786. [Google Scholar] [CrossRef] [Green Version]
- Laudadio, I.; Orso, F.; Azzalin, G.; Calabrò, C.; Berardinelli, F.; Coluzzi, E.; Gioiosa, S.; Taverna, D.; Sgura, A.; Carissimi, C.; et al. AGO2 Promotes Telomerase Activity and Interaction between the Telomerase Components TERT and TERC. EMBO Rep. 2019, 20, e45969. [Google Scholar] [CrossRef] [PubMed]
- Sauerwald, A.; Sandin, S.; Cristofari, G.; Scheres, S.H.W.; Lingner, J.; Rhodes, D. Structure of Active Dimeric Human Telomerase. Nat. Struct. Mol. Biol. 2013, 20, 454–460. [Google Scholar] [CrossRef]
- Tomaska, L.; Nosek, J.; Kramara, J.; Griffith, J.D. Telomeric Circles: Universal Players in Telomere Maintenance? Nat. Struct. Mol. Biol. 2009, 16, 1010–1015. [Google Scholar] [CrossRef] [PubMed]
- NCBI Assembly. Available online: https://www.ncbi.nlm.nih.gov/assembly (accessed on 1 April 2021).
- BLAST. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 1 April 2021).
- Corpet, F. Multiple Sequence Alignment with Hierarchical Clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef] [PubMed]
- Multalin. Available online: http://multalin.toulouse.inra.fr/multalin/ (accessed on 1 April 2021).
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef] [PubMed]
- Timetree. Available online: http://www.timetree.org (accessed on 1 April 2021).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Species | Chromosomal Localization | Starting Nucleotide of the TERC-Like Sequence (Length) | Position within the TERC Sequence | ITS Length (bp) | Orthologous TERC-ITS Loci | Orthologous Empty Loci |
|---|---|---|---|---|---|---|---|
| 1 | Homo sapiens | HSA6 | 33092493 (25) | 8–32 | 6 | PTR, GGG, PPA, MMU, CJA, SBO | MMR, OGA |
| 2 | Homo sapiens | HSA18 | 73830841 (31) | 15–45 | 14 | PTR, GGG, PPA, MMU, CJA | ND |
| 3 | Microcebus murinus | MMR3 | 9808477 (35) | 120–154 | 43 | ND | OGA |
| 4 | Microcebus murinus | MMR4 | 11270478 (27) | 142–168 | 43 | ND | HSA, PTR, GGG, PPA, MMU, CJA, SBO, OGA |
| 5 | Microcebus murinus | MMR6 | 96205947 (34) | 123–157 | 24 | ND | HSA, PTR, GGG, PPA, OGA |
| 6 | Microcebus murinus | MMR14 | 66686502 (36) | 127–162 | 32 | ND | HSA, PTR, GGG, PPA, MMU, CJA, SBO, OGA |
| 7 | Microcebus murinus | MMR32 | 2429016 (28) | 129–156 | 38 | ND | HSA, PTR, GGG, PPA, MMU, CJA, SBO, OGA |
| 8 | Mus musculus1 | Mus1 | 46771696 (31) | 341–374 | 57 | ND | RNO |
| 9 | Mus musculus1 | Mus1 | 69071165 (50) | 346–395 | 139 | ND | RNO |
| 10 | Mus musculus1 | Mus4 | 119841389 (42) | 351–392 | 53 | ND | RNO |
| 11 | Mus musculus1 | Mus5 | 25644087 (37) | 357–393 | 21 | ND | RNO |
| 12 | Mus musculus1 | Mus6 | 67988979 (44) | 351–394 | 93 | ND | ND |
| 13 | Mus musculus1 | Mus8 | 23257793 (38) | 351–388 | 213 | RNO | ND |
| 14 | Mus musculus1 | Mus9 | 47869714 (60) | 314–373 | 68 | ND | RNO |
| 15 | Mus musculus1 | Mus10 | 20335691 (98) | 289–388 | 108 | ND | RNO |
| 16 | Mus musculus1 | Mus10 | 58382382 (118) | 271–388 | 27 | ND | RNO |
| 17 | Mus musculus1 | Mus11 | 86905013 (38) | 343–381 | 55 | ND | ND |
| 18 | Mus musculus | Mus12 | 7976282 (68) | 316–384 | 77 | RNO | ND |
| 19 | Mus musculus1 | Mus12 | 111293353 (81) | 308–388 | 13 | ND | RNO |
| 20 | Mus musculus1 | Mus13 | 3641558 (54) | 331–384 | 22 | RNO | ND |
| 21 | Mus musculus | Mus17 | 77320142 (33) | 360–392 | 58 | ND | RNO |
| 22 | Mus musculus1 | MusX | 65734790 (74) | 322–395 | 23 | ND | RNO |
| 23 | Mus musculus1 | MusX | 99124249 (52) | 328–377 | 25 | RNO | ND |
| 24 | Cricetulus griseus | CGR_scaffold364 | 1907954 (49) | 332–380 | 60 | ND | Mus, RNO |
| 25 | Cricetulus griseus | CGR_scaffold477 | 2816499 (92) | 283–374 | 205 | ND | ND |
| 26 | Cricetulus griseus | CGR_scaffold628 | 177514 (44) | 336–379 | 35 | ND | ND |
| 27 | Cricetulus griseus | CGR_scaffold2535 | 334947 (98) | 283–380 | 113 | ND | Mus, RNO |
| 28 | Oryctolagus cuniculus | OCU20 | 24654676 (37) | 245–281 | 66 | ND | LTI |
| 29 | Oryctolagus cuniculus | OCU_chrUn0052 | 1868849 (20) | 148–167 | 39 | ND | LTI |
| 30 | Bos taurus | BTA12 | 22246021 (26) | 125–150 | 24 | ND | OAR, SSC, TTR |
| 31 | Bos taurus | BTA20 | 7319073 (24) | 124–147 | 23 | ND | OAR, SSC, TTR |
| 32 | Sus scrofa | SSC7 | 70772067 (29) | 139–167 | 40 | ND | BTA, OAR |
| 33 | Equus caballus1 | ECA19 | 10034300 (17) | 148–164 | 35 | ND | EAS |
| 34 | Tapirus indicus | TIN_scaffold2309 | 127238 (33) | 128–160 | 26 | ND | CSS, ECA |
| 35 | Loxodonta africana | LAF_scaffold21 | 34372782 (28) | 141–168 | 19 | ND | PCA |
| 36 | Macropus eugenii | MEU_scaffold107834 | 53871 (79) | 424–502 | 12 | ND | SHA |
| 37 | Hirundo rustica | HRU5 | 20102963 (42) | 106–146 | 10 | ND | PDO, CMO |
| 38 | Xenopus laevis | XLA_2L | 82565752 (11) | 54–64 | 28 | ND | XTR |
| 39 | Takifugu rubripes | TRU18 | 10170798 (8) | 7–14 | 46 | ND | ND |
| 40 | Scyliorhinus torazame | STO_scyto00000011 | 2181744 (31) | 233–263 | 30 | ND | ND |
| 41 | Scyliorhinus torazame | STO_scf_scyto00006538 | 68375 (16) | 4–19 | 27 | ND | ND |
| Flanking Sequence Modification | Number of Loci (%) |
|---|---|
| No modification | 5 (17.2) |
| Deletion | 8 (27.6) |
| Insertion | 4 (13.8) |
| Deletion and Insertion | 12 (41.4) |
| TOTAL | 29 (100) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sola, L.; Nergadze, S.G.; Cappelletti, E.; Piras, F.M.; Giulotto, E.; Santagostino, M. Telomeric-Like Repeats Flanked by Sequences Retrotranscribed from the Telomerase RNA Inserted at DNA Double-Strand Break Sites during Vertebrate Genome Evolution. Int. J. Mol. Sci. 2021, 22, 11048. https://doi.org/10.3390/ijms222011048
Sola L, Nergadze SG, Cappelletti E, Piras FM, Giulotto E, Santagostino M. Telomeric-Like Repeats Flanked by Sequences Retrotranscribed from the Telomerase RNA Inserted at DNA Double-Strand Break Sites during Vertebrate Genome Evolution. International Journal of Molecular Sciences. 2021; 22(20):11048. https://doi.org/10.3390/ijms222011048
Chicago/Turabian StyleSola, Lorenzo, Solomon G. Nergadze, Eleonora Cappelletti, Francesca M. Piras, Elena Giulotto, and Marco Santagostino. 2021. "Telomeric-Like Repeats Flanked by Sequences Retrotranscribed from the Telomerase RNA Inserted at DNA Double-Strand Break Sites during Vertebrate Genome Evolution" International Journal of Molecular Sciences 22, no. 20: 11048. https://doi.org/10.3390/ijms222011048
APA StyleSola, L., Nergadze, S. G., Cappelletti, E., Piras, F. M., Giulotto, E., & Santagostino, M. (2021). Telomeric-Like Repeats Flanked by Sequences Retrotranscribed from the Telomerase RNA Inserted at DNA Double-Strand Break Sites during Vertebrate Genome Evolution. International Journal of Molecular Sciences, 22(20), 11048. https://doi.org/10.3390/ijms222011048