
Two TGA Transcription Factor Members from Hyper-Susceptible Soybean Exhibiting Significant Basal Resistance to Soybean mosaic virus


Abstract
:1. Introduction
2. Results
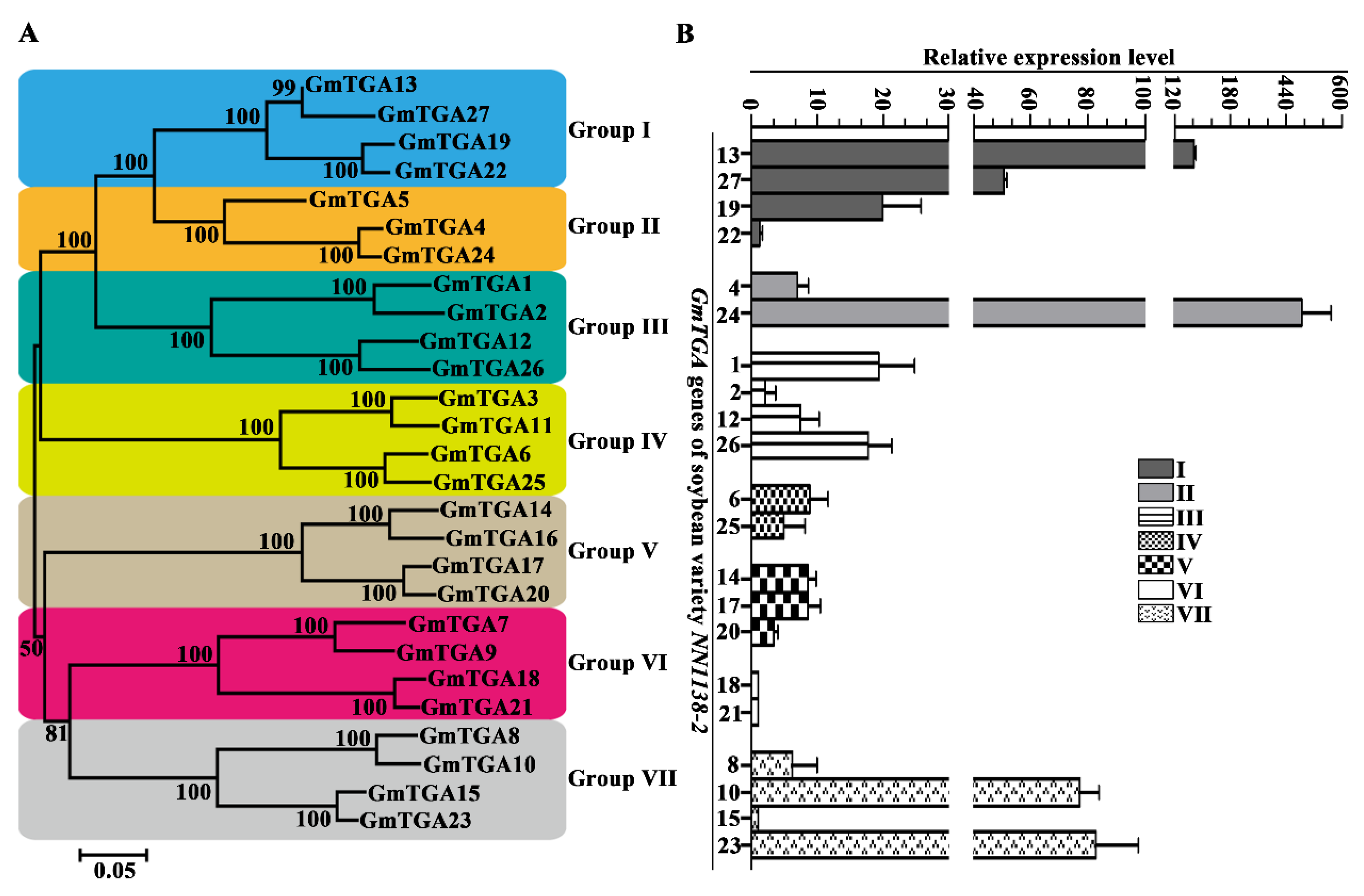
2.1. Identification and Phylogenetic Groups of GmTGAs in Soybeans
2.2. Differential Expression Profiles of GmTGAs in the Unifoliate Leaves of Soybean
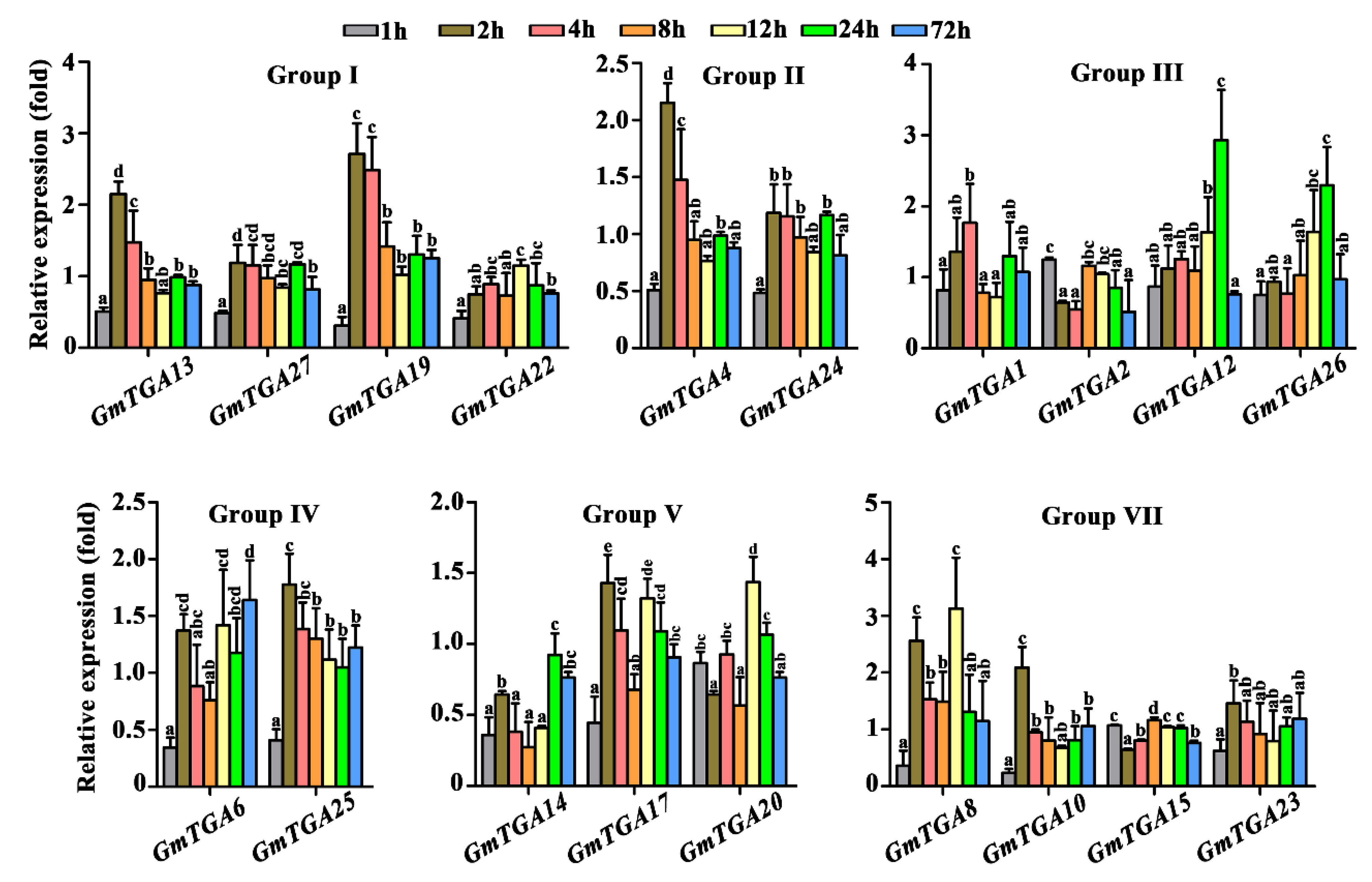
2.3. Differential Responses of GmTGA Genes to SMV Challenge
2.4. Transient Overexpression of GmTGA Genes in Nicotiana benthamiana Inhibiting but Not Promoting SMV-Multiplication
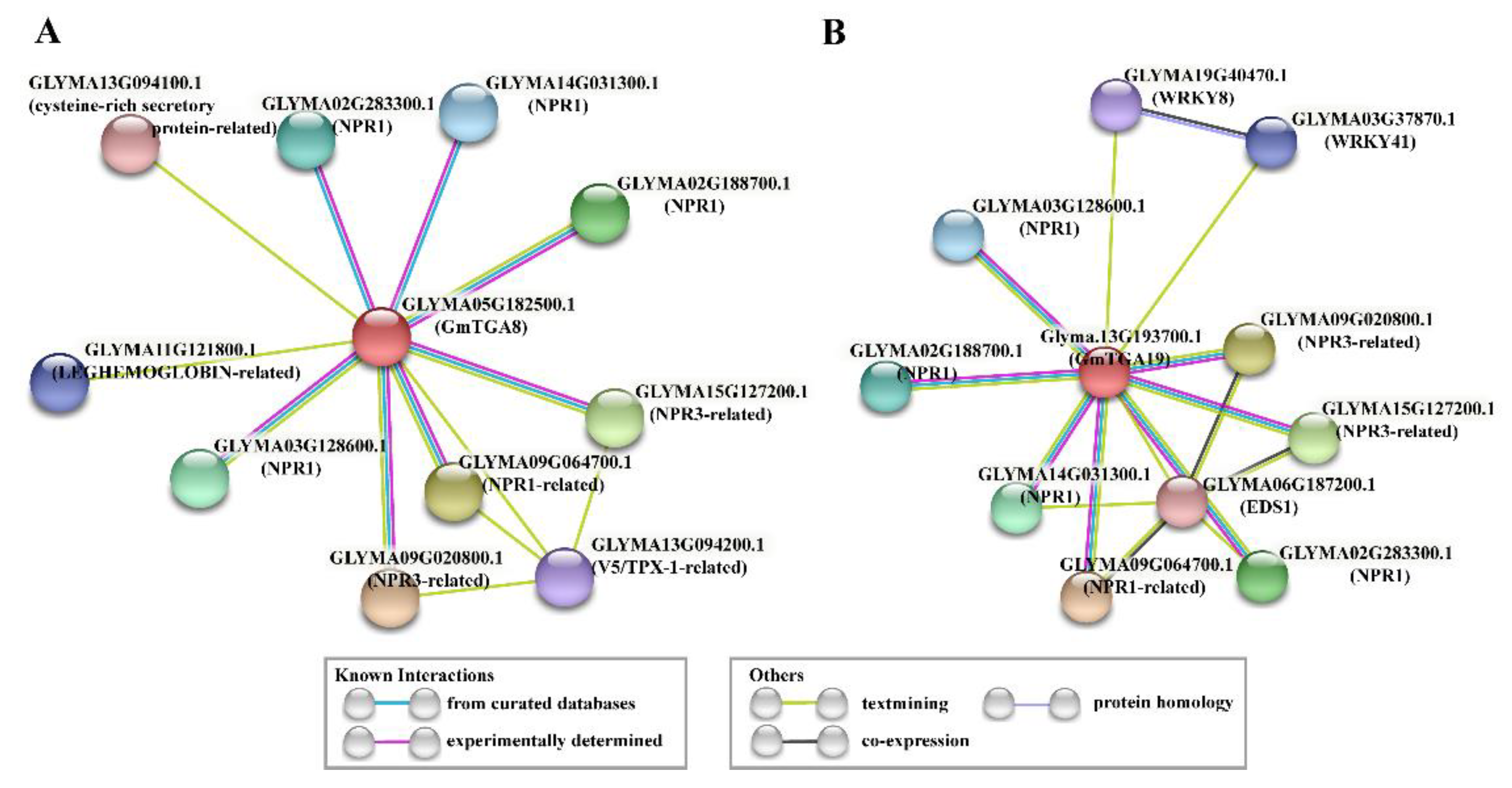
2.5. Characteristics and Interaction Network of GmTGA 8 and GmTGA19 Proteins
3. Discussion
3.1. Characteristics of the GmTGA Family in Response to SMV Stress
3.2. Potentials of GmTGA8 and GmTGA19 in Improving Soybean Resistance to SMV
4. Materials and Methods
4.1. Plant Materials and SMV Strain
4.2. Identification, Cloning and Sequence Analysis of GmTGA
4.3. Phylogenetic Analysis
4.4. SMV Inoculation and Sample Collection
4.5. The Quantitative Real-Time PCR Analysis
4.6. Construction of Transient Overexpression Constructs
4.7. Transient Assay in N. benthamiana, SMV Inoculation and ELISA Detection
4.8. Conserved Motifs and NLS Analysis in GmTGA Protein
4.9. Protein-Protein Interaction (PPI) Network Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Chittem, K.; Yajima, W.R.; Goswami, R.S.; del Río Mendoza, L.E. Transcriptome analysis of the plant pathogen Sclerotinia sclerotiorum interaction with resistant and susceptible canola (Brassica napus) lines. PLoS ONE 2020, 15, e0229844. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Shang, J.; Wang, W.; Du, J.; Li, K.; Wu, X.; Yu, L.; Liu, C.; Khaskheli, M.I.; Yang, W. Comparison of Transcriptome Differences in Soybean Response to Soybean Mosaic Virus under Normal Light and in the Shade. Viruses 2019, 11, 793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.B.; Foley, R.C.; Oñate-Sánchez, L. Transcription factors in plant defense and stress responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef]
- de-Leon, S.B.-T.; Davidson, E.H. Gene regulation: Gene control network in development. Annu. Rev. Biophys. Biomol. Struct. 2007, 36, 191–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef]
- Katagiri, F.; Lam, E.; Chua, N.-H. Two tobacco DNA-binding proteins with homology to the nuclear factor CREB. Nature 1989, 340, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, S.; Thurow, C.; Niggeweg, R.; Gatz, C. Analysis of the spacing between the two palindromes of activation sequence-1 with respect to binding to different TGA factors and transcriptional activation potential. Nucleic Acids Res. 2002, 30, 775–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, C.; Miao, Z.; Lam, E. DNA-binding properties, genomic organization and expression pattern of TGA6, a new member of the TGA family of bZIP transcription factors in Arabidopsis thaliana. Plant Mol. Biol. 1997, 34, 403–415. [Google Scholar] [CrossRef]
- Chern, M.; Bai, W.; Ruan, D.; Oh, T.; Chen, X.; Ronald, P.C. Interaction specificity and coexpression of rice NPR1 homologs 1 and 3 (NH1 and NH3), TGA transcription factors and Negative Regulator of Resistance (NRR) proteins. BMC Genom. 2014, 15, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Gatz, C. From pioneers to team players: TGA transcription factors provide a molecular link between different stress pathways. Mol. Plant-Microbe Interact. 2013, 26, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Fan, W.; Kinkema, M.; Li, X.; Dong, X. Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene. Proc. Natl. Acad. Sci. USA 1999, 96, 6523–6528. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Tessaro, M.J.; Lassner, M.; Li, X. Knockout analysis of Arabidopsis transcription factors TGA2, TGA5, and TGA6 reveals their redundant and essential roles in systemic acquired resistance. Plant Cell 2003, 15, 2647–2653. [Google Scholar] [CrossRef] [Green Version]
- Després, C.; Chubak, C.; Rochon, A.; Clark, R.; Bethune, T.; Desveaux, D.; Fobert, P.R. The Arabidopsis NPR1 disease resistance protein is a novel cofactor that confers redox regulation of DNA binding activity to the basic domain/leucine zipper transcription factor TGA1. Plant Cell 2003, 15, 2181–2191. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.-M.; Trifa, Y.; Silva, H.; Pontier, D.; Lam, E.; Shah, J.; Klessig, D.F. NPR1 differentially interacts with members of the TGA/OBF family of transcription factors that bind an element of the PR-1 gene required for induction by salicylic acid. Mol. Plant-Microbe Interact. 2000, 13, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Shearer, H.L.; Cheng, Y.T.; Wang, L.; Liu, J.; Boyle, P.; Després, C.; Zhang, Y.; Li, X.; Fobert, P.R. Arabidopsis clade I TGA transcription factors regulate plant defenses in an NPR1-independent fashion. Mol. Plant-Microbe Interact. 2012, 25, 1459–1468. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Busta, L.; Zhang, Q.; Ding, P.; Jetter, R.; Zhang, Y. TGACG-BINDING FACTOR 1 (TGA 1) and TGA 4 regulate salicylic acid and pipecolic acid biosynthesis by modulating the expression of SYSTEMIC ACQUIRED RESISTANCE DEFICIENT 1 (SARD 1) and CALMODULIN-BINDING PROTEIN 60g (CBP 60g). New Phytol. 2018, 217, 344–354. [Google Scholar] [CrossRef] [Green Version]
- Rahman, T.A.; Oirdi, M.E.; Gonzalez-Lamothe, R.; Bouarab, K. Necrotrophic pathogens use the salicylic acid signaling pathway to promote disease development in tomato. Mol. Plant Microbe Interact. 2012, 25, 1584–1593. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Huh, S.U.; Kojima, M.; Sakakibara, H.; Paek, K.-H.; Hwang, I. The cytokinin-activated transcription factor ARR2 promotes plant immunity via TGA3/NPR1-dependent salicylic acid signaling in Arabidopsis. Dev. Cell 2010, 19, 284–295. [Google Scholar] [CrossRef] [Green Version]
- Kesarwani, M.; Yoo, J.; Dong, X. Genetic interactions of TGA transcription factors in the regulation of pathogenesis-related genes and disease resistance in Arabidopsis. Plant Physiol. 2007, 144, 336–346. [Google Scholar] [CrossRef] [Green Version]
- Murmu, J.; Bush, M.J.; DeLong, C.; Li, S.; Xu, M.; Khan, M.; Malcolmson, C.; Fobert, P.R.; Zachgo, S.; Hepworth, S.R. Arabidopsis basic leucine-zipper transcription factors TGA9 and TGA10 interact with floral glutaredoxins ROXY1 and ROXY2 and are redundantly required for anther development. Plant Physiol. 2010, 154, 1492–1504. [Google Scholar] [CrossRef] [Green Version]
- Noshi, M.; Mori, D.; Tanabe, N.; Maruta, T.; Shigeoka, S. Arabidopsis clade IV TGA transcription factors, TGA10 and TGA9, are involved in ROS-mediated responses to bacterial PAMP flg22. Plant Sci 2016, 252, 12–21. [Google Scholar] [CrossRef]
- Chuang, C.-F.; Running, M.P.; Williams, R.W.; Meyerowitz, E.M. The PERIANTHIA gene encodes a bZIP protein involved in the determination of floral organ number in Arabidopsis thaliana. Genes Dev. 1999, 13, 334–344. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.H.; Whitham, S.A. Control of virus diseases in soybeans. Adv. Virus Res. 2014, 90, 355–390. [Google Scholar]
- Li, K.; Zhi, H.J. Advances in resistance to Soybean mosaic virus disease in soybean. Soybean Sci. 2016, 35, 525–530. [Google Scholar]
- Hill, J.H. Soybean mosaic virus. In Compendium of Soybean Diseases, 4th ed.; Hartman, G.L., Sinclair, J.B., Rupe, J.C., Eds.; APS Press: St. Paul, MN, USA, 1999; pp. 70–71. [Google Scholar]
- Ross, J. Effect of Aphid-Transmitted Soybean Mosaic Virus on Yields of Closely Related Resistant and Susceptible Soybean Lines 1. Crop Sci. 1977, 17, 869–872. [Google Scholar] [CrossRef]
- Hajimorad, M.R.; Domier, L.L.; Tolin, S.A.; Whitham, S.A.; Saghai Maroof, M.A. Soybean mosaic virus: A successful potyvirus with a wide distribution but restricted natural host range. Mol. Plant Pathol. 2018, 19, 1563–1579. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Li, K.; Dou, D.; Gai, J. Characterization of a soybean mosaic virus variant causing different diseases in Glycine max and Nicotiana benthamiana. Arch. Virol. 2017, 162, 549–553. [Google Scholar] [CrossRef]
- Almeida, Á.M.; Sakai, J.; Souto, E.R.; Kitajima, E.W.; Fukuji, T.S.; Hanada, K. Mosaic in Senna occidentalis in Southern Brazil induced by a new strain of Soybean mosaic virus. Fitopatol. Bras. 2002, 27, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Christie, S.R.; Crawford, W.E. Plant virus range of Nicotiana benthamiana. Plant Dis. Report. 1978, 62, 20–22. [Google Scholar]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178. [Google Scholar] [CrossRef] [Green Version]
- Ullah, I.; Magdy, M.; Wang, L.; Liu, M.; Li, X. Genome-wide identification and evolutionary analysis of TGA transcription factors in soybean. Sci. Rep. 2019, 9, 11186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen Ba, A.N.; Pogoutse, A.; Provart, N.; Moses, A.M. NLStradamus: A simple Hidden Markov Model for nuclear localization signal prediction. BMC Bioinform. 2009, 10, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieterse, C.M.; Van Loon, L. NPR1: The spider in the web of induced resistance signaling pathways. Curr. Opin. Plant Biol. 2004, 7, 456–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nijhawan, A.; Jain, M.; Tyagi, A.K.; Khurana, J.P. Genomic survey and gene expression analysis of the basic leucine zipper transcription factor family in rice. Plant Physiol. 2008, 146, 333–350. [Google Scholar] [CrossRef] [Green Version]
- Thurow, C.; Schiermeyer, A.; Krawczyk, S.; Butterbrodt, T.; Nickolov, K.; Gatz, C. Tobacco bZIP transcription factor TGA2.2 and related factor TGA2.1 have distinct roles in plant defense responses and plant development. Plant J. 2005, 44, 100–113. [Google Scholar] [CrossRef]
- Matthews, B.F.; Beard, H.; Brewer, E.; Kabir, S.; Macdonald, M.H.; Youssef, R.M. Arabidopsis genes, AtNPR1, AtTGA2 and AtPR-5, confer partial resistance to soybean cyst nematode (Heterodera glycines) when overexpressed in transgenic soybean roots. BMC Plant Biol. 2014, 14, 96. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Liu, Y.; Cui, X.-Y.; Fu, J.-D.; Zhou, Y.-B.; Zheng, W.-J.; Lan, J.-H.; Jin, L.-G.; Chen, M.; Ma, Y.-Z. Genome-Wide Characterization and Expression Analysis of Soybean TGA Transcription Factors Identified a Novel TGA Gene Involved in Drought and Salt Tolerance. Front. Plant Sci. 2019, 10, 549. [Google Scholar] [CrossRef]
- Wang, X.; Eggenberger, A.L.; Fwjr, N.; Hill, H.J. Pathogen-derived transgenic resistance to soybean mosaic virus in soybean. Mol. Breed. 2001, 8, 119–127. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Hussain, R.M.; Sheikh, A.H.; Haider, I.; Quareshy, M.; Linthorst, H.J. Arabidopsis WRKY50 and TGA transcription factors synergistically activate expression of PR1. Front. Plant Sci. 2018, 9, 930. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2017, 46, D493–D496. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A. The Pfam protein families database in 2019. Nucleic Acids Res. 2018, 47, D427–D432. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Niu, H.; Liu, C.; Zhang, J.; Hou, C.; Wang, D. Expression stabilities of candidate reference genes for RT-qPCR under different stress conditions in soybean. PLoS ONE 2013, 8, e75271. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Model Name (Wm82.a2.v1) | Location Coordinates | ORF (bp) | Size (aa) | MW (kDa) | pI |
|---|---|---|---|---|---|---|
| GmTGA1 | Glyma.01G084200 | Chr01:24682405..24689210 (−) | 1461 | 486 | 54.31 | 7.32 |
| GmTGA2 | Glyma.02G097900 | Chr02:8834516..8841154 (−) | 1404 | 467 | 51.85 | 7.32 |
| GmTGA3 | Glyma.02G176800 | Chr02:29019977..29028838 (−) | 1455 | 484 | 53.93 | 7.77 |
| GmTGA4 | Glyma.03G127600 | Chr03:34171878..34178336 (−) | 1383 | 460 | 50.93 | 7.00 |
| GmTGA5 | Glyma.03G128200 | Chr03:34259590..34264026 (+) | 873 | 290 | 32.37 | 8.82 |
| GmTGA6 | Glyma.03G142400 | Chr03:35810204..35817330 (−) | 1476 | 491 | 55.20 | 8.22 |
| GmTGA7 | Glyma.04G254800 | Chr04:52132354..52137159 (−) | 1089 | 362 | 40.93 | 8.69 |
| GmTGA8 | Glyma.05G182500 | Chr05:37029299..37037313 (+) | 1113 | 370 | 42.08 | 7.11 |
| GmTGA9 | Glyma.06G107300 | Chr06:8630095..8635111 (+) | 1068 | 355 | 39.98 | 6.21 |
| GmTGA10 | Glyma.08G140100 | Chr08:10719899..10725773 (+) | 1140 | 379 | 43.11 | 7.78 |
| GmTGA11 | Glyma.10G092100 | Chr10:12738719..12747206 (+) | 1554 | 517 | 57.92 | 6.61 |
| GmTGA12 | Glyma.10G276100 | Chr10:49858890..49863059 (−) | 1371 | 456 | 50.99 | 6.08 |
| GmTGA13 | Glyma.10G296200 | Chr10:51306677..51314923 (−) | 999 | 332 | 37.14 | 8.94 |
| GmTGA14 | Glyma.11G183700 | Chr11:25149986..25158384 (+) | 1476 | 491 | 54.01 | 6.04 |
| GmTGA15 | Glyma.11G236300 | Chr11:33112560..33118319 (−) | 1095 | 364 | 41.18 | 6.28 |
| GmTGA16 | Glyma.12G088700 | Chr12:7269137..7277440 (−) | 1506 | 501 | 55.25 | 6.61 |
| GmTGA17 | Glyma.12G184500 | Chr12:34579255..34588903 (−) | 1467 | 488 | 54.12 | 6.75 |
| GmTGA18 | Glyma.13G085100 | Chr13:19659235..19664662 (−) | 1113 | 370 | 41.92 | 7.13 |
| GmTGA19 | Glyma.13G193700 | Chr13:30682181..30695761 (−) | 1410 | 469 | 51.99 | 5.98 |
| GmTGA20 | Glyma.13G316900 | Chr13:41139769..41147330 (+) | 1473 | 490 | 54.63 | 6.96 |
| GmTGA21 | Glyma.14G167000 | Chr14:41307404..41312938 (−) | 1113 | 370 | 41.87 | 8.26 |
| GmTGA22 | Glyma.15G232000 | Chr15:43630502..43639894 (−) | 1344 | 447 | 49.45 | 5.86 |
| GmTGA23 | Glyma.18G020900 | Chr18:1529888..1535807 (+) | 1089 | 362 | 41.05 | 7.78 |
| GmTGA24 | Glyma.19G130200 | Chr19:38989153..38995505 (−) | 1380 | 459 | 50.72 | 8.53 |
| GmTGA25 | Glyma.19G145300 | Chr19:40604030..40611501 (−) | 1473 | 490 | 55.20 | 8.51 |
| GmTGA26 | Glyma.20G113600 | Chr20:35557820..35562522 (+) | 1368 | 455 | 50.70 | 5.88 |
| GmTGA27 | Glyma.20G246400 | Chr20:47647701..47654066 (−) | 1335 | 444 | 49.51 | 5.91 |
| Sample Name | EV | SMV-Induced GmTGA Transcription Factors | + | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Group I | Group II | Group III | Group IV | Group V | Group VII | |||||||||||||||||
| Treatment | #13 | #27 | #19 | #22 | #4 | #24 | #1 | #2 | #12 | #26 | #6 | #25 | #14 | #17 | #20 | #8 | #10 | #15 | #23 | |||
| Mock | 0.10 | 0.11 | 0.13 | 0.12 | 0.11 | 0.10 | 0.12 | 0.12 | 0.12 | 0.12 | 0.12 | 0.11 | 0.12 | 0.11 | 0.12 | 0.12 | 0.12 | 0.12 | 0.11 | 0.13 | 0.12 | |
| SMV | E1 | 0.74 d | 0.41 bc | 0.40 bc | 0.28 a | 0.43 bc | 041 bc | 0.45 c | 0.47 c | 0.41 bc | 0.47 c | 0.45 c | 0.74 d | 0.45 c | 0.41 bc | 0.47 c | 0.46 c | 0.30 a | 0.42 bc | 034 ab | 0.51 c | 0.40 bc |
| E2 | 0.81 c | 0.49 b | 0.48 b | 0.32 a | 0.51 b | 0.49 b | 0.52 b | 0.55 b | 0.48 b | 0.55 b | 0.53 b | 0.80 c | 0.53 b | 0.48 b | 0.54 b | 0.53 b | 0.32 a | 0.50 b | 0.46 b | 0.58 b | 0.47 b | |
| E3 | 0.72 c | 0.42 b | 0.45 b | 0.28 a | 0.41 b | 0.43 b | 0.40 b | 0.40 b | 0.39 b | 0.40 b | 0.46 b | 0.70 c | 0.44 b | 0.44 b | 0.45 b | 0.46 b | 0.27 a | 0.43 b | 0.40 b | 0.51 b | 0.42 b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, H.; Gu, S.; Li, K.; Gai, J. Two TGA Transcription Factor Members from Hyper-Susceptible Soybean Exhibiting Significant Basal Resistance to Soybean mosaic virus. Int. J. Mol. Sci. 2021, 22, 11329. https://doi.org/10.3390/ijms222111329
Jiang H, Gu S, Li K, Gai J. Two TGA Transcription Factor Members from Hyper-Susceptible Soybean Exhibiting Significant Basal Resistance to Soybean mosaic virus. International Journal of Molecular Sciences. 2021; 22(21):11329. https://doi.org/10.3390/ijms222111329
Chicago/Turabian StyleJiang, Hua, Shengyu Gu, Kai Li, and Junyi Gai. 2021. "Two TGA Transcription Factor Members from Hyper-Susceptible Soybean Exhibiting Significant Basal Resistance to Soybean mosaic virus" International Journal of Molecular Sciences 22, no. 21: 11329. https://doi.org/10.3390/ijms222111329
APA StyleJiang, H., Gu, S., Li, K., & Gai, J. (2021). Two TGA Transcription Factor Members from Hyper-Susceptible Soybean Exhibiting Significant Basal Resistance to Soybean mosaic virus. International Journal of Molecular Sciences, 22(21), 11329. https://doi.org/10.3390/ijms222111329