
Gene Doping with Peroxisome-Proliferator-Activated Receptor Beta/Delta Agonists Alters Immunity but Exercise Training Mitigates the Detection of Effects in Blood Samples

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. In Vitro Treatment of Human T Cells with GW0742 Increases Their FAO Potential and Polarization in Tregs
2.2. GW0742 Treatment Increases FAO Potential and Leads to Differential Changes in Treg Prevalence in Mouse Secondary Lymphoid Organs Depending on Training Status
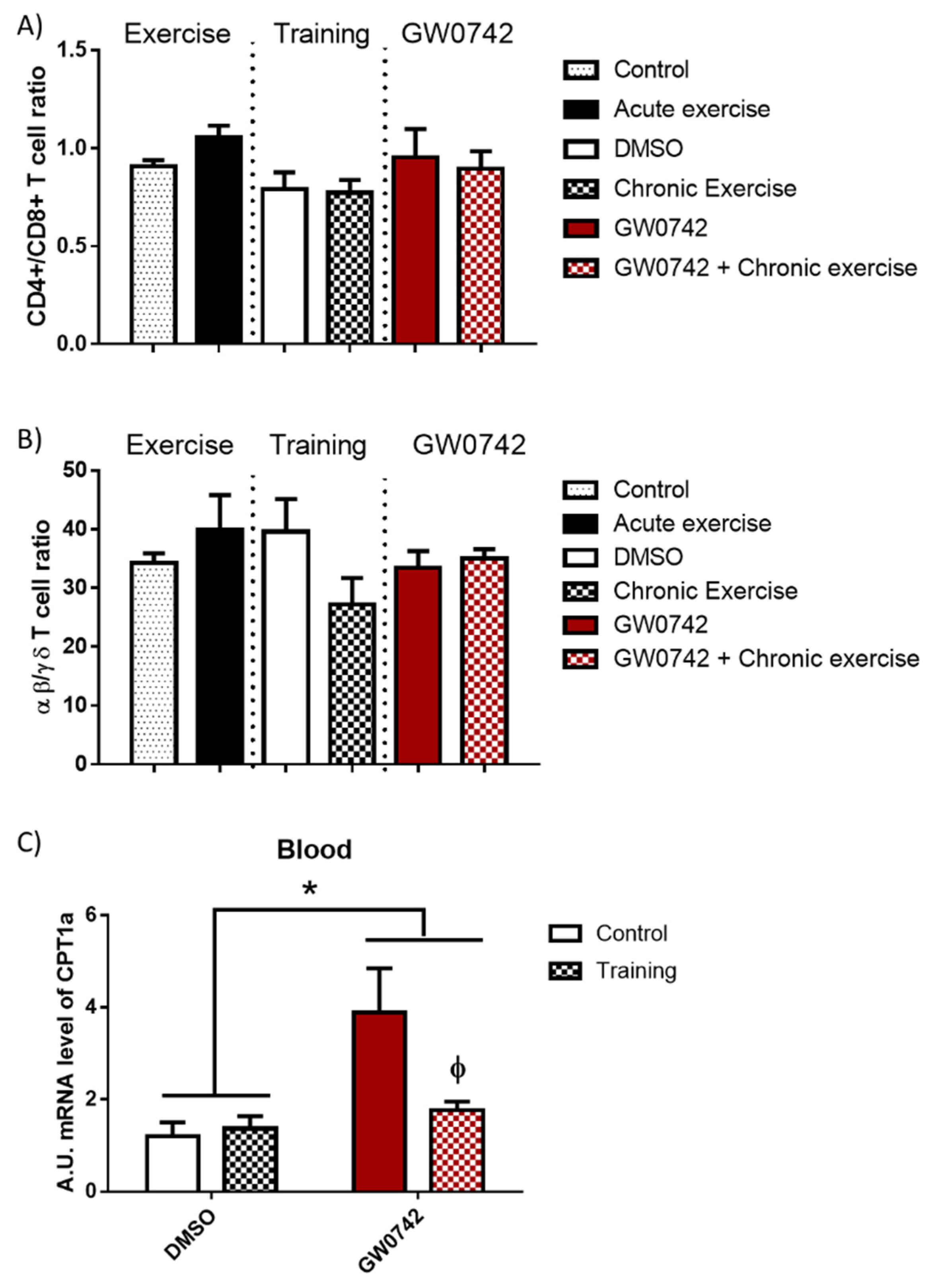
2.3. The Detection of GW0742 Effect on FAO Potential Is Masked in the Blood of Trained Mice
3. Discussion
4. Materials and Methods
4.1. Animal Experiments
4.1.1. Acute Treadmill Exercise
4.1.2. Physical Training and GW0742 Treatment of Mice
4.2. Mouse and Human T Cell Isolation and Treg Polarization
4.3. Measurement of β-Oxidation Using 3H-Labeled Palmitate
4.4. Cell Preparation and Flow Cytometry Analysis
4.5. RNA Extraction and Quantitative Real-Time PCR
4.6. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neels, J.G.; Grimaldi, P.A. Physiological functions of peroxisome proliferator-activated receptor beta. Physiol. Rev. 2014, 94, 795–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrieri, D.; Moon, H.Y.; van Praag, H. Exercise in a Pill: The Latest on Exercise-Mimetics. Brain Plast. 2017, 2, 153–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Gronde, T.; de Hon, O.; Haisma, H.J.; Pieters, T. Gene doping: An overview and current implications for athletes. Br. J. Sports Med. 2013, 47, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Trevisiol, S.; Moulard, Y.; Delcourt, V.; Jaubert, M.; Boyer, S.; Tendon, S.; Haryouli, H.; Taleb, W.; Caroff, M.; Chabot, B.; et al. Comprehensive characterization of the peroxisome proliferator activated receptor-delta agonist GW501516 for horse doping control analysis. Drug Test. Anal. 2021, 13, 1191–1202. [Google Scholar] [CrossRef]
- Pokrywka, A.; Cholbinski, P.; Kaliszewski, P.; Kowalczyk, K.; Konczak, D.; Zembron-Lacny, A. Metabolic modulators of the exercise response: Doping control analysis of an agonist of the peroxisome proliferator-activated receptor delta (GW501516) and 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR). J. Physiol. Pharmacol. 2014, 65, 469–476. [Google Scholar]
- Mothe-Satney, I.; Murdaca, J.; Sibille, B.; Rousseau, A.S.; Squillace, R.; Le Menn, G.; Rekima, A.; Larbret, F.; Pele, J.; Verhasselt, V.; et al. A role for Peroxisome Proliferator-Activated Receptor Beta in T cell development. Sci. Rep. 2016, 6, 34317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakaguchi, S.; Yamaguchi, T.; Nomura, T.; Ono, M. Regulatory T cells and immune tolerance. Cell 2008, 133, 775–787. [Google Scholar] [CrossRef] [Green Version]
- Newton, R.; Priyadharshini, B.; Turka, L.A. Immunometabolism of regulatory T cells. Nat. Immunol. 2016, 17, 618–625. [Google Scholar] [CrossRef] [PubMed]
- Le Garf, S.; Murdaca, J.; Mothe-Satney, I.; Sibille, B.; Le Menn, G.; Chinetti, G.; Neels, J.G.; Rousseau, A.S. Complementary Immunometabolic Effects of Exercise and PPARbeta/delta Agonist in the Context of Diet-Induced Weight Loss in Obese Female Mice. Int. J. Mol. Sci. 2019, 20, 5182. [Google Scholar] [CrossRef] [Green Version]
- Shaw, D.M.; Merien, F.; Braakhuis, A.; Dulson, D. T-cells and their cytokine production: The anti-inflammatory and immunosuppressive effects of strenuous exercise. Cytokine 2018, 104, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, M.; Bishop, N.C.; Stensel, D.J.; Lindley, M.R.; Mastana, S.S.; Nimmo, M.A. The anti-inflammatory effects of exercise: Mechanisms and implications for the prevention and treatment of disease. Nat. Rev. Immunol. 2011, 11, 607–615. [Google Scholar] [CrossRef]
- Padilha, C.S.; Figueiredo, C.; Minuzzi, L.G.; Chimin, P.; Deminice, R.; Kruger, K.; Rosa-Neto, J.C.; Lira, F.S. Immunometabolic responses according to physical fitness status and lifelong exercise during aging: New roads for exercise immunology. Ageing Res. Rev. 2021, 68, 101341. [Google Scholar] [CrossRef]
- Pearce, E.L.; Poffenberger, M.C.; Chang, C.H.; Jones, R.G. Fueling immunity: Insights into metabolism and lymphocyte function. Science 2013, 342, 1242454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blagih, J.; Coulombe, F.; Vincent, E.E.; Dupuy, F.; Galicia-Vazquez, G.; Yurchenko, E.; Raissi, T.C.; van der Windt, G.J.; Viollet, B.; Pearce, E.L.; et al. The energy sensor AMPK regulates T cell metabolic adaptation and effector responses in vivo. Immunity 2015, 42, 41–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blagih, J.; Hennequart, M.; Zani, F. Tissue Nutrient Environments and Their Effect on Regulatory T Cell Biology. Front. Immunol. 2021, 12, 637960. [Google Scholar] [CrossRef]
- Michalek, R.D.; Gerriets, V.A.; Jacobs, S.R.; Macintyre, A.N.; MacIver, N.J.; Mason, E.F.; Sullivan, S.A.; Nichols, A.G.; Rathmell, J.C. Cutting edge: Distinct glycolytic and lipid oxidative metabolic programs are essential for effector and regulatory CD4+ T cell subsets. J. Immunol. 2011, 186, 3299–3303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerriets, V.A.; Kishton, R.J.; Nichols, A.G.; Macintyre, A.N.; Inoue, M.; Ilkayeva, O.; Winter, P.S.; Liu, X.; Priyadharshini, B.; Slawinska, M.E.; et al. Metabolic programming and PDHK1 control CD4+ T cell subsets and inflammation. J. Clin. Investig. 2015, 125, 194–207. [Google Scholar] [CrossRef] [PubMed]
- Procaccini, C.; Carbone, F.; Di Silvestre, D.; Brambilla, F.; De Rosa, V.; Galgani, M.; Faicchia, D.; Marone, G.; Tramontano, D.; Corona, M.; et al. The Proteomic Landscape of Human Ex Vivo Regulatory and Conventional T Cells Reveals Specific Metabolic Requirements. Immunity 2016, 44, 712. [Google Scholar] [CrossRef] [Green Version]
- Raud, B.; Roy, D.G.; Divakaruni, A.S.; Tarasenko, T.N.; Franke, R.; Ma, E.H.; Samborska, B.; Hsieh, W.Y.; Wong, A.H.; Stuve, P.; et al. Etomoxir Actions on Regulatory and Memory T Cells Are Independent of Cpt1a-Mediated Fatty Acid Oxidation. Cell Metab. 2018, 28, 504–515.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, R.J.; Campbell, J.P.; Gleeson, M.; Kruger, K.; Nieman, D.C.; Pyne, D.B.; Turner, J.E.; Walsh, N.P. Can exercise affect immune function to increase susceptibility to infection? Exerc. Immunol. Rev. 2020, 26, 8–22. [Google Scholar] [PubMed]
- Belhaj, M.R.; Lawler, N.G.; Hoffman, N.J. Metabolomics and Lipidomics: Expanding the Molecular Landscape of Exercise Biology. Metabolites 2021, 11, 151. [Google Scholar] [CrossRef] [PubMed]
- Frandsen, J.; Poggi, A.I.; Ritz, C.; Larsen, S.; Dela, F.; Helge, J.W. Peak Fat Oxidation Rate Is Closely Associated With Plasma Free Fatty Acid Concentrations in Women; Similar to Men. Front. Physiol. 2021, 12, 696261. [Google Scholar] [CrossRef] [PubMed]
- Frandsen, J.; Vest, S.D.; Ritz, C.; Larsen, S.; Dela, F.; Helge, J.W. Plasma free fatty acid concentration is closely tied to whole body peak fat oxidation rate during repeated exercise. J. Appl. Physiol. 2019, 126, 1563–1571. [Google Scholar] [CrossRef] [PubMed]
- Andersson Hall, U.; Edin, F.; Pedersen, A.; Madsen, K. Whole-body fat oxidation increases more by prior exercise than overnight fasting in elite endurance athletes. Appl. Physiol. Nutr. Metab. 2016, 41, 430–437. [Google Scholar] [CrossRef] [Green Version]
- Astolfi, T.; Crettaz von Roten, F.; Kayser, B.; Saugy, M.; Faiss, R. The Influence of Training Load on Hematological Athlete Biological Passport Variables in Elite Cyclists. Front. Sports Act. Living 2021, 3, 618285. [Google Scholar] [CrossRef] [PubMed]
- Takata, Y.; Liu, J.; Yin, F.; Collins, A.R.; Lyon, C.J.; Lee, C.H.; Atkins, A.R.; Downes, M.; Barish, G.D.; Evans, R.M.; et al. PPARdelta-mediated antiinflammatory mechanisms inhibit angiotensin II-accelerated atherosclerosis. Proc. Natl. Acad. Sci. USA 2008, 105, 4277–4282. [Google Scholar] [CrossRef] [Green Version]
- Dorneles, G.P.; Dos Passos, A.A.Z.; Romao, P.R.T.; Peres, A. New Insights about Regulatory T Cells Distribution and Function with Exercise: The Role of Immunometabolism. Curr. Pharm. Des. 2020, 26, 979–990. [Google Scholar] [CrossRef]
- Dominguez-Villar, M.; Hafler, D.A. Regulatory T cells in autoimmune disease. Nat. Immunol. 2018, 19, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Sharif, K.; Watad, A.; Bragazzi, N.L.; Lichtbroun, M.; Amital, H.; Shoenfeld, Y. Physical activity and autoimmune diseases: Get moving and manage the disease. Autoimmun. Rev. 2018, 17, 53–72. [Google Scholar] [CrossRef]
- Feuerer, M.; Herrero, L.; Cipolletta, D.; Naaz, A.; Wong, J.; Nayer, A.; Lee, J.; Goldfine, A.B.; Benoist, C.; Shoelson, S.; et al. Lean, but not obese, fat is enriched for a unique population of regulatory T cells that affect metabolic parameters. Nat. Med. 2009, 15, 930–939. [Google Scholar] [CrossRef]
- Bapat, S.P.; Myoung Suh, J.; Fang, S.; Liu, S.; Zhang, Y.; Cheng, A.; Zhou, C.; Liang, Y.; LeBlanc, M.; Liddle, C.; et al. Depletion of fat-resident Treg cells prevents age-associated insulin resistance. Nature 2015, 528, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Jiang, P.; Wei, S.; Xu, X.; Wang, J. Regulatory T cells in tumor microenvironment: New mechanisms, potential therapeutic strategies and future prospects. Mol. Cancer 2020, 19, 116. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Franco, F.; Tsui, Y.C.; Xie, X.; Trefny, M.P.; Zappasodi, R.; Mohmood, S.R.; Fernandez-Garcia, J.; Tsai, C.H.; Schulze, I.; et al. CD36-mediated metabolic adaptation supports regulatory T cell survival and function in tumors. Nat. Immunol. 2020, 21, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Mothe-Satney, I.; Piquet, J.; Murdaca, J.; Sibille, B.; Grimaldi, P.A.; Neels, J.G.; Rousseau, A.S. Peroxisome Proliferator Activated Receptor Beta (PPARbeta) activity increases the immune response and shortens the early phases of skeletal muscle regeneration. Biochimie 2017, 136, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, A.S.; Murdaca, J.; Le Menn, G.; Sibille, B.; Wahli, W.; Le Garf, S.; Chinetti, G.; Neels, J.G.; Mothe-Satney, I. Invalidation of the Transcriptional Modulator of Lipid Metabolism PPARbeta/delta in T Cells Prevents Age-Related Alteration of Body Composition and Loss of Endurance Capacity. Front. Physiol. 2021, 12, 587753. [Google Scholar] [CrossRef] [PubMed]
- Le Garf, S.; Sibille, B.; Mothe-Satney, I.; Eininger, C.; Fauque, P.; Murdaca, J.; Chinetti, G.; Neels, J.G.; Rousseau, A.S. Alpha-lipoic acid supplementation increases the efficacy of exercise- and diet-induced obesity treatment and induces immunometabolic changes in female mice and women. FASEB J. 2021, 35, e21312. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sibille, B.; Mothe-Satney, I.; Le Menn, G.; Lepouse, D.; Le Garf, S.; Baudoin, E.; Murdaca, J.; Moratal, C.; Lamghari, N.; Chinetti, G.; et al. Gene Doping with Peroxisome-Proliferator-Activated Receptor Beta/Delta Agonists Alters Immunity but Exercise Training Mitigates the Detection of Effects in Blood Samples. Int. J. Mol. Sci. 2021, 22, 11497. https://doi.org/10.3390/ijms222111497
Sibille B, Mothe-Satney I, Le Menn G, Lepouse D, Le Garf S, Baudoin E, Murdaca J, Moratal C, Lamghari N, Chinetti G, et al. Gene Doping with Peroxisome-Proliferator-Activated Receptor Beta/Delta Agonists Alters Immunity but Exercise Training Mitigates the Detection of Effects in Blood Samples. International Journal of Molecular Sciences. 2021; 22(21):11497. https://doi.org/10.3390/ijms222111497
Chicago/Turabian StyleSibille, Brigitte, Isabelle Mothe-Satney, Gwenaëlle Le Menn, Doriane Lepouse, Sébastien Le Garf, Elodie Baudoin, Joseph Murdaca, Claudine Moratal, Noura Lamghari, Giulia Chinetti, and et al. 2021. "Gene Doping with Peroxisome-Proliferator-Activated Receptor Beta/Delta Agonists Alters Immunity but Exercise Training Mitigates the Detection of Effects in Blood Samples" International Journal of Molecular Sciences 22, no. 21: 11497. https://doi.org/10.3390/ijms222111497
APA StyleSibille, B., Mothe-Satney, I., Le Menn, G., Lepouse, D., Le Garf, S., Baudoin, E., Murdaca, J., Moratal, C., Lamghari, N., Chinetti, G., Neels, J. G., & Rousseau, A. -S. (2021). Gene Doping with Peroxisome-Proliferator-Activated Receptor Beta/Delta Agonists Alters Immunity but Exercise Training Mitigates the Detection of Effects in Blood Samples. International Journal of Molecular Sciences, 22(21), 11497. https://doi.org/10.3390/ijms222111497