
Corneal Lymphangiogenesis: Current Pathophysiological Understandings and Its Functional Role in Ocular Surface Disease


Abstract
:1. Cytokines and Molecular Markers for Lymphangiogenesis
1.1. Vascular Endothelial Growth Factor (VEGF)s; Focused on VEGF-C and -D
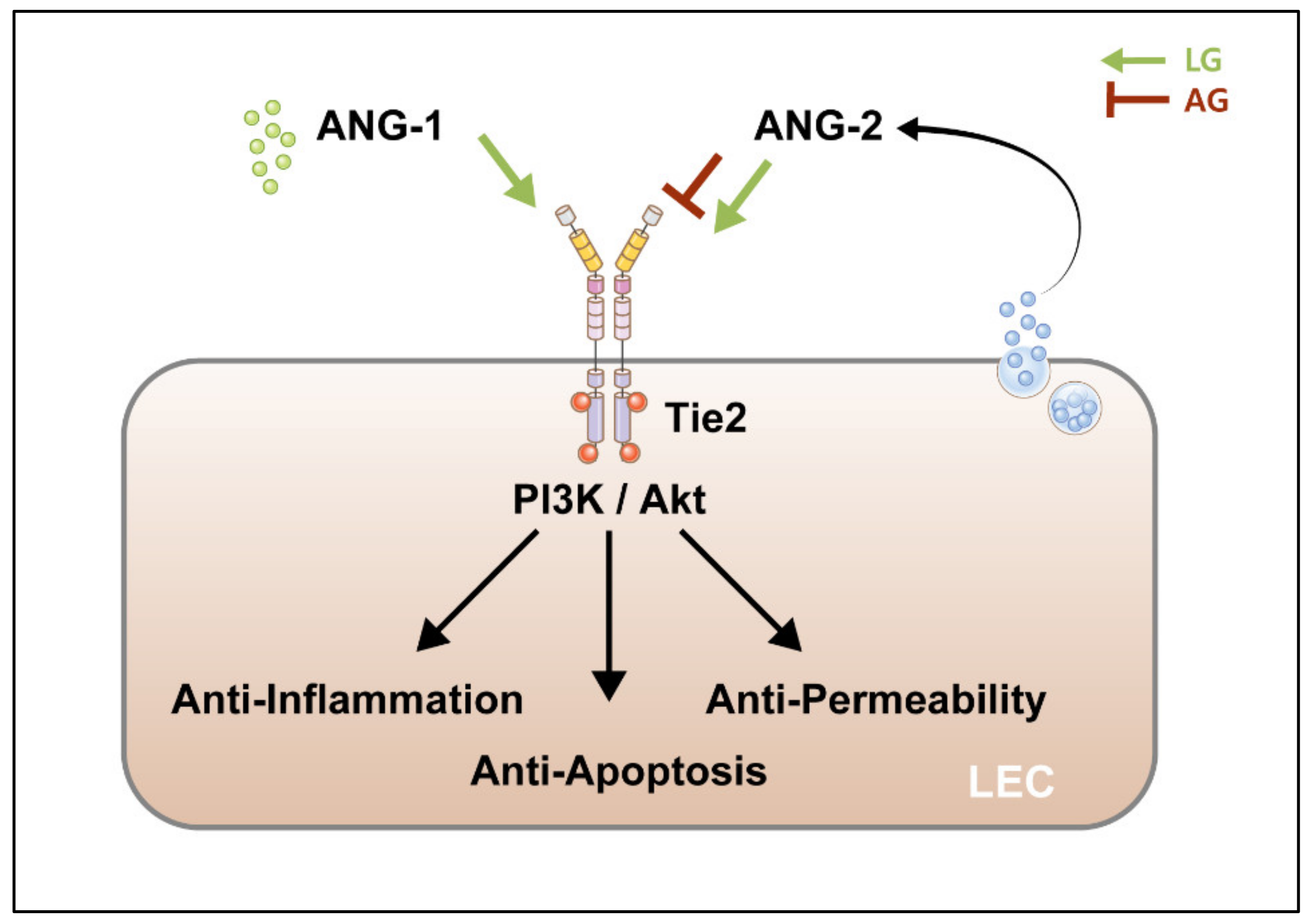
1.2. Angiopoietins (ANG)
1.3. Fibroblast Growth Factor2 (FGF2)
1.4. Hepatocyte Growth Factor (HGF), Platelet-Derived Growth Factors (PDGF), Insulin-Like Growth Factors (IGF), Endothelin, and Transforming Growth Factor (TGF)-β
1.5. Other Factors That Induce Lymphangiogenesis
2. Lymphatic Endothelial Cell (LEC) Markers
- PROX1In a knockout mouse model, Prox1 deletion resulted in complete loss of LV. Although LEC buds from veins were found morphologically, they did not express LEC markers and failed to migrate further under Prox1 knockdown conditions [65,66]. In accordance with the mouse model, Prox1 overexpression in human VECs showed reduced expression of VEC-specific genes with the upregulation of LEC-specific genes [2,62].
- LYVE-1LYVE-1 may be the most widely used marker for LEC research. LYVE-1 is the first marker of LEC development, and interestingly, it is expressed in a polarized manner in the venous endothelium since early lymphatic development. In mammals, LYVE-1 is mainly expressed in lymphatic capillaries and is downregulated in large LVs [2,67]. The functional role of LYVE-1 in the regulation of lymphatic development during embryogenesis and lymphangiogenesis in adults remains unclear.
- PodoplaninAlthough podoplanin is widely used as an LEC marker, it is also highly expressed in various cells, including LECs, podocytes, keratinocytes, and alveolar cells in the lungs. Podoplanin knockout mice showed paw lymphedema and abnormal lymphatic function and pattern, which may reflect impaired LEC migration [68].
- VEGFR3 and VEGF-C/D: see Section 1.1
- Chemokine (C-C motif) ligand 21CCL21 is secreted by LECs but not by VECs [69] and interacts with the CC chemokine receptor 7 (CCR-7), which is expressed on the surface of immune cells. It works as a guide to the immune cells, mainly dendritic cells bearing antigens and homing from the tissues into the LVs and the secondary lymphatic organs; thus, it plays an important role in immunoregulatory and inflammatory processes. CCL21 has also been shown to enhance LN metastasis in CCR-7-expressing malignant melanoma cells [70].
- DesmoplakinDesmoplakin is a cytoplasmic anchor protein of LEC adherens junctions that connect intermediate filaments to the plasma membrane. Desmoplakin is not expressed in VECs [71].
3. Cytokines Promote or Inhibit Lymphangiogenesis in Various Disease Models
4. Lymphangiogenesis in Ocular Surface Disease
4.1. Lymphangiogenesis in Allograft Rejection
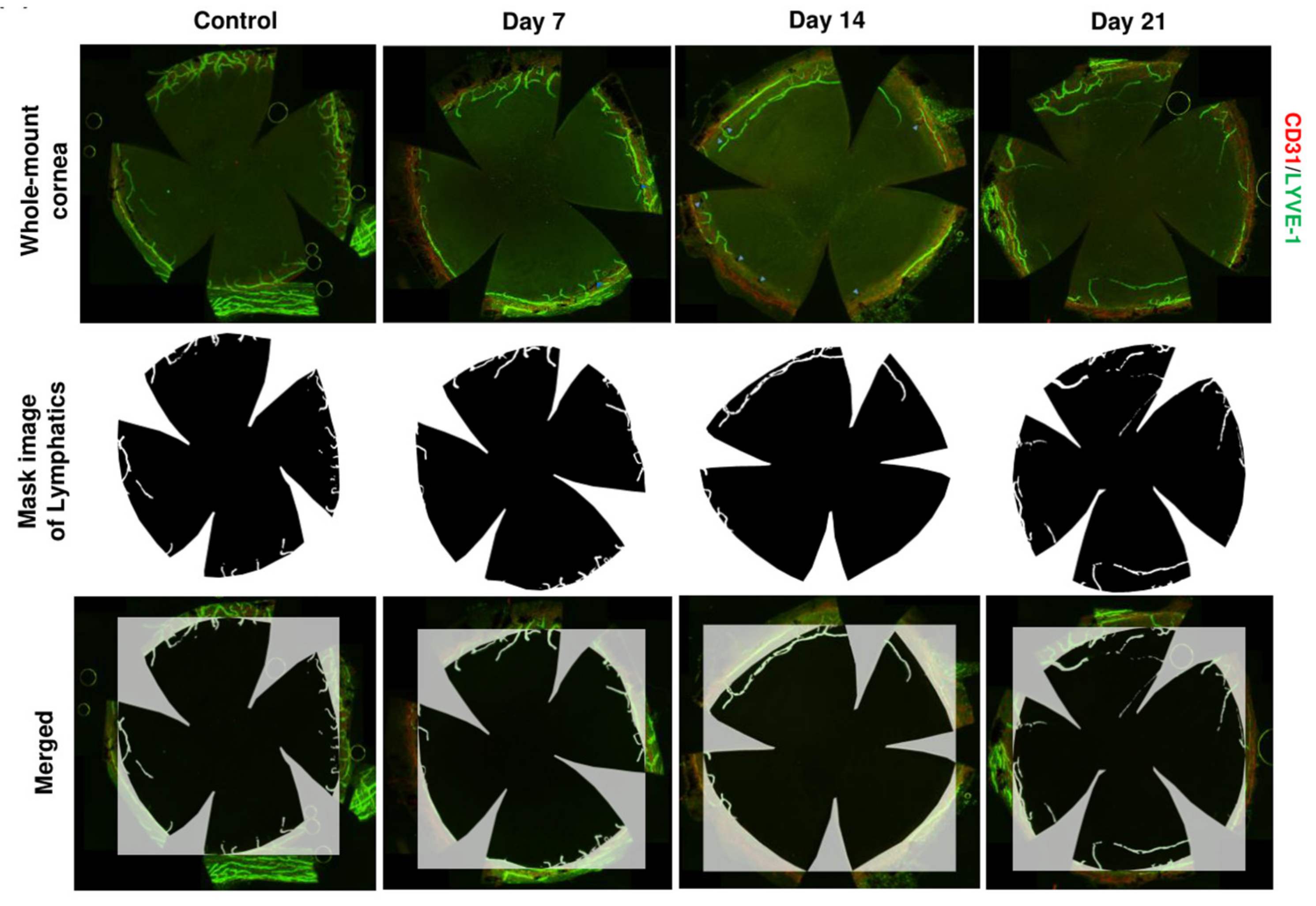
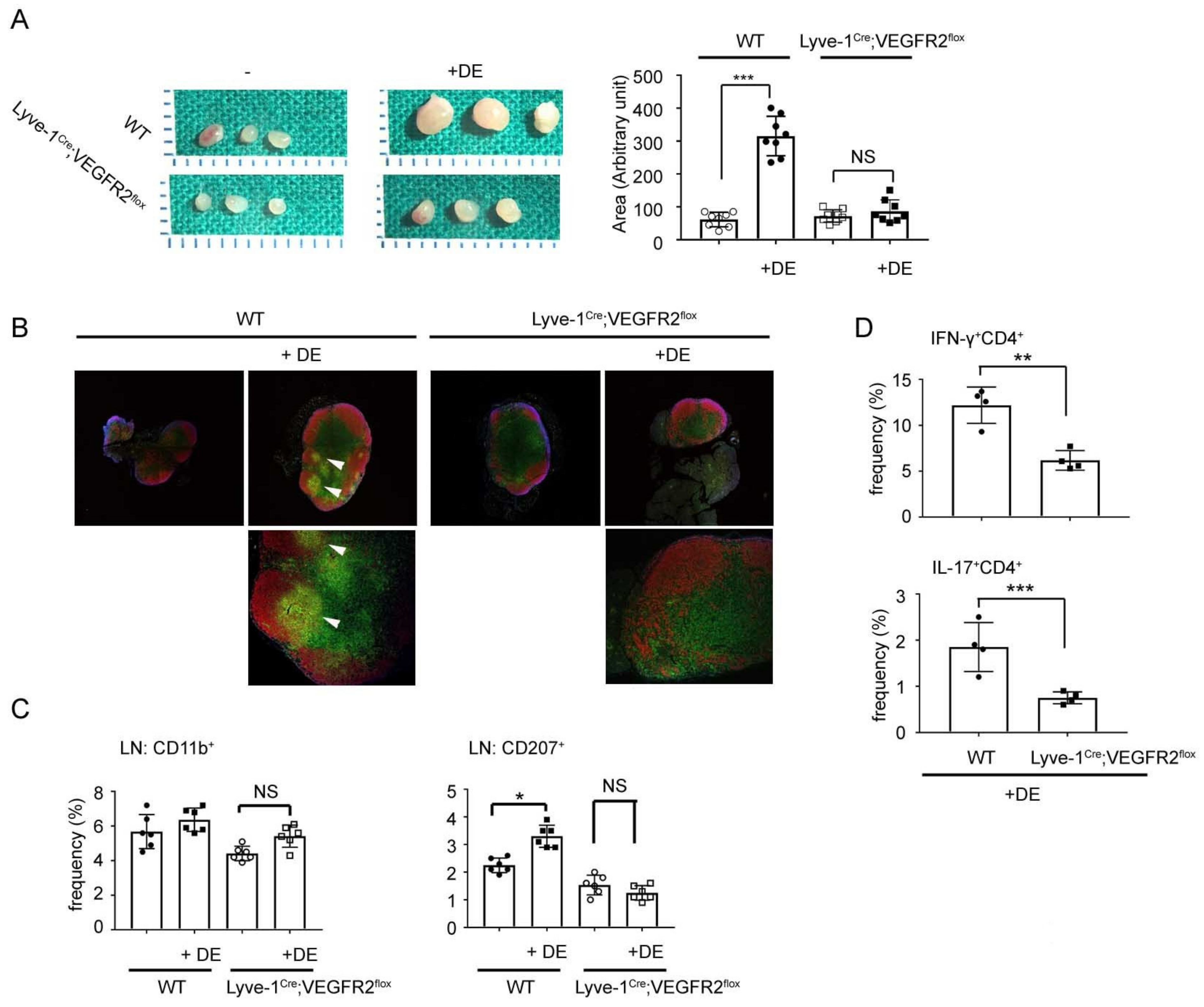
4.2. Lymphangiogenesis in Dry Eye Disease
4.3. Lymphangiogenesis in Other Ocular Diseases
5. Inhibition of Lymphangiogenesis
5.1. Inhibition of VEGFs
5.2. VEGFR-1/2 Inhibition
5.3. VEGFR-3 Inhibition
5.4. Non-VEGFs-Based Lymphangiogenesis Inhibition
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sáinz-Jaspeado, M.; Claesson-Welsh, L. Cytokines regulating lymphangiogenesis. Curr. Opin. Immunol. 2018, 53, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Alitalo, K.; Tammela, T.; Petrova, T.V. Lymphangiogenesis in development and human disease. Nature 2005, 438, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Hägerling, R.; Pollmann, C.; Andreas, M.; Schmidt, C.; Nurmi, H.; Adams, R.H.; Alitalo, K.; Andresen, V.; Schulte-Merker, S.; Kiefer, F. A novel multistep mechanism for initial lymphangiogenesis in mouse embryos based on ultramicroscopy. EMBO J. 2013, 32, 629–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Corral, I.; Ulvmar, M.H.; Stanczuk, L.; Tatin, F.; Kizhatil, K.; John, S.W.; Alitalo, K.; Ortega, S.; Makinen, T. Nonvenous origin of dermal lymphatic vasculature. Circ. Res. 2015, 116, 1649–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potente, M.; Mäkinen, T. Vascular heterogeneity and specialization in development and disease. Nat. Rev. Mol. Cell Biol. 2017, 18, 477–494. [Google Scholar] [CrossRef]
- Vaahtomeri, K.; Karaman, S.; Mäkinen, T.; Alitalo, K. Lymphangiogenesis guidance by paracrine and pericellular factors. Genes Dev. 2017, 31, 1615–1634. [Google Scholar] [CrossRef]
- Vempati, P.; Popel, A.S.; Mac Gabhann, F. Extracellular regulation of VEGF: Isoforms, proteolysis, and vascular patterning. Cytokine Growth Factor Rev. 2014, 25, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Moyon, D.; Pardanaud, L.; Bréant, C.; Karkkainen, M.J.; Alitalo, K.; Eichmann, A. Abnormal lymphatic vessel development in neuropilin 2 mutant mice. Development 2002, 129, 4797–4806. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Aspelund, A.; Alitalo, K. Lymphangiogenic factors, mechanisms, and applications. J. Clin. Investig. 2014, 124, 878–887. [Google Scholar] [CrossRef]
- Claesson-Welsh, L. VEGF receptor signal transduction—A brief update. Vasc. Pharmacol. 2016, 86, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Dellinger, M.T.; Meadows, S.M.; Wynne, K.; Cleaver, O.; Brekken, R.A. Vascular endothelial growth factor receptor-2 promotes the development of the lymphatic vasculature. PLoS ONE 2013, 8, e74686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karkkainen, M.J.; Haiko, P.; Sainio, K.; Partanen, J.; Taipale, J.; Petrova, T.V.; Jeltsch, M.; Jackson, D.G.; Talikka, M.; Rauvala, H.; et al. Vascular endothelial growth factor C is required for sprouting of the first lymphatic vessels from embryonic veins. Nat. Immunol. 2004, 5, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Mäkinen, T.; Veikkola, T.; Mustjoki, S.; Karpanen, T.; Catimel, B.; Nice, E.C.; Wise, L.; Mercer, A.; Kowalski, H.; Kerjaschki, D.; et al. Isolated lymphatic endothelial cells transduce growth, survival and migratory signals via the VEGF-C/D receptor VEGFR-3. EMBO J. 2001, 20, 4762–4773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coso, S.; Zeng, Y.; Opeskin, K.; Williams, E.D. Vascular endothelial growth factor receptor-3 directly interacts with phosphatidylinositol 3-kinase to regulate lymphangiogenesis. PLoS ONE 2012, 7, e39558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teichert, M.; Milde, L.; Holm, A.; Stanicek, L.; Gengenbacher, N.; Savant, S.; Ruckdeschel, T.; Hasanov, Z.; Srivastava, K.; Hu, J.; et al. Pericyte-expressed Tie2 controls angiogenesis and vessel maturation. Nat. Commun. 2017, 8, 16106. [Google Scholar] [CrossRef]
- Eklund, L.; Kangas, J.; Saharinen, P. Angiopoietin-Tie signalling in the cardiovascular and lymphatic systems. Clin. Sci. 2017, 131, 87–103. [Google Scholar] [CrossRef] [Green Version]
- Jeltsch, M.; Kaipainen, A.; Joukov, V.; Meng, X.; Lakso, M.; Rauvala, H.; Swartz, M.; Fukumura, D.; Jain, R.K.; Alitalo, K. Hyperplasia of lymphatic vessels in VEGF-C transgenic mice. Science 1997, 276, 1423–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, W.A.; Tzvetkova, D.; Nikolov, D.B. Structure of the angiopoietin-2 receptor binding domain and identification of surfaces involved in Tie2 recognition. Structure 2005, 13, 825–832. [Google Scholar] [CrossRef] [Green Version]
- Song, S.H.; Kim, K.L.; Lee, K.A.; Suh, W. Tie1 regulates the Tie2 agonistic role of angiopoietin-2 in human lymphatic endothelial cells. Biochem. Biophys. Res. Commun. 2012, 419, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Nurmi, H.; Appak, S.; Sabine, A.; Bovay, E.; Korhonen, E.A.; Orsenigo, F.; Lohela, M.; D’Amico, G.; Holopainen, T.; et al. Angiopoietin 2 regulates the transformation and integrity of lymphatic endothelial cell junctions. Genes Dev. 2014, 28, 1592–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morisada, T.; Oike, Y.; Yamada, Y.; Urano, T.; Akao, M.; Kubota, Y.; Maekawa, H.; Kimura, Y.; Ohmura, M.; Miyamoto, T.; et al. Angiopoietin-1 promotes LYVE-1-positive lymphatic vessel formation. Blood 2005, 105, 4649–4656. [Google Scholar] [CrossRef] [PubMed]
- Toyono, T.; Usui, T.; Yokoo, S.; Kimakura, M.; Nakagawa, S.; Yamagami, S.; Miyata, K.; Oike, Y.; Amano, S. Angiopoietin-like protein 2 is a potent hemangiogenic and lymphangiogenic factor in corneal inflammation. Investig. Ophthalmol. Vis. Sci. 2013, 54, 4278–4285. [Google Scholar] [CrossRef]
- Chauhan, S.K.; Lee, H.K.; Lee, H.S.; Park, E.Y.; Jeong, E.; Dana, R. PTK7+ Mononuclear Cells Express VEGFR2 and Contribute to Vascular Stabilization by Upregulating Angiopoietin-1. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1606–1615. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, G.; Sessa, R.; Kang, G.J.; Shi, M.; Ge, S.; Gong, A.J.; Wen, Y.; Chintharlapalli, S.; Chen, L. Angiopoietin-2 Blockade Promotes Survival of Corneal Transplants. Investig. Ophthalmol. Vis. Sci. 2017, 58, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, B.R.; Heinen, S.; Jeansson, M.; Ghosh, A.K.; Fatima, A.; Sung, H.K.; Onay, T.; Chen, H.; Yamaguchi, S.; Economides, A.N.; et al. A lymphatic defect causes ocular hypertension and glaucoma in mice. J. Clin. Investig. 2014, 124, 4320–4324. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Park, D.Y.; Bae, H.; Park, D.Y.; Kim, D.; Lee, C.K.; Song, S.; Chung, T.Y.; Lim, D.H.; Kubota, Y.; et al. Impaired angiopoietin/Tie2 signaling compromises Schlemm’s canal integrity and induces glaucoma. J. Clin. Investig. 2017, 127, 3877–3896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, R.; Ji, H.; Feng, N.; Zhang, Y.; Yang, X.; Andersson, P.; Sun, Y.; Tritsaris, K.; Hansen, A.J.; Dissing, S.; et al. Collaborative interplay between FGF-2 and VEGF-C promotes lymphangiogenesis and metastasis. Proc. Natl. Acad. Sci. USA 2012, 109, 15894–15899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, L.K.; Garcia-Cardeña, G.; Farnebo, F.; Fannon, M.; Chen, E.J.; Butterfield, C.; Moses, M.A.; Mulligan, R.C.; Folkman, J.; Kaipainen, A. Dose-dependent response of FGF-2 for lymphangiogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 11658–11663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, H.; Cao, R.; Brakenhielm, E.; Mäkinen, T.; Cao, Y.; Alitalo, K. Blockade of vascular endothelial growth factor receptor-3 signaling inhibits fibroblast growth factor-2-induced lymphangiogenesis in mouse cornea. Proc. Natl. Acad. Sci. USA 2002, 99, 8868–8873. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Tammela, T.; Yamamoto, M.; Anisimov, A.; Holopainen, T.; Kaijalainen, S.; Karpanen, T.; Lehti, K.; Ylä-Herttuala, S.; Alitalo, K. Notch restricts lymphatic vessel sprouting induced by vascular endothelial growth factor. Blood 2011, 118, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Zhang, X.; Luo, W.; Ge, H.; Sun, D.; Liu, P. Notch Signaling Pathway Is Involved in bFGF-Induced Corneal Lymphangiogenesis and Hemangiogenesis. J. Ophthalmol. 2019, 2019, 9613923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, J.H.; Lee, C.H.; Ji, Y.W.; Yeo, A.; Noh, H.; Song, I.; Kim, E.K.; Lee, H.K. Activation of Dll4/Notch Signaling and Hypoxia-Inducible Factor-1 Alpha Facilitates Lymphangiogenesis in Lacrimal Glands in Dry Eye. PLoS ONE 2016, 11, e0147846. [Google Scholar] [CrossRef] [PubMed]
- Platonova, N.; Miquel, G.; Regenfuss, B.; Taouji, S.; Cursiefen, C.; Chevet, E.; Bikfalvi, A. Evidence for the interaction of fibroblast growth factor-2 with the lymphatic endothelial cell marker LYVE-1. Blood 2013, 121, 1229–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Zhang, J.; Lao, X.; Jiang, H.; Yu, Y.; Deng, Y.; Zhong, J.; Liang, Y.; Xiong, L.; Deng, N. Construction of a disulfide-stabilized diabody against fibroblast growth factor-2 and the inhibition activity in targeting breast cancer. Cancer Sci. 2016, 107, 1141–1150. [Google Scholar] [CrossRef]
- Saito, Y.; Nakagami, H.; Morishita, R.; Takami, Y.; Kikuchi, Y.; Hayashi, H.; Nishikawa, T.; Tamai, K.; Azuma, N.; Sasajima, T.; et al. Transfection of human hepatocyte growth factor gene ameliorates secondary lymphedema via promotion of lymphangiogenesis. Circulation 2006, 114, 1177–1184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Xie, F.; Gao, W.; Yu, S.; Qiu, L.; Lin, W.; Sun, Y.; Jia, T. Expression of hepatocyte growth factor and c-Met in non-small-cell lung cancer and association with lymphangiogenesis. Mol. Med. Rep. 2015, 11, 2797–2804. [Google Scholar] [CrossRef] [Green Version]
- Onimaru, M.; Yonemitsu, Y.; Fujii, T.; Tanii, M.; Nakano, T.; Nakagawa, K.; Kohno, R.; Hasegawa, M.; Nishikawa, S.; Sueishi, K. VEGF-C regulates lymphangiogenesis and capillary stability by regulation of PDGF-B. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1685–H1696. [Google Scholar] [CrossRef] [PubMed]
- Kodama, M.; Kitadai, Y.; Sumida, T.; Ohnishi, M.; Ohara, E.; Tanaka, M.; Shinagawa, K.; Tanaka, S.; Yasui, W.; Chayama, K. Expression of platelet-derived growth factor (PDGF)-B and PDGF-receptor β is associated with lymphatic metastasis in human gastric carcinoma. Cancer Sci. 2010, 101, 1984–1989. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lv, J.; Zhang, T. Combination of IL-24 and cisplatin inhibits angiogenesis and lymphangiogenesis of cervical cancer xenografts in a nude mouse model by inhibiting VEGF, VEGF-C and PDGF-B. Oncol. Rep. 2015, 33, 2468–2476. [Google Scholar] [CrossRef] [Green Version]
- Björndahl, M.; Cao, R.; Nissen, L.J.; Clasper, S.; Johnson, L.A.; Xue, Y.; Zhou, Z.; Jackson, D.; Hansen, A.J.; Cao, Y. Insulin-like growth factors 1 and 2 induce lymphangiogenesis in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 15593–15598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achen, M.G.; Stacker, S.A. Tumor lymphangiogenesis and metastatic spread-new players begin to emerge. Int. J. Cancer 2006, 119, 1755–1760. [Google Scholar] [CrossRef]
- Spinella, F.; Garrafa, E.; Di Castro, V.; Rosanò, L.; Nicotra, M.R.; Caruso, A.; Natali, P.G.; Bagnato, A. Endothelin-1 stimulates lymphatic endothelial cells and lymphatic vessels to grow and invade. Cancer Res. 2009, 69, 2669–2676. [Google Scholar] [CrossRef] [Green Version]
- Caprara, V.; Scappa, S.; Garrafa, E.; Di Castro, V.; Rosanò, L.; Bagnato, A.; Spinella, F. Endothelin-1 regulates hypoxia-inducible factor-1α and -2α stability through prolyl hydroxylase domain 2 inhibition in human lymphatic endothelial cells. Life Sci. 2014, 118, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Clavin, N.W.; Avraham, T.; Fernandez, J.; Daluvoy, S.V.; Soares, M.A.; Chaudhry, A.; Mehrara, B.J. TGF-beta1 is a negative regulator of lymphatic regeneration during wound repair. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H2113–H2127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, J.M.; Nalbandian, A.; Mukouyama, Y.S. TGFβ signaling is required for sprouting lymphangiogenesis during lymphatic network development in the skin. Development 2013, 140, 3903–3914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.; Zhang, X.; Lu, Y.; Gong, L. TGFBIp mediates lymphatic sprouting in corneal lymphangiogenesis. J. Cell. Mol. Med. 2019, 23, 7602–7616. [Google Scholar] [CrossRef] [PubMed]
- Okanobo, A.; Chauhan, S.K.; Dastjerdi, M.H.; Kodati, S.; Dana, R. Efficacy of topical blockade of interleukin-1 in experimental dry eye disease. Am. J. Ophthalmol. 2012, 154, 63–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.; Chauhan, S.K.; El Annan, J.; Annan, J.E.; Sage, P.T.; Sharpe, A.H.; Dana, R. A novel function for programmed death ligand-1 regulation of angiogenesis. Am. J. Pathol. 2011, 178, 1922–1929. [Google Scholar] [CrossRef] [PubMed]
- Tsuru, S.; Ito, Y.; Matsuda, H.; Hosono, K.; Inoue, T.; Nakamoto, S.; Kurashige, C.; Mishima, T.; Tsujikawa, K.; Okamoto, H.; et al. RAMP1 signaling in immune cells regulates inflammation-associated lymphangiogenesis. Lab. Investig. 2020, 100, 738–750. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Im, S.T.; Wu, J.; Cho, C.S.; Jo, D.H.; Chen, Y.; Dana, R.; Kim, J.H.; Lee, S.M. Corneal lymphangiogenesis in dry eye disease is regulated by substance P/neurokinin-1 receptor system through controlling expression of vascular endothelial growth factor receptor 3. Ocul. Surf. 2021, 22, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Sabatino, F.; Di Zazzo, A.; De Simone, L.; Bonini, S. The Intriguing Role of Neuropeptides at the Ocular Surface. Ocul. Surf. 2017, 15, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Bignami, F.; Rama, P.; Ferrari, G. Substance P and its Inhibition in Ocular Inflammation. Curr. Drug Targets 2016, 17, 1265–1274. [Google Scholar] [CrossRef] [PubMed]
- Bignami, F.; Giacomini, C.; Lorusso, A.; Aramini, A.; Rama, P.; Ferrari, G. NK1 receptor antagonists as a new treatment for corneal neovascularization. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6783–6794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbariga, M.; Fonteyne, P.; Ostadreza, M.; Bignami, F.; Rama, P.; Ferrari, G. Substance P Modulation of Human and Murine Corneal Neovascularization. Investig. Ophthalmol. Vis. Sci. 2018, 59, 1305–1312. [Google Scholar] [CrossRef]
- Chauhan, S.K.; Jin, Y.; Goyal, S.; Lee, H.S.; Fuchsluger, T.A.; Lee, H.K.; Dana, R. A novel pro-lymphangiogenic function for Th17/IL-17. Blood 2011, 118, 4630–4634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Paiva, C.S.; Chotikavanich, S.; Pangelinan, S.B.; Pitcher, J.D., 3rd; Fang, B.; Zheng, X.; Ma, P.; Farley, W.J.; Siemasko, K.F.; Niederkorn, J.Y.; et al. IL-17 disrupts corneal barrier following desiccating stress. Mucosal Immunol. 2009, 2, 243–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Numasaki, M.; Fukushi, J.; Ono, M.; Narula, S.K.; Zavodny, P.J.; Kudo, T.; Robbins, P.D.; Tahara, H.; Lotze, M.T. Interleukin-17 promotes angiogenesis and tumor growth. Blood 2003, 101, 2620–2627. [Google Scholar] [CrossRef] [PubMed]
- Pickens, S.R.; Volin, M.V.; Mandelin, A.M., 2nd; Kolls, J.K.; Pope, R.M.; Shahrara, S. IL-17 contributes to angiogenesis in rheumatoid arthritis. J. Immunol. 2010, 184, 3233–3241. [Google Scholar] [CrossRef]
- Suryawanshi, A.; Veiga-Parga, T.; Reddy, P.B.; Rajasagi, N.K.; Rouse, B.T. IL-17A differentially regulates corneal vascular endothelial growth factor (VEGF)-A and soluble VEGF receptor 1 expression and promotes corneal angiogenesis after herpes simplex virus infection. J. Immunol. 2012, 188, 3434–3446. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, S.K.; Dohlman, T.H.; Dana, R. Corneal Lymphatics: Role in Ocular Inflammation as Inducer and Responder of Adaptive Immunity. J. Clin. Cell. Immunol. 2014, 5, 1000256. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Yuk, C.M.; Shin, K.; Lee, S.H. Interleukin-17A negatively regulates lymphangiogenesis in T helper 17 cell-mediated inflammation. Mucosal Immunol. 2018, 11, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Petrova, T.V.; Mäkinen, T.; Mäkelä, T.P.; Saarela, J.; Virtanen, I.; Ferrell, R.E.; Finegold, D.N.; Kerjaschki, D.; Ylä-Herttuala, S.; Alitalo, K. Lymphatic endothelial reprogramming of vascular endothelial cells by the Prox-1 homeobox transcription factor. EMBO J. 2002, 21, 4593–4599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalucka, J.; de Rooij, L.; Goveia, J.; Rohlenova, K.; Dumas, S.J.; Meta, E.; Conchinha, N.V.; Taverna, F.; Teuwen, L.A.; Veys, K.; et al. Single-Cell Transcriptome Atlas of Murine Endothelial Cells. Cell 2020, 180, 764–779.e720. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, N.; He, Y.; D’Addio, M.; Tacconi, C.; Detmar, M.; Dieterich, L.C. Single-cell mapping reveals new markers and functions of lymphatic endothelial cells in lymph nodes. PLoS Biol. 2020, 18, e3000704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wigle, J.T.; Harvey, N.; Detmar, M.; Lagutina, I.; Grosveld, G.; Gunn, M.D.; Jackson, D.G.; Oliver, G. An essential role for Prox1 in the induction of the lymphatic endothelial cell phenotype. EMBO J. 2002, 21, 1505–1513. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kang, J.; Yoo, J.; Ganesan, S.K.; Cook, S.C.; Aguilar, B.; Ramu, S.; Lee, J.; Hong, Y.K. Prox1 physically and functionally interacts with COUP-TFII to specify lymphatic endothelial cell fate. Blood 2009, 113, 1856–1859. [Google Scholar] [CrossRef] [Green Version]
- Jackson, D.G. Hyaluronan in the lymphatics: The key role of the hyaluronan receptor LYVE-1 in leucocyte trafficking. Matrix Biol. J. Int. Soc. Matrix Biol. 2019, 78–79, 219–235. [Google Scholar] [CrossRef]
- Schacht, V.; Ramirez, M.I.; Hong, Y.K.; Hirakawa, S.; Feng, D.; Harvey, N.; Williams, M.; Dvorak, A.M.; Dvorak, H.F.; Oliver, G.; et al. T1alpha/podoplanin deficiency disrupts normal lymphatic vasculature formation and causes lymphedema. EMBO J. 2003, 22, 3546–3556. [Google Scholar] [CrossRef] [PubMed]
- Kriehuber, E.; Breiteneder-Geleff, S.; Groeger, M.; Soleiman, A.; Schoppmann, S.F.; Stingl, G.; Kerjaschki, D.; Maurer, D. Isolation and characterization of dermal lymphatic and blood endothelial cells reveal stable and functionally specialized cell lineages. J. Exp. Med. 2001, 194, 797–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiley, H.E.; Gonzalez, E.B.; Maki, W.; Wu, M.T.; Hwang, S.T. Expression of CC chemokine receptor-7 and regional lymph node metastasis of B16 murine melanoma. J. Natl. Cancer Inst. 2001, 93, 1638–1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebata, N.; Nodasaka, Y.; Sawa, Y.; Yamaoka, Y.; Makino, S.; Totsuka, Y.; Yoshida, S. Desmoplakin as a specific marker of lymphatic vessels. Microvasc. Res. 2001, 61, 40–48. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, J.; Qin, L. VEGF-C mediated enhancement of lymphatic drainage reduces intestinal inflammation by regulating IL-9/IL-17 balance and improving gut microbiota in experimental chronic colitis. Am. J. Transl. Res. 2017, 9, 4772–4784. [Google Scholar] [PubMed]
- Chung, E.S.; Saban, D.R.; Chauhan, S.K.; Dana, R. Regulation of blood vessel versus lymphatic vessel growth in the cornea. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1613–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, Y.; Ji, Y.W.; Lee, S.M.; Shim, J.; Noh, H.; Yeo, A.; Park, C.; Park, M.S.; Chang, E.J.; Lee, H.K. Activation of HIF-1α (hypoxia inducible factor-1α) prevents dry eye-induced acinar cell death in the lacrimal gland. Cell Death Dis. 2014, 5, e1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.S.; Hos, D.; Blanco, T.; Bock, F.; Reyes, N.J.; Mathew, R.; Cursiefen, C.; Dana, R.; Saban, D.R. Involvement of corneal lymphangiogenesis in a mouse model of allergic eye disease. Investig. Ophthalmol. Vis. Sci. 2015, 56, 3140–3148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dale, S.B.; Saban, D.R. Linking immune responses with fibrosis in allergic eye disease. Curr. Opin. Allergy Clin. Immunol. 2015, 15, 467–475. [Google Scholar] [CrossRef] [Green Version]
- Shin, K.; Kataru, R.P.; Park, H.J.; Kwon, B.I.; Kim, T.W.; Hong, Y.K.; Lee, S.H. TH2 cells and their cytokines regulate formation and function of lymphatic vessels. Nat. Commun. 2015, 6, 6196. [Google Scholar] [CrossRef] [Green Version]
- Savetsky, I.L.; Ghanta, S.; Gardenier, J.C.; Torrisi, J.S.; García Nores, G.D.; Hespe, G.E.; Nitti, M.D.; Kataru, R.P.; Mehrara, B.J. Th2 cytokines inhibit lymphangiogenesis. PLoS ONE 2015, 10, e0126908. [Google Scholar] [CrossRef]
- Zhong, W.; Montana, M.; Santosa, S.M.; Isjwara, I.D.; Huang, Y.H.; Han, K.Y.; O’Neil, C.; Wang, A.; Cortina, M.S.; de la Cruz, J.; et al. Angiogenesis and lymphangiogenesis in corneal transplantation—A review. Surv. Ophthalmol. 2018, 63, 453–479. [Google Scholar] [CrossRef]
- Hos, D.; Matthaei, M.; Bock, F.; Maruyama, K.; Notara, M.; Clahsen, T.; Hou, Y.; Le, V.N.H.; Salabarria, A.C.; Horstmann, J.; et al. Immune reactions after modern lamellar (DALK, DSAEK, DMEK) versus conventional penetrating corneal transplantation. Prog. Retin. Eye Res. 2019, 73, 100768. [Google Scholar] [CrossRef]
- Horstmann, J.; Schulz-Hildebrandt, H.; Bock, F.; Siebelmann, S.; Lankenau, E.; Hüttmann, G.; Steven, P.; Cursiefen, C. Label-Free In Vivo Imaging of Corneal Lymphatic Vessels Using Microscopic Optical Coherence Tomography. Investig. Ophthalmol. Vis. Sci. 2017, 58, 5880–5886. [Google Scholar] [CrossRef] [Green Version]
- Le, V.N.H.; Hou, Y.; Horstmann, J.; Bock, F.; Cursiefen, C. Novel Method to Detect Corneal Lymphatic Vessels In Vivo by Intrastromal Injection of Fluorescein. Cornea 2018, 37, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, T.; Bock, F.; Yuen, D.; Hos, D.; Bachmann, B.O.; Zahn, G.; Wiegand, S.; Chen, L.; Cursiefen, C. Cutting edge: Lymphatic vessels, not blood vessels, primarily mediate immune rejections after transplantation. J. Immunol. 2010, 184, 535–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regenfuss, B.; Bock, F.; Cursiefen, C. Corneal angiogenesis and lymphangiogenesis. Curr. Opin. Allergy Clin. Immunol. 2012, 12, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.A.; Chan, S.W.S.; Zhou, X.; Glinka, Y.; Girard, E.; Yucel, Y.; Gupta, N. Lymphatic vessels identified in failed corneal transplants with neovascularisation. Br. J. Ophthalmol. 2019, 103, 421–427. [Google Scholar] [CrossRef] [Green Version]
- Seo, Y.; Kim, M.K.; Lee, J.H.; Chang, E.J.; Kim, E.K.; Lee, H.K. Expression of Lymphangiogenic Markers in Rejected Human Corneal Buttons after Penetrating Keratoplasty. Curr. Eye Res. 2015, 40, 902–912. [Google Scholar] [CrossRef] [PubMed]
- Goyal, S.; Chauhan, S.K.; Dana, R. Blockade of prolymphangiogenic vascular endothelial growth factor C in dry eye disease. Arch. Ophthalmol. 2012, 130, 84–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bock, F.; Maruyama, K.; Regenfuss, B.; Hos, D.; Steven, P.; Heindl, L.M.; Cursiefen, C. Novel anti(lymph)angiogenic treatment strategies for corneal and ocular surface diseases. Prog. Retin. Eye Res. 2013, 34, 89–124. [Google Scholar] [CrossRef]
- Narimatsu, A.; Hattori, T.; Koike, N.; Tajima, K.; Nakagawa, H.; Yamakawa, N.; Usui, Y.; Kumakura, S.; Matsumoto, T.; Goto, H. Corneal lymphangiogenesis ameliorates corneal inflammation and edema in late stage of bacterial keratitis. Sci. Rep. 2019, 9, 2984. [Google Scholar] [CrossRef] [PubMed]
- Goyal, S.; Chauhan, S.K.; El Annan, J.; Nallasamy, N.; Zhang, Q.; Dana, R. Evidence of corneal lymphangiogenesis in dry eye disease: A potential link to adaptive immunity? Arch. Ophthalmol. 2010, 128, 819–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chennakesavalu, M.; Somala, S.R.R.; Dommaraju, S.R.; Peesapati, M.P.; Guo, K.; Rosenblatt, M.I.; Chang, J.H.; Azar, D.T. Corneal lymphangiogenesis as a potential target in dry eye disease—A systematic review. Surv. Ophthalmol. 2021, 66, 960–976. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.W.; Lee, J.L.; Kang, H.G.; Gu, N.; Byun, H.; Yeo, A.; Noh, H.; Kim, S.; Choi, E.Y.; Song, J.S.; et al. Corneal lymphangiogenesis facilitates ocular surface inflammation and cell trafficking in dry eye disease. Ocul. Surf. 2018, 16, 306–313. [Google Scholar] [CrossRef]
- Ji, Y.W.; Lee, J.H.; Choi, E.Y.; Kang, H.G.; Seo, K.Y.; Song, J.S.; Kim, H.C.; Lee, H.K. HIF1α-mediated TRAIL Expression Regulates Lacrimal Gland Inflammation in Dry Eye Disease. Investig. Ophthalmol. Vis. Sci. 2020, 61, 3. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Ji, Y.W.; Hwang, H.S.; Oh, J.W.; Kim, H.C.; Lee, H.K.; Kim, K.P. Proteomic analysis of human lacrimal and tear fluid in dry eye disease. Sci. Rep. 2017, 7, 13363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cursiefen, C.; Maruyama, K.; Bock, F.; Saban, D.; Sadrai, Z.; Lawler, J.; Dana, R.; Masli, S. Thrombospondin 1 inhibits inflammatory lymphangiogenesis by CD36 ligation on monocytes. J. Exp. Med. 2011, 208, 1083–1092. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Kim, J.; Ahn, J.H.; Hong, Y.K.; Mäkinen, T.; Lim, D.S.; Koh, G.Y. YAP and TAZ Negatively Regulate Prox1 During Developmental and Pathologic Lymphangiogenesis. Circ. Res. 2019, 124, 225–242. [Google Scholar] [CrossRef] [PubMed]
- Bryant-Hudson, K.; Conrady, C.D.; Carr, D.J. Type I interferon and lymphangiogenesis in the HSV-1 infected cornea—Are they beneficial to the host? Prog. Retin. Eye Res. 2013, 36, 281–291. [Google Scholar] [CrossRef] [Green Version]
- Wuest, T.R.; Carr, D.J. VEGF-A expression by HSV-1-infected cells drives corneal lymphangiogenesis. J. Exp. Med. 2010, 207, 101–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurung, H.R.; Carr, M.M.; Bryant, K.; Chucair-Elliott, A.J.; Carr, D.J. Fibroblast growth factor-2 drives and maintains progressive corneal neovascularization following HSV-1 infection. Mucosal Immunol. 2018, 11, 172–185. [Google Scholar] [CrossRef] [Green Version]
- Hos, D.; Bukowiecki, A.; Horstmann, J.; Bock, F.; Bucher, F.; Heindl, L.M.; Siebelmann, S.; Steven, P.; Dana, R.; Eming, S.A.; et al. Transient Ingrowth of Lymphatic Vessels into the Physiologically Avascular Cornea Regulates Corneal Edema and Transparency. Sci. Rep. 2017, 7, 7227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryant-Hudson, K.M.; Chucair-Elliott, A.J.; Conrady, C.D.; Cohen, A.; Zheng, M.; Carr, D.J.J. HSV-1 targets lymphatic vessels in the eye and draining lymph node of mice leading to edema in the absence of a functional type I interferon response. Am. J. Pathol. 2013, 183, 1233–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, P.J.; Chang, M.; Garg, N.; Zhu, J.; Chang, J.H.; Shukla, D. Corneal lymphangiogenesis in herpetic stromal keratitis. Surv. Ophthalmol. 2015, 60, 60–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cursiefen, C.; Chen, L.; Borges, L.P.; Jackson, D.; Cao, J.; Radziejewski, C.; D’Amore, P.A.; Dana, M.R.; Wiegand, S.J.; Streilein, J.W. VEGF-A stimulates lymphangiogenesis and hemangiogenesis in inflammatory neovascularization via macrophage recruitment. J. Clin. Investig. 2004, 113, 1040–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clauss, M.; Weich, H.; Breier, G.; Knies, U.; Röckl, W.; Waltenberger, J.; Risau, W. The vascular endothelial growth factor receptor Flt-1 mediates biological activities. Implications for a functional role of placenta growth factor in monocyte activation and chemotaxis. J. Biol. Chem. 1996, 271, 17629–17634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dastjerdi, M.H.; Saban, D.R.; Okanobo, A.; Nallasamy, N.; Sadrai, Z.; Chauhan, S.K.; Hajrasouliha, A.R.; Dana, R. Effects of topical and subconjunctival bevacizumab in high-risk corneal transplant survival. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2411–2417. [Google Scholar] [CrossRef] [Green Version]
- Fasciani, R.; Mosca, L.; Giannico, M.I.; Ambrogio, S.A.; Balestrazzi, E. Subconjunctival and/or intrastromal bevacizumab injections as preconditioning therapy to promote corneal graft survival. Int. Ophthalmol. 2015, 35, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.H.; Shen, E.P. Long-term topical bevacizumab for prevention of corneal graft rejections. Eur. J. Ophthalmol. 2020, 1120672120939504. [Google Scholar] [CrossRef]
- Harooni, H.; Reddy, V.; Root, T.; Ambati, B. Bevacizumab for graft rejection. Ophthalmology 2007, 114, 1950. [Google Scholar] [CrossRef]
- Dohlman, T.H.; Omoto, M.; Hua, J.; Stevenson, W.; Lee, S.M.; Chauhan, S.K.; Dana, R. VEGF-trap aflibercept significantly improves long-term graft survival in high-risk corneal transplantation. Transplantation 2015, 99, 678–686. [Google Scholar] [CrossRef]
- Hayashi, T.; Usui, T.; Yamagami, S. Suppression of Allograft Rejection with Soluble VEGF Receptor 2 Chimeric Protein in a Mouse Model of Corneal Transplantation. Tohoku J. Exp. Med. 2016, 239, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, Y.K.; Zhang, X.; Uehara, H.; Young, J.R.; Archer, B.; Ambati, B. Vascular Endothelial Growth Factor Receptor 1 morpholino increases graft survival in a murine penetrating keratoplasty model. Investig. Ophthalmol. Vis. Sci. 2012, 53, 8458–8471. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Tiem, M.; Watkins, R.; Cho, Y.K.; Wang, Y.; Olsen, T.; Uehara, H.; Mamalis, C.; Luo, L.; Oakey, Z.; et al. Soluble vascular endothelial growth factor receptor 3 is essential for corneal alymphaticity. Blood 2013, 121, 4242–4249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emami-Naeini, P.; Dohlman, T.H.; Omoto, M.; Hattori, T.; Chen, Y.; Lee, H.S.; Chauhan, S.K.; Dana, R. Soluble vascular endothelial growth factor receptor-3 suppresses allosensitization and promotes corneal allograft survival. Graefe’s Arch. Clin. Exp. Ophthalmol. 2014, 252, 1755–1762. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Le, V.N.H.; Tóth, G.; Siebelmann, S.; Horstmann, J.; Gabriel, T.; Bock, F.; Cursiefen, C. UV light crosslinking regresses mature corneal blood and lymphatic vessels and promotes subsequent high-risk corneal transplant survival. Am. J. Transplant. 2018, 18, 2873–2884. [Google Scholar] [CrossRef]
- Bucher, F.; Bi, Y.; Gehlsen, U.; Hos, D.; Cursiefen, C.; Bock, F. Regression of mature lymphatic vessels in the cornea by photodynamic therapy. Br. J. Ophthalmol. 2014, 98, 391–395. [Google Scholar] [CrossRef]
- Hou, Y.; Le, V.N.H.; Clahsen, T.; Schneider, A.C.; Bock, F.; Cursiefen, C. Photodynamic Therapy Leads to Time-Dependent Regression of Pathologic Corneal (Lymph) Angiogenesis and Promotes High-Risk Corneal Allograft Survival. Investig. Ophthalmol. Vis. Sci. 2017, 58, 5862–5869. [Google Scholar] [CrossRef] [Green Version]
- Le, V.N.H.; Schneider, A.C.; Scholz, R.; Bock, F.; Cursiefen, C. Fine Needle-Diathermy Regresses Pathological Corneal (Lymph)Angiogenesis and Promotes High-Risk Corneal Transplant Survival. Sci. Rep. 2018, 8, 5707. [Google Scholar] [CrossRef]
- Hos, D.; Le, V.N.H.; Hellmich, M.; Siebelmann, S.; Roters, S.; Bachmann, B.O.; Cursiefen, C. Risk of Corneal Graft Rejection After High-risk Keratoplasty Following Fine-needle Vessel Coagulation of Corneal Neovascularization Combined With Bevacizumab: A Pilot Study. Transplant. Direct 2019, 5, e452. [Google Scholar] [CrossRef] [PubMed]
- Bignami, F.; Lorusso, A.; Rama, P.; Ferrari, G. Growth inhibition of formed corneal neovascularization following Fosaprepitant treatment. Acta Ophthalmol. 2017, 95, e641–e648. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Authors | Title | Journal | PMID |
|---|---|---|---|
| Schönberg A | Immunomodulatory Strategies Targeting Dendritic Cells to Improve Corneal Graft Survival | J Clin Med. 2020 Apr 28;9(5):1280. doi:10.3390/jcm9051280. | 32354200 |
| Hos D | Immune reactions after modern lamellar (DALK, DSAEK, DMEK) versus conventional penetrating corneal transplantation | Prog Retin Eye Res. 2019 Nov;73:100768. doi:10.1016/j.preteyeres.2019.07.001. Epub 2019 Jul 3. | 31279005 |
| Hori J | Immune privilege in corneal transplantation | Prog Retin Eye Res. 2019 Sep;72:100758. doi:10.1016/j.preteyeres.2019.04.002. Epub 2019 Apr 20. | 31014973 |
| Yu T | The atypical chemokine receptor-2 does not alter corneal graft survival but regulates early stage of corneal graft-induced lymphangiogenesis | Graefes Arch Clin Exp Ophthalmol. 2018 Oct;256(10):1875–1882. doi:10.1007/s00417-018-4070-1. Epub 2018 Jul 27. | 30054731 |
| Le VNH | Fine Needle-Diathermy Regresses Pathological Corneal (Lymph)Angiogenesis and Promotes High-Risk Corneal Transplant Survival | Sci Rep. 2018 Apr 9;8(1):5707. doi:10.1038/s41598-018-24037-3. | 29632336 |
| Su W | Pharmacological inhibition of caspase-8 suppresses inflammation-induced lymphangiogenesis and allograft rejection in the cornea | J Allergy Clin Immunol. 2018 Jul;142(1):290–294.e9. doi:10.1016/j.jaci.2018.02.005. Epub 2018 Mar 2. | 29477723 |
| Zhong W | Angiogenesis and lymphangiogenesis in corneal transplantation-A review | Surv Ophthalmol. 2018 Jul-Aug;63(4):453–479. doi:10.1016/j.survophthal.2017.12.008. Epub 2017 Dec 27. | 29287709 |
| Hou Y | Photodynamic Therapy Leads to Time-Dependent Regression of Pathologic Corneal (Lymph) Angiogenesis and Promotes High-Risk Corneal Allograft Survival | Invest Ophthalmol Vis Sci. 2017 Nov 1;58(13):5862–5869. doi:10.1167/iovs.17-22904. | 29145577 |
| Zhang L | Angiopoietin-2 Blockade Promotes Survival of Corneal Transplants | Invest Ophthalmol Vis Sci. 2017 Jan 1;58(1):79–86. doi:10.1167/iovs.16-20485. | 28061513 |
| Chen WS | Pathological lymphangiogenesis is modulated by galectin-8-dependent crosstalk between podoplanin and integrin-associated VEGFR-3 | Nat Commun. 2016 Apr 12;7:11302. doi:10.1038/ncomms11302. | 27066737 |
| Schöllhorn L | Thrombospondin-1 as a Regulator of Corneal Inflammation and Lymphangiogenesis: Effects on Dry Eye Disease and Corneal Graft Immunology | J Ocul Pharmacol Ther. 2015 Sep;31(7):376–85. doi:10.1089/jop.2015.0020. Epub 2015 Jul 8. | 26154823 |
| Seo Y | Expression of Lymphangiogenic Markers in Rejected Human Corneal Buttons after Penetrating Keratoplasty | Curr Eye Res. 2015 Sep;40(9):902–12. doi:10.3109/02713683.2014.969809. Epub 2014 Oct 20. | 25330436 |
| Emami-Naeini P | Soluble vascular endothelial growth factor receptor-3 suppresses allosensitization and promotes corneal allograft survival | Graefes Arch Clin Exp Ophthalmol. 2014 Nov;252(11):1755–62. doi:10.1007/s00417-014-2749-5. Epub 2014 Aug 5. | 25091513 |
| Hos D | Lymphatic vessels in the development of tissue and organ rejection | Adv Anat Embryol Cell Biol. 2014;214:119–41. doi:10.1007/978-3-7091-1646-3_10. | 24276891 |
| Flynn TH | The effect of perioperative allergic conjunctivitis on corneal lymphangiogenesis after corneal transplantation | BrJ Ophthalmol. 2011 Oct;95(10):1451–6. doi:10.1136/bjo.2010.201939.Epub 2011 Jun 7. | 21653212 |
| Dietrich T | Cutting edge: lymphatic vessels, not blood vessels, primarily mediate immune rejections after transplantation | J Immunol. 2010 Jan 15;184(2):535–9. doi:10.4049/jimmunol.0903180. Epub 2009 Dec 16. | 20018627 |
| Maruyama K | The maintenance of lymphatic vessels in the cornea is dependent on the presence of macrophages | Invest Ophthalmol Vis Sci. 2012 May 31;53(6):3145–53. doi:10.1167/iovs.11-8010. | 22511631 |
| Author | Title | Journal | PMID |
|---|---|---|---|
| Seo, Y | Activation of HIF-1α (hypoxia inducible factor-1α) prevents dry eye-induced acinar cell death in the lacrimal gland | Cell death & disease 2014, 5, e1309. doi:10.1038/cddis.2014.260. | 24967971 |
| Goyal, S | Blockade of prolymphangiogenic vascular endothelial growth factor C in dry eye disease | Arch ophthalmol 2012, 130, 84–89. doi:10.1001/archophthalmol.2011.266. | 21911653 |
| Chennakesavalu, M | Corneal lymphangiogenesis as a potential target in dry eye disease–a systematic review | Sur ophthalmol 20212021 Mar 31;S0039-6257(21)00080-1. doi:10.1016 /j.survophthal.2021.03.007 | 33811911 |
| Goyal, S | Evidence of corneal lymphangiogenesis in dry eye disease: a potential link to adaptive immunity? | Arch ophthalmol 2010, 128, 819–824, doi:10.1001/archophthalmol.2010.124. | 20625040 |
| Lee, SJ | Corneal lymphangiogenesis in dry eye disease is regulated by substance P/neurokinin-1 receptor system through controlling expression of vascular endothelial growth factor receptor 3 | Ocul surf 2021, 22, 72–79. doi:10.1016/j.jtos.2021.07.003. | 34311077 |
| Min JH | Activation of Dll4/Notch Signaling and Hypoxia-Inducible Factor-1 Alpha Facilitates Lymphangiogenesis in Lacrimal Glands in Dry Eye | PLoS One. 2016 Feb 1;11(2):e0147846. doi:10.1371/jounal.pone.0147846. eCollection 2016. | 26828208 |
| Ji YW | Corneal lymphangiogenesis facilitates ocular surface inflammation and cell trafficking in dry eye disease | Ocul Surf. 2018 Jul;16(3):306–313 doi:10.1016/j.jtos.2018.03.008. Epub 2018 Mar 27. | 29601983 |
| Okanobo A | Efficacy of topical blockade of interleukin-1 in experimental dry eye disease | Am J Ophthalmol 2012, 154, 63–71. doi:10.1016/j.ajo.2012.01.034. | 22541929 |
| Authors | Title | Journal | PMID |
|---|---|---|---|
| Narimatsu A. | Corneal lymphangiogenesis ameliorates corneal inflammation and edema in late stage of bacterial keratitis | Sci Rep. 2019 Feb 27;9(1):2984. doi:10.1038/s41598-019-39876-x. | 30814667 |
| Gurung HR | Fibroblast growth factor-2 drives and maintains progressive corneal neovascularization following HSV-1 infection | Mucosal Immunol. 2018 Jan;11(1):172–185. doi:10.1038/mi.2017.26. Epub 2017 Apr 5. | 28378806 |
| Gurung HR | Cornea lymphatics drive the CD8+ T-cell response to herpes simplex virus-1 | Immunol Cell Biol. 2017 Jan;95(1):87–98. doi:10.1038/icb.2016.80. Epub 2016 Aug 31. | 27577867 |
| Lee HS | Involvement of corneal lymphangiogenesis in a mouse model of allergic eye disease | Invest Ophthalmol Vis Sci. 2015 May;56(5):3140–8. doi:10.1167/iovs.14-16186. | 26024097 |
| Park PJ | Corneal lymphangiogenesis in herpetic stromal keratitis | Surv Ophthalmol. 2015 Jan–Feb; 60(1):60–71. doi:10.1016/j.survophthal.2014.06.001. Epub 2014 Jun 10. | 25444520 |
| Wuest TR | VEGF-A expression by HSV-1-infected cells drives corneal lymphangiogenesis | J Exp Med. 2010 Jan 18;207(1):101–15. doi:10.1084/jem.20091385. Epub 2009 Dec 21. | 20026662 |
| Suryawanshi A | IL-17A differentially regulates corneal vascular endothelial growth factor (VEGF)-A and soluble VEGF receptor 1 expression and promotes corneal angiogenesis after herpes simplex virus infection | J Immunol 2012, 188, 3434-3446. doi:10.4049/jimmunol.1102602. | 22379030 |
| Title | Remarks |
|---|---|
| Cross-talk between infiltrating myeloid cells, T cells, and LECs | Determine the role of infiltrating myeloid cells at the early stage of lymphangiogenesis |
| Effectiveness of VEGF inhibitors in allograft survival and lymphangiogenesis | Large scale clinical research for VEGF inhibitors is needed |
| Transcriptome analysis of lymphangiogenesis at the single-cell level | Cell-cell interaction is more clearly defined with scRNA-seq at each step of lymphangiogenesis |
| Proteomic analysis of lymphangiogenesis in each pathologic condition | OMICs study is essential to clearly understand and define druggable targets for lymphangiogenesis |
| Role of lymphangiogenesis in incurable ocular surface disease (e.g., OCP, GVHD) | Studies of lymphangiogenesis on severe ocular vascular diseases are sparse |
| Role of cells residing on the ocular surface and immune cells | Determine the role of corneal epithelium, keratocyte, and endothelial cells in lymphangiogenesis |
| Effective molecular target for developing drugs | Besides VEGFs, more effective and durable targets for drug development should be investigated |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-K.; Lee, S.-M.; Lee, D.-I. Corneal Lymphangiogenesis: Current Pathophysiological Understandings and Its Functional Role in Ocular Surface Disease. Int. J. Mol. Sci. 2021, 22, 11628. https://doi.org/10.3390/ijms222111628
Lee H-K, Lee S-M, Lee D-I. Corneal Lymphangiogenesis: Current Pathophysiological Understandings and Its Functional Role in Ocular Surface Disease. International Journal of Molecular Sciences. 2021; 22(21):11628. https://doi.org/10.3390/ijms222111628
Chicago/Turabian StyleLee, Hyung-Keun, Sang-Mok Lee, and Dong-Ihll Lee. 2021. "Corneal Lymphangiogenesis: Current Pathophysiological Understandings and Its Functional Role in Ocular Surface Disease" International Journal of Molecular Sciences 22, no. 21: 11628. https://doi.org/10.3390/ijms222111628
APA StyleLee, H. -K., Lee, S. -M., & Lee, D. -I. (2021). Corneal Lymphangiogenesis: Current Pathophysiological Understandings and Its Functional Role in Ocular Surface Disease. International Journal of Molecular Sciences, 22(21), 11628. https://doi.org/10.3390/ijms222111628