
Recent Progress in Rice Broad-Spectrum Disease Resistance



Abstract
:1. Introduction
2. Identified Rice Broad-Spectrum Disease Resistance Genes in Past 10 Years
2.1. R Genes Confer Broad-Spectrum Disease Resistance in Rice
2.2. Defense Regulator Genes Contribute to Broad-Spectrum Disease Resistance
2.3. Identification of Broad-Spectrum Disease Resistance Loci by QTL Mapping and GWAS Analysis
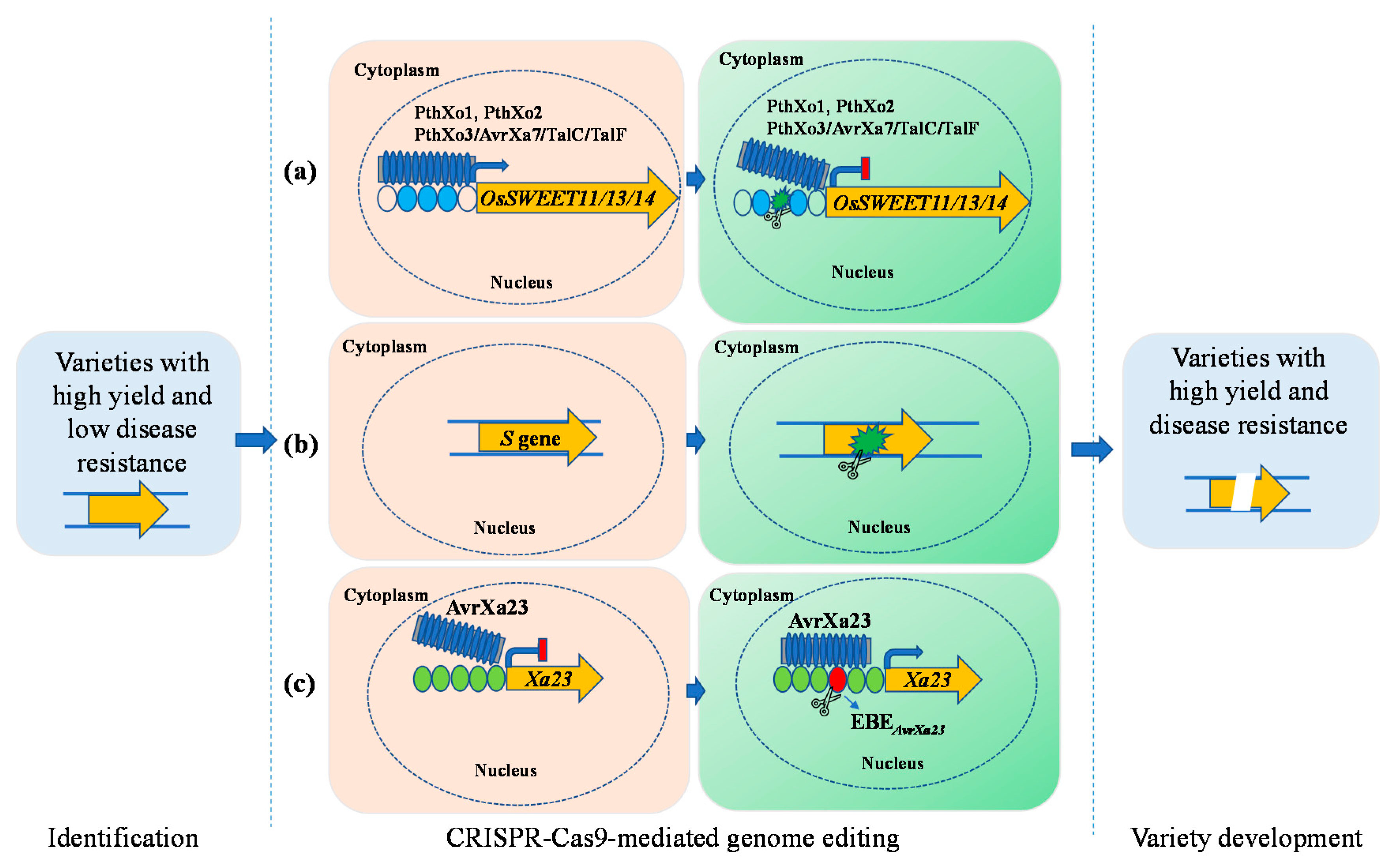
3. Strategies for Broad-Spectrum Disease Resistance Rice Breeding
3.1. Gene Pyramiding Breeding Is an Effective Way to Obtain Broad-Spectrum Disease Resistance Rice Varieties
3.2. Engineering Broad-Spectrum Disease Resistance Rice by Editing Susceptibility and Executor R Genes Is a New Method with Broad Application Prospects
3.3. Transgenic Rice Expressing Genes from Other Species Shows Broad-Spectrum Disease Resistance
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kou, Y.; Wang, S. Broad-spectrum and durability: Understanding of quantitative disease resistance. Curr. Opin. Plant Biol. 2010, 13, 181–185. [Google Scholar] [CrossRef]
- Li, W.; Chern, M.; Yin, J.; Wang, J.; Chen, X. Recent advances in broad-spectrum resistance to the rice blast disease. Curr. Opin. Plant Biol. 2019, 50, 114–120. [Google Scholar] [CrossRef]
- Peng, Y.; Yang, J.; Li, X.; Zhang, Y. Salicylic acid: Biosynthesis and signaling. Annu. Rev. Plant Biol. 2021, 72, 761–791. [Google Scholar] [CrossRef]
- Das, A.; Soubam, D.; Singh, P.K.; Thakur, S.; Singh, N.K.; Sharma, T.R. A novel blast resistance gene, Pi54rh cloned from wild species of rice, Oryza rhizomatis confers broad spectrum resistance to Magnaporthe oryzae. Funct. Integr. Genom. 2012, 12, 215–228. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, B.; Zhu, X.; Yang, J.; Bordeos, A.; Wang, G.; Leach, J.E.; Leung, H. Fine-mapping and molecular marker development for Pi56(t), a NBS-LRR gene conferring broad-spectrum resistance to Magnaporthe oryzae in rice. Theor. Appl. Genet. 2013, 126, 985–998. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Lei, C.; Xu, X.; Hao, K.; Wang, J.; Cheng, Z.; Ma, X.; Ma, J.; Zhou, K.; Zhang, X.; et al. Pi64, encoding a novel CC-NBS-LRR protein, confers resistance to leaf and neck blast in rice. Mol. Plant-Microbe Interact. 2015, 28, 558–568. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Zhai, K.; Xie, Z.; Yang, D.; Zhu, X.; Liu, J.; Wang, X.; Qin, P.; Yang, Y.; Zhang, G. Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance. Science 2017, 355, 962–965. [Google Scholar] [CrossRef]
- Xie, Z.; Yan, B.; Shou, J.; Tang, J.; Wang, X.; Zhai, K.; Liu, J.; Li, Q.; Luo, M.; Deng, Y.; et al. A nucleotide-binding site-leucine-rich repeat receptor pair confers broad-spectrum disease resistance through physical association in rice. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180308. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Wang, X.; Jia, Y.; Minkenberg, B.; Wheatley, M.; Fan, J.; Jia, M.H.; Famoso, A.; Edwards, J.D.; Wamishe, Y.; et al. The rice blast resistance gene Ptr encodes an atypical protein required for broad-spectrum disease resistance. Nat. Commun. 2018, 9, 2039. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Xiao, G.; Telebanco-Yanoria, M.J.; Siazon, P.M.; Padilla, J.; Opulencia, R.; Bigirimana, J.; Habarugira, G.; Wu, J.; Li, M.; et al. The broad-spectrum rice blast resistance (R) gene Pita2 encodes a novel R protein unique from Pita. Rice 2020, 13, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasudevan, K.; Vera Cruz, C.M.; Gruissem, W.; Bhullar, N.K. Geographically distinct and domain-specific sequence variations in the alleles of rice blast resistance gene Pib. Front. Plant Sci. 2016, 7, 915. [Google Scholar] [CrossRef] [Green Version]
- Roychowdhury, M.; Jia, Y.; Jia, M.H.; Fjellstrom, R.; Cartwright, R.D. Identification of the rice blast resistance gene Pib in the national small grains collection. Phytopathology 2012, 102, 700–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, L.; Wu, J.; Chen, C.; Wu, W.; He, X.; Lin, F.; Wang, L.; Ashikawa, I.; Matsumoto, T.; Wang, L.; et al. The isolation of Pi1, an allele at the Pik locus which confers broad spectrum resistance to rice blast. Theor. Appl. Genet. 2012, 125, 1047–1055. [Google Scholar] [CrossRef]
- Thakur, S.; Singh, P.K.; Das, A.; Rathour, R.; Variar, M.; Prashanthi, S.K.; Singh, A.K.; Singh, U.D.; Chand, D.; Singh, N.K.; et al. Extensive sequence variation in rice blast resistance gene Pi54 makes it broad spectrum in nature. Front. Plant Sci. 2015, 6, 345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, F.; He, Y.; Chen, J.; Long, X.; Wang, H.; Zhu, M.; Liu, S.; Cai, Q.; Zhang, Z. Analysis of natural variation of the rice blast resistance gene Pike and identification of a novel allele Pikg. Mol. Genet. Genom. 2021, 296, 939–952. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.; Chi, W.; Huang, L.; Qu, M.; Zhang, S.; Chen, Z.Q.; Chen, Z.J.; Tian, D.; Gui, Y.; Chen, X.; et al. Bulked segregant analysis coupled with whole-genome sequencing (BSA-seq) mapping identifies a novel pi21 haplotype conferring basal resistance to rice blast disease. Int. J. Mol. Sci. 2020, 21, 2162. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Lin, K.; Zhang, W.; Nan, J.; Zhang, X.; Wang, C.; Wang, R.; Jiang, G.; Yuan, Q.; Lin, S. Improving the blast resistance of the elite rice variety Kongyu-131 by updating the pi21 locus. BMC Plant Biol. 2019, 19, 1–12. [Google Scholar] [CrossRef]
- Zhou, Y.; Lei, F.; Wang, Q.; He, W.; Yuan, B.; Yuan, W. Identification of novel alleles of the rice blast-resistance gene Pi9 through sequence-based allele mining. Rice 2020, 13, 1–12. [Google Scholar] [CrossRef]
- Xiao, G.; Borja, F.N.; Mauleon, R.; Padilla, J.; Telebanco-Yanoria, M.J.; Yang, J.; Lu, G.; Dionisio-Sese, M.; Zhou, B. Identification of resistant germplasm containing novel resistance genes at or tightly linked to the Pi2/9 locus conferring broad-spectrum resistance against rice blast. Rice 2017, 10, 1–12. [Google Scholar] [CrossRef]
- Tian, D.; Chen, Z.; Chen, Z.; Zhou, Y.; Wang, Z.; Wang, F.; Chen, S. Allele-specific marker-based assessment revealed that the rice blast resistance genes Pi2 and Pi9 have not been widely deployed in Chinese indica rice cultivars. Rice 2016, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Imam, J.; Mandal, N.P.; Variar, M.; Shukla, P. Allele mining and selective patterns of Pi9 gene in a set of rice landraces from india. Front. Plant Sci. 2016, 7, 1846. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Wang, W.; Han, J.; Chen, S.; Wang, C.; Zeng, L.; Feng, A.; Yang, J.; Zhou, B.; Zhu, X. Functional divergence of duplicated genes results in a novel blast resistance gene Pi50 at the Pi2/9 locus. Theor. Appl. Genet. 2015, 128, 2213–2225. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Bao, Y.M.; Xie, L.J.; Su, Y.Y.; Chu, R.Z.; He, W.W.; Huang, J.; Wang, J.F.; Zhang, H.S. Fine mapping and identification of blast resistance gene Pi-hk1 in a broad-spectrum resistant japonica rice landrace. Phytopathology 2013, 103, 1162–1168. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Liu, S.; Xu, P.; Deng, W.; Li, X.; Tharreau, D.; Li, J.; Zhou, J.; Wang, Q.; Tao, D.; et al. Fine mapping of Pi57(t) conferring broad spectrum resistance against Magnaporthe oryzae in introgression line IL-E1454 derived from Oryza longistaminata. PLoS ONE 2017, 12, e0186201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, W.; Wang, Y.; Wang, L.; Ma, Z.; Zhao, J.; Wang, P.; Zhang, L.; Liu, Z.; Lu, X. Genetic mapping and molecular marker development for Pi65(t), a novel broad-spectrum resistance gene to rice blast using next-generation sequencing. Theor. Appl. Genet. 2016, 129, 1035–1044. [Google Scholar] [CrossRef]
- Dong, L.; Liu, S.; Kyaing, M.S.; Xu, P.; Tharreau, D.; Deng, W.; Li, X.; Bi, Y.; Zeng, L.; Li, J.; et al. Identification and fine mapping of Pi69(t), a new gene conferring broad-spectrum resistance against Magnaporthe oryzae from Oryza glaberrima steud. Front. Plant Sci. 2020, 11, 1190. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, P.; Mei, L.; He, X.; Chen, L.; Liu, H.; Shen, S.; Ji, Z.; Zheng, X.; Zhang, Y.; et al. Xa7, a new executor R gene that confers durable and broad-spectrum resistance to bacterial blight disease in rice. Plant Commun. 2021, 2, 100143. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, X.; Fan, Y.; Gao, Y.; Zhu, Q.; Zheng, C.; Qin, T.; Li, Y.; Che, J.; Zhang, M.; et al. XA23 is an executor R protein and confers broad-spectrum disease resistance in rice. Mol. Plant 2015, 8, 290–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutin, M.; Sabot, F.; Ghesquiere, A.; Koebnik, R.; Szurek, B. A knowledge-based molecular screen uncovers a broad-spectrum OsSWEET14 resistance allele to bacterial blight from wild rice. Plant J. 2015, 84, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Zhang, D.; Yin, F.; Zhong, Q.; Wang, B.; Xiao, S.; Ke, X.; Wang, L.; Zhang, Y.; Zhao, C.; et al. Identification and fine-mapping of a new bacterial blight resistance gene, Xa47(t), in G252, an introgression line of yuanjiang common wild rice (Oryza rufipogon). Plant Dis. 2021. [Google Scholar] [CrossRef]
- Du, D.; Zhang, C.; Xing, Y.; Lu, X.; Cai, L.; Yun, H.; Zhang, Q.; Zhang, Y.; Chen, X.; Liu, M.; et al. The CC-NB-LRR OsRLR1 mediates rice disease resistance through interaction with OsWRKY19. Plant Biotechnol. J. 2021, 19, 1052–1064. [Google Scholar] [CrossRef]
- Li, D.; Li, S.; Wei, S.; Sun, W. Strategies to manage rice sheath blight: Lessons from interactions between rice and Rhizoctonia solani. Rice 2021, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Fan, J.; Fang, A.; Li, Y.; Tariqjaveed, M.; Li, D.; Hu, D.; Wang, W.M. Ustilaginoidea virens: Insights into an emerging rice pathogen. Annu. Rev. Phytopathol. 2020, 58, 363–385. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Y.; He, J.; Zheng, X.; Hu, J.; Liu, Y.; Dai, H.; Zhang, Y.; Wang, B.; Wu, W.; et al. STV11 encodes a sulphotransferase and confers durable resistance to rice stripe virus. Nat. Commun. 2014, 5, 4768. [Google Scholar] [CrossRef]
- Wu, J.; Kou, Y.; Bao, J.; Li, Y.; Tang, M.; Zhu, X.; Ponaya, A.; Xiao, G.; Li, J.; Li, C.; et al. Comparative genomics identifies the Magnaporthe oryzae avirulence effector AvrPi9 that triggers Pi9-mediated blast resistance in rice. New Phytol. 2015, 206, 1463–1475. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Yin, J.; Chern, M.; Zhu, X.; Yang, C.; He, K.; Liu, Y.; He, M.; Wang, J.; Song, L.; et al. New insights into bsr-d1-mediated broad-spectrum resistance to rice blast. Mol. Plant Pathol. 2020, 21, 951–960. [Google Scholar] [CrossRef]
- Li, W.; Zhu, Z.; Chern, M.; Yin, J.; Yang, C.; Ran, L.; Cheng, M.; He, M.; Wang, K.; Wang, J.; et al. A natural allele of a transcription factor in rice confers broad-spectrum blast resistance. Cell 2017, 170, 114–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.T.; Wang, K.; Chern, M.; Liu, Y.C.; Zhu, Z.W.; Liu, J.; Zhu, X.B.; Yin, J.J.; Ran, L.; Xiong, J.; et al. Sclerenchyma cell thickening through enhanced lignification induced by OsMYB30 prevents fungal penetration of rice leaves. New Phytol. 2020, 226, 1850–1863. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xia, Y.; Lin, S.; Wang, Y.; Guo, B.; Song, X.; Ding, S.; Zheng, L.; Feng, R.; Chen, S.; et al. Osa-miR164a targets OsNAC60 and negatively regulates rice immunity against the blast fungus Magnaporthe oryzae. Plant J. 2018, 95, 584–597. [Google Scholar] [CrossRef] [Green Version]
- Inoue, H.; Hayashi, N.; Matsushita, A.; Xinqiong, L.; Nakayama, A.; Sugano, S.; Jiang, C.J.; Takatsuji, H. Blast resistance of CC-NB-LRR protein Pb1 is mediated by WRKY45 through protein-protein interaction. Proc. Natl. Acad. Sci. USA 2013, 110, 9577–9582. [Google Scholar] [CrossRef] [Green Version]
- Zhai, K.R.; Deng, Y.W.; Liang, D.; Tang, J.; Liu, J.; Yan, B.X.; Yin, X.; Lin, H.; Chen, F.D.; Yang, D.Y.; et al. RRM transcription factors Interact with NLRs and regulate broad-spectrum blast resistance in rice. Mol. Cell 2019, 74, 996–1009.e7. [Google Scholar] [CrossRef]
- Li, W.; Zhong, S.; Li, G.; Li, Q.; Mao, B.; Deng, Y.; Zhang, H.; Zeng, L.; Song, F.; He, Z. Rice RING protein OsBBI1 with E3 ligase activity confers broad-spectrum resistance against Magnaporthe oryzae by modifying the cell wall defence. Cell Res. 2011, 21, 835–848. [Google Scholar] [CrossRef]
- Liu, M.X.; Zhang, S.B.; Hu, J.X.; Sun, W.X.; Padilla, J.; He, Y.L.; Li, Y.; Yin, Z.Y.; Liu, X.Y.; Wang, W.H.; et al. Phosphorylation-guarded light-harvesting complex II contributes to broad-spectrum blast resistance in rice. Proc. Natl. Acad. Sci. USA 2019, 116, 17572–17577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Chen, X.; Ding, X.; Wang, Y.; Chen, Q.; Song, W.Y. The XA21 binding protein XB25 is required for maintaining XA21-mediated disease resistance. Plant J. 2013, 73, 814–823. [Google Scholar] [CrossRef]
- Park, C.-J.; Song, M.-Y.; Kim, C.-Y.; Jeon, J.-S.; Ronald, P.C. Rice bip3 regulates immunity mediated by the PRRs XA3 and XA21 but not immunity mediated by the NB-LRR protein, Pi5. Biochem. Biophys. Res. Commun. 2014, 448, 70–75. [Google Scholar] [CrossRef]
- Feng, J.X.; Cao, L.; Li, J.; Duan, C.J.; Luo, X.M.; Le, N.; Wei, H.H.; Liang, S.J.; Chu, C.C.; Pan, Q.H.; et al. Involvement of OsNPR1/NH1 in rice basal resistance to blast fungus Magnaporthe oryzae. Eur. J. Plant Pathol. 2011, 131, 221–235. [Google Scholar] [CrossRef]
- Chern, M.; Xu, Q.; Bart, R.S.; Bai, W.; Ruan, D.; Sze-To, W.H.; Canlas, P.E.; Jain, R.; Chen, X.; Ronald, P.C. A genetic screen identifies a requirement for cysteine-rich-receptor-like kinases in rice NH1 (OsNPR1)-mediated immunity. PLoS Genet. 2016, 12, e1006049. [Google Scholar] [CrossRef]
- He, S.L.; Jiang, J.Z.; Chen, B.H.; Kuo, C.H.; Ho, S.L. Overexpression of a constitutively active truncated form of OsCDPK1 confers disease resistance by affecting OsPR10a expression in rice. Sci. Rep. 2018, 8, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Wang, L.; Yang, Z.; Liu, H.; Chu, C.; Zhang, Z.; Zhang, Q.; Li, X.; Xiao, J.; Wang, S.; et al. The rice Raf-like MAPKKK OsILA1 confers broad-spectrum resistance to bacterial blight by suppressing the OsMAPKK4-OsMAPK6 cascade. J. Integr. Plant Biol. 2021, 63, 1815–1842. [Google Scholar] [CrossRef]
- Chen, H.; Li, C.; Liu, L.; Zhao, J.; Cheng, X.; Jiang, G.; Zhai, W. The Fd-GOGAT1 mutant gene lc7 confers resistance to Xanthomonas oryzae pv. oryzae in rice. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Li, J.F.; Ao, Y.; Qu, J.W.; Li, Z.Q.; Su, J.B.; Zhang, Y.; Liu, J.; Feng, D.R.; Qi, K.B.; et al. Lysin motif-containing proteins LYP4 and LYP6 play dual roles in peptidoglycan and chitin perception in rice innate immunity. Plant Cell 2012, 24, 3406–3419. [Google Scholar] [CrossRef] [Green Version]
- Vo, K.T.X.; Kim, C.Y.; Hoang, T.V.; Lee, S.K.; Shirsekar, G.; Seo, Y.S.; Lee, S.W.; Wang, G.L.; Jeon, J.S. OsWRKY67 plays a positive role in basal and XA21-mediated resistance in rice. Front. Plant Sci. 2017, 8, 2220. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.M.; Shi, Z.Y.; Zhang, X.H.; Wang, M.X.; Zhang, L.; Zheng, K.Z.; Liu, J.Y.; Hu, X.M.; Di, C.R.; Qian, Q.; et al. Inducible overexpression of Ideal Plant Architecture1 improves both yield and disease resistance in rice. Nat. Plants 2019, 5, 902. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, L.; Shi, H.; Chern, M.; Yu, H.; Yi, H.; He, M.; Yin, J.J.; Zhu, X.B.; Li, Y.; et al. A single transcription factor promotes both yield and immunity in rice. Science 2018, 361, 1026–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, S.; Liu, H.; Zhang, M.; Chen, D.; Li, Q.; Tian, J.; Xiao, J.; Li, X.; Wang, S.; Yuan, M. The host basal transcription factor IIA subunits coordinate for facilitating infection of TALEs-carrying bacterial pathogens in rice. Plant Sci. 2019, 284, 48–56. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, J.; Yan, S.; Zhang, S.; Zhao, J.; Wang, W.; Yang, T.; Wang, X.; Mao, X.; Dong, J.; et al. The germin-like protein OsGLP2-1 enhances resistance to fungal blast and bacterial blight in rice. Plant Mol. Biol. 2016, 92, 411–423. [Google Scholar] [CrossRef]
- Filipe, O.; De Vleesschauwer, D.; Haeck, A.; Demeestere, K.; Hofte, M. The energy sensor OsSnRK1a confers broad-spectrum disease resistance in rice. Sci. Rep. 2018, 8, 3864. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.-Y.; Vo, K.T.X.; An, G.; Jeon, J.-S. A rice sucrose non-fermenting-1 related protein kinase 1, OSK35, plays an important role in fungal and bacterial disease resistance. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 669–675. [Google Scholar] [CrossRef]
- Wang, J.; Wang, S.; Hu, K.; Yang, J.; Xin, X.; Zhou, W.; Fan, J.; Cui, F.; Mou, B.; Zhang, S.; et al. The calcium-dependent protein kinase OsCPK4 regulates a buffering mechanism that fine-tunes innate immunity in rice. Phytopathology 2018, 108, 35. [Google Scholar]
- Kanda, Y.; Nishizawa, Y.; Kamakura, T.; Mori, M. Overexpressed BSR1-mediated enhancement of disease resistance depends on the MAMP-recognition system. Int. J. Mol. Sci. 2020, 21, 5397. [Google Scholar] [CrossRef]
- You, Q.; Zhai, K.; Yang, D.; Yang, W.; Wu, J.; Liu, J.; Pan, W.; Wang, J.; Zhu, X.; Jian, Y.; et al. An E3 ubiquitin ligase-bag protein module controls plant innate immunity and broad-spectrum disease resistance. Cell Host Microbe 2016, 20, 758–769. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Park, C.H.; He, F.; Nagano, M.; Wang, M.; Bellizzi, M.; Zhang, K.; Zeng, X.; Liu, W.; Ning, Y.; et al. The RhoGAP SPIN6 associates with SPL11 and OsRac1 and negatively regulates programmed cell death and innate immunity in rice. PLoS Pathog. 2015, 11, e1004629. [Google Scholar] [CrossRef]
- Harkenrider, M.; Sharma, R.; De Vleesschauwer, D.; Tsao, L.; Zhang, X.T.; Chern, M.; Canlas, P.; Zuo, S.M.; Ronald, P.C. Overexpression of rice wall-associated kinase 25 (OsWAK25) alters resistance to bacterial and fungal pathogens. PLoS ONE 2016, 11, e0147310. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.E.; Ning, Y.S.; Zhang, Y.X.; Yu, N.; Zhao, C.D.; Zhan, X.D.; Wu, W.X.; Chen, D.B.; Wei, X.J.; Wang, G.L.; et al. OsCUL3a negatively regulates cell death and immunity by degrading OsNPR1 in rice. Plant Cell 2017, 29, 345–359. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Ding, B.; Zhou, X.; Wang, G.L. The rice dynamin-related protein OsDRP1E negatively regulates programmed cell death by controlling the release of cytochrome c from mitochondria. PLoS Pathog. 2017, 13, e1006157. [Google Scholar] [CrossRef]
- Wang, S.; Lei, C.; Wang, J.; Ma, J.; Tang, S.; Wang, C.; Zhao, K.; Tian, P.; Zhang, H.; Qi, C.; et al. SPL33, encoding an eEF1A-like protein, negatively regulates cell death and defense responses in rice. J. Exp. Bot. 2017, 68, 899–913. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, P.; Li, C.; Wang, Y.; Guo, L.; Jiang, G.; Zhai, W. LMM5.1 and LMM5.4, two eukaryotic translation elongation factor 1A-like gene family members, negatively affect cell death and disease resistance in rice. J. Genet. Genom. 2017, 44, 107–118. [Google Scholar] [CrossRef]
- Qin, P.; Fan, S.; Deng, L.; Zhong, G.; Zhang, S.; Li, M.; Chen, W.; Wang, G.; Tu, B.; Wang, Y.; et al. LML1, encoding a conserved eukaryotic release factor 1 protein, regulates cell death and pathogen resistance by forming a conserved complex with SPL33 in rice. Plant Cell Physiol. 2018, 59, 887–902. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Bai, Q.; Xu, P.; Wu, T.; Guo, D.; Peng, Y.; Zhang, H.; Deng, X.; Chen, X.; Luo, M.; et al. Mutation in rice abscisic acid2 results in cell death, enhanced disease-resistance, altered seed dormancy and development. Front. Plant Sci. 2018, 9, 405. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Wang, Y.; Ma, X.; Meng, L.; Jing, R.; Wang, F.; Wang, S.; Cheng, Z.; Zhang, X.; Jiang, L.; et al. Disruption of gene SPL35, encoding a novel CUE domain-containing protein, leads to cell death and enhanced disease response in rice. Plant Biotechnol. J. 2019, 17, 1679–1693. [Google Scholar] [CrossRef] [Green Version]
- Ding, B.; Bellizzi Mdel, R.; Ning, Y.; Meyers, B.C.; Wang, G.L. HDT701, a histone H4 deacetylase, negatively regulates plant innate immunity by modulating histone H4 acetylation of defense-related genes in rice. Plant Cell 2012, 24, 3783–3794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Y.; Liu, Q.; Cao, Y.; Zhang, Y.; Chen, D.; Lou, X.; Cheng, S.; Cao, L. The Osmpk15 negatively regulates Magnaporthe oryza and Xoo disease resistance via SA and JA signaling pathway in rice. Front. Plant Sci. 2019, 10, 752. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Liao, H.; Chern, M.; Yin, J.; Chen, Y.; Wang, J.; Zhu, X.; Chen, Z.; Yuan, C.; Zhao, W.; et al. Loss of function of a rice TPR-domain RNA-binding protein confers broad-spectrum disease resistance. Proc. Natl. Acad. Sci. USA 2018, 115, 3174–3179. [Google Scholar] [CrossRef] [Green Version]
- Ke, Y.; Yuan, M.; Liu, H.; Hui, S.; Qin, X.; Chen, J.; Zhang, Q.; Li, X.; Xiao, J.; Zhang, Q.; et al. The versatile functions of OsALDH2B1 provide a genic basis for growth-defense trade-offs in rice. Proc. Natl. Acad. Sci. USA 2020, 117, 3867–3873. [Google Scholar] [CrossRef] [PubMed]
- Tonnessen, B.W.; Manosalva, P.; Lang, J.M.; Baraoidan, M.; Bordeos, A.; Mauleon, R.; Oard, J.; Hulbert, S.; Leung, H.; Leach, J.E. Rice phenylalanine ammonia-lyase gene OsPAL4 is associated with broad spectrum disease resistance. Plant Mol. Biol. 2015, 87, 273–286. [Google Scholar] [CrossRef]
- Yang, W.; Ju, Y.; Zuo, L.; Shang, L.; Li, X.; Li, X.; Feng, S.; Ding, X.; Chu, Z. OsHsfB4d binds the promoter and regulates the expression of oshsp18.0-ci to resistant against Xanthomonas oryzae. Rice 2020, 13, 28. [Google Scholar] [CrossRef]
- Ju, Y.; Tian, H.; Zhang, R.; Zuo, L.; Jin, G.; Xu, Q.; Ding, X.; Li, X.; Chu, Z. Overexpression of OsHSP18.0-CI enhances resistance to bacterial leaf streak in rice. Rice 2017, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Ke, Y.; Liu, H.; Li, X.; Xiao, J.; Wang, S. Rice OsPAD4 functions differently from Arabidopsis AtPAD4 in host-pathogen interactions. Plant J. 2014, 78, 619–631. [Google Scholar] [CrossRef]
- Son, S.; Kim, H.; Lee, K.S.; Kim, S.; Park, S.R. Rice glutaredoxin GRXS15 confers broad-spectrum resistance to Xanthomonas oryzae pv. oryzae and Fusarium fujikuroi. Biochem. Biophys. Res. Commun. 2020, 533, 1385–1392. [Google Scholar] [CrossRef]
- Son, S.; Moon, S.J.; Kim, H.; Lee, K.S.; Park, S.R. Identification of a novel NPR1 homolog gene, OsNH5N16, which contributes to broad-spectrum resistance in rice. Biochem. Biophys. Res. Commun. 2021, 549, 200–206. [Google Scholar] [CrossRef]
- Li, N.; Wei, S.; Chen, J.; Yang, F.; Kong, L.; Chen, C.; Ding, X.; Chu, Z. OsASR2 regulates the expression of a defence-related gene, Os2H16, by targeting the GT-1 cis-element. Plant Biotechnol. J. 2018, 16, 771–783. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Kong, L.; Zhou, W.; Zhang, X.; Wei, S.; Ding, X.; Chu, Z. Overexpression of Os2H16 enhances resistance to phytopathogens and tolerance to drought stress in rice. Plant Cell Tissue Organ Cult. 2013, 115, 429–441. [Google Scholar] [CrossRef]
- Manosalva, P.M.; Bruce, M.; Leach, J.E. Rice 14-3-3 protein (GF14e) negatively affects cell death and disease resistance. Plant J. 2011, 68, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Hu, Y.; Tang, X.; Zhou, P.; Deng, X.; Wang, H.; Guo, Z. Constitutive expression of rice WRKY30 gene increases the endogenous jasmonic acid accumulation, PR gene expression and resistance to fungal pathogens in rice. Planta 2012, 236, 1485–1498. [Google Scholar] [CrossRef]
- Helliwell, E.E.; Wang, Q.; Yang, Y. Transgenic rice with inducible ethylene production exhibits broad-spectrum disease resistance to the fungal pathogens Magnaporthe oryzae and Rhizoctonia solani. Plant Biotechnol. J. 2013, 11, 33–42. [Google Scholar] [CrossRef]
- Hu, B.; Zhou, Y.; Zhou, Z.; Sun, B.; Zhou, F.; Yin, C.; Ma, W.; Chen, H.; Lin, Y. Repressed OsMESL expression triggers reactive oxygen species-mediated broad-spectrum disease resistance in rice. Plant Biotechnol. J. 2021, 19, 1511–1522. [Google Scholar] [CrossRef]
- Yin, X.; Zou, B.; Hong, X.; Gao, M.; Yang, W.; Zhong, X.; He, Y.; Kuai, P.; Lou, Y.; Huang, J.; et al. Rice copine genes OsBON1 and OsBON3 function as suppressors of broad-spectrum disease resistance. Plant Biotechnol. J. 2018, 16, 1476–1487. [Google Scholar] [CrossRef]
- Bossa-Castro, A.M.; Tekete, C.; Raghavan, C.; Delorean, E.E.; Dereeper, A.; Dagno, K.; Koita, O.; Mosquera, G.; Leung, H.; Verdier, V.; et al. Allelic variation for broad-spectrum resistance and susceptibility to bacterial pathogens identified in a rice MAGIC population. Plant Biotechnol. J. 2018, 16, 1559–1568. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Wang, J.; Bao, Y.; Li, P.; Xie, L.; Huang, J.; Zhang, H. Identification of the quantitative trait loci in japonica rice landrace Heikezijing responsible for broad-spectrum resistance to rice blast. Phytopathology 2010, 100, 822–829. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Qi, X.; Gealy, D.R.; Olsen, K.M.; Caicedo, A.L.; Jia, Y. QTL analysis for resistance to blast disease in U.S. weedy rice. Mol. Plant-Microbe Interact. 2015, 28, 834–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sattayachiti, W.; Wanchana, S.; Arikit, S.; Nubankoh, P.; Patarapuwadol, S.; Vanavichit, A.; Darwell, C.T.; Toojinda, T. Genome-wide association analysis identifies resistance loci for bacterial leaf streak resistance in rice (Oryza sativa L.). Plants 2020, 9, 1673. [Google Scholar] [CrossRef]
- Korinsak, S.; Darwell, C.T.; Wanchana, S.; Praphaisal, L.; Korinsak, S.; Thunnom, B.; Patarapuwadol, S.; Toojinda, T. Identification of bacterial blight resistance loci in rice (Oryza sativa L.) against diverse Xoo thai strains by genome-wide association study. Plants 2021, 10, 518. [Google Scholar] [CrossRef] [PubMed]
- Kumar, I.S.; Nadarajah, K. A meta-analysis of quantitative trait loci associated with multiple disease resistance in rice (Oryza sativa L.). Plants 2020, 9, 1491. [Google Scholar] [CrossRef] [PubMed]
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.B.; Rahim, H.A.; Latif, M.A. Marker-assisted introgression of broad-spectrum blast resistance genes into the cultivated MR219 rice variety. J. Sci. Food Agric. 2017, 97, 2810–2818. [Google Scholar] [CrossRef]
- Jiang, H.; Li, Z.; Liu, J.; Shen, Z.; Gao, G.; Zhang, Q.; He, Y. Development and evaluation of improved lines with broad-spectrum resistance to rice blast using nine resistance genes. Rice 2019, 12, 29. [Google Scholar] [CrossRef] [Green Version]
- Jain, P.; Dubey, H.; Singh, P.K.; Solanke, A.U.; Singh, A.K.; Sharma, T.R. Deciphering signalling network in broad spectrum near isogenic lines of rice resistant to Magnaporthe oryzae. Sci. Rep. 2019, 9, 16939. [Google Scholar] [CrossRef]
- Wu, Y.; Xiao, N.; Chen, Y.; Yu, L.; Pan, C.; Li, Y.; Zhang, X.; Huang, N.; Ji, H.; Dai, Z.; et al. Comprehensive evaluation of resistance effects of pyramiding lines with different broad-spectrum resistance genes against Magnaporthe oryzae in rice (Oryza sativa L.). Rice 2019, 12, 1–13. [Google Scholar] [CrossRef]
- Yang, D.; Tang, J.; Yang, D.; Chen, Y.; Ali, J.; Mou, T. Improving rice blast resistance of Feng39S through molecular marker-assisted backcrossing. Rice 2019, 12, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhao, L.; Zhang, X.; Zhang, Q.; Jia, Y.; Wang, G.; Li, S.; Tian, D.; Li, W.H.; Yang, S. Large-scale identification and functional analysis of NLR genes in blast resistance in the Tetep rice genome sequence. Proc. Natl. Acad. Sci. USA 2019, 116, 18479–18487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalingam, J.; Palanisamy, S.; Alagarasan, G.; Renganathan, V.G.; Ramanathan, A.; Saraswathi, R. Improvement of stable restorer lines for blast resistance through functional marker in rice (Oryza sativa L.). Genes 2020, 11, 1266. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Lin, X.; Xiang, X.; Ren, H.; Fan, X.; Chen, K. Characterization and evaluation of transgenic rice pyramided with the Pi Genes Pib, Pi25 and Pi54. Rice 2021, 14, 1–14. [Google Scholar] [CrossRef]
- Suh, J.P.; Jeung, J.U.; Noh, T.H.; Cho, Y.C.; Park, S.H.; Park, H.S.; Shin, M.S.; Kim, C.K.; Jena, K.K. Development of breeding lines with three pyramided resistance genes that confer broad-spectrum bacterial blight resistance and their molecular analysis in rice. Rice 2013, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, S.K.; Nayak, D.K.; Mohanty, S.; Behera, L.; Barik, S.R.; Pandit, E.; Lenka, S.; Anandan, A. Pyramiding of three bacterial blight resistance genes for broad-spectrum resistance in deepwater rice variety, Jalmagna. Rice 2015, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, J.; Savitha, P.; Alagarasan, G.; Saraswathi, R.; Chandrababu, R. Functional marker assisted improvement of stable cytoplasmic male sterile lines of rice for bacterial blight resistance. Front. Plant Sci. 2017, 8, 1131. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Fang, Z.; Zhou, J.; Li, L.; Lu, L.; Li, L.; Li, T.; Chen, L.; Zhang, W.; Zhai, W.; et al. Transcriptional insights into the pyramided resistance to rice bacterial blight. Sci. Rep. 2018, 8, 12358. [Google Scholar] [CrossRef] [PubMed]
- Ch, B.; Hk, M.S.; Sk, H.; Rr, K.; Mbvn, K.; Vp, B.; As, H.; Gs, L.; Sm, B.; Ms, M. Marker-assisted pyramiding of two major, broad-spectrum bacterial blight resistance genes, Xa21 and Xa33 into an elite maintainer line of rice, DRR17B. PLoS ONE 2018, 13, e0201271. [Google Scholar]
- Wang, S.; Liu, W.; Lu, D.; Lu, Z.; Wang, X.; Xue, J.; He, X. Distribution of bacterial blight resistance genes in the main cultivars and application of Xa23 in rice breeding. Front. Plant Sci. 2020, 11, 555228. [Google Scholar] [CrossRef]
- Hsu, Y.C.; Chiu, C.H.; Yap, R.; Tseng, Y.C.; Wu, Y.P. Pyramiding bacterial blight resistance genes in tainung82 for broad-spectrum resistance using marker-assisted selection. Int. J. Mol. Sci. 2020, 21, 1281. [Google Scholar] [CrossRef] [Green Version]
- Mi, J.; Yang, D.; Chen, Y.; Jiang, J.; Mou, H.; Huang, J.; Ouyang, Y.; Mou, T. Accelerated molecular breeding of a novel P/TGMS line with broad-spectrum resistance to rice blast and bacterial blight in two-line hybrid rice. Rice 2018, 11, 1–12. [Google Scholar] [CrossRef]
- Ramalingam, J.; Raveendra, C.; Savitha, P.; Vidya, V.; Chaithra, T.L.; Velprabakaran, S.; Saraswathi, R.; Ramanathan, A.; Arumugam Pillai, M.P.; Arumugachamy, S.; et al. Gene pyramiding for achieving enhanced resistance to bacterial blight, blast, and sheath blight diseases in rice. Front. Plant Sci. 2020, 11, 591457. [Google Scholar] [CrossRef]
- Ashkani, S.; Rafii, M.Y.; Shabanimofrad, M.; Miah, G.; Sahebi, M.; Azizi, P.; Tanweer, F.A.; Akhtar, M.S.; Nasehi, A. Molecular breeding strategy and challenges towards improvement of blast disease resistance in rice crop. Front. Plant Sci. 2015, 6, 886. [Google Scholar] [CrossRef] [Green Version]
- Ning, X.; Yunyu, W.; Aihong, L. Strategy for use of rice blast resistance genes in rice molecular breeding. Rice Sci. 2020, 27, 263–277. [Google Scholar] [CrossRef]
- Zaidi, S.S.; Mukhtar, M.S.; Mansoor, S. Genome editing: Targeting susceptibility genes for plant disease resistance. Trends Biotechnol. 2018, 36, 898–906. [Google Scholar] [CrossRef]
- Fukuoka, S.; Saka, N.; Koga, H.; Ono, K.; Shimizu, T.; Ebana, K.; Hayashi, N.; Takahashi, A.; Hirochika, H.; Okuno, K.; et al. Loss of function of a proline-containing protein confers durable disease resistance in rice. Science 2009, 325, 998–1001. [Google Scholar] [CrossRef] [PubMed]
- Iyer, A.S.; McCouch, S.R. The rice bacterial blight resistance gene xa5 encodes a novel form of disease resistance. Mol. Plant-Microbe Interact. 2004, 17, 1348–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, G.H.; Xia, Z.H.; Zhou, Y.L.; Wan, J.; Li, D.Y.; Chen, R.S.; Zhai, W.X.; Zhu, L.H. Testifying the rice bacterial blight resistance gene xa5 by genetic complementation and further analyzing xa5 (Xa5) in comparison with its homolog TFIIAγ1. Mol. Genet. Genom. 2006, 275, 354–366. [Google Scholar] [CrossRef]
- Antony, G.; Zhou, J.H.; Huang, S.; Li, T.; Liu, B.; White, F.; Yang, B. Rice xa13 recessive resistance to bacterial blight is defeated by induction of the disease susceptibility gene Os-11N3. Plant Cell 2010, 22, 3864–3876. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Sugio, A.; White, F.F. Os8N3 is a host disease-susceptibility gene for bacterial blight of rice. Proc. Natl. Acad. Sci. USA 2006, 103, 10503–10508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, Z.; Fu, B.; Yang, H.; Xu, C.; Li, Z.; Sanchez, A.; Park, Y.J.; Bennetzen, J.L.; Zhang, Q.; Wang, S. Targeting xa13, a recessive gene for bacterial blight resistance in rice. Theor. Appl. Genet. 2006, 112, 455–461. [Google Scholar] [CrossRef]
- Liu, Q.S.; Yuan, M.; Zhou, Y.; Li, X.H.; Xiao, J.H.; Wang, S.P. A paralog of the MtN3/saliva family recessively confers race-specific resistance to Xanthomonas oryzae in rice. Plant Cell Environ. 2011, 34, 1958–1969. [Google Scholar] [CrossRef]
- Zhou, J.; Peng, Z.; Long, J.; Sosso, D.; Liu, B.; Eom, J.S.; Huang, S.; Liu, S.; Vera Cruz, C.; Frommer, W.B.; et al. Gene targeting by the TAL effector PthXo2 reveals cryptic resistance gene for bacterial blight of rice. Plant J. 2015, 82, 632–643. [Google Scholar] [CrossRef]
- Tian, J.; Xu, G.; Yuan, M. Precise editing enables crop broad-spectrum resistance. Mol. Plant 2019, 12, 1542–1544. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Xu, X.; Gong, Q.; Li, Z.; Li, Y.; Wang, S.; Yang, Y.; Ma, W.; Liu, L.; Zhu, B.; et al. Engineering broad-spectrum bacterial blight resistance by simultaneously disrupting variable TALE-binding elements of multiple susceptibility genes in rice. Mol. Plant 2019, 12, 1434–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliva, R.; Ji, C.; Atienza-Grande, G.; Huguet-Tapia, J.C.; Perez-Quintero, A.; Li, T.; Eom, J.S.; Li, C.; Nguyen, H.; Liu, B.; et al. Broad-spectrum resistance to bacterial blight in rice using genome editing. Nat. Biotechnol. 2019, 37, 1344–1350. [Google Scholar] [CrossRef] [Green Version]
- Zafar, K.; Khan, M.Z.; Amin, I.; Mukhtar, Z.; Yasmin, S.; Arif, M.; Ejaz, K.; Mansoor, S. Precise CRISPR-Cas9 mediated genome editing in super basmati rice for resistance against bacterial blight by targeting the major susceptibility gene. Front. Plant Sci. 2020, 11, 575. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Luo, Y.; Vu, N.T.Q.; Shen, S.; Xia, K.; Zhang, M. CRISPR/Cas9-mediated mutation of OsSWEET14 in rice cv. Zhonghua11 confers resistance to Xanthomonas oryzae pv. oryzae without yield penalty. BMC Plant Biol. 2020, 20, 1–11. [Google Scholar] [CrossRef]
- Tao, H.; Shi, X.; He, F.; Wang, D.; Xiao, N.; Fang, H.; Wang, R.; Zhang, F.; Wang, M.; Li, A.; et al. Engineering broad-spectrum disease-resistant rice by editing multiple susceptibility genes. J. Integr. Plant Biol. 2021, 63, 1639–1648. [Google Scholar] [CrossRef]
- Li, W.; Deng, Y.; Ning, Y.; He, Z.; Wang, G.L. Exploiting broad-spectrum disease resistance in crops: From molecular dissection to breeding. Annu. Rev. Plant Biol. 2020, 71, 575–603. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Abdelrahman, M.; Gao, Y.; Ji, Z.; Mishra, R.; Sun, H.; Sui, Y.; Wu, C.; Wang, C.; Zhao, K. Engineering broad-spectrum resistance to bacterial blight by CRISPR-Cas9-mediated precise homology directed repair in rice. Mol. Plant 2021, 14, 1215–1218. [Google Scholar] [CrossRef]
- Molla, K.A.; Karmakar, S.; Chanda, P.K.; Sarkar, S.N.; Datta, S.K.; Datta, K. Tissue-specific expression of Arabidopsis NPR1 gene in rice for sheath blight resistance without compromising phenotypic cost. Plant Sci. 2016, 250, 105–114. [Google Scholar] [CrossRef]
- Xu, G.; Yuan, M.; Ai, C.; Liu, L.; Zhuang, E.; Karapetyan, S.; Wang, S.; Dong, X. uORF-mediated translation allows engineered plant disease resistance without fitness costs. Nature 2017, 545, 491–494. [Google Scholar] [CrossRef]
- Li, Z.; Huang, J.; Wang, Z.; Meng, F.; Zhang, S.; Wu, X.; Zhang, Z.; Gao, Z. Overexpression of Arabidopsis nucleotide-binding and leucine-rich repeat genes RPS2 and RPM1(D505V) confers broad-spectrum disease resistance in rice. Front. Plant Sci. 2019, 10, 417. [Google Scholar] [CrossRef] [PubMed]
- Krattinger, S.G.; Sucher, J.; Selter, L.L.; Chauhan, H.; Zhou, B.; Tang, M.; Upadhyaya, N.M.; Mieulet, D.; Guiderdoni, E.; Weidenbach, D.; et al. The wheat durable, multipathogen resistance gene Lr34 confers partial blast resistance in rice. Plant Biotechnol. J. 2016, 14, 1261–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pokhrel, S.; Ponniah, S.K.; Jia, Y.; Yu, O.; Manoharan, M. Transgenic rice expressing isoflavone synthase gene from soybean shows resistance against blast fungus (Magnaporthe oryzae). Plant Dis. 2021. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene Name | Protein Type | Isolates or Pathogens 1 | Chromosome | Reference |
|---|---|---|---|---|
| Pi64 | NBS-LRR | 9 Mo isolates | 1 | [6] |
| Pizh | NBS-LRR | 31 Mo isolates | 6 | [8] |
| Pigm | NBS-LRR | 30 Mo isolates | 6 | [7] |
| Pi50 | NBS-LRR | 20 Mo isolates | 6 | [22] |
| Pi56(t) | NBS-LRR | 19 Mo isolates | 9 | [5] |
| Pi54rh | NBS-LRR | 7 Mo isolates | 11 | [4] |
| Ptr | ARM repeat protein | 331 Mo isolates | 12 | [9] |
| Pita2 | ARM repeat protein | 64 Mo isolates | 12 | [10] |
| Xa7 | Executor R protein | 8 Xoo isolates | 6 | [27] |
| Xa23 | Executor R protein | 39 Xoo isolates | 11 | [28] |
| xa41(t) | Sugar transporter (SWEET) | 18 Xoo isolates | 11 | [29] |
| Xa47(t) | NBS-LRR | 10 Xoo isolates | 11 | [30] |
| Gene Name | Protein Type | Isolates or Pathogens 1 | Chr 2 | Reference |
|---|---|---|---|---|
| Bsr-d1 | C2H2-type transcription factor | 9 Mo isolates | 3 | [36,37] |
| OsMYB30 | MYB transcription factor | 5 Mo isolates | 2 | [38] |
| OsNAC60 | NAC transcription factor | 20 Mo isolates | 12 | [39] |
| OsWRKY45 | WRKY transcription factor | 1 Mo isolate | 5 | [40] |
| PIBP1 | RRM (RNA recognition motif) protein | 3 Mo isolates | 3 | [41] |
| OsBBI1 | RING E3 ubiquitin ligase | 7 Mo isolates | 6 | [42] |
| LHCB5 | Light-harvesting complex II protein | 21 Mo isolates | 11 | [43] |
| OsXB25 | Plant-specific ankyrin-repeat (PANK) protein | 1 Xoo isolate | 9 | [44] |
| OsBiP3 | Endoplasmic reticulum (ER) chaperone, luminal-binding protein 3 | 2 Xoo isolates | 5 | [45] |
| OsNPR1 | BTB/POZ-ankyrin repeat protein | 1 Mo isolate, 2 Xoo isolates | 1 | [46] |
| OsCRK6 | Cysteine-rich-receptor-like kinases | 1 Xoo isolate | 7 | [47] |
| OsCRK10 | Cysteine-rich-receptor-like kinases | 1 Xoo isolate | 7 | [47] |
| OsCDPK1 | Calcium-dependent protein kinases | Xoo | 3 | [48] |
| OsILA1 | Raf-like MAPKKK | 9 Xoo isolates | 6 | [49] |
| lc7 | Ferredoxin-dependent glutamate synthase1 | 7 Xoo isolates | 7 | [50] |
| OsLYP4 | Lysin motif-containing proteins | 1 Mo isolate, 1 Xoo isolate, 1 Xoc isolate | 9 | [51] |
| OsLYP6 | Lysin motif-containing proteins | 1 Mo isolate, 1 Xoo isolate, 1 Xoc isolate | 6 | [51] |
| OsWRKY67 | WRKY transcription factor | 2 Mo isolates, 2 Xoo isolates | 5 | [52] |
| IPA1 | Transcription factors | 12 Mo isolates | 8 | [53,54] |
| OsTFIIAα, | Transcription factor IIA subunits | 10 Xoo isolates, 6 Xoc isolates | 5 | [55] |
| OsTFIIAβ | Transcription factor IIA subunits | 10 Xoo isolates, 6 Xoc isolates | 5 | [55] |
| OsGLP2-1 | Germin-like protein | 1 Mo isolate, 1 Xoo isolate | 2 | [56] |
| OsSnRK1a | Sucrose nonfermenting 1-related protein kinase 1 | 1 Mo isolate, 1 Xoo isolate, 1 Cm isolate and 1 Rs isolate | 5 | [57] |
| OSK35/OsSnRK1b | Sucrose nonfermenting 1-related protein kinases | 1 Mo isolate, 1 Xoo isolate | 3 | [58] |
| OsCPK4 | Calcium-dependent protein kinase | 1 Mo isolate, 1 Xoo isolate | 2 | [59] |
| BSR1 | BIK1-like receptor-like cytoplasmic kinase | 2 Mo isolates, 3 Xoo isolates, 1 Bg isolate, 1 Cm isolate, rice stripe virus | 9 | [60] |
| OsBAG4 | Ubiquitin-like and BAG domain | 1 Mo isolate, 1 Xoo isolate | 1 | [61] |
| EBR1 | RING-Type E3 Ligase | 1 Mo isolate, 6 Xoo isolates | 5 | [61] |
| SPIN6 | Rho GTPase-activating protein (RhoGAP) | 1 Mo isolate, 1 Xoo isolate | 7 | [62] |
| OsWAK25 | Wall-associated kinases | 2 Mo isolates, 1 Xoo isolate | 3 | [63] |
| OsCUL3a | Cullin 3-based RING E3 ubiquitin ligases | 1 Mo isolate, 3 Xoo isolates | 2 | [64] |
| OsDRP1E | Dynamin-related protein | 1 Mo isolate, 1 Xoo isolate | 9 | [65] |
| SPL33 | Eukaryotic translation elongation factor 1 alpha (eEF1A)-like protein | 12 Mo isolates, 11 Xoo isolates | 1 | [66] |
| LMM5.1 | Eukaryotic translation elongation factor 1A (eEF1A)-like protein | 6 Mo isolates, 5 Xoo isolates | 1 | [67] |
| LMM5.4 | Eukaryotic translation elongation factor 1A (eEF1A)-like protein | 6 Mo isolates, 5 Xoo isolates | 4 | [67] |
| LML1 | Eukaryotic release factor 1 (eRF1) protein | 4 Mo isolates, 6 Xoo isolates | 4 | [68] |
| OsABA2 | Xanthoxin dehydrogenase | 2 Mo isolates, 4 Xoo isolates | 3 | [69] |
| SPL35 | CUE (coupling of ubiquitin conjugation to ER degradation) domain-containing protein | 8 Mo isolates, 4 Xoo isolates | 3 | [70] |
| OsHDT701 | Histone deacetylase | 4 Mo isolates, 1 Xoo isolate | 5 | [71] |
| OsMPK15 | Mitogen-activated protein kinase | 2 Mo isolates, 2 Xoo isolates | 11 | [72] |
| Bsr-k1 | Tetratricopeptide repeats (TPRs)containing protein | 7 Mo isolates, 10 Xoo isolates | 10 | [73] |
| OsALDH2B1 | Aldehyde dehydrogenase | 1 Mo isolate, 1 Xoo isolate, 1 Xoc isolate | 6 | [74] |
| OsPAL4 | Phenylalanine ammonia-lyase | 1 Mo isolate, 1 Xoo isolate, 1 Xoc isolate | 2 | [75] |
| OsHsfB4d | Class B heat-shock factor | 1 Xoo isolate, 1 Xoc isolate | 3 | [76] |
| OsHsp18.0-CI | Heat-shock proteins | 5 Xoc isolates | 3 | [77] |
| OsPAD4 | Phytoalexin-deficient 4 | 2 Xoo isolates, 1 Xoc isolate | 11 | [78] |
| OsGRXS15 | Glutaredoxins family proteins | 1 Xoo isolate, 1 Ff isolate | 1 | [79] |
| OsNH5N16 | Pathogenesis-related genes 1 homologs (NHs) | 1 Xoo isolates, 1 Ff isolate | 11 | [80] |
| OsASR2 | Abscisic acid, stress, and ripening 2 protein | 1 Xoo isolate, 1 Xoc isolate | 11 | [81] |
| Os2H16 | Short-chain peptide-encoding protein | 1 Xoo isolate, 1 Xoc isolate | 6 | [82] |
| OsGF14e | 14-3-3 protein | 1 Xoo isolate, 1 Rs isolate | 2 | [83] |
| OsWRKY30 | WRKY transcription factors | 1 Mo isolate, 1 Rs isolate | 8 | [84] |
| OsACS2 | 1-aminocyclopropane-1-carboxylic acid synthase | 2 Mo isolates, 1 Rs isolate | 4 | [85] |
| OsMESL | Methyl esterase-like protein | 1 Mo isolate, 1 Xoo isolate, 1 Rs isolate | 7 | [86] |
| OsBON1 | Copine protein | 1 Mo isolate, 3 Xoo isolates, 1 Rs isolate | 2 | [87] |
| OsBON3 | Copine protein | 1 Mo isolate, 3 Xoo isolates, 1 Rs isolate | 5 | [87] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Zhu, Y.; Shi, H.; Qiu, J.; Ding, X.; Kou, Y. Recent Progress in Rice Broad-Spectrum Disease Resistance. Int. J. Mol. Sci. 2021, 22, 11658. https://doi.org/10.3390/ijms222111658
Liu Z, Zhu Y, Shi H, Qiu J, Ding X, Kou Y. Recent Progress in Rice Broad-Spectrum Disease Resistance. International Journal of Molecular Sciences. 2021; 22(21):11658. https://doi.org/10.3390/ijms222111658
Chicago/Turabian StyleLiu, Zhiquan, Yujun Zhu, Huanbin Shi, Jiehua Qiu, Xinhua Ding, and Yanjun Kou. 2021. "Recent Progress in Rice Broad-Spectrum Disease Resistance" International Journal of Molecular Sciences 22, no. 21: 11658. https://doi.org/10.3390/ijms222111658
APA StyleLiu, Z., Zhu, Y., Shi, H., Qiu, J., Ding, X., & Kou, Y. (2021). Recent Progress in Rice Broad-Spectrum Disease Resistance. International Journal of Molecular Sciences, 22(21), 11658. https://doi.org/10.3390/ijms222111658