
Agroinfiltration Mediated Scalable Transient Gene Expression in Genome Edited Crop Plants

 ,
, 
 and
and 
Abstract
:1. Introduction
2. Genome Editing Methods
3. Priority towards Agrobacterium-Based Genetic Transformation
4. Types of Agrobacterium-Based Genetic Transformation
5. Stable Genetic Transformation
6. Transient (Temporary) Genetic Transformation
7. Stable vs. Transient Genetic Transformation
8. Agroinfiltration
9. Methods of Agroinfiltration
10. Factors Affecting Agroinfiltration

| Plant | Family | Cultivar/Genotype (s) | Target Gene | Tissue | Method | Agrobacterium Strain (s) | Binary Vector (s) | Optimization for Agroinfiltration | Detection Methods | Remarks | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Model Plants | |||||||||||
| Arabidopsis thaliana (Arabidopsis) | Brassicaceae | Columbia (Col-0) ecotype | GUS | Leaf | Syringe (needleless) | LBA4404, C58C1, GV3101, EHA105 and AGL-1 | pCAMBIA1304 | 0.01% Triton X-100 or 0.01% Tween-20 LAB4404—Best Agrobacterium strain | GUS staining | Incubation of the infiltrated plants under short day conditions at high relative humidity maximize the gene expression | [91] |
| Cucumis melo L. (Melon) | Cucurbitaceae | - | Nattokinase (NK) | Fruit | Syringe (needle) | LBA4404 | pPZP35S, pPZP35SN, pPZP35SNi, pPZPE8, pPZPE8N and pPZPE8Ni | Acetosyringone 0.2 mm and Codon-optimized synthetic NK gene | Quantitative Real Time PCR (qRT-PCR) analysis and fibrinolytic activity | High expression of recombinant NK gene | [154] |
| Glycine max (Soybean) | Fabaceae | Williams 82, Jack, JackX, ‘Peking’, L77-1863 and Williams | GUS | Leaf and seedlings | Syringe (needleless) and vacuum | A281 EHA105 LBA4404 Ach5 and J2 | pCambia1305.1 | Infiltration buffer (10 mM 2-(N-morpholino) ethanesulfonic acid sodium salt, 10 mM MgCl2, 100 µM acetosyringone) with dithiothreitol and 30 s sonication | GUS assay | Increase in the agroinfiltration-mediated GUS expression | [155] |
| Nicotiana benthamiana (Tobacco) | Solanaceae | - | GUS | Leaf | Syringe (needleless) | EHA105 | pCAMBIA1301 | 20 µM azacytidine, 0.56 mM ascorbate and 0.03% (v/v) Tween-20 | qRT-PCR | At about 6-fold higher transient gene expression | [98] |
| - | GUS | Leaf | Syringe | AGL1, C58C1 and LBA4404 | pEAQ-GSN | Acetosyringone (500 μM), Lipoic acid (5 μM), Pluronic F-68 (0.002%) and 37 °C heat shock | GUS assay, Enzyme-linked immunoassay (ELISA) and Polyacrylamide gel electrophoresis (PAGE) analysis | Around 3.5-fold higher levels of absolute GUS protein compared to the pEAQ-HT deconstructed virus vector platform | [118] | ||
| Wild-type | Green fluorescent protein (GFP) | Leaf | Syringe (needleless) | EHA105, LBA4404, AGL0 and AGL1 | pCAMBIA(gfp)1302 | Best Agrobacterium strain-AGL0 and EHA105, acetosyringone 450–600 μM, viral protein HC-Pro, Leaf ageing | GFP imaging | High gene expression was observed in the youngest leaf | [156] | ||
| Wild type non-transgenic plants | Anthrax receptor decoy protein (immunoadhesin) and CMG2-Fc | Whole plants and detached Leaf | Vacuum | - | pBIN and pCB302 | Number of viral suppressors of post-transcriptional gene silencing constructs: p1, p10, p19, p21, p24, p25, p38, 2b, and HCPro | ELISA, Bradford assay and Western Blotting | p1 exhibit maximum gene expression contributing towards post transcriptional gene silencing | [157] | ||
| - | Ave1 and Ve1 | Leaf | Syringe | GV3101 | Gateway-compatible binary vectors | Gateway-compatible binary vectors improve agroinfiltration efficiency | Polymerase Chain Reaction (PCR) | Ve1-mediated resistance against verticillium | [158] | ||
| - | Firefly luciferase | Leaf | Syringe (needleless) | C58C1 (pGV2260) | pExp35S-LUC | Hierarchical design of promoter, leaf, plant and sampling position | Luciferase activity | Best result through sampling more positions on the same leaf | [159] | ||
| Transgenic plants | AC1, AC2, AC4 from DNA-A and BC1 from DNA-B of African cassava mosaic virus (ACMV) | Leaf | Syringe (needleless) | GV3101 | RNA interference constructs | ACMV-Cameroon:DO2:1998 transient protection assay | Electrophoresis, southern and northern hybridizations | Systemic movement of the silencing signal | [160] | ||
| - | Hemagglutinin ectodomain derived from influenza A virus strain A | Detached leaf | Vacuum | GV3101 | pMP90 | Variation in the duration of water removal treatment from 0.7 to 4.4 h | Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) | Improvement in recombinant hemagglutinin yield | [161] | ||
| - | Human epidermal growth factor | Leaf | Syringe (needleless) | GV3101 | pBYR2e-hEGF | Expression vector carrying different hEGF constructs, Agrobacterium cell density (0.2, 0.4, 0.6, and 0.8) at OD600 | ELISA | Production of recombinant hEGF protein | [162] | ||
| - | GFP, DsRed fluorescent protein, Yellow fluorescent protein (YFP) and Cyan Fluorescent Protein (CFP) | Leaf | Syringe (needleless) | EHA105 | pBYKEAM or pBYKEAM2 | Plant expression vectors | SDS-PAGE, Fluorescence Imaging, Western Blotting, ELISA | High level production of monoclonal antibodies | [163] | ||
| Nicotiana tabacum (Tobacco) | Solanaceae | N. tabacum cv. Samsun and Xanthi, and N. benthamiana | Human interferon-γ (hIFN-γ) protein | Leaf | Syringe (needleless) | EHA101, GV3101, and LBA4404 | pGEM-hIFN-γ | Best Agrobacterium strain GV3101 with OD600 of 1.0 and acetosyringone 200 µM at 4 days post agroinfiltration | Reverse Transcription polymerase chain reaction (RT-PCR), qRT-PCR, SDS-PAGE, Western Blotting, ELISA | Bioactive hIFN-γ protein production | [164] |
| Pisum sativum (Pea) | Fabaceae | Pisum sativum and Medicago sativa plants | Salivary gene | Leaf | Syringe and vacuum | AGL-1 | pEAQ-HT-DEST1 | Screening of a range of pea cultivars | Protein extraction and Western-Blotting | Increase aphids fecundity | [165] |
| Solanum lycopersicum (Tomato) and Nicotiana benthamiana (Tobacco) | Solanaceae | MicroTom, a dwarf tomato cultivar | GFP | Leaf | Syringe (needleless) | EHA105 | pCASGFPt (control GFP), pOsAPP1GFP (pGFPTag16) and pOsZF1GFP (pZF1gfp) | Testing of agroinfiltration by expressing GFP fusions of the putative antiphagocytic protein 1 (APP1) (OsAPP, LOC_Os03g56930) and ZOS3-18—C2H2 zinc-finger protein (OsZF1, LOC_Os03g55540) | GUS staining | Subcellular localization of proteins | [166] |
| Floricultural crops | |||||||||||
| Cannabis sativa L. (Hemp) | Cannabaceae | Fedora 17, Felina 32, Ferimon, Futura 75, Santhica 27 and USO31 | Phytoene Desaturase (PDS) and GUS | Plant tissue-mature leaf discs, mature leaf, pollen sacs, anthers, sepals, pollen sac clusters, filaments, pollen grains, nonglandular trichomes, female flowers and pistil | Vacuum | EHA105, LBA4404 and GV3101 | pEarleyGate 101-uidA | Silwett L-770 (0.015%), ascorbic acid (5 mm) and sonication of 30 s followed by a 10-min vacuum treatment | qRT-PCR | Highest GUS expression in the leaf, stem, root tissues, male and female flowers | [167] |
| Eustoma russullianus (Lisianthus) | Gentianaceae | - | GUS | Pollen | Vacuum | LBA4404 | pBI121 | Sucrose 7–15% pH 5.5–7.0 Temperature 20–27 °C | GUS assay, Southern hybridization and RT-PCR | Pollen transformation | [168] |
| Gerbera jemosonii (Gerbera) | Asteraceae | Express and White Grizzly | GUS, GFP, iris-dfr and petunia-f3′5′h | Flower | Syringe and vacuum | GV3101 | pCambia/dfr and pFGC5941 | Vacuum infiltration prove to be the best method | GFP and GUS assay | Change in the anthocyanin pigment | [169] |
| Piper colubrinum Link (Black pepper) | Piperaceae | - | GUS and Serine threonine protein kinase (STPK) gene | Detached Leaf | Vacuum | EHA 105 | pCAMBIA 1305.2 and pHELLSGATE | Higher vacuum up to 400–600 mm Hg increased infiltration transformation efficiency | qRT-PCR | Silencing of STPK gene | [170] |
| Vitis vinifera L. (Grapevine) | Vitaceae | Sugraone, Aleatico, Moscato Giallo and Aglianico | Free GFP and (mRFP1), GFP::HDEL, GAPA1::YFP and b::GFP | Leaf | Syringe (needleless) | LBA4404, GV3101 and AGL1 | pBI121, pBIN-m-gfp5-ER, pAVA554, pRSET-mRFP1, pAVA554 and pGreen 0029 | Combination of sugraone cultivar and the GV3101 showed high gene expression | GFP imaging | Compatibility between Agrobacterium strain and genotype exhibited high transient gene expression | [139] |
| Vitis vinifera (Grapevine) | Vitaceae | Cabernet Sauvignon, Cinsault, Muscat Ottonel and Syrah | GUS, GFP and stilbene synthase | Leaf | Syringe (needleless) and vacuum | C58C1 | pBIN19 and pBINY53 | Presence of additional virulence factors like virG and virE promote infiltration | RT-PCR, GUS staining and Fluorescence microscopy | Vacuum infiltration better than syringe infiltration | [171] |
| Horticultural crops | |||||||||||
| Maesa lanceolata (False assegai) | Primulaceae | - | GFP | Leaf | Syringe | C58, EHA101, EHA105, LBA4404, GV3301, GV2260 and pMP90 | pK7FWGF2 | A. tumefaciens strain LBA4404 at an OD600 = 1.0 in the presence of 100 µM acetosyringone and in the absence of viral suppressor construct | PCR | Saponin production | [172] |
| Malus domestica Borkh-(Apple), Pyrus communis L. (Pear) | Rosaceae | Apple ‘Gala’ and Pear ‘Conference’ | GUS | Leaf | Vacuum | EHA105 | pBBR1MCS-5 | Silwet L-77 at a low concentration (0.002% v/v) | Optimising through 10 different binary plasmids and A. tumefaciens inoculations | Transformation efficiency between 50 and 80% | [173] |
| Leguminous crop | |||||||||||
| Mucuna bracteata | Fabaceae | - | Anti-toxoplasma immunoglobulin | Leaf | Vacuum | GV3101 | pTRAkcHcLcTg130 | High expression in bottom trifoliate leaf at 2 days post-infiltration | Western blotting and ELISA | Transient expression in M. bracteata, was two-fold higher than the model Nicotiana benthamiana plant | [174] |
| Vegetable Crop | |||||||||||
| Spinacia oleracea (Spinach) | Chenopodiaceae | Korean cultivar Sakyechul | GUS | Leaf | Syringe (needleless) and vacuum | EHA105, LBA4404 and GV2260 | pB7WG2D-GUS | Agrobacterium GV2260 strain suspension at OD600 of 1.0 | qRT-PCR | Increased efficiency, duration of gene expression and protein accumulation | [97] |
| Vegetable and model crops | |||||||||||
| Lactuca serriola and L. sativa (Lettuce), Lycopersicon esculentum (Tomato), N. benthamiana (Tobacco) and Arabidopsis thaliana (Arabidopsis) | Asteraceae, Solanaceae and Brassicaceae | Wild lettuce LS102, cultivated lettuce cv. Valmaine and cv. Mariska, tomato-Rio Grande 76R, Arabidopsis-Columbia-0 | GUS | Leaf | Syringe (needleless) | 42 wild strains | pCB301‘empty’, tobacco etch virus-P1/HcPro, turnip mosaic virus-P1/HcPro and P19 from tomato bushy stunt virus | Best Agrobacterium strain C58C1 | GUS assay | High gene expression in lettuce as compared to Nicotiana benthamiana | [88] |
| Nicotiana tobacum (Tobacco), Solanum tuberosum (Potato) and Lactuca sativa (Lettuce) | Solanaceae and Asteraceae | Nicotiana tobacum cv. Xanthi, Solanum tuberosum cv. Agria | Human growth hormone | Leaf | Vacuum | pGV3850 | pBin19 | Time span of infiltration upto 35 min | Western blotting and ELISA | High production of recombinant hGH protein in tobacco and potato as compared to lettuce | [175] |
11. Advantages of Agroinfiltration Method over Other Transient Genetic Transformation Methods
12. CRISPR-Cas9 Based Genome Editing via Agroinfiltration
| Plant | Family | Cultivar/Genotype | Target | Tissue | Stage | Method | Agrobacterium Strain | Promoter | Cas9-Codon Optimised | Detection Assay | Mutation Rate | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Model plants | ||||||||||||
| Arabidopsis thaliana (Arabidopsis) | Brassicaceae | Columbia-0 | PDS | Seedlings | 2-week-old | Syringe (needleless) | GV3101 | CaMV35SPDK and AtU6 | Yes | PCR and Sanger sequencing | 2.7% | [185] |
| Arabidopsis thaliana (Arabidopsis) | Brassicaceae | Transgenic plants | GFP | Leaf | 4-week-old | Syringe (needleless) | C58 and EHA105 | CaMV35S and AtU6 | Yes | Fluorescence confocal microscopy and Sangersequencing | - | [186] |
| Nicotiana benthamiana (Tobacco) | Solanaceae | - | PDS | Leaf | 5-week-old | Syringe (needleless) | GV3101 | CaMV35SPDK and AtU6 | Yes | PCR and Sanger sequencing | 4.8% | [185] |
| - | PDS | Leaf | 3–4 weeks | Syringe (needleless) | AGL1 | CaMV35S and AtU6 | Yes | PCR and restriction enzyme assay | 2.1% | [187] | ||
| - | PDS | Leaf | 3–4 weeks | Syringe | GV3101 | CaMVE35S | - | RT-PCR and Sanger sequencing | 12.7–13.8% | [188] | ||
| - | PDS | Leaf | 3–4 weeks | Syringe | GV3101 | BS3 and uid | Yes | qRT-PCR and Sanger sequencing | - | [189] | ||
| Transgenic KQ334 plant | NbPDS3 and isopentenyl/dimethylallyl diphosphate synthase (NbIspH) genes | Leaf | Six-leaf stage | Syringe | GV3101 | CaMV35S and U6 | Yes | RT-PCR and Sanger sequencing | PDS-85%, IspH-75% | [190] | ||
| - | PDS | Leaf | 3-week-old | Syringe (needleless) | AGL1 | J23119 | - | PCR, Sanger and Illumina sequencing | 3–18% | [191] | ||
| Transgenic plants | Six sites Bean yellow dwarf virus genome—Rep binding site (RBS), hairpin, nonanucleotide sequence and three Rep motifs | Leaf tips | 5-week-old | Syringe | GV3101 | Double 35S promoter and AtU6 or At7SL RNA polymerase III promoter | - | qRT-PCR and Illumina sequencing | 0.03–70.01% | [192] | ||
| - | Xylosyltransferase gene | Leaf | 5–6 weeks | Syringe (needleless) | GV3101 | Nopaline synthase | Yes | PCR, restriction digestion analysis and Sanger sequencing | 12.1%—XT1 and 9.9%—XT2 | [193] | ||
| Nicotiana tabacum (Tobacco) | Solanaceae | Cas9-overexpressing transgenic lines | PDS and proliferating cell nuclear antigen gene (PCNA) genes | Leaf | - | Syringe | GV3101 | CaMV 35S | Yes | T7 endonuclease 1 based assay, restriction digestion analysis and Sanger sequencing | - | [194] |
| - | PDS | Leaf | - | Syringe (needleless) | EHA105 and GV3101 | AtU6 | Yes | PCR and Sanger sequencing | - | [195] | ||
| - | PDS | Leaf | 3–4 weeks | Syringe | GV3101 | 35SPDK | Yes | Flourescent microscopy and PCR analysis | - | [196] | ||
| Wild-type | GFP | Leaf | 4-week-old | Syringe (needleless) | C58 and EHA105 | CaMV35S and AtU6 | Yes | Fluorescence confocal microscopy and Sanger sequencing | - | [186] | ||
| Solanum lycopersicum (Tomato) | Solanaceae | Transgenic plants | Immunity associated genes | Leaf | 4-week-old | Syringe (needleless) | 1D1249 | U6 promoter | - | PCR and Sanger sequencing | 61.5% | [96] |
| Cereal crop | ||||||||||||
| Sorghum bicolor (Sorghum) | Poaceae | Tx430 plants | GFP | Leaf | 3–4 weeks | Syringe (needleless) | GV3101 | CaMV 35S and maize Ubiquitin 1 | Yes | Fluorescence microscopy | - | [184] |
| Horticultural crops | ||||||||||||
| Citrus sinensis (Sweet Orange) | Rutaceae | Valencia cultivar | PDS | Leaf | Three-year-old | Syringe | - | CaMV 35S | - | PCR and Sanger sequencing | 3.2–3.9% | [197] |
| Citrus paradisi (Grapefruit) | Rutaceae | Wild type Duncan and transgenic plants | Canker susceptibility gene (CsLOB1) | Leaf | - | Syringe (needleless) | EHA105 | Cassava vein mosaic virus promoter and CaMV 35S promoter | - | Illumina sequencing | 3.58–88.78% | [198] |
| Fragaria × ananassa (Strawberry) | Rosaceae | Fragaria vesca (cv. Reine des Vallées) and F. × ananassa Duch. (cv. Camarosa) | Tomato MADS box gene6 | Fruit | Green stage | - | AGL-0 | 35SCaMV and AtU6-26 | - | PCR and Sanger sequencing | - | [199] |
| Floricultural crop | ||||||||||||
| Papaver somniferum L. (Opium poppy) | Papaveraceae | P. somniferum (cv. Ofis-95) | 3′-hydroxyl-N-methylcoclaurine 4′-O-methyltransferase (4′OMT2) | Leaf | - | Syringe (needleless) | EHA105 and GV3101 | AtU6 | Yes | PCR and Sanger sequencing | - | [195] |
| Leguminous crop | ||||||||||||
| Vigna unguiculata [L.] Walp. (Cowpea) | Fabaceae | Transgenic plants | Meiosis genes i.e., SPO11-1, REC8, and OSD1 | Detached leaflets | 3–4 weeks | Syringe (with and without needle) | AGL-1 | Arabidopsis ubiquitin 3, RPS5a and AtU6-26 | - | PCR and Illumina sequencing | 1% | [144] |
| Vegetable crops | ||||||||||||
| Brassica oleracea var. capitata f. Rubra (Purple cabbage) | Brassicaceae | Rebecca F1 and Huzaro F1 | Centromere-specific histone H3 | Leaf | 6-week-old | Syringe | GV3101 | CaMV35S and AtU6 | Yes | PCR and Illumina HiSeq sequencing | 0.07–14.42% | [200] |
| Dioscorea alata (Yam) | Dioscoreaceae | Transgenic plants | PDS | Leaf | 2 months old | Syringe | EHA105 and LBA4404 | DaU6 promoter, maize ubiquiti and CaMV35S | - | PCR and Sanger sequencing | 83.3% | [201] |
13. Modifications of Agroinfiltration-Based Genome Editing
14. Other Applications in Plants
| Plant | Target | Family | Material | Promoter | Agrobacterium Strain | Method | Outcome | Reference |
|---|---|---|---|---|---|---|---|---|
| Model plants | ||||||||
| Medicago truncatula (Barrel clover) | LEGUME ANTHOCYANIN PRODUCTION 1 (LAP1) transcription factor | Fabaceae | Medicago truncatula cv. R108 | CaMV35S | GV3101 | Syringe | Accumulation of anthocyanin pigment | [209] |
| Nicotiana benthamiana (Tobacco) | C5-1 murine antibody | Solanaceae | Seeds obtained from National tobacco germplasm | CaMV 35S | AGL1 | Syringe | Recombinant protein production at lab scale | [211] |
| GFP transgene | Solanaceae | - | CaMV35S | - | Syringe | Systemic silencing of a GFP transgene | [214] | |
| epiGFP (without GFP integration) | Solanaceae | Stably integrated GFP transgene (intGFP) transgenic plants | CaMV35S | - | Syringe | Systemic silencing through interaction between epiGFP and intGFP | [215] | |
| Ubiquitin ligase-associated protein SGT1 | Solanaceae | Transgenic plants | CaMV35S | - | Syringe | Proof of SGT1 that is required for host and nonhost disease resistance in plants | [222] | |
| mGFP5-er, Bt Cry1Ac, and BoPI transgene genes | Solanaceae | - | CaMV 35S | GV3850 | Syringe | Ease the detection of candidate insect resistance transgenes | [223] | |
| Chimaeric human β1,4-galactosyltransferase | Solanaceae | Wild-type | CaMV 35S, Rubisco, plastocyanin | R612, R610, R621, R622 and 35SHcPro | Vacuum | High-yield antibodies production with human-like N-glycans | [230] | |
| Haemagglutinin gene | Solanaceae | - | Plastocyanin, chimeric double 35S | AGL1 | Vacuum | Production of an influenza vaccine | [235] | |
| E3 ligase Constitutive photomorphogenic1 (COP1) and its substrate HY5 | Solanaceae | Wild-type | - | EHA105 and ABI | Syringe | Detection of protein ubiquitination | [232] | |
| Foot-and-mouth disease (FMD) virus P1-polyprotein (P1) and VP1 (viral capsid protein 1) and E. coli glutathione reductase (GOR) | Solanaceae | - | CaMV35S | LBA4404 | Syringe | Recombinant VP1 protein degradation | [233] | |
| Nicotiana species (Tobacco) | Cauliflower mosaic virus Gene VI | Solanaceae | N. edwardsonii and N. clevelandii | CaMV35S | C58 | Syringe (needleless) | Identification of gene VI protein elicitor | [219] |
| Nicotiana sylvestris (Tobacco) | Class I chitinase A gene CHN48 transgene | Solanaceae | Wild type and transgenic plants | CaMV35S | - | Syringe | Transgene silencing | [213] |
| Nicotiana tabacum (Tobacco) | N gene | Solanaceae | Samsun NN and nn plants | - | - | Syringe | Identification of the Tobacco Mosaic Virus elicitor | [218] |
| Avr9 and Avr4-Tobacco Cf-9 and Cf-4-tomato | Solanaceae | N. tabacum cv. Petite Havana and transgenic tobacco lines | CaMV35S | MOG101 | Syringe (needleless) | Co-expression of the Avr4/Cf-4 gene pair confer resistance | [220] | |
| Rx2, AC15 | Solanaceae | Tetraploid potato cultivars BZURA (Rx2 genotype) and three susceptible potato accessions (rx genotype) | CaMV35S and Rx1 | C58C1 | Isolation of Rx resistance genes | [221] | ||
| Stress-responsive as-1 and heat shock elements, yeast GAL4 transactivation system, two promoters of pathogenesis-related genes as well as a heat shock promoter | Solanaceae | Nicotiana tabacum var. Xanthi nc | EHA 105 | Syringe | Identification of the cis-regulatory regions in promoters | [224] | ||
| Human lactoferrin | Solanaceae | - | MPr1163 and CaMV E-35S | LBA4404 | Syringe | Efficient use of chimeric promoter MPr1163 for the expression of heterologous protein | [225] | |
| Collagen and chimeric P4H genes | Solanaceae | - | L3, 1287 | A1286 and A1284 | Vacuum | Improvement in expression of collagen | [226] | |
| Pseudomonas syringae pv. phaseolicola harpin (HrpZPsph) gene | Solanaceae | N. tabacum cv. W38 TetR, N. tabacum cv. Xanthi, and N. benthamiana | CaMV35S | C58C1 | - | Detection of plant cellular receptor(s) for harpin is extracellular | [227] | |
| T84.66/GS8 diabody | Solanaceae | N. tabacum cv. Petit Havana SR1 | CaMV35S | GV3101 | Vacuum | Production of A carcinoembryonic antigen-specific diabody | [228] | |
| Human chorionic gonadotropin (hCG) | Solanaceae | Nicotiana tabacum cv. Petite Havana SR1 | - | GV3101 | Vacuum | Production of recombinant antibodies against hCG | [229] | |
| Foot and Mouth Disease Virus (FMDV) Coat Protein | Solanaceae | - | CaMV35S | GV3101 | Syringe | Production of recombinant antigen of FMD | [231] | |
| Glycine max (Soybean) | Coatomer subunit alpha (COPA) and aquaporin 9 (AQ9) genes | Fabaceae | - | CaMV35S | EHA 105 | Mechanical abrasion using carborundum | RNA interference against Tetranychus urticae | [217] |
| Floricultural crops | ||||||||
| Vitis vinifera L. (Grapevine) | Grapevine gene VvPGIP1 | Vitaceae | Cabernet franc | CaMV35S | GV3101 | Vacuum | Transient gene silencing | [216] |
| Antirrhinum majus (Snapdragon) | AS1 and 4′CGT genes | Plantaginaceae | - | - | LBA4404 | Syringe | Conversion from white to pale yellow petals | [236] |
| Horticultural crop | ||||||||
| Fragaria × ananassa (Strawberry) | Chalcone synthase gene | Rosaceae | F. × ananassa cv. Elsanta | CaMV35 S | AGL0 | Syringe | Gene silencing | [122] |
| Vegetable crops | ||||||||
| Raphanus sativus L. (Radish) | Staphylococcal enterotoxin B (SEB) genes | Brassicaceae | CaMV35S | LBA4404 | Syringe | Production of Leaf-Encapsulated Vaccines | [237] | |
| Solanum melongena L. (Eggplant) | Hydroxycinnamoyl CoA-quinate transferase gene | Solanaceae | p19 protein of Tomato bushy stunt virus (native promoter) | GV3101 | Syringe (needle) | Improvement in chlorogenic content | [238] | |
15. Limitations
16. Bio-Safety and Commercialization Aspects
17. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adli, M. The CRISPR tool kit for genome editing and beyond. Nat. Commun. 2018, 9, 1911. [Google Scholar] [CrossRef]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and applications of CRISPR-Cas9 for genome engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zlobin, N.E.; Lebedeva, M.V.; Taranov, V.V. CRISPR/Cas9 genome editing through in planta transformation. Crit. Rev. Biotechnol. 2020, 40, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Anami, S.; Njuguna, E.; Coussens, G.; Aesaert, S.; Van Lijsebettens, M. Higher plant transformation: Principles and molecular tools. Int. J. Dev. Biol. 2013, 57, 483–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzfira, T.; Citovsky, V. Agrobacterium-mediated genetic transformation of plants: Biology and biotechnology. Curr. Opin. Biotechnol. 2006, 17, 147–154. [Google Scholar] [CrossRef]
- Kapila, J.; De Rycke, R.; van Montagu, M.; Angenon, G. An Agrobacterium-mediated transient gene expression system for intact leaves. Plant Sci. 1997, 122, 101–108. [Google Scholar] [CrossRef]
- Klein, T.M.; Wolf, E.D.; Sanford, J.C. High-velocity microprojectiles for delivering nucleic acids into living cells. Nature 1987, 327, 70–73. [Google Scholar] [CrossRef]
- Paszkowski, J.; Shillito, R.D.; Saul, M.; Mandák, V.; Hohn, T.; Hohn, B.; Potrykus, I. Direct gene transfer to plants. EMBO J. 1984, 3, 2717–2722. [Google Scholar] [CrossRef]
- D’Halluin, K.; Bonne, E.; Bossut, M.; De Beuckeleer, M.; Leemans, J. Transgenic maize plants by tissue electroporation. Plant Cell 1992, 4, 1495–1505. [Google Scholar]
- De la Peña, A.; Lörz, H.; Schell, J. Transgenic rye plants obtained by injecting DNA into young floral tillers. Nature 1987, 325, 274–276. [Google Scholar] [CrossRef]
- Krens, F.A.; Molendijk, L.; Wullems, G.J.; Schilperoort, R.A. In vitro transformation of plant protoplasts with Ti-plasmid DNA. Nature 1982, 296, 72–74. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Sandhya, D.; Jogam, P.; Allini, V.R.; Abbagani, S.; Alok, A. The present and potential future methods for delivering CRISPR/Cas9 components in plants. J. Genet. Eng. Biotechnol. 2020, 18, 1–11. [Google Scholar] [CrossRef]
- Guidarelli, M.; Baraldi, E. Transient transformation meets gene function discovery: The strawberry fruit case. Front. Plant Sci. 2015, 6, 444. [Google Scholar] [CrossRef] [Green Version]
- Twyman, R.M.; Kohli, A.; Stoger, E.; Christou, P. Foreign DNA: Integration and Expression in Transgenic Plants. In Genetic Engineering: Principles and Methods; Setlow, J.K., Ed.; Springer: Boston, MA, USA, 2002; Volume 24, pp. 107–136. [Google Scholar]
- Chhikara, S.; Chaudhary, D.; Yadav, M.; Sainger, M.; Jaiwal, P.K. A non-tissue culture approach for developing transgenic Brassica juncea L. plants with Agrobacterium tumefaciens. In Vitro Cell Dev. Biol. Plant 2012, 48, 7–14. [Google Scholar] [CrossRef]
- Jones, H.D. Future of breeding by genome editing is in the hands of regulators. GM Crops Food 2015, 6, 223–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashraf, A.; Rahman, A. CRISPR/Cas9 and Biosafety Issues. In Conference: Workshop Hand Book for 1st National Conference on CRISPR-Cas9 Genome Editing Technology; 2017; pp. 1–10. Available online: https://www.researchgate.net/publication/333557456_CRISPRCas9_and_Biosafety_Issues (accessed on 10 September 2021).
- Lee, M.W.; Yang, Y. Transient Expression Assay by Agroinfiltration of Leaves. Methods Mol. Biol. 2006, 323, 225–229. [Google Scholar] [CrossRef]
- González, M.N.; Massa, G.A.; Andersson, M.; Andrea, C.; Oneto, D.; Turesson, H.; Storani, L.; Olsson, N.; Fält, A.S.; Hofvander, P.; et al. Comparative potato genome editing: Agrobacterium tumefaciens-mediated transformation and protoplasts transfection delivery of CRISPR/Cas9 components directed to StPPO2 gene. Plant Cell Tiss. Organ Cult. 2021, 145, 291–305. [Google Scholar] [CrossRef]
- Zhang, Z.; Mao, Y.; Ha, S.; Liu, W.; Botella, J.R.; Zhu, J.K. A multiplex CRISPR/Cas9 platform for fast and efficient editing of multiple genes in Arabidopsis. Plant Cell Rep. 2016, 35, 1519–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Choi, J.; Won, K.H. A stable DNA-free screening system for CRISPR/RNPs-mediated gene editing in hot and sweet cultivars of Capsicum annuum. BMC Plant Biol. 2020, 20, 449. [Google Scholar] [CrossRef]
- Malzahn, A.; Lowder, L.; Qi, Y. Plant genome editing with TALEN and CRISPR. Cell Biosci. 2017, 7, 21. [Google Scholar] [CrossRef] [Green Version]
- Donini, M.; Marusic, C. Current state-of-the-art in plant-based antibody production systems. Biotechnol. Lett. 2019, 41, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liang, Z.; Zong, Y.; Wang, Y.; Liu, J.; Chen, K.; Qiu, J.-L.; Gao, C. Efficient and transgene-free genome editing in wheat through transient expression of CRISPR/Cas9 DNA or RNA. Nat. Commun. 2016, 7, 12617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Z.; Chen, K.; Li, T.; Zhang, Y.; Wang, Y.; Zhao, Q.; Liu, J.; Zhang, H.; Liu, C.; Ran, Y.; et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nat. Commun. 2017, 8, 14261. [Google Scholar] [CrossRef] [PubMed]
- Sheludko, Y.V. Agrobacterium-mediated transient expression as an approach to production of recombinant proteins in plants. Recent Pat. Biotechnol. 2008, 2, 198–208. [Google Scholar] [CrossRef]
- Krenek, P.; Samajova, O.; Luptovciak, I.; Doskocilova, A.; Komis, G.; Samaj, J. Transient plant transformation mediated by Agrobacterium tumefaciens: Principles, methods and applications. Biotechnol. Adv. 2015, 33, 1024–1042. [Google Scholar] [CrossRef]
- Lacroix, B.; Citovsky, V. The roles of bacterial and host plant factors in Agrobacterium mediated genetic transformation. Int. J. Dev. Biol. 2013, 57, 467–481. [Google Scholar] [CrossRef]
- Chen, Q.; Lai, H.; Hurtado, J.; Stahnke, J.; Leuzinger, K.; Dent, M. Agroinfiltration as an effective and scalable strategy of gene delivery for production of pharmaceutical proteins. Adv. Tech. Biol. Med. 2013, 1, 103. [Google Scholar] [CrossRef] [Green Version]
- Tyurin, A.A.; Suhorukova, A.V.; Kabardaeva, K.V.; Goldenkova-Pavlova, I.V. Transient gene expression is an effective experimental tool for the research into the fine mechanisms of plant gene function: Advantages, limitations, and solutions. Plants 2020, 9, 1187. [Google Scholar] [CrossRef]
- Ramkumar, T.R.; Lenka, S.K.; Arya, S.; Bansal, K.C. A Short History and Perspectives on Plant Genetic Transformation. In Biolistic DNA Delivery in Plants; Methods in Molecular Biology; Rustgi, S., Luo, H., Eds.; Humana: New York, NY, USA, 2020; p. 2124. [Google Scholar] [CrossRef]
- Low, L.Y.; Yang, S.K.; Andrew, K.D.X.; Ong-Abdullah, J.; Tan, N.P.; Lai, K.S. Transgenic plants: Gene constructs, vector and transformation method. In New Visions in Plant Science; Çelik, Ö., Ed.; Intech Open: London, UK, 2018; pp. 41–61. [Google Scholar]
- Zambryski, P. Basic processes underlying Agrobacterium-mediated DNA transfer to plant cells. Annu. Rev. Genet. 1988, 22, 1–30. [Google Scholar] [CrossRef]
- Christou, P.; Ford, T.L.; Kofron, M. Rice genetic engineering: A review. Trends Biotechnol. 1992, 10, 239–246. [Google Scholar] [CrossRef]
- Crossway, A.; Oakes, J.V.; Irvine, J.M.; Ward, B.; Knauf, V.C.; Shewmaker, C.K. Integration of foreign DNA following microinjection of tobacco mesophyll protoplasts. Mol. Gen. Genet. 1986, 202, 179–185. [Google Scholar] [CrossRef]
- Negrutiu, I.; Shillito, R.D.; Potrykus, I.; Biasini, G.; Sala, F. Hybrid genes in the analysis of transformation conditions. I. Setting up a simple method for direct gene transfer in plant protoplasts. Plant Mol. Biol. 1987, 8, 363–373. [Google Scholar] [CrossRef]
- Datta, S.K.; Peterhans, A.; Datta, K.; Potrykus, I. Genetically engineered fertile Indica-rice plants recovered from protoplasts. Nat. Biotechnol. 1990, 8, 736–740. [Google Scholar] [CrossRef]
- Shillito, R.D.; Saul, M.W.; Paszkowski, J.; Muller, M.; Potrykus, I. High efficiency direct gene transfer to plants. Nat. Biotechnol. 1985, 3, 1099–1103. [Google Scholar] [CrossRef]
- Fromm, M.E.; Taylor, L.P.; Walbot, V. Stable transformation of maize after gene-transfer by electroporation. Nature 1986, 319, 791–793. [Google Scholar] [CrossRef] [PubMed]
- Frame, B.R.; Drayton, P.R.; Bagnall, S.V.; Lewnau, C.J.; Bullock, W.P.; Wilson, H.M.; Dunwell, J.M.; Thompson, J.A.; Wang, K. Production of fertile transgenic maize plants by silicon-carbide whisker-mediated transformation. Plant J. 1994, 6, 941–948. [Google Scholar] [CrossRef]
- Koncz, C.; Németh, K.; Rédei, G.P.; Schell, J. Homologous recombination and gene silencing in plants. In Homology Recognition during T-DNA Integration into the Plant Genome; Springer: Berlin/Heidelberg, Germany, 1994; pp. 167–189. [Google Scholar]
- Pawlowski, W.P.; Somers, D.A. Transgene inheritance in plants genetically engineered by microprojectile bombardment. Mol. Biotechnol. 1996, 6, 17–30. [Google Scholar] [CrossRef]
- Hansen, G.; Shillito, R.D.; Chilton, M.D. T-strand integration in maize protoplasts after codelivery of a T-DNA substrate and virulence genes. Proc. Natl. Acad. Sci. USA 1997, 94, 11726–11730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enríquez-Obregón, G.A.; Vázquez-Padrón, R.I.; Prieto-Samsonov, D.L.; De la Riva, G.A.; Selman-Housein, G. Herbicide-resistant sugarcane (Saccharum officinarum L.) plants by Agrobacterium mediated transformation. Planta 1998, 206, 20–27. [Google Scholar] [CrossRef]
- Shou, H.; Frame, B.R.; Whitham, S.A.; Wang, K. Assessment of transgenic maize events produced by particle bombardment or Agrobacterium mediated transformation. Mol. Breed. 2004, 13, 201–208. [Google Scholar] [CrossRef]
- Travella, S.; Ross, S.M.; Harden, J.; Everett, C.; Snape, J.W.; Harwood, W.A. A comparison of transgenic barley lines produced by particle bombardment and Agrobacterium mediated techniques. Plant Cell Rep. 2005, 23, 780–789. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yin, X.; Yang, A.; Li, G.; Zhang, J. Stability of inheritance of transgenes in maize (Zea mays L.) lines produced using different transformation methods. Euphytica 2005, 144, 11–22. [Google Scholar] [CrossRef]
- Gao, C.; Long, D.; Lenk, I.; Nielsen, K.K. Comparative analysis of transgenic tall fescue (Festuca arundinacea Schreb.) plants obtained by Agrobacterium mediated transformation and particle bombardment. Plant Cell Rep. 2008, 27, 1601–1609. [Google Scholar] [CrossRef]
- Hwang, H.H.; Yu, M.; Lai, E.M. Agrobacterium-mediated plant transformation: Biology and applications. Arab. Book 2017, 15, e0186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntosh, K.B.; Hulm, J.L.; Young, L.W.; Bonham-Smith, P.C. A rapid Agrobacterium-mediated Arabidopsis thaliana transient assay system. Plant Mol. Biol. Rep. 2004, 22, 53–61. [Google Scholar] [CrossRef]
- Sheng, J.; Citovsky, V. Agrobacterium-plant cell interaction: Have virulence proteins, will travel. Plant Cell 1996, 8, 1699–1710. [Google Scholar]
- Horsch, R.B.; Fry, J.E.; Hoffmann, N.L.; Eïchholtz, D.; Rogers, S.G.; Fraley, R.T. A simple and general method for transferring genes into plants. Science 1985, 227, 1229–1231. [Google Scholar]
- Chilton, M.D.; Drummond, M.H.; Merlo, D.J.; Sciaky, D.; Montoya, A.L.; Gordon, M.P.; Nester, E.W. Stable incorporation of plasmid DNA into higher plant cells: The molecular basis of crown gall tumorigenesis. Cell 1977, 11, 263–271. [Google Scholar] [CrossRef]
- De Cleene, M.; De Ley, J. The host range of crown gall. Bot. Rev. 1976, 42, 389–466. [Google Scholar] [CrossRef]
- Li, W.; Guo, G.; Zheng, G. Agrobacterium-mediated transformation: State of the art and future prospect. Chin. Sci. Bull. 2000, 45, 1537–1546. [Google Scholar] [CrossRef]
- Bundock, P.; den Dulk-Ras, A.; Beijersbergen, A.; Hooykaas, P.J. Trans-kingdom T-DNA transfer from Agrobacterium tumefaciens to Saccharomyces cerevisiae. EMBO J. 1995, 14, 3206–3214. [Google Scholar] [CrossRef] [PubMed]
- Bundock, P.; Mroczek, K.; Winkler, A.A.; Steensma, H.Y.; Hooykaas, P.J.J. T-DNA from Agrobacterium tumefaciens as an efficient tool for gene targeting in Kluyveromyces lactis. Mol. Gen. Genet. 1999, 261, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Piers, K.L.; Heath, J.D.; Liang, X.; Stephens, K.M.; Nester, E.W. Agrobacterium tumefaciens mediated transformation of yeast. Proc. Natl. Acad. Sci. USA 1996, 93, 1613–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Groot, M.J.A.; Bundock, P.; Hooykaas, P.J.J.; Beijersbergen, A.G.M. Agrobacterium tumefaciens mediated transformation of filamentous fungi. Nat. Biotechnol. 1998, 16, 839–842. [Google Scholar] [CrossRef]
- Kunik, T.; Tzfira, T.; Kapulnik, Y.; Gafni, Y.; Dingwall, C.; Citovsky, V. Genetic transformation of HeLa cells by Agrobacterium. Proc. Natl. Acad. Sci. USA 2001, 98, 1871–1876. [Google Scholar] [CrossRef]
- Kelly, B.A.; Kado, C.I. Agrobacterium mediated T-DNA transfer and integration into the chromosome of Streptomyces lividans. Mol. Plant Pathol. 2002, 3, 125–134. [Google Scholar] [CrossRef]
- Hooykaas, P.J.J. Transformation mediated by Agrobacterium tumefaciens. In Advances in Fungal Biotechnology for Industry, Agriculture, and Medicine; Tkacz, J.S., Lange, L., Eds.; Springer: Boston, MA, USA, 2004; pp. 41–65. [Google Scholar]
- Kumar, S.V.; Misquitta, R.W.; Reddy, V.S.; Rao, B.J.; Rajam, M.V. Genetic transformation of the green alga—Chlamydomonas reinhardtii by Agrobacterium tumefaciens. Plant Sci. 2004, 166, 731–738. [Google Scholar] [CrossRef]
- Pelczar, P.; Kalck, V.; Gomez, D.; Hohn, B. Agrobacterium proteins VirD2 and VirE2 mediate precise integration of synthetic TDNA complexes in mammalian cells. EMBO Rep. 2004, 5, 632–637. [Google Scholar] [CrossRef] [Green Version]
- Michielse, C.B.; Hooykaas, P.J.J.; van den Hondel, C.A.; Ram, A.F. Agrobacterium mediated transformation as a tool for functional genomics in fungi. Curr. Genet. 2005, 48, 1–17. [Google Scholar] [CrossRef]
- Bulgakov, V.P.; Kiselev, K.V.; Yakovlev, K.V.; Zhuravlev, Y.N.; Gontcharov, A.A.; Odintsova, N.A. Agrobacterium mediated transformation of sea urchin embryos. Biotechnol. J. 2006, 1, 454–461. [Google Scholar] [CrossRef]
- Lacroix, B.; Li, J.; Tzfira, T.; Citovsky, V. Will you let me use your nucleus? How Agrobacterium gets its T-DNA expressed in the host plant cell. Can. J. Physiol. Pharmacol. 2006, 84, 333–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomashow, M.; Panagopoulos, C.; Gordon, M.; Nester, E.W. Host range of Agrobacterium tumefaciens is determined by the Ti plasmid. Nature 1980, 283, 794–796. [Google Scholar] [CrossRef]
- Herrera-Estrella, L.; Simpson, J.; Martinez-Trujillo, M. Transgenic plants: An historical perspective. Methods Mol. Biol. 2005, 286, 3–32. [Google Scholar] [PubMed]
- Draper, J.; Scott, R.; Hamill, J. Transformation of dicotyledonous plant cells using the Ti plasmid of Agrobacterium tumefaciens and the Ri plasmid of A. rhizogenes. In Plant Genetic Transformation and Gene Expression: A Laboratory Manual; Draper, J., Scott, R., Armitage, P., Walden, R., Eds.; Blackwell Scientific Publishers: Oxford, UK, 1988; pp. 69–160. [Google Scholar]
- Tinland, B. The integration of T-DNA into plant genomes. Trends Plant Sci. 1996, 1, 178–184. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Sood, P.; Citovsky, V. The roles of plant phenolics in defence and communication during Agrobacterium and Rhizobium infection. Mol. Plant Pathol. 2010, 11, 705–719. [Google Scholar] [CrossRef] [PubMed]
- Gelvin, S.B. Plant proteins involved in Agrobacterium mediated genetic transformation. Annu. Rev. Phytopathol. 2010, 48, 45–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelvin, S.B. Traversing the cell: Agrobacterium T-DNA’s journey to the host genome. Front. Plant Sci. 2012, 3, 52. [Google Scholar] [CrossRef] [Green Version]
- Pitzschke, A. Agrobacterium infection and plant defense-transformation success hangs by a thread. Front. Plant Sci. 2013, 4, 519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christie, P.J.; Whitaker, N.; González-Rivera, C. Mechanism and structure of the bacterial type IV secretion systems. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 1578–1591. [Google Scholar] [CrossRef] [Green Version]
- Hwang, H.H.; Liu, Y.T.; Huang, S.C.; Tung, C.Y.; Huang, F.C.; Tsai, Y.L.; Cheng, T.F.; Lai, E.M. Overexpression of the HspL promotes Agrobacterium tumefaciens virulence in Arabidopsis under heat shock conditions. Phytopathology 2015, 105, 160–168. [Google Scholar] [CrossRef] [Green Version]
- Nonaka, S.; Someya, T.; Kadota, Y.; Nakamura, K.; Ezura, H. Super-Agrobacterium ver. 4: Improving the transformation frequencies and genetic engineering possibilities for crop plants. Front. Plant Sci. 2019, 10, 1204. [Google Scholar] [CrossRef]
- Hoshikawa, K.; Fujita, S.; Renhu, N.; Ezura, K.; Yamamoto, T.; Nonaka, S.; Ezura, H.; Miura, K. Efficient transient protein expression in tomato cultivars and wild species using agroinfiltration-mediated high expression system. Plant Cell Rep. 2019, 38, 75–84. [Google Scholar] [CrossRef]
- Knoch, E.; Sugawara, S.; Mori, T.; Poulsen, C.; Fukushima, A.; Harholt, J.; Fujimoto, Y.; Umemoto, N.; Saito, K. Third DWF1 paralog in Solanaceae, sterol ∆24-isomerase, branches withanolide biosynthesis from the general phytosterol pathway. Proc. Natl. Acad. Sci. USA 2019, 115, E8096–E8103. [Google Scholar] [CrossRef] [Green Version]
- Bent, A. Arabidopsis thaliana Floral Dip Transformation Method. In Agrobacterium Protocols; Methods in Molecular, Biology; Wang, K., Ed.; Humana Press: New York, NY, USA, 2006; Volume 343. [Google Scholar]
- Gelvin, S.B. Agrobacterium Transformation of Arabidopsis thaliana Roots. In Agrobacterium Protocols; Methods in Molecular, Biology; Wang, K., Ed.; Humana Press: New York, NY, USA, 2006; Volume 343. [Google Scholar]
- Tague, B.W.; Mantis, J. In Planta Agrobacterium-Mediated transformation by vacuum infiltration. In Arabidopsis Protocols; Methods in Molecular, Biology™; Salinas, J., Sanchez-Serrano, J.J., Eds.; Humana Press: New York, NY, USA, 2006; Volume 323. [Google Scholar]
- Rivero, L.; Scholl, R.; Holomuzki, N.; Crist, D.; Grotewold, E.; Brkljacic, J. Handling Arabidopsis Plants: Growth, Preservation of Seeds, Transformation, and Genetic Crosses. In Arabidopsis Protocols; Methods in Molecular Biology (Methods and, Protocols); Sanchez-Serrano, J., Salinas, J., Eds.; Humana Press: Totowa, NJ, USA, 2014. [Google Scholar]
- Wroblewski, T.; Tomczak, A.; Michelmore, R. Optimization of Agrobacterium mediated transient assays of gene expression in lettuce, tomato and Arabidopsis. Plant Biotechnol. J. 2005, 3, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Marion, J.; Bach, L.; Bellec, Y.; Meyer, C.; Gissot, L.; Faure, J.D. Systematic analysis of protein subcellular localization and interaction using high-throughput transient transformation of Arabidopsis seedlings. Plant J. 2008, 56, 169–179. [Google Scholar] [CrossRef]
- Jones, H.D.; Doherty, A.; Sparks, C.A. Transient transformation of plants. Methods Mol. Biol. 2009, 513, 131–152. [Google Scholar]
- Kim, M.J.; Baek, K.; Park, C.M. Optimization of conditions for transient Agrobacterium mediated gene expression assays in Arabidopsis. Plant Cell Rep. 2009, 28, 1159–1167. [Google Scholar] [CrossRef] [PubMed]
- Li, J.F.; Park, E.; von Arnim, A.G.; Nebenfuhr, A. The FAST technique: A simplified Agrobacterium-based transformation method for transient gene expression analysis in seedlings of Arabidopsis and other plant species. Plant Methods 2009, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, K.; Qi, Y.; Nguyen, L.V.; Bethke, G.; Tsuda, Y.; Glazebrook, J.; Katagiri, F. An efficient Agrobacterium-mediated transient transformation of Arabidopsis. Plant J. 2012, 69, 713–719. [Google Scholar] [CrossRef]
- Wu, H.Y.; Liu, K.H.; Wang, Y.C.; Wu, J.F.; Chiu, W.L.; Chen, C.Y.; Wu, S.H.; Sheen, J.; Lai, E.M. AGROBEST: An efficient Agrobacterium mediated transient expression method for versatile gene function analyses in Arabidopsis seedlings. Plant Methods 2014, 10, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, B.J.; Gardner, R.C. Localized transient expression of GUS in leaf discs following cocultivation with Agrobacterium. Plant Mol. Biol. 1989, 14, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Roberts, H.M.; Eck, J.V.; Martin, G.B. Generation and molecular characterization of CRISPR/Cas9-induced mutations in 63 immunity-associated genes in tomato reveals specificity and a range of gene modifications. Front. Plant Sci. 2020, 11, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, D.V.; Pamplona, R.S.; Kim, J.; Oh, Y.K.; Cho, S.K.; Ahn, J.; Yang, S.W.; Riu, K.Z.; Boo, K.H. Optimization of Agrobacterium-mediated transient expression of heterologous genes in spinach. Plant Biotechnol. Rep. 2017, 11, 397–405. [Google Scholar] [CrossRef]
- Zhao, H.; Tan, Z.; Wen, X.; Wang, Y. An improved syringe agroinfiltration protocol to enhance transformation efficiency by combinative use of 5-azacytidine, ascorbate acid and tween-20. Plants 2017, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- Heenatigala, P.P.M.; Yang, J.J.; Bishopp, A.; Sun, Z.L.; Li, G.J.; Kumar, S.; Hu, S.; Wu, Z.; Lin, W.; Yao, L.; et al. Development of efficient protocols for stable and transient gene transformation for Wolffia globosa using Agrobacterium. Front. Chem. 2018, 6, 227. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.; Li, Z.; Wang, S. Production of bioactive recombinant reteplase by virus-based transient expression system in Nicotiana benthamiana. Front. Plant Sci. 2019, 10, 1225. [Google Scholar] [CrossRef] [PubMed]
- Garabagi, F.; McLean, M.D.; Hall, J.C. Transient and stable expression of antibodies in Nicotiana species. Methods Mol. Biol. 2012, 907, 389–408. [Google Scholar]
- Sánchez-Álvarez, A.; Ruíz-López, N.; Moreno-Pérez, A.J.; Martínez-Force, E.; Garcés, R.; Salas, J.J. Agrobacterium-mediated transient gene expression in developing Ricinus communis seeds: A first step in making the castor oil plant a chemical biofactory. Front. Plant Sci. 2019, 10, 1410. [Google Scholar] [CrossRef] [PubMed]
- Sainsbury, F.; Lomonossoff, G.P. Transient expressions of synthetic biology in plants. Curr. Opin. Plant Biol. 2014, 19, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osakabe, Y.; Osakabe, K. Genome editing to improve abiotic stress responses in plants. Prog. Mol. Biol. Transl. Sci. 2017, 149, 99–109. [Google Scholar] [PubMed]
- Gleba, Y.; Klimyu, V.; Marillonnet, S. Magnifection—A new platform for expressing recombinant vaccines in plants. Vaccine 2005, 23, 2047–2048. [Google Scholar] [CrossRef] [PubMed]
- Gleba, Y.; Klimyuk, V.; Marillonnet, S. Viral vectors for the expression of proteins in plants. Curr. Opin. Biotechnol. 2007, 18, 134–141. [Google Scholar] [CrossRef]
- Potrykus, I. Gene transfer to plants: Assessment of published approaches and results. Annu. Rev. Plant Biol. 1991, 42, 205–225. [Google Scholar] [CrossRef]
- Dai, S.; Zheng, P.; Marmey, P.; Zhang, S.; Tian, W.; Chen, S.; Beachy, R.N.; Fauquet, C. Comparative analysis of transgenic rice plants obtained by Agrobacterium mediated transformation and particle bombardment. Mol. Breed. 2001, 7, 25–33. [Google Scholar] [CrossRef]
- Zheng, L.; Liu, G.; Meng, X.; Li, Y.; Wang, Y. A versatile Agrobacterium-mediated transient gene expression system for herbaceous plants and trees. Biochem. Genet. 2012, 50, 761–769. [Google Scholar] [CrossRef]
- Burman, N.; Chandran, D.; Khurana, J.P. A rapid and highly efficient method for transient gene expression in rice plants. Front. Plant Sci. 2020, 11, 584011. [Google Scholar] [CrossRef]
- Fischer, R.; Vaquero-Martin, C.; Sack, M.; Drossard, J.; Emans, N.; Commandeur, U. Towards molecular farming in the future: Transient protein expression in plants. Biotechnol. Appl. Biochem. 1999, 30, 113–116. [Google Scholar]
- Sparkes, I.A.; Runions, J.; Kearns, A.; Hawes, C. Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat. Protoc. 2006, 1, 2019–2025. [Google Scholar] [CrossRef]
- Zhou, D.D.; Yu, J.N. The progress of establishing transient expression system in plant cell. Chin. Agric. Sci. Bull. 2013, 29, 151–156. [Google Scholar]
- Liu, S.; Ma, J.; Liu, H.; Guo, Y.; Li, W.; Niu, S. An efficient system for Agrobacterium-mediated transient transformation in Pinus tabuliformis. Plant Methods 2020, 16, 52. [Google Scholar] [CrossRef] [PubMed]
- Xia, P.; Hu, W.; Liang, T.; Yang, D.; Liang, Z. An attempt to establish an Agrobacterium-mediated transient expression system in medicinal plants. Protoplasma 2020, 257, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Kusnadi, A.R.; Nikolov, Z.L.; Howard, J.A. Production of recombinant proteins in transgenic plants: Practical considerations. Biotechnol. Bioeng. 1997, 56, 473–484. [Google Scholar] [CrossRef]
- Marsian, J.; Lomonossoff, G.P. Molecular pharming-VLPs made in plants. Curr. Opin. Biotechnol. 2016, 37, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Norkunas, K.; Harding, R.; Dale, J.; Dugdale, B. Improving agroinfiltration-based transient gene expression in Nicotiana benthamiana. Plant Methods 2018, 14, 71. [Google Scholar] [CrossRef]
- Yuasa, T.; Sugiki, M.; Watanabe, Y. Activation of SIPK in response to UV-C irradiation, utility of a glutathione-S transferase-tagged plant MAP kinase by transient expression with agroinfiltration. Plant Biotechnol. 2005, 22, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Joh, L.D.; Wroblewski, T.; Ewing, N.N.; VanderGheynst, J.S. High-level transient expression of recombinant protein in lettuce. Biotechnol. Bioeng. 2005, 91, 861–871. [Google Scholar] [CrossRef]
- Rico, A.; Bennett, M.H.; Forcat, S.; Huang, W.E.; Preston, G.M. Agroinfiltration reduces ABA levels and suppresses Pseudomonas syringae-elicited salicylic acid production in Nicotiana tabacum. PLoS ONE 2010, 5, e8977. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, T.; Kalinowski, G.; Schwab, W. RNAi-induced silencing of gene expression in strawberry fruit (Fragaria × ananassa) by agroinfiltration: A rapid assay for gene function analysis. Plant J. 2006, 48, 818–826. [Google Scholar] [CrossRef]
- Menassa, R.; Ahmad, A.; Joensuu, J.J. Transient expression using agroinfiltration and its applications in molecular farming. In Molecular Farming in Plants: Recent Advances and Future Prospects; Wang, A., Ma, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar] [CrossRef]
- Tan, L.W.; Rahman, Z.A.; Goh, H.H.; Hwang, D.; Ismanizan, I.; Zamri, Z. Production of transgenic rice (indica cv. MR219) overexpressing Abp57 gene through Agrobacterium-mediated transformation. Sains. Malays. 2017, 46, 703–711. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, L. Transient expression and analysis of fluorescent reporter proteins in plant pollen tubes. Nat. Protoc. 2011, 6, 419–426. [Google Scholar] [CrossRef]
- Del Toro, F.; Tenllado, F.; Chung, B.N.; Canto, T.A. Procedure for the transient expression of genes by agroinfiltration above the permissive threshold to study temperature-sensitive processes in plant–pathogen interactions. Mol. Plant Pathol. 2014, 15, 848–857. [Google Scholar] [CrossRef]
- Voinnet, O.; Rivas, S.; Mestre, P.; Baulcombe, D. An enhanced transient expression system in plants based on suppression of gene silencing by the p19 protein of tomato bushy stunt virus. Plant J. 2003, 33, 949–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, K.; Yang, Q.; Yang, T.; Wu, Y.; Wang, G.; Yang, F.; Wang, R.; Lin, X.; Li, G. Development of Agrobacterium-mediated transient expression system in Caragana intermedia and characterization of CiDREB1C in stress response. BMC Plant Biol. 2019, 19, 237. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Lai, H. Gene delivery into plant cells for recombinant protein production. BioMed Res. Int. 2015, 2015, 932161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoji, Y.; Chichester, J.A.; Bi, H.; Musiychuk, K.; de la Rosa, P.; Goldschmidt, L.; Horsey, A.; Ugulava, N.; Palmer, G.A.; Mett, V.; et al. Plant-expressed HA as a seasonal influenza vaccine candidate. Vaccine 2008, 26, 2930–2934. [Google Scholar] [CrossRef]
- Ma, J.; Xiang, H.; Donnelly, D.J.; Meng, F.-R.; Xu, H.; Durnford, D.; Li, X.-Q. Genome editing in potato plants by agrobacterium-mediated transient expression of transcription activator-like effector nucleases. Plant Biotechnol. Rep. 2017, 11, 249–258. [Google Scholar] [CrossRef]
- Krishnan, V.; Jose, J.; Jolly, M.; Vinutha, T.; Kumar, R.; Manickavasagam, M.; Praveen, S.; Sachdev, A. ‘AGRODATE’: A rapid Agrobacterium-mediated transient expression tool for gene function analysis in leaf discs. J. Plant Biochem. Biotechnol. 2019, 29, 294–304. [Google Scholar] [CrossRef]
- Fraley, R.; Horsch, R.; Matzke, A.; Chilton, M.D.; Chilton, W.S.; Sanders, P.R. In vitro transformation of petunia cells by an improved method of co-cultivation with A. tumefaciens strains. Plant Mol. Biol. 1984, 3, 371–378. [Google Scholar] [CrossRef]
- Horsch, R.B.; Klee, H.J. Rapid assay of foreign gene expression in leaf disc transformed by Agrobacterium tumefaciens: Role of T-DNA borders in the transfer process. Proc. Natl. Acad. Sci. USA 1986, 83, 4428–4432. [Google Scholar] [CrossRef] [Green Version]
- Vaghchhipawala, Z.; Rojas, C.M.; Senthil-Kumar, M.; Mysore, K.S. Agroinoculation and Agroinfiltration: Simple Tools for Complex Gene Function Analyses. In Plant Reverse Genetics; Methods in Molecular Biology (Methods and, Protocols); Pereira, A., Ed.; Humana Press: Totowa, NJ, USA, 2011; Volume 678. [Google Scholar]
- Jia, H.; Liao, M.; Verbelen, J.P.; Vissenberg, K. Direct creation of marker-free tobacco plants from agroinfiltrated leaf discs. Plant Cell Rep. 2007, 26, 1961–1965. [Google Scholar] [CrossRef]
- Orzaez, D.; Mirabel, S.; Wieland, W.H.; Granell, A. Agroinjection of tomato fruits. A tool for rapid functional analysis of transgenes directly in fruit. Plant Physiol. 2006, 140, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Qian, W.; Yu, C.; Qin, H.; Liu, X.; Zhang, A.; Johansen, I.E.; Wang, D. Molecular and functional analysis of phosphomannomutase (PMM) from higher plants and genetic evidence for the involvement of PMM in ascorbic acid biosynthesis in Arabidopsis and Nicotiana benthamiana. Plant J. 2007, 49, 399–413. [Google Scholar] [CrossRef]
- Zottini, M.; Barizza, E.; Costa, A.; Formentin, E.; Ruberti, C.; Carimi, F.; Schiavo, F.L. Agroinfiltration of grapevine leaves for fast transient assays of gene expression and for long-term production of stable transformed cells. Plant Cell Rep. 2008, 27, 845–853. [Google Scholar] [CrossRef]
- Matsuo, K.; Fukuzawa, N.; Matsumura, T. A simple agroinfiltration method for transient gene expression in plant leaf discs. J. Biosci. Bioeng. 2016, 122, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Huang, X.; Wu, M.; Wang, Y.; Chang, Y.; Liu, K.; Zhang, J.; Zhang, Y.; Zhang, F.; Yi, L.; et al. A rapid, highly efficient and economical method of Agrobacterium-mediated In Planta transient transformation in living onion epidermis. PLoS ONE 2014, 9, e83556. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.I.; Park, H.S. Transient expression in chinese cabbage by hydrogen peroxide-aided agroinfiltration. Agric. Chem. Biotechnol. 2005, 48, 229–230. [Google Scholar]
- Naji-Talakar, S. Plant-derived biopharmaceuticals: Overview and success of agroinfiltration. Trends Capstone 2017, 2, 1–12. [Google Scholar]
- Juranić, M.; Nagahatenna, D.S.K.; Salinas-Gamboa, R.; Hand, M.L.; Sánchez-León, N.; Leong, W.H.; How, T.; Bazanova, N.; Spriggs, A.; Vielle-Calzada, J.-P.; et al. A detached leaf assay for testing transient gene expression and gene editing in cowpea (Vigna unguiculata [L.] Walp.). Plant Methods 2020, 16, 88. [Google Scholar] [CrossRef] [PubMed]
- Amoah, B.K.; Wu, H.; Sparks, C.; Jones, H.D. Factors influencing Agrobacterium-mediated transient expression of uidA in wheat inflorescence tissue. J. Exp. Bot. 2001, 52, 1135–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, T.; Bhattacharya, A.; Ahuja, P.; Chand, P. Transgenic tea [Camellia sinensis (L.) O. Kuntze cv. Kangra Jat] plants obtained by Agrobacterium-mediated transformation of somatic embryos. Plant Cell Rep. 2001, 20, 712–720. [Google Scholar] [CrossRef]
- Dong, J.Z.; McHughen, A. An improved procedure for production of transgenic flax plants using Agrobacterium tumefaciens. Plant Sci. 1993, 88, 61–71. [Google Scholar] [CrossRef]
- Matsuda, D.; Dreher, T.W. The tRNA-like structure of Turnip yellow mosaic virus RNA is a 3′-translational enhancer. Virology 2004, 321, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Sepahdoost, S.; Haghighi, H.; Sohi, H.H.; Jourabchi, E.; Ghorbanli, M. Agroinfiltration of Human Growth Hormone in Medicago sativa and Trifolium Alexanderium Leaves; National Congress of Biotechnology of Iran; Iranian Biotechnology Association: Trieste, Italy, 2005; Volume 4. [Google Scholar]
- Vargas-Guevara, C.; Vargas-Segura, C.; Villalta-Villalobos, J.; Pereira, L.F.P.; Gatica-Arias, A. A simple and efficient agroinfiltration method in coffee leaves (Coffea arabica L.): Assessment of factors affecting transgene expression. 3 Biotech 2018, 8, 471. [Google Scholar] [CrossRef]
- Ahlquist, P. RNA-dependent RNA polymerases, viruses, and RNA silencing. Science 2002, 296, 1270–1273. [Google Scholar] [CrossRef] [Green Version]
- Fischer, R.; Stoger, E.; Schillberg, S.; Christou, P.; Twyman, R.M. Plant-based production of biopharmaceuticals. Curr. Opin. Plant Biol. 2004, 7, 152–158. [Google Scholar] [CrossRef]
- Delgadillo, M.O.; Saenz, P.; Salvador, B.; García, J.A.; Simón-Mateo, C. Human influenza virus NS1 protein enhances viral pathogenicity and acts as an RNA silencing suppressor in plants. J. Gen. Virol. 2004, 85, 993–999. [Google Scholar] [CrossRef]
- Han, L.; Zhang, L.; Liu, J.; Li, H.; Wang, Y.; Hasi, A. Transient expression of optimized and synthesized nattokinase gene in melon (Cucumis melo L.) fruit by agroinfiltration. Plant Biotechnol. 2015, 15, 0430a. [Google Scholar] [CrossRef] [Green Version]
- King, J.L.; Finer, J.J.; McHale, L.K. Development and optimization of agroinfiltration for soybean. Plant Cell Rep. 2015, 34, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Wydro, M.; Kozubek, E.; Lehmann, P. Optimization of transient Agrobacterium-mediated gene expression system in leaves of Nicotiana benthamiana. Acta Biochim Pol. 2006, 53, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Arzola, L.; Chen, J.; Rattanaporn, K.; Maclean, J.M.; McDonald, K.A. Transient co-expression of post-transcriptional gene silencing suppressors for increased In Planta expression of a recombinant anthrax receptor fusion protein. Int. J. Mol. Sci. 2011, 12, 4975–4990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Fradin, E.; de Jonge, R.; van Esse, H.P.; Smit, P.; Liu, C.-M.; Thomma, B.P.H.J. Optimized agroinfiltration and virus-induced gene silencing to study Ve1-mediated Verticillium resistance in tobacco. Mol. Plant Microbe Interact. 2013, 26, 182–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashandy, H.; Jalkanen, S.; Teeri, T.H. Within leaf variation is the largest source of variation in agroinfiltration of Nicotiana benthamiana. Plant Methods 2015, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Patil, B.L.; Fauquet, C.M. Light intensity and temperature affect systemic spread of silencing signal in transient agroinfiltration studies. Mol. Plant Pathol. 2015, 16, 484–494. [Google Scholar] [CrossRef]
- Fujiuchi, N.; Matsuda, R.; Matoba, N.; Fujiwara, K. Removal of bacterial suspension water occupying the intercellular space of detached leaves after agroinfiltration improves the yield of recombinant hemagglutinin in a Nicotiana benthamiana transient gene expression system. Biotechnol Bioeng. 2016, 113, 901–906. [Google Scholar] [CrossRef]
- Hanittinan, O.; Oo, Y.; Chaotham, C.; Rattanapisit, K.; Shanmugaraj, B.; Phoolcharoen, W. Expression optimization, purification and in vitro characterization of human epidermal growth factor produced in Nicotiana benthamiana. Biotechnol. Rep. 2020, 28, e00524. [Google Scholar] [CrossRef] [PubMed]
- Diamos, A.G.; Hunter, J.G.L.; Pardhe, M.D.; Rosenthal, S.H.; Sun, H.; Foster, B.C.; DiPalma, M.P.; Chen, Q.; Mason, H.S. High level production of monoclonal antibodies using an optimized plant expression system. Front. Bioeng. Biotechnol. 2020, 7, 472. [Google Scholar] [CrossRef]
- Heidari-Japelaghi, R.; Valizadeh, M.; Haddad, R.; Dorani-Uliaiea, E.; Jalali-Javaran, M. Production of bioactive human IFN-γ protein by agroinfiltration in tobacco. Protein Expr. Purif. 2020, 173, 105616. [Google Scholar] [CrossRef]
- Guy, E.; Boulain, H.; Aigu, Y.; Pennec, C.L.; Chawki, K.; Morlière, S.; Schädel, K.; Kunert, G.; Simon, J.-C.; Sugio, A. Optimization of agroinfiltration in Pisum sativum provides a new tool for studying the salivary protein functions in the pea aphid complex. Front Plant Sci. 2016, 7, 1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokkirala, V.R.; Peng, Y.; Abbagani, S.; Zhu, Z.; Umate, P. Subcellular localization of proteins of Oryza sativa L. in the model tobacco and tomato plants. Plant Signal Behav. 2010, 5, 1336–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deguchi, M.; Bogush, D.; Weeden, H.; Spuhler, Z.; Potlakayala, S.; Kondo, T.; Zhang, Z.J.; Rudrabhatla, S. Establishment and optimization of a hemp (Cannabis sativa L.) agroinfiltration system for gene expression and silencing studies. Sci. Rep. 2020, 10, 3504. [Google Scholar] [CrossRef] [Green Version]
- Sung, H.P. In Vitro culture and transformation by agroinfiltration of lisianthus (Eustoma russellianus) pollen. J. Life Sci. 2004, 14, 1018–1022. [Google Scholar]
- Hussein, G.M.; Abu El-Heba, G.A.; Abdou, S.M.; Abdallah, N.A. Optimization of transient gene expression system in Gerbera jamesonii petals. GM Crops Food 2013, 4, 50–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mani, T.; Manjula, S. Optimization of Agrobacterium-mediated transient gene expression and endogenous gene silencing in Piper colubrinum Link. by vacuum infiltration. Plant Cell Tiss. Organ Cult. 2011, 105, 113–119. [Google Scholar] [CrossRef]
- Santos-Rosa, M.; Poutaraud, A.; Merdinoglu, D.; Mestre, P. Development of a transient expression system in grapevine via agro-infiltration. Plant Cell Rep. 2008, 27, 1053–1063. [Google Scholar] [CrossRef]
- Faizal, A.; Geelen, D. Agroinfiltration of intact leaves as a method for the transient and stable transformation of saponin producing Maesa lanceolata. Plant Cell Rep. 2012, 31, 1517–1526. [Google Scholar] [CrossRef]
- Chevreau, E.; Dousset, N.; Joffrion, C.; Richer, A.; Charrier, A.; Vergne, E. Agroinfiltration is a key factor to improve the efficiency of apple and pear transformation. Sci. Hortic. 2019, 251, 150–154. [Google Scholar] [CrossRef]
- Abd-Aziz, N.; Tan, B.C.; Rejab, N.A.; Othman, R.Y.; Khalid, N. A new plant expression system for producing pharmaceutical proteins. Mol. Biotechnol. 2020, 62, 240–251. [Google Scholar] [CrossRef]
- Sohi, H.H.; Jourabchi, E.; Khodabandeh, M. Transient expression of human growth hormone in potato (Solanum tuberosum), tobacco (Nicotiana tobacum) and lettuce (Lactuca sativa) leaves by agroinfiltration. Iranian J. Biotechnol. 2005, 3, 109–113. [Google Scholar]
- Koscianska, E.; Kalantidis, K.; Wypijewski, K.; Sadowski, J.; Tabler, M. Analysis of RNA silencing in agroinfiltrated leaves of Nicotiana benthamiana and Nicotiana tabacum. Plant Mol. Biol. 2005, 59, 647–661. [Google Scholar] [CrossRef]
- Yamamoto, T.; Hoshikawa, K.; Ezura, K.; Okazawa, R.; Fujita, S.; Takaoka, M.; Mason, H.S.; Ezura, H.; Miura, K. Improvement of the transient expression system for production of recombinant proteins in plants. Sci. Rep. 2018, 8, 4755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Affandi, N.D.; Mostaffa, N.H.; Al-Idrus, A. Interactomics: Development of an efficient and improved Agrobacterium tumefaciens-mediated transformation method for transient expression of heterologous protein in recalcitrant plant tissues in planta. Res. Square 2020. [Google Scholar] [CrossRef] [Green Version]
- Belhaj, K.; Chaparro-Garcia, A.; Kamoun, S.; Nekrasov, V. Plant genome editing made easy: Targeted mutagenesis in model and crop plants using the CRISPR/Cas system. Plant Methods 2013, 9, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, R.; Emans, N. Molecular farming of pharmaceutical proteins. Transgenic Res. 2000, 9, 279–299. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Yamaguchi, K. Efficient agroinfiltration-mediated transient GUS expression for assaying different promoters in rice. Plant Biotechnol. 2003, 20, 235–239. [Google Scholar] [CrossRef]
- Coura, R.; Nardi, N. A role for adeno-associated viral vectors in gene therapy. Genet. Mol. Biol. 2008, 31, 1–11. [Google Scholar] [CrossRef]
- Soyars, C.L.; Peterson, B.A.; Burr, C.A.; Nimchuk, Z.L. Cutting edge genetics: CRISPR/Cas9 editing of plant genomes. Plant Cell Physiol. 2018, 59, 1608–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, R.; Liang, Y.; Lee, M.Y.; Pidatala, V.R.; Mortimer, J.C.; Scheller, H.V. Agrobacterium-mediated transient transformation of sorghum leaves for accelerating functional genomics and genome editing studies. BMC Res. Notes 2020, 13, 116. [Google Scholar] [CrossRef] [Green Version]
- Li, J.F.; Norville, J.; Aach, J.; McCormack, M.; Zhang, D.; Bush, J.; Church, G.M.; Sheen, J. Multiplex and homologous recombination–mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9. Nat. Biotechnol. 2013, 31, 688–691. [Google Scholar] [CrossRef]
- Jiang, W.; Zhou, H.; Bi, H.; Fromm, M.; Yang, B.; Weeks, D.P. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucleic Acids Res. 2013, 41, e188. [Google Scholar] [CrossRef] [PubMed]
- Nekrasov, V.; Staskawicz, B.; Weigel, D.; Jones, J.D.G.; Kamoun, S. Targeted mutagenesis in the model plant Nicotiana benthamiana using Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 691–693. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Kumar, J.; Alok, A.; Tuli, R. RNA-guided genome editing for target gene mutations in wheat. G3 2013, 3, 2233–2238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piatek, A.; Ali, Z.; Baazim, H.; Li, L.; Abulfaraj, A.; Al-Shareef, S.; Aouida, M.; Mahfouz, M.M. RNA-guided transcriptional regulation in planta via synthetic dCas9-based transcription factors. Plant Biotechnol. J. 2015, 13, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Han, T.; Liu, G.; Chen, T.; Wang, Y.; Yu, A.Y.L.; Liu, Y. A geminivirus-based guide RNA delivery system for CRISPR/Cas9 mediated plant genome editing. Sci. Rep. 2015, 5, 14926. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, D.J.; Ali, Z.; Wang, C.; Aljedaani, F.; Hooykaas, P.J.J.; Mahfouz, M.; de Pater, S. CRISPR/Cas9 Mutagenesis by translocation of Cas9 protein into plant cells via the Agrobacterium Type IV secretion system. Front. Genome Ed. 2020, 2, 6. [Google Scholar] [CrossRef]
- Baltes, N.J.; Hummel, A.W.; Konecna, E.; Cegan, R.; Bruns, A.N.; Bisaro, D.M.; Voytas, D.F. Conferring resistance to geminiviruses with the CRISPR–Cas prokaryotic immune system. Nat. Plants 2015, 1, 1–4. [Google Scholar] [CrossRef]
- Vazquez-Vilar, M.; Bernabé-Orts, J.M.; Fernandez-del-Carmen, A.; Ziarsolo, P.; Blanca, J.; Granell, A.; Orzaez, D. A modular toolbox for gRNA–Cas9 genome engineering in plants based on the GoldenBraid standard. Plant Methods 2016, 12, 10. [Google Scholar] [CrossRef] [Green Version]
- Ali, Z.; Abul-faraj, A.; Li, L.; Ghosh, N.; Piatek, M.; Mahjoub, A.; Aouida, M.; Piatek, A.; Baltes, N.J.; Voytas, D.F.; et al. Efficient virus-mediated genome editing in plants using the CRISPR/Cas9 system. Mol. Plant 2015, 8, 1288–1291. [Google Scholar] [CrossRef] [Green Version]
- Alagoz, Y.; Gurkok, T.; Zhang, B.; Unver, T. Manipulating the biosynthesis of bioactive compound alkaloids for next-generation metabolic engineering in opium poppy using CRISPR-Cas9 genome editing technology. Sci. Rep. 2016, 6, 30910. [Google Scholar] [CrossRef] [Green Version]
- Mubarik, M.S.; Khan, S.H.; Ahmad, A.; Khan, Z.; Sajjad, M.; Khan, I.A. Disruption of phytoene desaturase gene using transient expression of Cas9: gRNA Complex. Int. J. Agric. Biol. 2016, 18. [Google Scholar] [CrossRef]
- Jia, H.; Wang, N. Targeted genome editing of sweet orange using Cas9/sgRNA. PLoS ONE 2014, 9, e93806. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Zhang, Y.; Orbovic, V.; Xu, J.; White, F.; Jones, J.; Wang, N. Genome editing of the disease susceptibility gene CsLOB1 in citrus confers resistance to citrus canker. Plant Biotechnol. J. 2016, 15, 817–823. [Google Scholar] [CrossRef] [Green Version]
- Martín-Pizarro, C.; Triviño, J.C.; Posé, D. Functional analysis of the TM6 MADS-box gene in the octoploid strawberry by CRISPR/Cas9-directed mutagenesis. J. Exp. Bot. 2019, 70, 885–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stajič, E.; Kiełkowska, A.; Murovec, J.; Bohanec, B. Deep sequencing analysis of CRISPR/Cas9 induced mutations by two delivery methods in target model genes and the CENH3 region of red cabbage (Brassica oleracea var. capitata f. rubra). Plant Cell Tissue Organ Cult. 2019, 139, 227–235. [Google Scholar] [CrossRef]
- Syombua, E.D.; Zhang, Z.; Tripathi, J.N. A CRISPR/Cas9-based genome-editing system for yam (Dioscorea spp.). Plant Biotechnol. J. 2020, 19, 645–647. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Gao, C. Targeted genome modification technologies and their applications in crop improvements. Plant Cell Rep. 2014, 33, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Bortesi, L.; Fischer, R. The CRISPR/Cas9 system for plant genome editing and beyond. Biotechnol. Adv. 2015, 33, 41–52. [Google Scholar] [CrossRef]
- Goodin, M.M.; Dietzgen, R.G.; Schichnes, D.; Ruzin, S.; Jackson, A.O. pGD vectors: Versatile tools for the expression of green and red fluorescent protein fusions in agroinfiltrated plant leaves. Plant J. 2002, 31, 375–383. [Google Scholar] [CrossRef]
- Jia, H.; Wang, N. Xcc-facilitated agroinfiltration of citrus leaves: A tool for rapid functional analysis of transgenes in citrus leaves. Plant Cell Rep. 2014, 33, 1993–2001. [Google Scholar] [CrossRef]
- Poles, L.; Licciardello, C.; Distefano, G.; Nicolosi, E.; Gentile, A.; Malfa, S.L. Recent advances of in vitro culture for the application of new breeding techniques in citrus. Plants 2020, 9, 938. [Google Scholar] [CrossRef]
- Li, X.; Li, H.; Zhao, Y.; Zong, P.; Zhan, Z.; Piao, Z. Establishment of Agrobacterium-mediated genetic transformation and application of CRISPR/Cas9 gene editing system to Chinese cabbage (Brassica rapa L. ssp. pekinensis). Res. Square 2020. [Google Scholar] [CrossRef]
- Grefen, C.; Donald, N.; Hashimoto, K.; Kudla, J.; Schumacher, K.; Blatt, M.R. A ubiquitin-10 promoter-based vector set for fluorescent protein tagging facilitates temporal stability and native protein distribution in transient and stable expression studies. Plant J. 2010, 64, 355–365. [Google Scholar] [CrossRef]
- Picard, K.; Lee, R.; Hellens, R.; Macknight, R. Transient gene expression in Medicago truncatula leaves via agroinfiltration. In Legume Genomics Methods Protocols; Rose, J.R., Ed.; Humana Press: Totowa, NJ, USA, 2013; pp. 215–226. [Google Scholar]
- Lombardi, R.; Villani, M.E.; Di Carli, M.; Brunetti, P.; Benvenuto, E.; Donini, M. Optimisation of the purification process of a tumour-targeting antibody produced in N. benthamiana using vacuum-agroinfiltration. Transgenic Res. 2010, 19, 1083–1097. [Google Scholar] [CrossRef] [PubMed]
- D’Aoust, M.A.; Lavoie, P.O.; Belles-Isles, J.; Bechtold, N.; Martel, M.; Vézina, L.P. Transient expression of antibodies in plants using syringe agroinfiltration. Methods Mol. Biol. 2009, 483, 41–50. [Google Scholar] [PubMed]
- Du, J.; Rietman, H.; Vleeshouwers, V.G.A.A. Agroinfiltration and PVX agroinfection in potato and Nicotiana benthamiana. J. Visual Exp. 2013, 83, e50971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schöb, H.; Kunz, C.; Meins, J.F. Silencing of transgenes introduced into leaves by agroinfiltration: A simple, rapid method for investigating sequence requirements for gene silencing. Mol. Gen. Genet. 1997, 256, 581–585. [Google Scholar] [CrossRef]
- Voinnet, O.; Baulcombe, D. Systemic signalling in gene silencing. Nature 1997, 389, 553. [Google Scholar] [CrossRef]
- Voinnet, O.; Vain, P.; Angell, S.; Baulcombe, D.C. Systemic spread of sequence-specific transgene RNA degradation in plants is initiated by localized introduction of ectopic promoterless DNA. Cell 1998, 95, 177–187. [Google Scholar] [CrossRef] [Green Version]
- Bertazzon, N.; Raiola, A.; Castiglioni, C.; Gardiman, M.; Angelini, E.; Borgo, M.; Ferrari, S. Transient silencing of the grapevine gene VvPGIP1 by agroinfiltration with a construct for RNA interference. Plant Cell Rep. 2012, 31, 133–143. [Google Scholar] [CrossRef]
- Dubey, V.K.; Lee, U.G.; Kwon, D.H.; Lee, S.H. Agroinfiltration-based expression of hairpin RNA in soybean plants for RNA interference against Tetranychus urticae. Pestic. Biochem. Physiol. 2017, 142, 53–58. [Google Scholar] [CrossRef]
- Abbink, T.E.M.; Tjernberg, P.A.; Bol, J.F.; Linthorst, H.J.M. Tobacco mosaic virus helicase domain induces necrosis in N gene-carrying tobacco in the absence of virus replication. Mol. Plant Microbe Interact. 1998, 11, 1242–1246. [Google Scholar] [CrossRef] [Green Version]
- Palanichelvam, K.; Cole, A.B.; Shababi, M.; Schoelz, J.E. Agroinfiltration of cauliflower mosaic virus gene vi elicits hypersensitive response in Nicotiana species. Mol. Plant Microbe Interact. 2000, 13, 1275–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Hoorn, R.A.L.; Laurent, F.; Roth, R.; De Wit, P.J.G.M. Agroinfiltration is a versatile tool that facilitates comparative analyses of Avr9/Cf-9-induced and Avr4/Cf-4-induced necrosis. Mol. Plant-Microbe Interact. 2000, 13, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Bendahmane, M.; Lynch, C., II; Tulsiani, D.R. Calmodulin signals capacitation and triggers the agonist-induced acrosome reaction in mouse spermatozoa. Arch. Biochem Biophys. 2001, 390, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Peart, J.R.; Lu, R.; Sadanandom, A.; Malcuit, I.; Moffett, P.; Brice, D.C.; Schauser, L.; Jaggard, D.A.W.; Xiao, S.; Coleman, M.J.; et al. Ubiquitin ligase-associated protein SGT1 is required for host and nonhost disease resistance in plants. Proc. Natl. Acad. Sci. USA 2002, 99, 10865–10869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leckie, B.M.; Stewart, C.N. Agroinfiltration as a technique for rapid assays for evaluating candidate insect resistance transgenes in plants. Plant Cell Rep. 2011, 30, 325–334. [Google Scholar] [CrossRef]
- Yang, Y.; Li, R.; Qi, M. In vivo analysis of plant promoters and transcription factors by agroinfiltration of tobacco leaves. Plant J. 2000, 22, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Rancé, I.; Norre, F.; Gruber, V.; Theisen, M. Combination of viral promoter sequences to generate highly active promoters for heterologous therapeutic protein over-expression in plants. Plant Sci. 2002, 162, 833–842. [Google Scholar] [CrossRef]
- Merle, C.; Perret, S.; Lacour, T.; Jonval, V.; Hudaverdian, S.; Garrone, R.; Ruggiero, F.; Theisen, M. Hydroxylated human homotrimeric collagen I in Agrobacterium tumefaciens-mediated transient expression and in transgenic tobacco plant. FEBS Lett. 2002, 515, 114–118. [Google Scholar] [CrossRef] [Green Version]
- Tampakaki, A.P.; Panopoulos, N.J. Elicitation of hypersensitive cell death by extracellularly targeted HrpZPsph produced In Planta. Mol. Plant Microbe Interact. 2000, 13, 1366–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaquero, C.; Sack, M.; Schuster, F. A carcinoembryonic antigen-specific diabody produced in tobacco. FASEB J. 2002, 16, 408–410. [Google Scholar] [CrossRef] [PubMed]
- Kathuria, S.; Sriraman, R.; Nath, R.; Sack, M.; Pal, R.; Artsaenko, O.; Talwar, G.P.; Fischer, R.; Finnern, R. Efficacy of plant-produced recombinant antibodies against HCG. Hum. Reprod. 2002, 17, 2054–2061. [Google Scholar] [CrossRef] [Green Version]
- Vézina, L.P.; Faye, L.; Lerouge, P.; D’Aoust, M.-A.; Marquet-Blouin, E.; Burel, C.; Lavoie, P.-O.; Bardor, M.; Gomord, V. Transient co-expression for fast and high-yield production of antibodies with human-like N-glycans in plants. Plant Biotechnol. J. 2009, 7, 442–455. [Google Scholar] [CrossRef]
- Habibi-Pirkoohi, M.; Malekzadeh-Shafaroudi, S.; Marashi, H.; Moshtaghi, N.; Nassiri, M.; Zibaee, S. Transient expression of foot and mouth disease virus (FMDV) coat protein in tobacco (Nicotiana tabacom) via agroinfiltration. Iranian J. Biotechnol. 2014, 12, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Zhang, Y.; Tang, S.; Zhao, Q.; Zhang, Z.; Zhang, H.; Dong, L.; Guo, H.; Xie, Q. An efficient system to detect protein ubiquitination by agroinfiltration in Nicotiana benthamiana. Plant J. 2010, 61, 893–903. [Google Scholar] [CrossRef]
- Pillay, P.; Kunert, K.J.; van Wyk, S.; Makgopa, M.E.; Cullis, C.A.; Vorster, B.J. Agroinfiltration contributes to VP1 recombinant protein degradation. Bioengineered 2016, 7, 459–477. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Mazarei, M.; Rudis, M.R.; Fethe, M.H.; Stewart, J.C.N. Rapid in vivo analysis of synthetic promoters for plant pathogen phytosensing. BMC Biotechnol. 2011, 11, 108. [Google Scholar] [CrossRef] [Green Version]
- D’Aoust, M.A.; Lavoie, P.O.; Couture, M.M.-J.; Trépanier, S.; Guay, J.-M.; Dargis, M.; Mongrand, S.; Landry, N.; Ward, B.J.; Vézina, L.-P. Influenza virus-like particles produced by transient expression in Nicotiana benthamiana induce a protective immune response against a lethal viral challenge in mice. Plant Biotechnol. J. 2008, 6, 930–940. [Google Scholar] [CrossRef]
- Rajabi, A.; Fahmideh, L.; Keikhasaber, M.; Omran, V.G. Evaluation of transient expression of AS1 and 4′CGT genes in African violets petals by agroinfiltration for production new color in the flower. Iranian J. Biol. 2021, 2021, 2383–2738. [Google Scholar]
- Liu, P.; Wang, Y.; Ulrich, R.G.; Simmons, C.W.; VanderGheynst, J.S.; Gallo, R.L.; Huang, C.-M. Leaf-Encapsulated vaccines: Agroinfiltration and transient expression of the antigen staphylococcal endotoxin B in radish leaves. J. Immunol. Res. 2018, 2018, 3710961. [Google Scholar] [CrossRef] [Green Version]
- Kaushik, P. Standardisation of an agroinfiltration protocol for eggplant fruits and proving its usefulness by over-expressing the SmHQT gene. Preprints 2019. [Google Scholar] [CrossRef] [Green Version]
- Hesami, M.; Alizadeh, M.; Naderi, R.; Tohidfar, M. Forecasting and optimizing Agrobacterium-mediated genetic transformation via ensemble model- fruit fly optimization algorithm: A data mining approach using chrysanthemum databases. PLoS ONE 2020, 15, e0239901. [Google Scholar] [CrossRef] [PubMed]
- Leuzinger, K.; Dent, M.; Hurtado, J.; Stahnke, J.; Lai, H.; Zhou, X.; Chen, Q. Efficient agroinfiltration of plants for high-level transient expression of recombinant proteins. J. Vis. Exp. 2013, 77, 50521. [Google Scholar] [CrossRef] [PubMed]
- Mirakhorli, N.; Reyhani, A.H.; Shiran, B.; Hooshmand, S. Optimization of transient expression by Agrobacterium in almond. J. Genet. Eng. Biotechnol. 2014, 3, 9–18. [Google Scholar]
- Kumar, S. Biosafety and Biosecurity Issues in Biotechnology Research. Biosafety 2015, 4, 1. [Google Scholar] [CrossRef]
- Breyer, D.; Kopertekh, L.; Reheul, D. Alternatives to antibiotic resistance marker genes for in vitro selection of genetically modified plants—Scientific developments, current use, operational access and biosafety considerations. Crit. Rev. Plant Sci. 2014, 33, 286–330. [Google Scholar] [CrossRef] [Green Version]
- Shanmugaraj, B.; Ramalingam, S. Plant expression platform for the production of recombinant pharmaceutical proteins. Austin J. Biotechnol. Bioeng. 2014, 1, 4. [Google Scholar]
- Mahesh, S. The state of art of new transgenic techniques in plant breeding: A review. J. Adv. Biol. Biotechnol. 2016, 9, 1–11. [Google Scholar] [CrossRef]
- Grosse-Holz, F.; Madeira, L.; Zahid, M.A.; Songer, M.; Kourelis, J.; Fesenko, M.; Ninck, S.; Kaschani, F.; Kaiser, M.; van der Hoorn, R.A.L. Three unrelated protease inhibitors enhance accumulation of pharmaceutical recombinant proteins in Nicotiana benthamiana. Plant Biotechnol. J. 2018, 16, 1797–1810. [Google Scholar] [CrossRef] [Green Version]


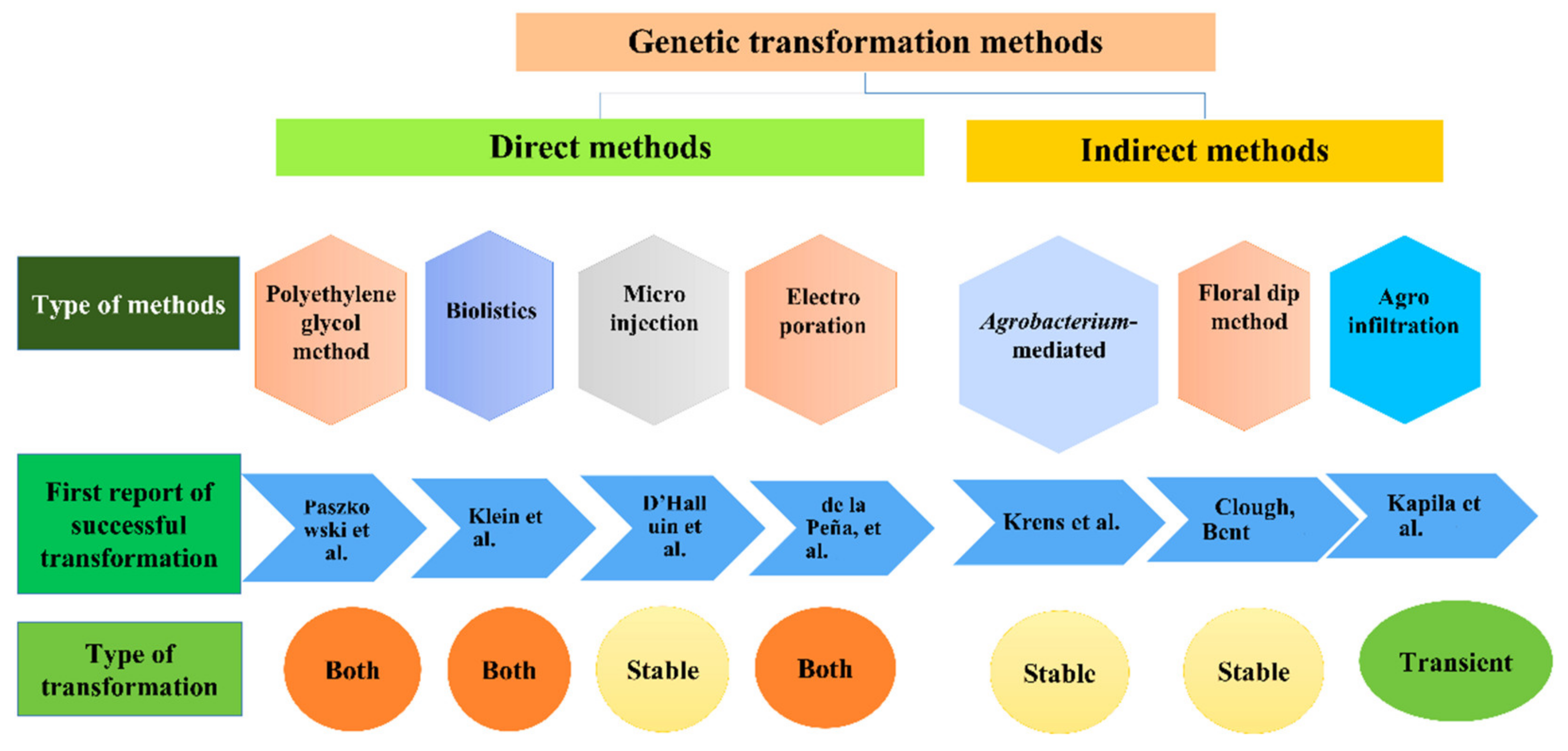{kind=link}
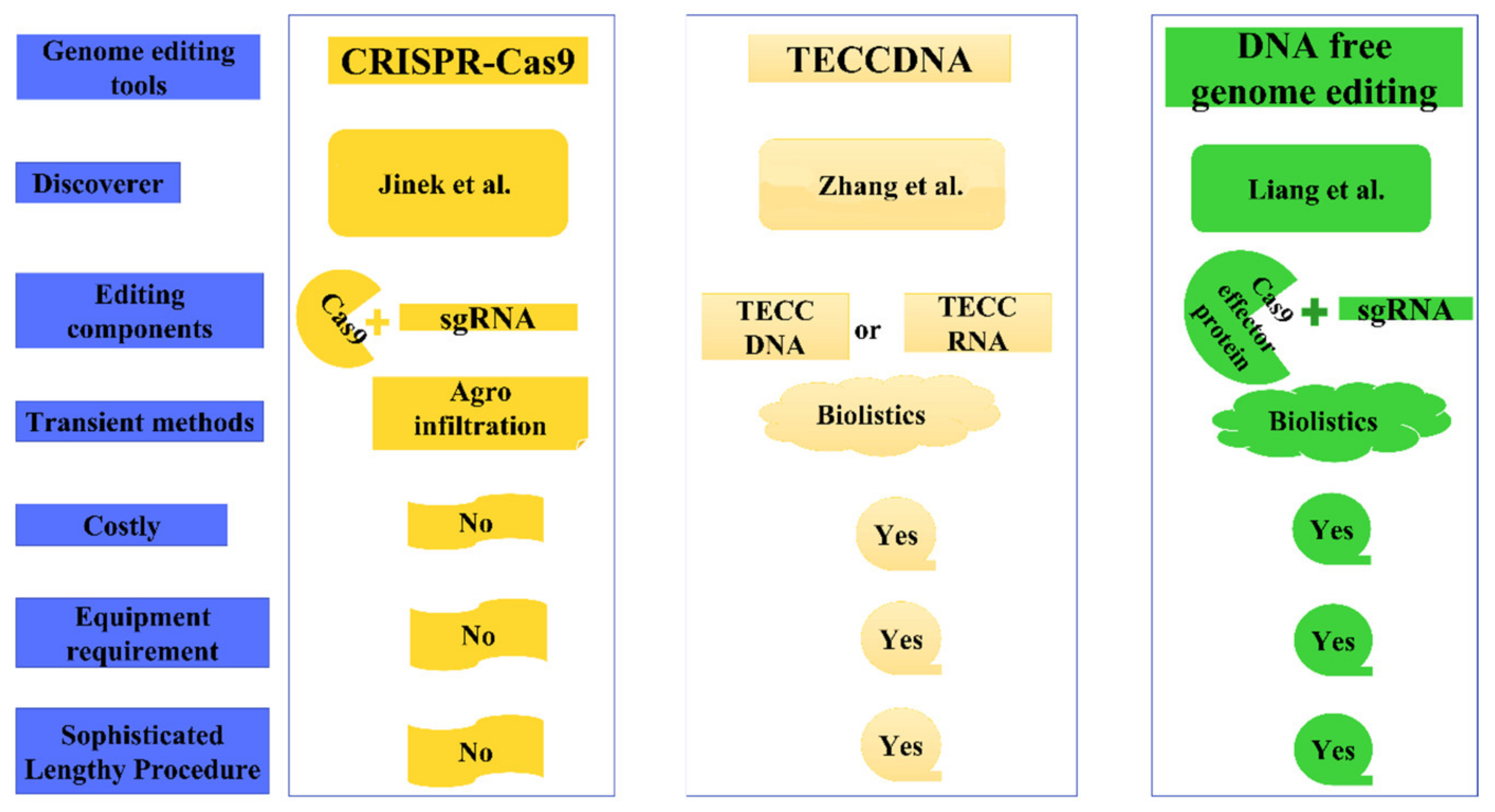{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaur, M.; Manchanda, P.; Kalia, A.; Ahmed, F.K.; Nepovimova, E.; Kuca, K.; Abd-Elsalam, K.A. Agroinfiltration Mediated Scalable Transient Gene Expression in Genome Edited Crop Plants. Int. J. Mol. Sci. 2021, 22, 10882. https://doi.org/10.3390/ijms221910882
Kaur M, Manchanda P, Kalia A, Ahmed FK, Nepovimova E, Kuca K, Abd-Elsalam KA. Agroinfiltration Mediated Scalable Transient Gene Expression in Genome Edited Crop Plants. International Journal of Molecular Sciences. 2021; 22(19):10882. https://doi.org/10.3390/ijms221910882
Chicago/Turabian StyleKaur, Maninder, Pooja Manchanda, Anu Kalia, Farah K. Ahmed, Eugenie Nepovimova, Kamil Kuca, and Kamel A. Abd-Elsalam. 2021. "Agroinfiltration Mediated Scalable Transient Gene Expression in Genome Edited Crop Plants" International Journal of Molecular Sciences 22, no. 19: 10882. https://doi.org/10.3390/ijms221910882
APA StyleKaur, M., Manchanda, P., Kalia, A., Ahmed, F. K., Nepovimova, E., Kuca, K., & Abd-Elsalam, K. A. (2021). Agroinfiltration Mediated Scalable Transient Gene Expression in Genome Edited Crop Plants. International Journal of Molecular Sciences, 22(19), 10882. https://doi.org/10.3390/ijms221910882