
Stress Driven Discovery of Natural Products From Actinobacteria with Anti-Oxidant and Cytotoxic Activities Including Docking and ADMET Properties

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
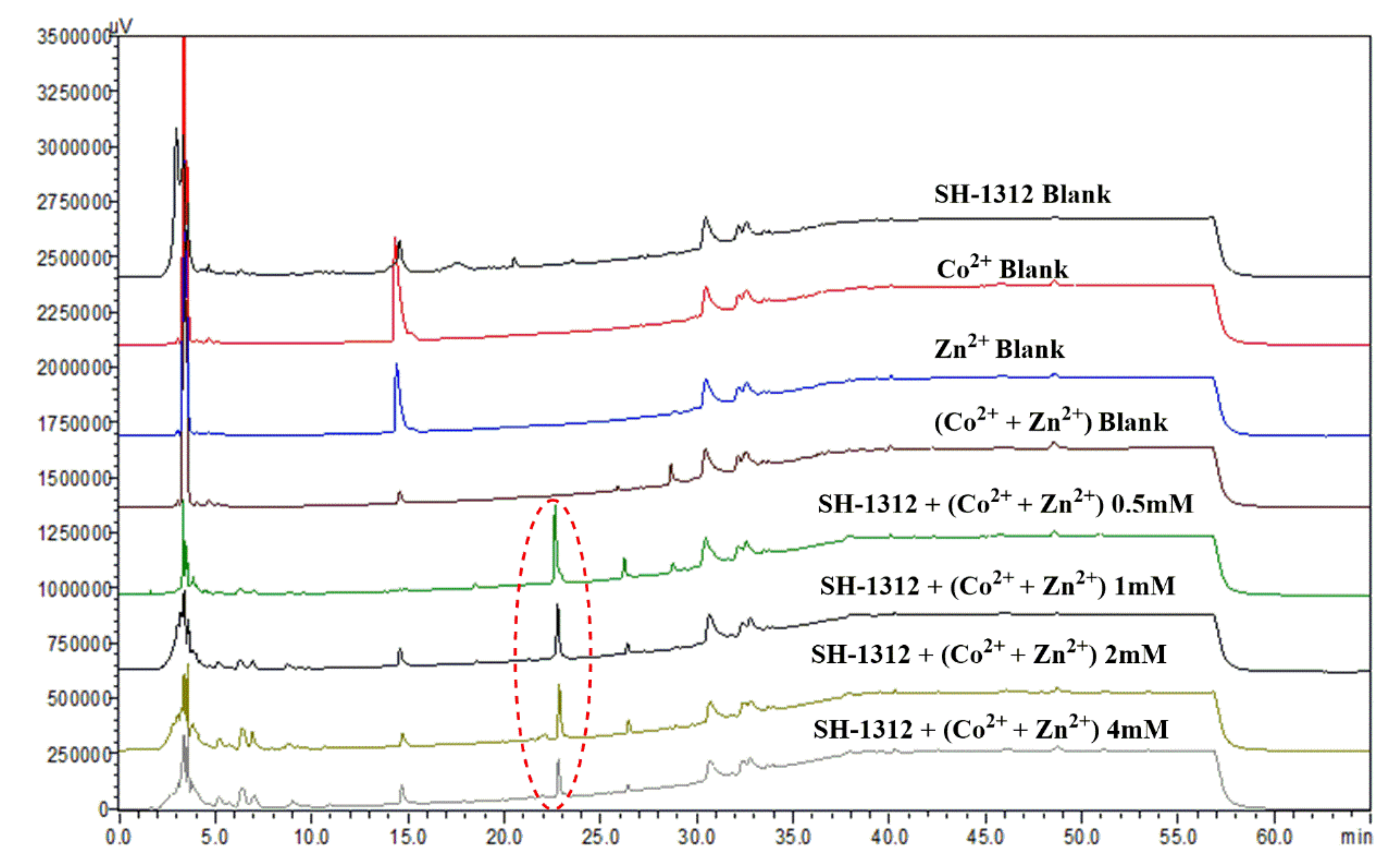
2.1. HPLC Evaluation of Secondary Metabolites of Metal Treated and Untreated Extracts
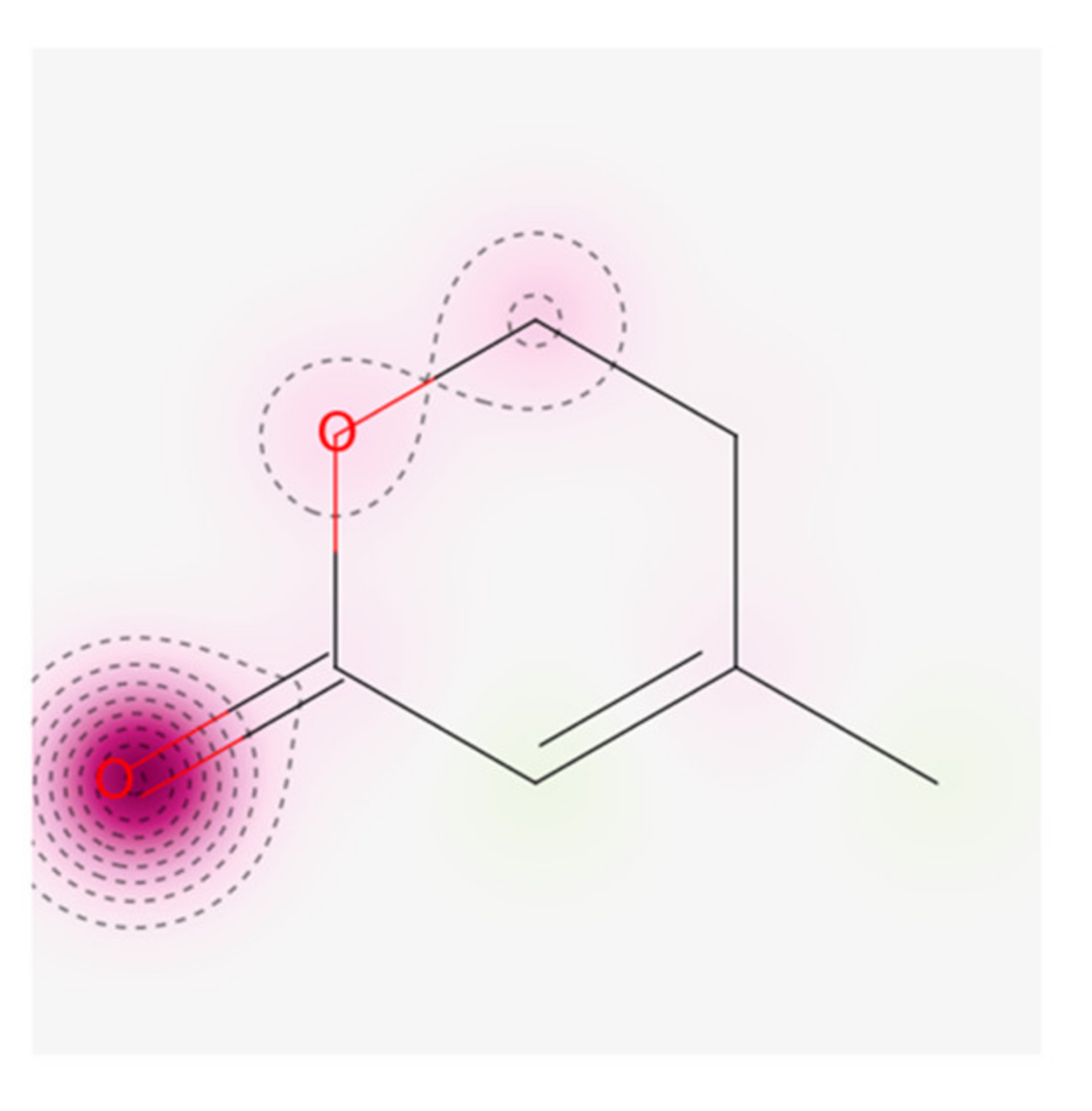
2.2. Identification and Structure Determination of Metal-Induced Secondary Metabolite
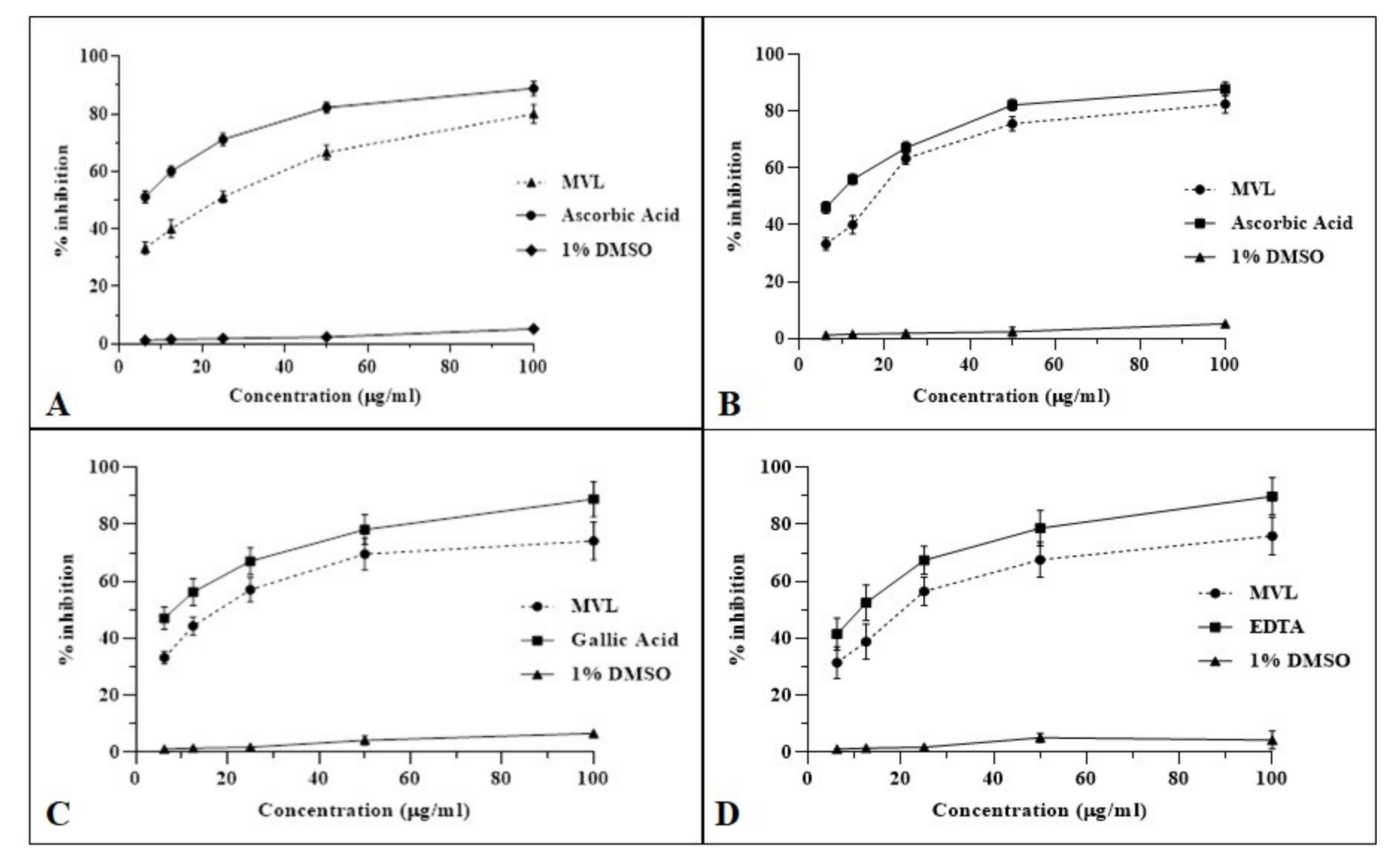
2.3. Anti-Oxidant Assay
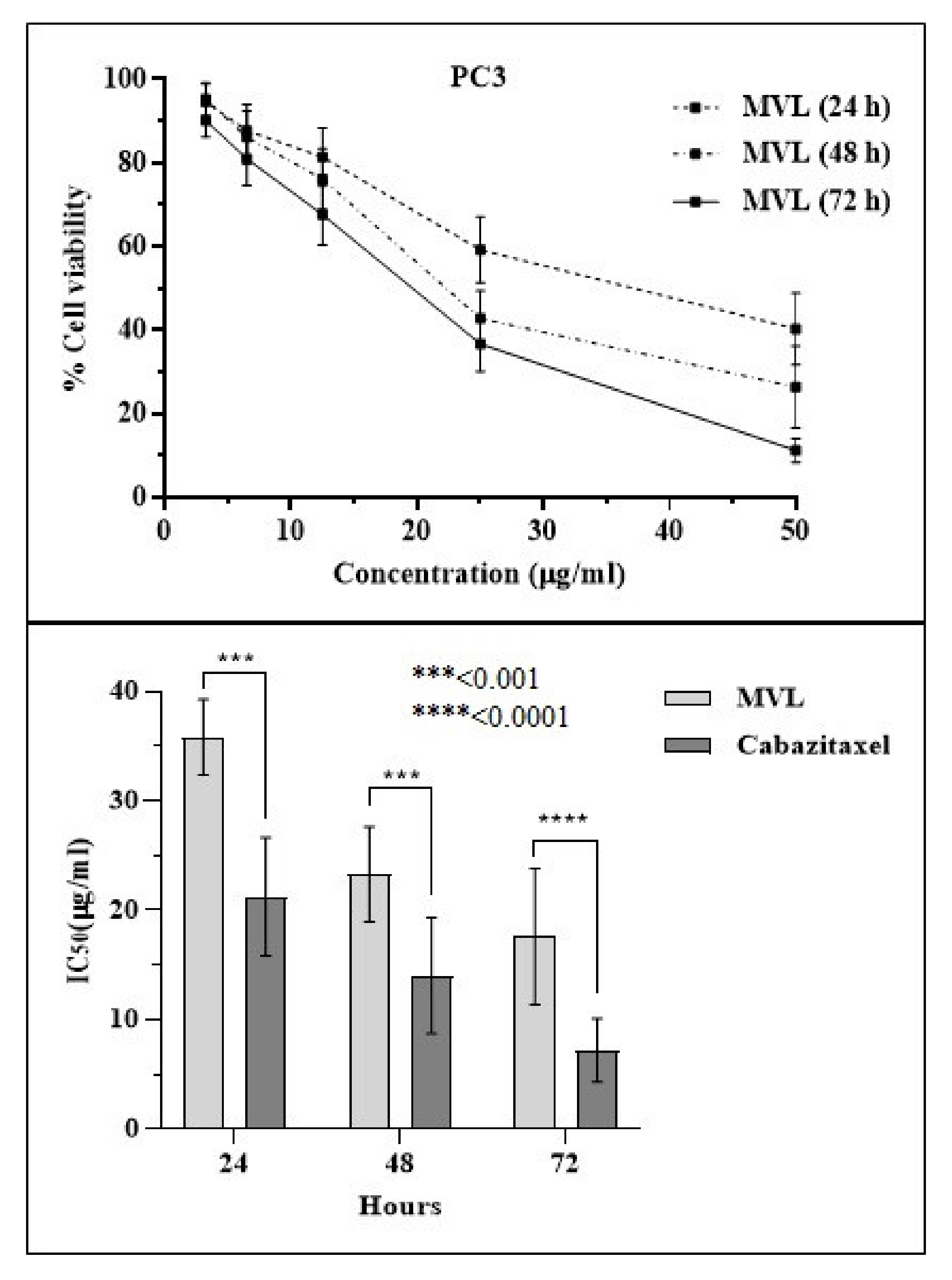
2.4. Cytotoxicity Assessment
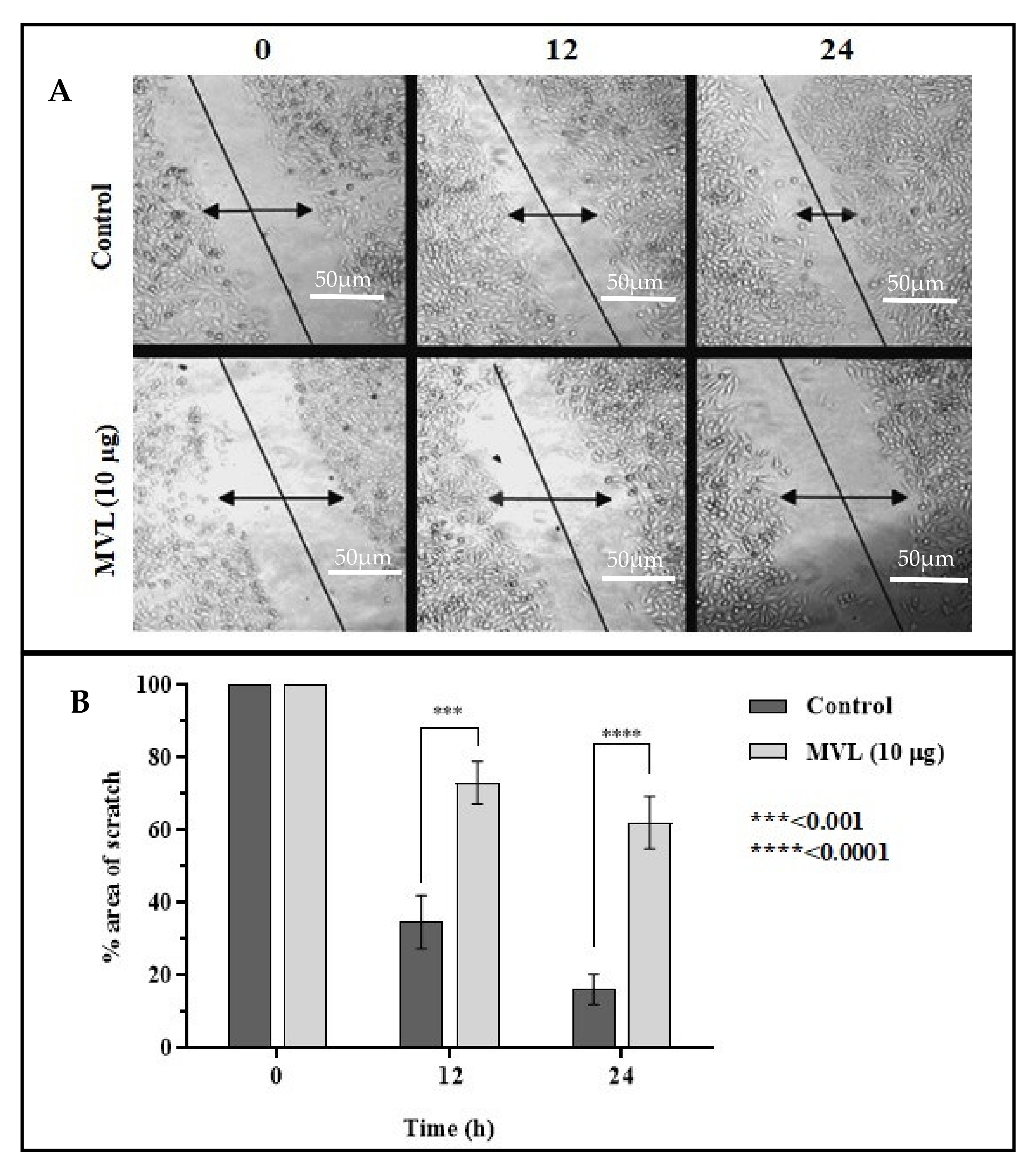
2.5. Cell Migration Assay
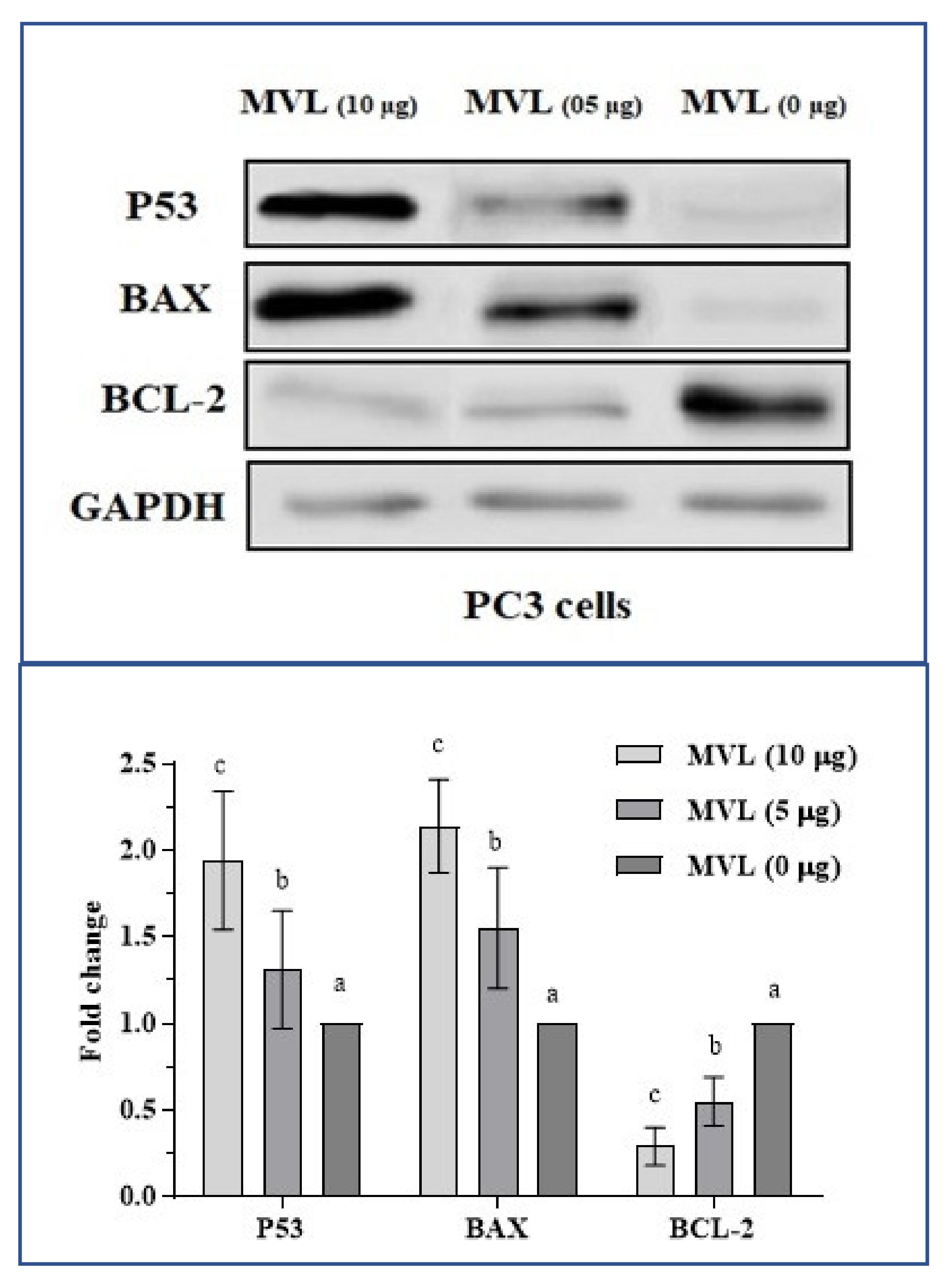
2.6. Molecular Expression Assessment
2.7. Toxicity Assessment (Safety Profiling)
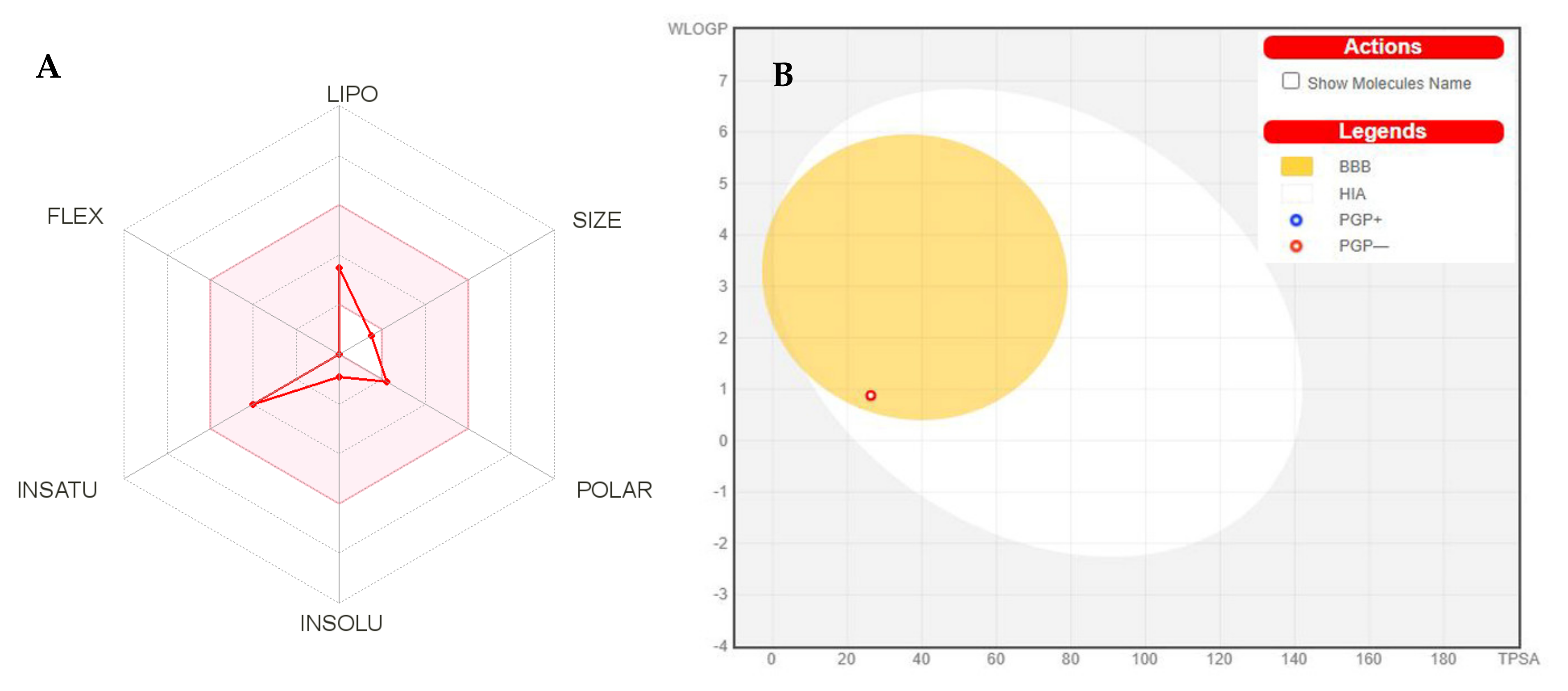
2.8. Pharmacokinetic and Toxicological Properties
2.8.1. Pharmacokinetic Properties
2.8.2. Toxicity Assessment
2.8.3. Cardiac Toxicity
2.9. Molecular Docking
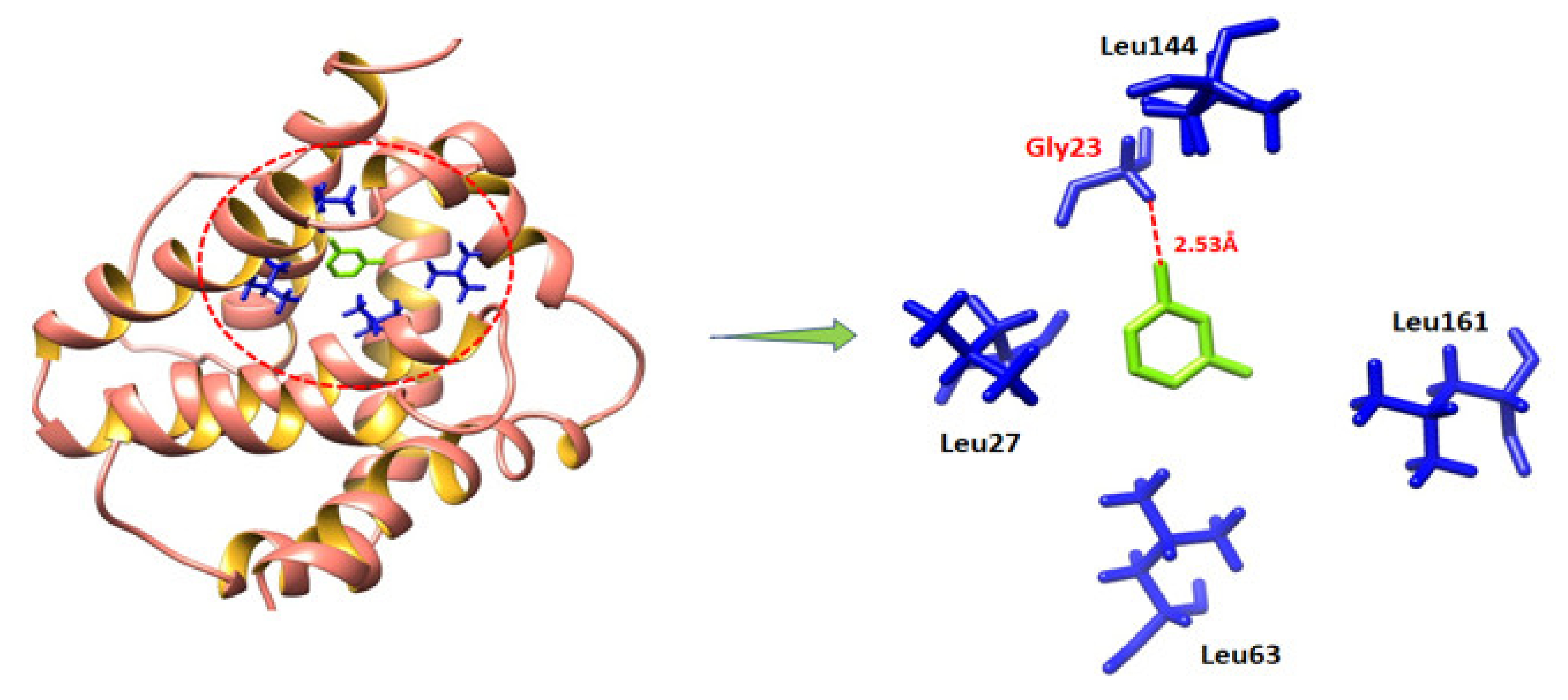
2.9.1. Molecular Docking and Binding Energy Analysis
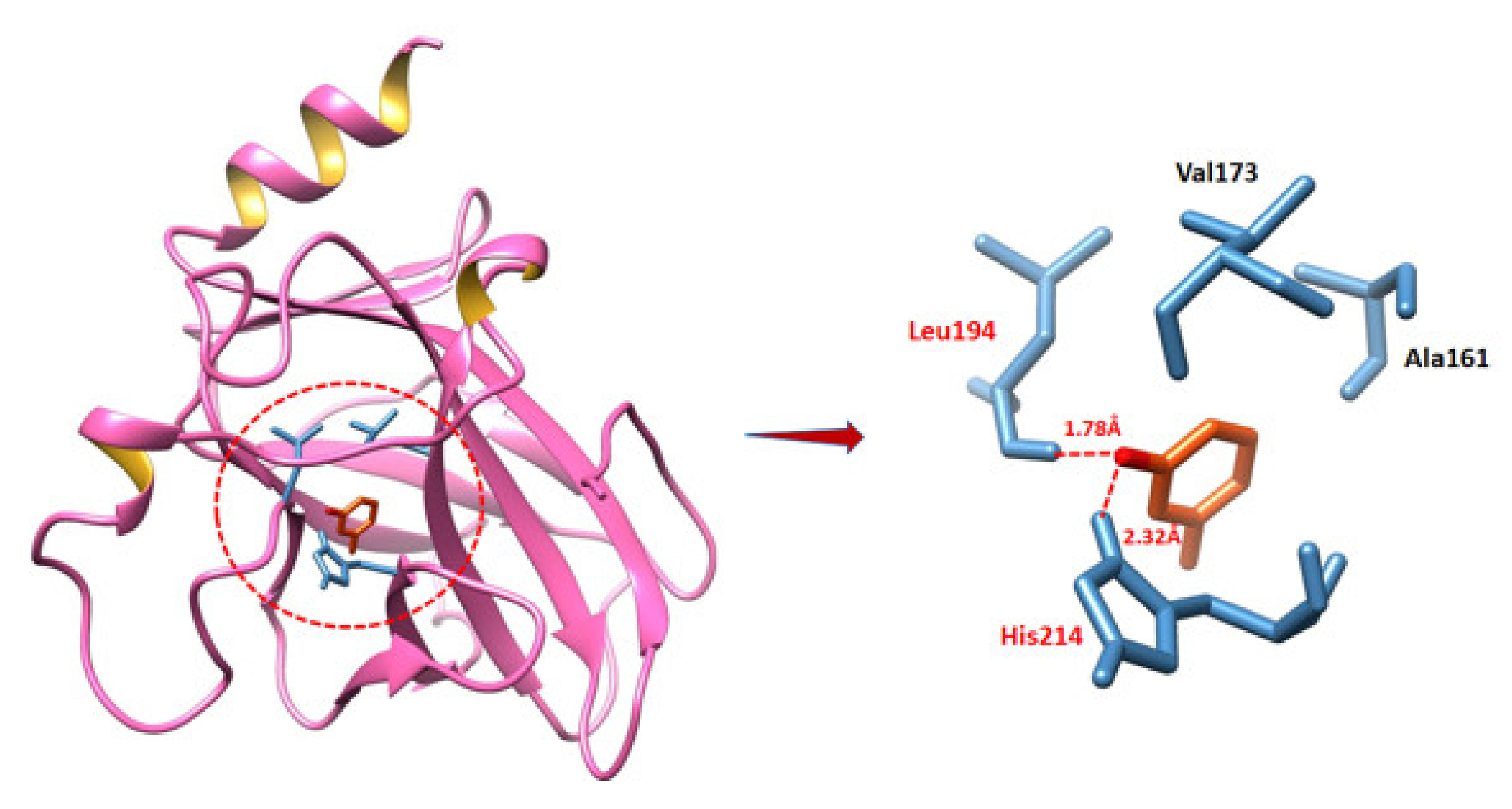
2.9.2. Binding Analyses of MVL against BAX and P53
3. Discussion
4. Materials and Methods
4.1. General
4.2. Soil Sample Collection
4.3. Isolation and Storage of SH-1312 Strain
4.4. Metal Stress and Normal Cultivation
4.5. HPLC Analysis and Purification of Stress Metabolite
4.6. Extraction and Isolation
4.7. Anti-Oxidant Evaluation
4.8. Cytotoxicity Analysis against Human Prostate PC3 Cell Line
4.9. In Vitro Wound Assay
4.10. Western Blotting
4.11. Comet Assay
4.12. ADMET Analysis
4.13. Prediction of Cardiac Toxicity
4.14. Molecular Docking
Designing of Ligand and Molecular Docking Simulation Using Autodock
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shams ul Hassan, S.; Jin, H.-z.; Abu-Izneid, T.; Rauf, A.; Ishaq, M.; Suleria, H.A.R. Stress-driven discovery in the natural products: A gateway towards new drugs. Biomed. Pharmacother. 2019, 109, 459–467. [Google Scholar] [CrossRef]
- Khalid, M.; Hassani, S.; Abdollahi, M. Metal-induced oxidative stress: An evidence-based update of advantages and disadvantages. Curr. Opin. Toxicol. 2020, 20–21, 55–68. [Google Scholar] [CrossRef]
- Pan, C.; Shi, Y.; Auckloo, B.N.; Hassan, S.S.u.; Akhter, N.; Wang, K.; Ye, Y.; Arthur Chen, C.-T.; Tao, X.; Wu, B. Isolation and Antibiotic Screening of Fungi from a Hydrothermal Vent Site and Characterization of Secondary Metabolites from a Penicillium Isolate. Mar. Biotechnol. 2017, 19, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Syed Shams ul, H.; Muhammad, I.; Wei-dong, Z.; Hui-Zi, J. An Overview of the Mechanisms of Marine Fungi-Derived Anti-Inflammatory and Anti-Tumor Agents and their Novel Role in Drug Targeting. Curr. Pharm. Des. 2021, 27, 2605–2614. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.S.; Shah, S.A.A.; Pan, C.; Fu, L.; Cao, X.; Shi, Y.; Wu, X.; Wang, K.; Wu, B. Production of an antibiotic enterocin from a marine actinobacteria strain H1003 by metal-stress technique with enhanced enrichment using response surface methodology. Pak. J. Pharm. Sci. 2017, 30, 313–324. [Google Scholar] [PubMed]
- Zhang, X.; Elliot, M.A. Unlocking the trove of metabolic treasures: Activating silent biosynthetic gene clusters in bacteria and fungi. Curr. Opin. Microbiol. 2019, 51, 9–15. [Google Scholar] [CrossRef]
- Dąbrowska, G.B.; Turkan, S.; Tylman-Mojżeszek, W.; Mierek-Adamska, A. In Silico Study of the RSH (RelA/SpoT Homologs) Gene Family and Expression Analysis in Response to PGPR Bacteria and Salinity in Brassica napus. Int. J. Mol. Sci. 2021, 22, 10666. [Google Scholar] [CrossRef]
- Molina-Risco, M.; Ibarra, O.; Faion-Molina, M.; Kim, B.; Septiningsih, E.M.; Thomson, M.J. Optimizing Agrobacterium-Mediated Transformation and CRISPR-Cas9 Gene Editing in the tropical japonica Rice Variety Presidio. Int. J. Mol. Sci. 2021, 22, 10909. [Google Scholar] [CrossRef]
- Alhadrami, H.A.; Thissera, B.; Hassan, M.H.A.; Behery, F.A.; Ngwa, C.J.; Hassan, H.M.; Pradel, G.; Abdelmohsen, U.R.; Rateb, M.E. Bio-Guided Isolation of Antimalarial Metabolites from the Coculture of Two Red Sea Sponge-Derived Actinokineospora and Rhodococcus spp. Mar. Drugs 2021, 19, 109. [Google Scholar] [CrossRef]
- Xie, Y.; Zhao, X.-C.; Hassan, S.; Zhen, X.-Y.; Ishaq, M.; Yan, S.; Yuan, X.; Li, H.-L.; Jin, H.-Z. One new sesquiterpene and one new iridoid derivative from Valeriana amurensis. Phytochem. Lett. 2019, 32, 6–9. [Google Scholar] [CrossRef]
- Khan, I.; Abbas, T.; Anjum, K.; Abbas, S.Q.; Shagufta, B.I.; Shah, S.A.A.; Akhter, N.; Hassan, S.S. Antimicrobial potential of aqueous extract of Camellia sinensis against representative microbes. Pak. J. Pharm. Sci. 2019, 32, 631–636. [Google Scholar]
- Makkar, R.; Behl, T.; Bungau, S.; Zengin, G.; Mehta, V.; Kumar, A.; Uddin, M.S.; Ashraf, G.M.; Abdel-Daim, M.M.; Arora, S.; et al. Nutraceuticals in Neurological Disorders. Int. J. Mol. Sci. 2020, 21, 4424. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhu, S.; Wu, G.; ul Hassan, S.S.; Xie, Y.; Ishaq, M.; Sun, Y.; Yan, S.-k.; Qian, X.-p.; Jin, H.-z. Chemical Constituents of Vernonia parishii. Chem. Nat. Compd. 2020, 56, 134–136. [Google Scholar] [CrossRef]
- Shah, S.A.A.; Hassan, S.S.u.; Bungau, S.; Si, Y.; Xu, H.; Rahman, M.H.; Behl, T.; Gitea, D.; Pavel, F.-M.; Corb Aron, R.A.; et al. Chemically Diverse and Biologically Active Secondary Metabolites from Marine Phylum chlorophyta. Mar Drugs 2020, 18, 493. [Google Scholar] [CrossRef] [PubMed]
- Tomm, H.A.; Ucciferri, L.; Ross, A.C. Advances in microbial culturing conditions to activate silent biosynthetic gene clusters for novel metabolite production. J. Ind. Microbiol. Biotechnol. 2019, 46, 1381–1400. [Google Scholar] [CrossRef] [PubMed]
- Mohammadipanah, F.; Kermani, F.; Salimi, F. Awakening the Secondary Metabolite Pathways of Promicromonospora kermanensis Using Physicochemical and Biological Elicitors. Appl. Biochem. Biotechnol. 2020, 192, 1224–1237. [Google Scholar] [CrossRef] [PubMed]
- Haferburg, G.; Groth, I.; Möllmann, U.; Kothe, E.; Sattler, I. Arousing sleeping genes: Shifts in secondary metabolism of metal tolerant actinobacteria under conditions of heavy metal stress. Biometals 2009, 22, 225. [Google Scholar] [CrossRef]
- Chmielowska-Bąk, J.; Deckert, J. Plant Recovery after Metal Stress—A Review. Plants 2021, 10, 450. [Google Scholar] [CrossRef]
- Liu, Z.Q. Bridging free radical chemistry with drug discovery: A promising way for finding novel drugs efficiently. Eur. J. Med. Chem. 2020, 189, 112020. [Google Scholar] [CrossRef]
- Memariani, Z.; Abbas, S.Q.; ul Hassan, S.S.; Ahmadi, A.; Chabra, A. Naringin and naringenin as anticancer agents and adjuvants in cancer combination therapy: Efficacy and molecular mechanisms of action, a comprehensive narrative review. Pharmacol. Res. 2020, 171, 105264. [Google Scholar] [CrossRef]
- Varejão, E.V.V.; Demuner, A.J.; Barbosa, L.C.d.A.; Barreto, R.W. Phytotoxic effects of metabolites from Alternaria euphorbiicola against its host plant Euphorbia heterophylla. Química Nova 2013, 36, 1004–1007. [Google Scholar] [CrossRef] [Green Version]
- Hassan, S.S.u.; Zhang, W.-D.; Jin, H.-z.; Basha, S.H.; Priya, S.V.S.S. In-silico anti-inflammatory potential of guaiane dimers from Xylopia vielana targeting COX-2. J. Biomol. Struct. Dyn. 2020, 1–15. [Google Scholar] [CrossRef]
- Cruz, J.V.; Serafim, R.B.; da Silva, G.M.; Giuliatti, S.; Rosa, J.M.C.; Araújo Neto, M.F.; Leite, F.H.A.; Taft, C.A.; da Silva, C.; Santos, C.B.R. Computational design of new protein kinase 2 inhibitors for the treatment of inflammatory diseases using QSAR, pharmacophore-structure-based virtual screening, and molecular dynamics. J. Mol. Model. 2018, 24, 225. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Zoete, V. A BOILED-Egg To Predict Gastrointestinal Absorption and Brain Penetration of Small Molecules. Chem. Med. Chem. 2016, 11, 1117–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Tian, Y.; Gao, Y.; Wu, F.; Luo, X.; Ju, X.; Liu, G. In silico Design of Novel HIV-1 NNRTIs Based on Combined Modeling Studies of Dihydrofuro[3,4-d]pyrimidines. Front. Chem. 2020, 8, 164. [Google Scholar] [CrossRef] [Green Version]
- Barclay, L.A.; Wales, T.E.; Garner, T.P.; Wachter, F.; Lee, S.; Guerra, R.M.; Stewart, M.L.; Braun, C.R.; Bird, G.H.; Gavathiotis, E.; et al. Inhibition of Pro-apoptotic BAX by a noncanonical interaction mechanism. Mol. Cell. 2015, 57, 873–886. [Google Scholar] [CrossRef] [Green Version]
- Joerger, A.C.; Ang, H.C.; Veprintsev, D.B.; Blair, C.M.; Fersht, A.R. Structures of p53 cancer mutants and mechanism of rescue by second-site suppressor mutations. J. Biol. Chem. 2005, 280, 16030–16037. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, I.; Luo, W.; Shoaib, R.M.; Li, G.L.; Shams Ul Hassan, S.; Yang, Z.H.; Xiao, X.; Tu, G.L.; Yan, S.K.; Ma, X.P.; et al. Guaiane-type sesquiterpenoids from Cinnamomum migao HW Li: And their anti-inflammatory activities. Phytochemistry 2021, 190, 112850. [Google Scholar] [CrossRef]
- Muhammad, I.; Shams Ul Hassan, S.; Cheung, S.; Li, X.; Wang, R.; Zhang, W.D.; Yan, S.K.; Zhang, Y.; Jin, H.Z. Phytochemical study of Ligularia subspicata and valuation of its anti-inflammatory activity. Fitoterapia 2021, 148, 104800. [Google Scholar] [CrossRef]
- Spengler, E.K.; Kleiner, D.E.; Fontana, R.J. Vemurafenib-induced granulomatous hepatitis. Hepatology 2017, 65, 745–748. [Google Scholar] [CrossRef] [Green Version]
- Truong, J.; Yan, A.T.; Cramarossa, G.; Chan, K.K. Chemotherapy-induced cardiotoxicity: Detection, prevention, and management. Can. J. Cardiol. 2014, 30, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Raschi, E.; Vasina, V.; Poluzzi, E.; De Ponti, F. The hERG K+ channel: Target and antitarget strategies in drug development. Pharm. Res. 2008, 57, 181–195. [Google Scholar] [CrossRef]
- Nielsen, E.; Ostergaard, G.; Larsen, J. Toxicological Risk Assessment of Chemicals: A Practical Guide; CRC Press: Florida, FL, USA, 2008. [Google Scholar]
- Majid, M.; Nasir, B.; Zahra, S.S.; Khan, M.R.; Mirza, B.; Haq, I.-u. Ipomoea batatas L. Lam. ameliorates acute and chronic inflammations by suppressing inflammatory mediators, a comprehensive exploration using in vitro and in vivo models. BMC Complementary Altern. Med. 2018, 18, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Stamm, A.; Reimers, K.; Strauß, S.; Vogt, P.; Scheper, T.; Pepelanova, I. In vitro wound healing assays–state of the art. BioNanoMaterials 2016, 17, 79–87. [Google Scholar] [CrossRef]
- Møller, P.; Azqueta, A.; Boutet-Robinet, E.; Koppen, G.; Bonassi, S.; Milić, M.; Gajski, G.; Costa, S.; Teixeira, J.P.; Costa Pereira, C.; et al. Minimum Information for Reporting on the Comet Assay (MIRCA): Recommendations for describing comet assay procedures and results. Nat. Protoc. 2020, 15, 3817–3826. [Google Scholar] [CrossRef] [PubMed]
- Braga, R.C.; Alves, V.M.; Silva, M.F.; Muratov, E.; Fourches, D.; Lião, L.M.; Tropsha, A.; Andrade, C.H. Pred-hERG: A novel web-accessible computational tool for predicting cardiac toxicity. Mol. Inform. 2015, 34, 698–701. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | IC50 (µg/mL) | |||
|---|---|---|---|---|
| DPPH Scavenging | NO Inhibition | OH● Inhibition | Iron Chelation | |
| MVL | 19.65 ± 5.7 *** | 15.49 ± 4.8 **** | 19.65 ± 5.22 *** | 19.38 ± 7.11 *** |
| Ascorbic acid | 6.52 ± 4.92 | 8.44 ± 4.17 | -- | -- |
| Gallic acid | -- | -- | 6.26 ± 6.39 | -- |
| EDTA | -- | -- | -- | 10.20 ± 6.54 |
| 1% DMSO | -- | -- | -- | -- |
| Compound | IC50 (µg/mL) | ||
|---|---|---|---|
| 24 h | 48 h | 72 h | |
| MVL | 35.81 ± 4.2 *** | 23.29 ± 3.8 **** | 16.25 ± 6.5 **** |
| Cabazitaxel | 21.16 ± 5.1 | 15.09 ± 5.7 | 9.25 ± 3.4 |
| Sample | Comet Length (µm) | Head Length (µm) | Tail Length (µm) | % DNA in Head | % DNA in Tail | Tail Moment (µm) |
|---|---|---|---|---|---|---|
| Control | 40.4 ± 4.2 | 35.9 ± 1.8 | 4.5 ± 0.5 | 88.8 ± 2.1 | 11.2 ± 1.3 β | 0.11 ± 0.04 β |
| EMS (20 µg/mL) | 42.6 ± 3.6 | 24.1 ± 2.7 | 18.5 ± 1.4 | 56.6 ± 3.5 | 43.4 ± 1.8 ¥ | 1.37 ± 0.11 ¥ |
| MVL (10 µg/mL) | 42.6 ± 3.1 | 36.7 ± 2.5 | 5.9 ± 0.3 | 86.1 ± 2.8 | 13.9 ± 1.8 β | 0.11 ± 0.03 β |
| MVL (20 µg/mL) | 41.4 ± 2.4 | 31.7 ± 1.8 | 8.5 ± 1.3 | 80.6 ± 1.8 | 19.4 ± 2.6 β, ¥ | 0.24 ± 0.03 β, ¥ |
| Properties | Parameters | MVL |
|---|---|---|
| Physicochemical properties | MW a (g/mol) | 112.13 |
| Rotatable bonds | 0 | |
| HBA b | 2 | |
| HBD c | 0 | |
| Fraction Csp3 | 0.50 | |
| TPSA d | 26.30 | |
| Lipophilicity Log Po/w | iLOGP | 1.54 |
| XLOGP3 | 0.60 | |
| MLOGP | 0.88 | |
| Consensus | 1.07 |
| Properties | Parameters | MVL |
|---|---|---|
| Absorption | Water Solubility | −0.509 |
| GI a | 100 | |
| Log Kp (Skin permeation) cm/s | −6.56 | |
| P-gp substrate | No | |
| Distribution | BBB b | −0.031 |
| CNS permeation (Log PS) | −2.633 | |
| VD c (human) | −0.037 | |
| Metabolism | CYP1A2 inhibitor | No |
| CYP2C19 inhibitor | No | |
| CYP2C9 inhibitor | No | |
| CYP2D6 inhibitor | No | |
| CYP3A4 inhibitor | No | |
| Excretion | Total Clearance (log mL/min/kg) | 0.814 |
| Renal OCT2 substrate | No |
| Toxicity | Parameters | MVL |
|---|---|---|
| Human | Ames toxicity | No |
| hERG I inhibitor | No | |
| hERG II inhibitor | No | |
| Hepatotoxicity | No | |
| Max. tolerated dose (human) (log mg/kg/day) | 1.01 | |
| Oral Rat | Oral Toxicity (LD50) (mg/kg) | 1890 |
| Oral Toxicity classification * | IV | |
| Environmental | Daphnia magna LC50 -Log10 (mol/L) | 3.347 |
| Bioaccumulation factor Log10 (BCF) | 0.487 | |
| Tetrahymena pyriformis IGC50 -Log10 (mol/L) | −0.867 | |
| Fathead Minnow LC50 Log10 (mmol/L) | 0.452 |
| Docking Complexes | Binding Energy (Kcal/mol) |
|---|---|
| P53_Ligand | −5.6 |
| BAX_Ligand | −6.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, S.S.u.; Muhammad, I.; Abbas, S.Q.; Hassan, M.; Majid, M.; Jin, H.-Z.; Bungau, S. Stress Driven Discovery of Natural Products From Actinobacteria with Anti-Oxidant and Cytotoxic Activities Including Docking and ADMET Properties. Int. J. Mol. Sci. 2021, 22, 11432. https://doi.org/10.3390/ijms222111432
Hassan SSu, Muhammad I, Abbas SQ, Hassan M, Majid M, Jin H-Z, Bungau S. Stress Driven Discovery of Natural Products From Actinobacteria with Anti-Oxidant and Cytotoxic Activities Including Docking and ADMET Properties. International Journal of Molecular Sciences. 2021; 22(21):11432. https://doi.org/10.3390/ijms222111432
Chicago/Turabian StyleHassan, Syed Shams ul, Ishaq Muhammad, Syed Qamar Abbas, Mubashir Hassan, Muhammad Majid, Hui-Zi Jin, and Simona Bungau. 2021. "Stress Driven Discovery of Natural Products From Actinobacteria with Anti-Oxidant and Cytotoxic Activities Including Docking and ADMET Properties" International Journal of Molecular Sciences 22, no. 21: 11432. https://doi.org/10.3390/ijms222111432
APA StyleHassan, S. S. u., Muhammad, I., Abbas, S. Q., Hassan, M., Majid, M., Jin, H. -Z., & Bungau, S. (2021). Stress Driven Discovery of Natural Products From Actinobacteria with Anti-Oxidant and Cytotoxic Activities Including Docking and ADMET Properties. International Journal of Molecular Sciences, 22(21), 11432. https://doi.org/10.3390/ijms222111432