
Immortalizing Cells for Human Consumption


{kind=link}
{kind=link}
{kind=link}
Abstract
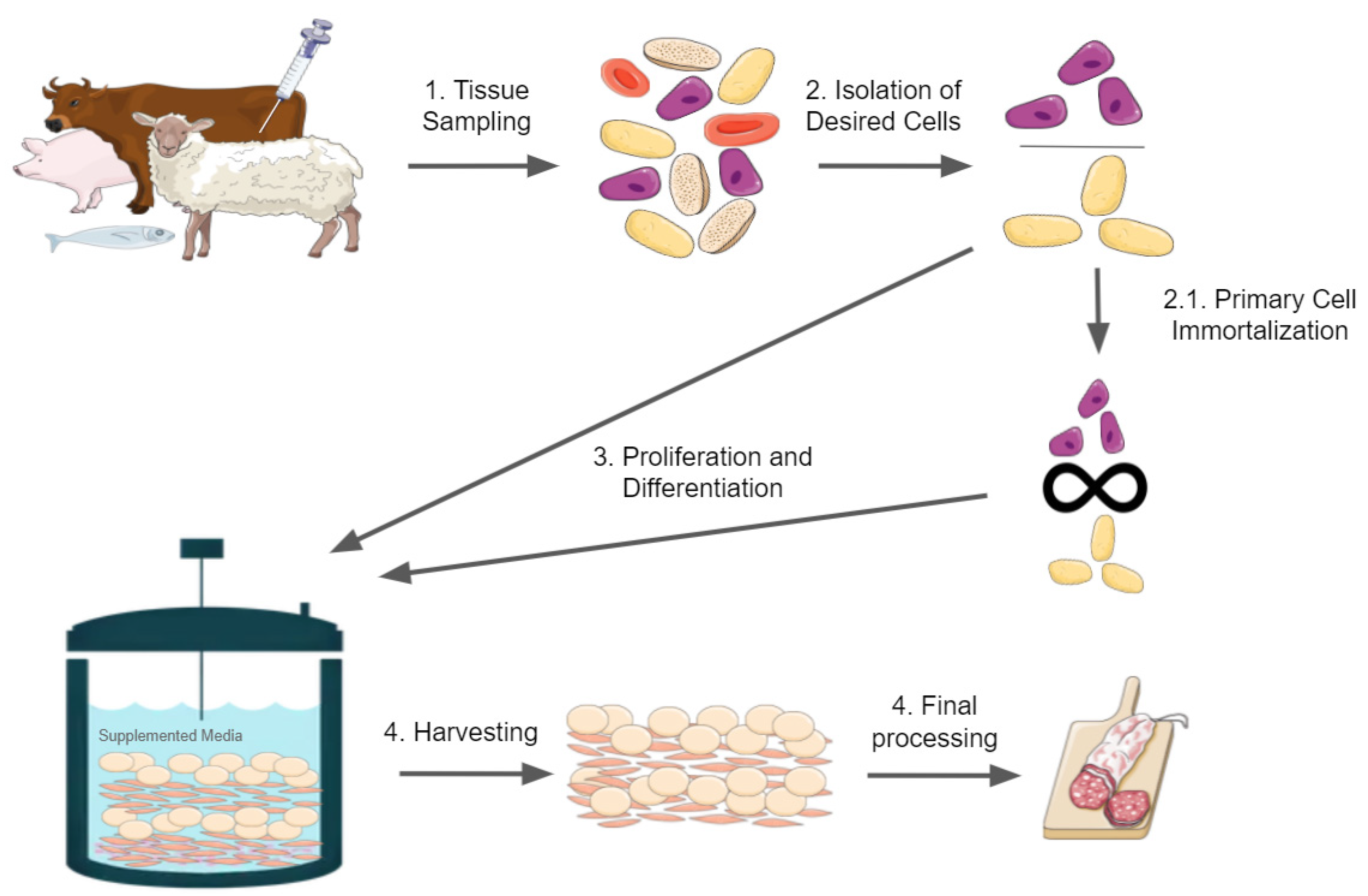
:1. Introduction
2. Methods for Establishing Immortal Cell Lines
2.1. Spontaneous Immortalization of Cell Lines
2.2. Establishment by Telomerase
2.3. Establishment by Inactivation of the p53/p16/Rb Stress Response

2.4. Combined Approaches to Establishment
3. Unique Challenges for Cell Lines Used in Cellular Agriculture
3.1. Species-to-Species Variation in Establishment Difficulty
3.2. The Food Safety of Cell Lines
3.3. Cell Lines and the Consumer Acceptance of Cultured Meat
3.4. Other Unique Attributes to Select for in Cultured Meat Cell Lines
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Post, M.J.; Levenberg, S.; Kaplan, D.L.; Genovese, N.; Fu, J.; Bryant, C.; Negowetti, N.; Verzijden, K.; Moutsatsou, P. Scientific, sustainability and regulatory challenges of cultured meat. Nat. Food 2020, 1, 403–415. [Google Scholar] [CrossRef]
- Hallman, W.K.; Hallman, W.K., II. An empirical assessment of common or usual names to label cell-based seafood products. J. Food Sci. 2020, 85, 2267–2277. [Google Scholar] [CrossRef] [PubMed]
- Labeling of Meat or Poultry Products Comprised of or Containing Cultured Animal Cells; Federal Register: Washington, DC, USA, 2021.
- Boler, D.; Woerner, D. What is meat? A perspective from the American Meat Science Association. Anim. Front. 2017, 7, 8–11. [Google Scholar] [CrossRef] [Green Version]
- Alexandratos, N.; Bruinsma, J. World Agriculture Towards 2030/2050: The 2012 Revision; ESA Working Paper No. 12-03; FAO: Rome, Italy, 2012; Available online: http://www.fao.org/3/ap106e/ap106e.pdf (accessed on 24 October 2021).
- DiMaio, T.; This Scientist Is Developing New Cell Lines for Slaughter-Free Meat. The Good Food Institute. 2019. Available online: https://gfi.org/blog/gareth-sullivan-cell-lines-research-grant/ (accessed on 24 October 2021).
- Maqsood, M.I.; Matin, M.M.; Bahrami, A.R.; Ghasroldasht, M.M. Immortality of Cell Lines: Challenges and Advantages of Establishment. Cell Biol. Int. 2013, 37, 1038–1045. [Google Scholar] [CrossRef] [PubMed]
- Ben-Arye, T.; Levenberg, S. Tissue Engineering for Clean Meat Production. Front. Sustain. Food Syst. 2019, 3, 46. [Google Scholar] [CrossRef]
- ATCC. Available online: https://atcc.org/ (accessed on 27 June 2021).
- Listrat, A.; Lebret, B.; Louveau, I.; Astruc, T.; Bonnet, M.; Lefaucheur, L.; Picard, B.; Bugeon, J. How Muscle Structure and Composition Influence Meat and Flesh Quality. Sci. World J. 2020, 2016, 3182746. [Google Scholar] [CrossRef] [Green Version]
- Sie, C.; Barnett, J. Consumer Acceptance of Cultured Meat: An Updated Review (2018–2020). Appl. Sci. 2020, 10, 5201. [Google Scholar] [CrossRef]
- Liu, J.; Hocquette, É.; Ellies-Oury, M.P.; Chriki, S.; Hocquette, J.F. Chinese Consumers’ Attitudes and Potential Acceptance toward Artificial Meat. Foods 2021, 10, 353. [Google Scholar] [CrossRef] [PubMed]
- Kerafast’s Partnership with The Good Food Institute Links Researchers to Cell Lines. Kerafast. 2021. Available online: https://news.kerafast.com/2021/07/13/kerafasts-partnership-with-the-good-food-institute-links-researchers-to-cell-lines/ (accessed on 24 October 2021).
- Jedrzejczak-Silicka, M. History of Cell Culture. In New Insights into Cell Culture Technology; IntechOpen: London, UK, 2017; pp. 1–41. [Google Scholar] [CrossRef] [Green Version]
- Yao, T.; Asayama, Y. Animal-cell culture media: History, characteristics, and current issues. Reprod. Med. Biol. 2017, 16, 99–117. [Google Scholar] [CrossRef] [PubMed]
- Foster, D.N.; Foster, L.K. Immortalized Cell Lines Protein Production. U.S. Patent US6207415B1, 27 March 2001. [Google Scholar]
- Court of Justice of the European Union. Organisms Obtained by Mutagenesis Are GMOs and Are, in Principle, Subject to the Obligations Laid Down by the GMO Directive. Press Release No 111/18. Luxembourg. 25 July 2018. Available online: https://curia.europa.eu/jcms/upload/docs/application/pdf/2018-07/cp180111en.pdf (accessed on 24 October 2021).
- Rakers, S. Fish Matters—Characterization of Fish Cell Cultures for Cell-Based Fish Production. ISCCM. 2020. Available online: https://caos.community/isccm/ (accessed on 16 December 2020).
- Rangarajan, A.; Hong, S.J.; Gifford, A.; Weinberg, R.A. Species- and Cell Type-Specific Requirements for Cellular Transformation. Cancer Cell 2004, 6, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Shefferson, R.; Jones, O.; Salguero-Gómez, R. (Eds.) The Evolution of Senescence in the Tree of Life; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar] [CrossRef]
- Hayflick, L. The Limited in Vitro Lifetime of Human Diploid Cell Strains. Exp. Cell Res. 1965, 37, 614–636. [Google Scholar] [CrossRef]
- Holt, S.E.; Wright, W.E.; Shay, J.W. Regulation of telomerase activity in immortal cell lines. Mol. Cell Biol. 1996, 16, 2932–2939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Jiang, J.; Sottile, V.; McWhir, J.; Lebkowski, J.; Carpenter, M.K. Immortalized Fibroblast-Like Cells Derived from Human Embryonic Stem Cells Support Undifferentiated Cell Growth. Stem Cells J. 2004, 22, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Harada, H.; Nakagawa, H.; Oyama, K.; Takaoka, M.; Andl, C.D.; Jacobmeier, B.; Rustgi, A.K. Telomerase Induces Immortalization of Human Esophageal Keratinocytes Without P16INK4a Inactivation. Mol. Cancer Res. 2003, 11, 729–738. Available online: https://mcr.aacrjournals.org/content/molcanres/1/10/729.full.pdf (accessed on 24 October 2021).
- Chang, M.W.-F.; Grillari, J.; Mayrhofer, C.; Fortschegger, K.; Allmaier, G.; Marzban, G.; Katinger, H.; Voglauer, R. Comparison of Early Passage, Senescent and HTERT Immortalized Endothelial Cells. Exp. Cell Res. 2005, 309, 121–136. [Google Scholar] [CrossRef]
- Chen, J. The Cell-Cycle Arrest and Apoptotic Functions of p53 in Tumor Initiation and Progression. Cold Spring Harb. Perspect. Med. 2016, 6, a026104. [Google Scholar] [CrossRef]
- Takahashi, A.; Ohtani, N.; Hara, E. Irreversibility of cellular senescence: Dual roles of p16INK4a/Rb-pathway in cell cycle control. Cell Div. 2007, 2, 10. [Google Scholar] [CrossRef] [Green Version]
- Yamada, Y.; Liao, G.-R.; Tseng, C.-Y.; Tseng, Y.-Y.; Hsu, W.-L. Establishment and Characterization of Transformed Goat Primary Cells by Expression of Simian Virus 40 Large T Antigen for Orf Virus Propagations. PLoS ONE 2019, 14, e0226105. [Google Scholar] [CrossRef]
- Jin, X.; Lee, J.S.; Kwak, S.; Lee, S.Y.; Jung, J.E.; Kim, T.K.; Xu, C.; Hong, Z.; Li, Z.; Kim, S.M.; et al. Establishment and Characterization of Three Immortal Bovine Muscular Epithelial Cell Lines. Mol. Cells 2006, 21, 29–33. [Google Scholar]
- Zhang, X.; Yu, D.; Wu, Y.; Gu, T.; Ma, N.; Dong, S.; Yao, Y.G. Establishment and transcriptomic features of an immortalized hepatic cell line of the Chinese tree shrew. Appl. Microbiol. Biotechnol. 2020, 104, 8813–8823. [Google Scholar] [CrossRef]
- Ahuja, D.; Sáenz-Robles, M.; Pipas, J. SV40 large T antigen targets multiple cellular pathways to elicit cellular transformation. Oncogene 2005, 24, 7729–7745. [Google Scholar] [CrossRef] [Green Version]
- Chou, J.Y. Differentiated Mammalian Cell Lines Immortalized by Temperature Sensitive Tumor Viruses. Mol. Endocrinol. 1989, 3, 1511–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klingelhutz, A.J.; Barber, S.A.; Smith, P.P.; Dyer, K.; McDougall, J.K. Restoration of telomeres in human papillomavirus-immortalized human anogenital epithelial cells. Mol. Cell. Biol. 1994, 14, 961–969. [Google Scholar] [CrossRef] [Green Version]
- Oh, H.M.; Oh, J.M.; Choi, S.C.; Kim, S.W.; Han, W.C.; Kim, T.H.; Park, D.S.; Jun, C.D. An efficient method for the rapid establishment of Epstein–Barr virus immortalization of human B lymphocytes. Cell Prolif. 2003, 36, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Counter, C.M.; Avilion, A.A.; LeFeuvre, C.E.; Stewart, N.G.; Greider, C.W.; Harley, C.B.; Bacchetti, S. Telomere shortening associated with chromosome instability is arrested in immortal cells which express telomerase activity. EMBO J. 1992, 11, 1921–1929. [Google Scholar] [CrossRef]
- Shay, J.W.; Wright, W.E.; Werbin, H. Defining the molecular mechanisms of human cell immortalization. Biochim. Biophys. Acta 1991, 1072, 1–7. [Google Scholar] [CrossRef]
- Toss, A.; Venturelli, M.; Peterle, C.; Piacentini, F.; Cascinu, S.; Cortesi, L. Molecular Biomarkers for Prediction of Targeted Therapy Response in Metastatic Breast Cancer: Trick or Treat? Int. J. Mol. Sci. 2017, 18, 85. [Google Scholar] [CrossRef] [Green Version]
- Ryu, W. Molecular Virology of Human Pathogenic Viruses; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Thorley, M.; Duguez, S.; Mazza, E.; Valsoni, S.; Bigot, A.; Mamchaoui, K.; Harmon, B.; Voit, T.; Mouly, V.; Duddy, W. Skeletal Muscle Characteristics Are Preserved in hTERT/cdk4 human myogenic cell lines. Skelet. Muscle 2016, 6, 43. [Google Scholar] [CrossRef] [Green Version]
- Genovese, N.; Desmet, D.N.; Schulze, E. Methods for Extending the Replicative Capacity of Somatic Cells during an Ex Vivo Cultivation Process. Patent WO2017124100A1, 20 July 2017. [Google Scholar]
- Stadler, G.; Chen, J.C.; Wagner, K.; Robin, J.; Shay, J.; Emerson, C.; Wright, W. Establishment of clonal myogenic cell lines from severely affected dystrophic muscles—CDK4 maintains the myogenic population. Skelet. Muscle 2011, 1, 12. [Google Scholar] [CrossRef] [Green Version]
- Bomkamp, C. GFI’s Sustainable Seafood Initiative Announces New Strategic Partnership and Grantees. 2020. Available online: https://www.gfi.org/blog-gfi-ssi-announcements (accessed on 24 October 2021).
- Wang, Y.; Chen, S.; Yan, Z.; Pei, M. A prospect of cell immortalization combined with matrix microenvironmental optimization strategy for tissue engineering and regeneration. Cell Biosci 2019, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Ong, K.; Johnston, J.; Datar, I.; Sewalt, V.; Holmes, D.; Shatkin, J.A. Food Safety Considerations and Research Priorities for the Cultured Meat and Seafood Industry. Compr. Rev. Food Sci. Food Saf. 2021, 1–28. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Mesnage, R.; Tsatsakis, A.M.; Golokhvast, K.S.; Yang, S.H.; Antoniou, M.N.; Chung, G. Addressing concerns over the fate of DNA derived from genetically modified food in the human body: A review. Food Chem. Toxicol. 2019, 124, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Routray, I.; Mahmood, A.; Ngwa, N.E.; Tasleem, M.; Sahin, K.; Kucuk, O.; Ali, S. Cell line cross-contamination and accidental co-culture. J. Stem Cell Res. Ther. 2016, 1, 179–185. [Google Scholar] [CrossRef]
- Geraghty, R.J.; Capes-Davis, A.; Davis, J.M.; Downward, J.; Freshney, R.I.; Knezevic, I.; Lovell-Badge, R.; Masters, J.R.W.; Meredith, J.; Stacey, G.N.; et al. Guidelines for the Use of Cell Lines in Biomedical Research. Br. J. Cancer 2014, 111, 1021–1046. [Google Scholar] [CrossRef] [Green Version]
- Ben-David, U.; Siranosian, B.; Ha, G.; Tang, H.; Oren, Y.; Hinohara, K.; Strathdee, C.A.; Dempster, J.; Lyons, N.J.; Burns, R.; et al. Genetic and Transcriptional Evolution Alters Cancer Cell Line Drug Response. Nature 2018, 560, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Fountain, D.; Ralston, M.; Higgins, N.; Gorlin, J.; Uhl, L.; Wheeler, C.; Antin, J.; Churchill, W.; Benjamin, R. Liquid nitrogen freezers: A potential source of microbial contamination of hematopoietic stem cell components. Transfusion 1997, 37, 585–591. [Google Scholar] [CrossRef]
- FDA. Characterization and Qualification of Cell Substrates and Other Biological Materials Used in the Production of Viral Vaccines for Infectious Disease Indications. Guidance for Industry. 2010. Available online: https://www.fda.gov/media/78428/download (accessed on 24 October 2021).
- FDA. Points to Consider in the Characterization of Cell Lines Used to Produce Biologicals. 1993. Available online: https://www.fda.gov/media/76255/download (accessed on 24 October 2021).
- FDA. Points to Consider in the Manufacture and Testing of Therapeutic Products for Human Use Derived from Transgenic Animals. 1995. Available online: https://www.fda.gov/media/76253/download (accessed on 24 October 2021).
- FDA. Approved Cellular and Gene Therapy Products. 2021. Available online: https://www.fda.gov/vaccines-blood-biologics/cellular-gene-therapy-products/approved-cellular-and-gene-therapy-products (accessed on 24 October 2021).
- Waltz, E. Club-goers take first bites of lab-made chicken. Nat. Biotechnol. 2021, 39, 257–258. [Google Scholar] [CrossRef] [PubMed]
- SFA. Requirements for the Safety Assessment of Novel Foods. 2020. Available online: https://www.sfa.gov.sg/docs/default-source/food-import-and-export/Requirements-on-safety-assessment-of-novel-foods_23-Nov-2020.pdf (accessed on 24 October 2021).
- FDA and USDA. USDA and FDA Joint Public Meeting on the Use of Cell Culture Technology to Develop Products Derived from Livestock and Poultry. 2018. Available online: https://www.fda.gov/food/workshops-meetings-webinars-food-and-dietary-supplements/joint-public-meeting-use-cell-culture-technology-develop-products-derived-livestock-and-poultry (accessed on 24 October 2021).
- Froggatt, A.; Wellesley, L.; Meat Analogues: Considerations for the EU. Chatham House, the Royal Institute of International Affairs. 2019. Available online: https://www.chathamhouse.org/sites/default/files/2020-12/2019-02-18-meat-analogues.pdf (accessed on 24 October 2021).
- EU. Regulation (EU) 2015/2283 of the European Parliament and of the Council of 25 November 2015 on Novel Foods, Amending Regulation (EU) No 1169/2011 of the European Parliament and of the Council and Repealing Regulation (EC) No 258/97 of the European Parliament and of the Council and Commission Regulation (EC) No 1852/2001. 2015. Available online: http://data.europa.eu/eli/reg/2015/2283/2021-03-27 (accessed on 24 October 2021).
- Szejda, K.; Bryant, C.J.; Urbanovich, T. US and UK Consumer Adoption of Cultivated Meat: A Segmentation Study. Foods 2021, 10, 1050. [Google Scholar] [CrossRef]
- Bryant, C.; van Nek, L.; Rolland, N.C.M. European Markets for Cultured Meat: A Comparison of Germany and France. Foods 2020, 9, 1152. [Google Scholar] [CrossRef]
- Siegrist, M.; Hartmann, C. Consumer acceptance of novel food technologies. Nat. Food 2020, 1, 343–350. [Google Scholar] [CrossRef]
- Verbeke, W.; Hung, Y.; Baum, C.M.; De Steur, H. The power of initial perceived barriers versus motives shaping consumers’ willingness to eat cultured meat as a substitute for conventional meat. Livest. Sci. 2021, 253, 104705. [Google Scholar] [CrossRef]
- Siegrist, M.; Sütterlin, B.; Hartmann, C. Perceived naturalness and evoked disgust influence acceptance of cultured meat. Meat Sci. 2018, 139, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Gaskell, G.; Allansdottir, A.; Allum, N.; Castro, P.; Esmer, Y.; Fischler, C.; Jackson, J.; Kronberger, N.; Hampel, J.; Mejlgaard, N.; et al. The 2010 Eurobarometer on the life sciences. Nat. Biotechnol. 2011, 29, 113–114. [Google Scholar] [CrossRef] [PubMed]
- Kronberger, N.; Wagner, W.; Nagata, M. How Natural Is “More Natural”? The Role of Method, Type of Transfer, and Familiarity for Public Perceptions of Cisgenic and Transgenic Modification. Sci. Commun. 2014, 36, 106–130. [Google Scholar] [CrossRef]
- Scott, S.; Rozin, P. Are Additives Unnatural? Generality and Mechanisms of Additivity Dominance. Judgm. Decis. Mak. 2017, 12, 572–583. Available online: http://journal.sjdm.org/17/17608b/jdm17608b.pdf (accessed on 24 October 2021).
- Gaskell, G.; Allum, N.; Bauer, M.; Durant, J.; Allansdottir, A.; Bonfadelli, H.; Boy, D.; de Cheveigné, S.; Fjaestad, B.; Gutteling, J.M.; et al. Biotechnology and the European public. Nat. Biotechnol. 2000, 18, 935–938. [Google Scholar] [CrossRef]
- Ritchie, H.; Roser, M.; Meat and Dairy Production. OurWorldInData.Org. 2017. Available online: https://ourworldindata.org/meat-production (accessed on 15 October 2021).
- Bryant, C.; Szejda, K.; Parekh, N.; Deshpande, V.; Tse, B. A Survey of Consumer Perceptions of Plant-Based and Clean Meat in the USA, India, and China. Front. Sustain. Food Syst. 2019, 3, 11. [Google Scholar] [CrossRef]
- Wilks, M.; Phillips, C.; Fielding, K.; Hornsey, M. Testing potential psychological predictors of attitudes towards cultured meat. Appetite 2019, 136, 137–145. [Google Scholar] [CrossRef]
- Baum, C.; Bröring, S.; Lagerkvistc, C.J. Information, attitudes, and consumer evaluations of cultivated meat. Food Qual. Prefer. 2021, 92, 104225. [Google Scholar] [CrossRef]
- Hwang, J.; You, J.; Moon, J.; Jeong, J. Factors Affecting Consumers’ Alternative Meats Buying Intentions: Plant-Based Meat Alternative and Cultured Meat. Sustainability 2020, 12, 5662. [Google Scholar] [CrossRef]
- Dempsey, C.; Bryant, C.J. Cultured meat: Do Chinese consumers have an appetite? OSF Prepr. 2020, 1–40. [Google Scholar] [CrossRef]
- Abdullah, F.A.A.; Borilova, G.; Steinhauserova, I. Halal Criteria Versus Conventional Slaughter Technology. Animals 2019, 9, 530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chriki, S.; Hocquette, J.-F. The Myth of Cultured Meat: A Review. Front. Nutr. 2020, 7, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryant, C. Culture, meat, and cultured meat. J. Anim. Sci. 2020, 98, 1–7. [Google Scholar] [CrossRef]
- Kenigsberg, J.A.; Zivotofsky, A.Z. A Jewish Religious Perspective on Cellular Agriculture. Front. Sustain. Food Syst. 2020, 3, 128. [Google Scholar] [CrossRef]
- Baharuddin, A.; Ghazali, M.; Harun, M.; Mutalib, L.; Shaarani, I.; Alauddin, M.; Razali, M. Three-Dimensional (3D) Printed Halal Meat: Do We Need a New Regulatory Framework? INSLA E-Proc. 2020, 3, 438–449. Available online: https://insla.usim.edu.my/index.php/eproceeding/article/view/49 (accessed on 24 October 2021).
- Hamdan, M.; Ramli, M.; Huri, N.; Rahman, N.; Abdullah, A. Will Muslim consumers replace livestock slaughter with cultured meat in the market? Trends Food Sci. Technol. 2021, 109, 729–732. [Google Scholar] [CrossRef]
- Elfenbein, A.; Kolbeck, J. Ex Vivo Meat Production. Patent WO2018227016A1, 13 December 2018. [Google Scholar]
- Genovese, N.J.; Roberts, R.M.; Telugu, B.P.V.L. Method for Scalable Skeletal Muscle Lineage Specification and Cultivation. U.S. Patent US20160227830A1, 11 August 2016. [Google Scholar]
- Beurrier, C.; Anticipatory Life Cycle and Cost of Goods Assessment of Cultured Meat Manufacturing Processes. ISCCM. 2020. Available online: https://caos.community/isccm/ (accessed on 15 December 2020).
- Genovese, N.J.; Schulze, E.N.; Desmet, D.N. Compositions and Methods for Increasing the Efficiency of Cell Cultures Used for Food Production. Patent WO2019014652A1, 17 January 2019. [Google Scholar]
- Schneider, M.; Marison, I.; von Stockar, U. The importance of ammonia in mammalian cell culture. J. Biotechnol. 1996, 46, 161–185. [Google Scholar] [CrossRef]
- Fraeye, I.; Kratka, M.; Vandenburgh, H.; Thorrez, L. Sensorial and Nutritional Aspects of Cultured Meat in Comparison to Traditional Meat: Much to Be Inferred. Front. Nutr. 2020, 7, 35. [Google Scholar] [CrossRef] [Green Version]
- Rowat, A. (University of California, Los Angeles, California, USA). Personal communication, 2020.
- Kang, D.H.; Louis, F.; Liu, H.; Shimoda, H.; Nishiyama, Y.; Nozawa, H.; Kakitani, M.; Takagi, D.; Kasa, D.; Nagamori, E.; et al. Engineered whole cut meat-like tissue by the assembly of cell fibers using tendon-gel integrated bioprinting. Nat. Commun. 2020, 12, 5059. [Google Scholar] [CrossRef] [PubMed]
- Stout, A.; Mirliani, A.; Soule-Albridge, E.; Cohen, J.; Kaplan, D. Engineering carotenoid production in mammalian cells for nutritionally enhanced cell-cultured foods. Metab. Eng. 2020, 62, 126–137. [Google Scholar] [CrossRef]
- Simsa, R.; Yuen, J.; Stout, A.; Rubio, N.; Fogelstrand, P.; Kaplan, D.L. Extracellular Heme Proteins Influence Bovine Myosatellite Cell Proliferation and the Color of Cell-Based Meat. Foods 2019, 8, 521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fish, K.; Rubio, N.; Stout, A.; Yuen, J.; Kaplan, D. Prospects and Challenges for Cell-Cultured Fat as a Novel Food Ingredient. Trends Food Sci. Technol. 2020, 98, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Ramayo-Caldas, Y.; Renand, G.; Ballester, M.; Saintilan, R.; Rocha, D. Multi-breed and multi-trait co-association analysis of meat tenderness and other meat quality traits in three French beef cattle breeds. Genet. Sel. Evol. 2016, 48, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warner, R.; Greenwood, P.; Pethick, D.; Ferguson, D. Genetic and environmental effects on meat quality. Meat Sci. 2010, 86, 171–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, K.; Shoji, N.; Honda, T.; Oyama, K. Genetic relationships between meat quality traits and fatty acid composition in Japanese black cattle. Anim. Sci. J. 2016, 88, 11–18. [Google Scholar] [CrossRef]
- Cell Farm Food Tech. Available online: https://cellfarmfoodtech.com/ (accessed on 11 December 2020).
- Eat JUST, Inc. JUST Makes History with Toriyama Wagyu Beef. Available online: https://medium.com/eatjust/a-new-tradition-721e5039de4 (accessed on 11 December 2020).
- Bubner, P.; YC Alum Orbillion Bio Raises an Oversubscribed $5 Million Seed Round, Demonstrates Intense Demand for Lab-Grown Meat that Puts Taste First. Orbillion Bio. 2021. Available online: https://www.orbillion.com/news/release-yc-alum-orbillion-bio-raises-an-oversubscribed-5-million-seed-round-demonstrates-intense-demand-for-lab-grown-meat-that-puts-taste-first (accessed on 24 October 2021).
- Sghaier, C.; Picard, B.; Faulconnier, Y.; Micol, D.; Brun, J.-P.; Reichstadt, M.; Jurie, C.; Durand, D.; Renand, G.; Journaux, L.; et al. A Data Warehouse of Muscle Characteristics and Beef Quality in France and A Demonstration of Potential Applications. Ital. J. Anim. Sci. 2013, 12, e41. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAOSTAT Statistical Database: New Food Balances; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018; Available online: http://www.fao.org/faostat/en/#data/FBS (accessed on 14 September 2021).


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soice, E.; Johnston, J. Immortalizing Cells for Human Consumption. Int. J. Mol. Sci. 2021, 22, 11660. https://doi.org/10.3390/ijms222111660
Soice E, Johnston J. Immortalizing Cells for Human Consumption. International Journal of Molecular Sciences. 2021; 22(21):11660. https://doi.org/10.3390/ijms222111660
Chicago/Turabian StyleSoice, Emily, and Jeremiah Johnston. 2021. "Immortalizing Cells for Human Consumption" International Journal of Molecular Sciences 22, no. 21: 11660. https://doi.org/10.3390/ijms222111660
APA StyleSoice, E., & Johnston, J. (2021). Immortalizing Cells for Human Consumption. International Journal of Molecular Sciences, 22(21), 11660. https://doi.org/10.3390/ijms222111660