
The Role of BMP Signaling in Female Reproductive System Development and Function



Abstract
:1. Introduction
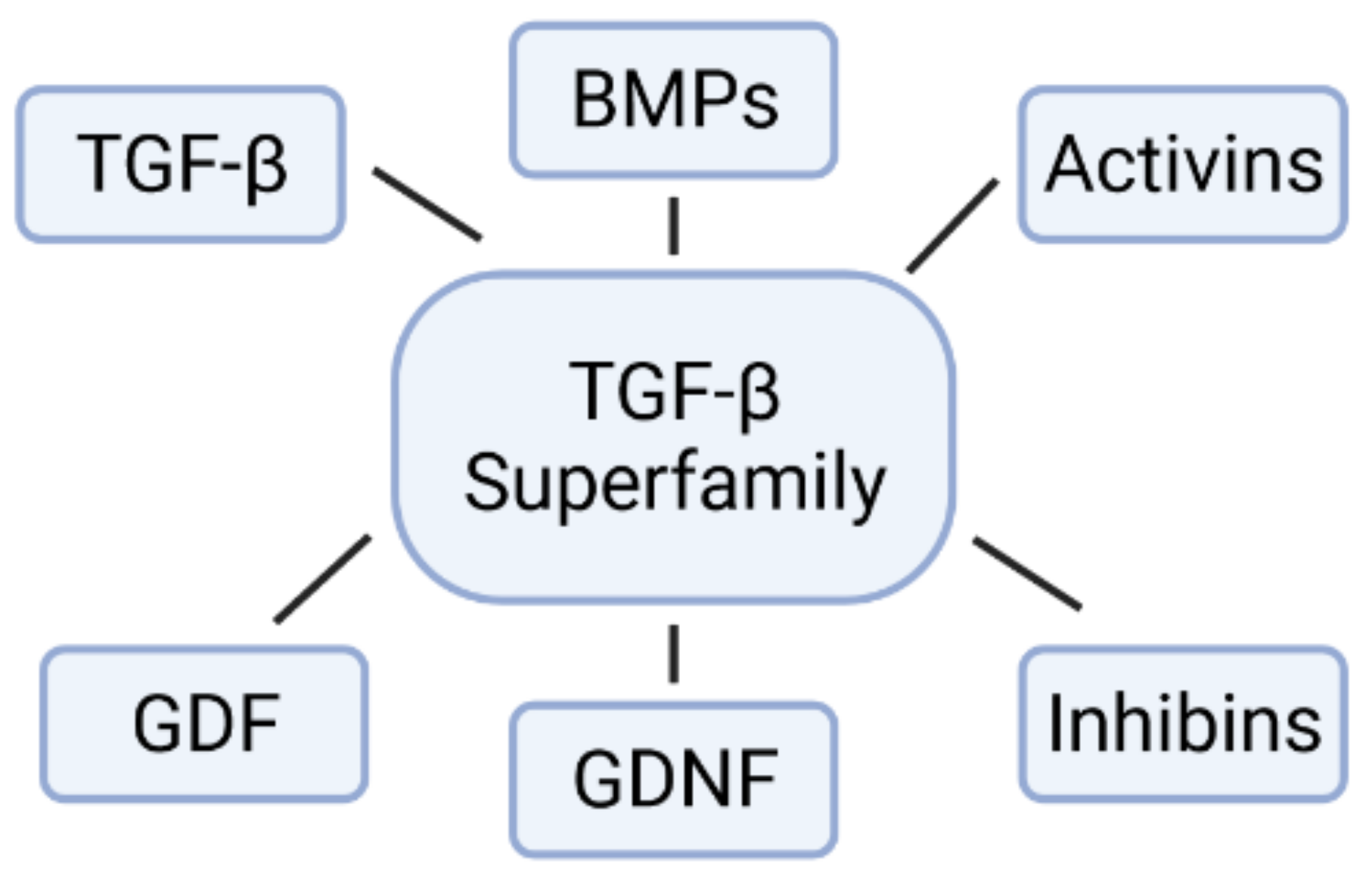
2. The Family of Bone Morphogenetic Proteins (BMPs)
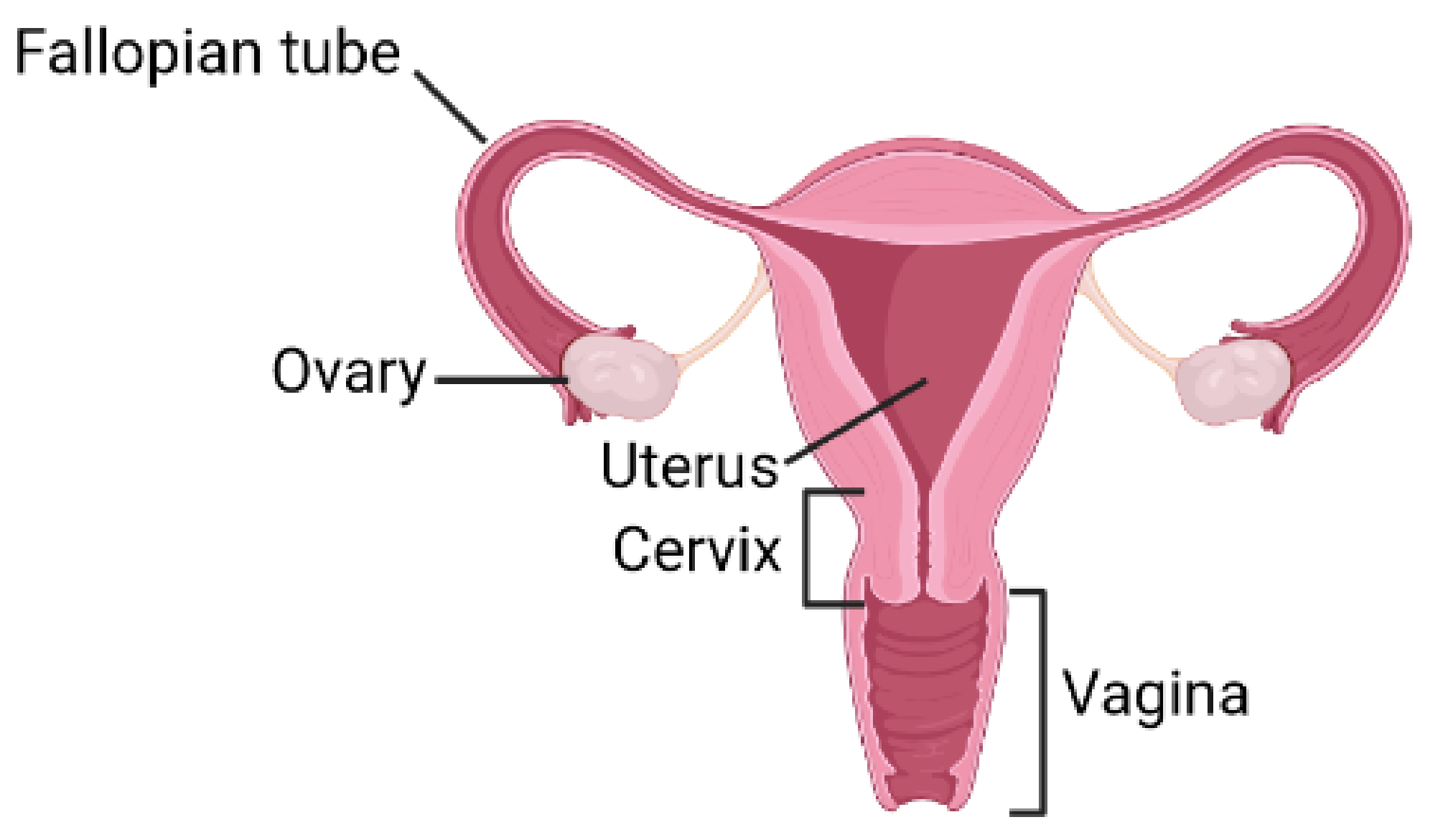
3. BMPs Signaling in Female Reproductive System
3.1. BMPs in the Ovarian Function and Follicular Development
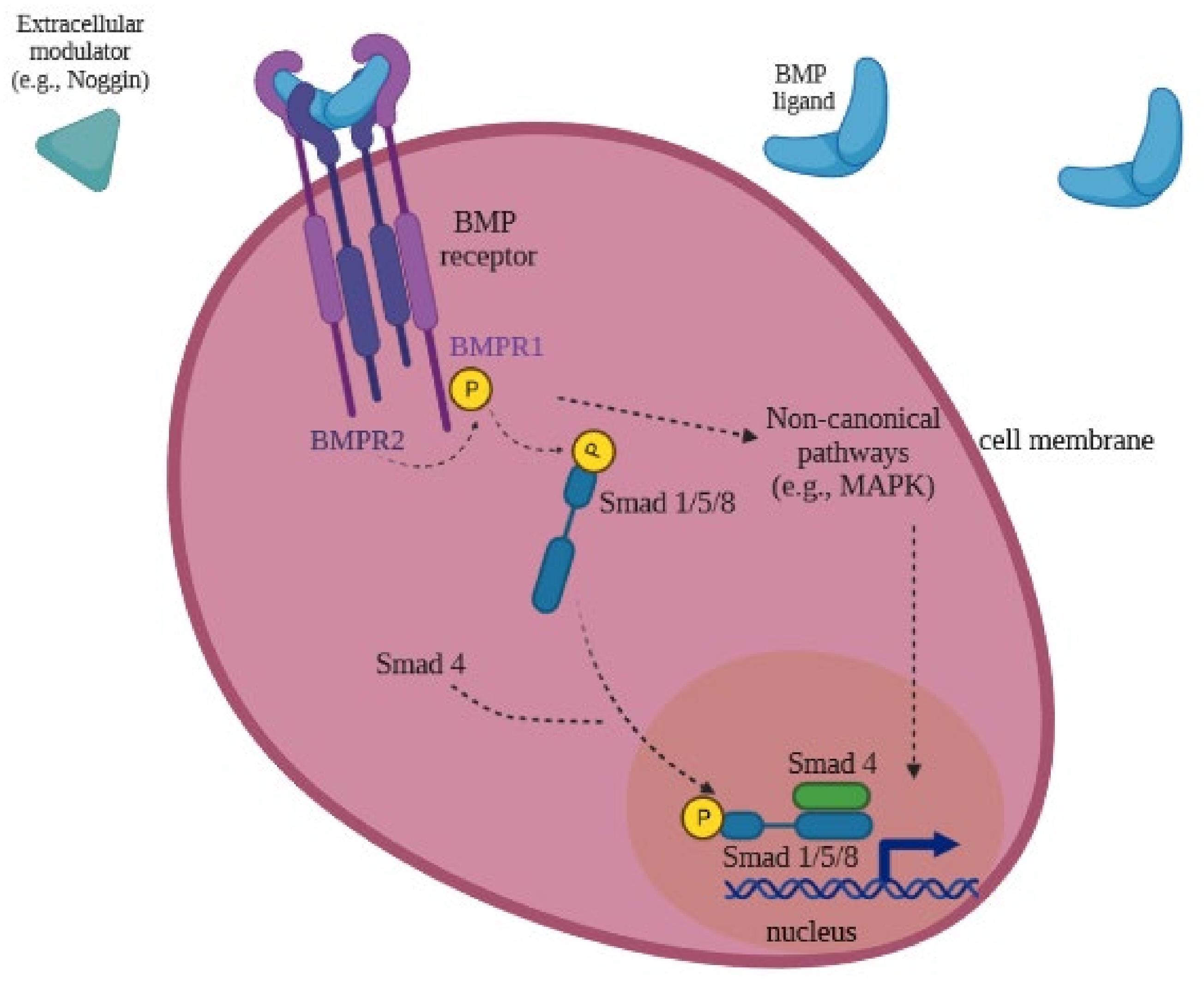
3.2. BMP Receptors Signaling in the Female Reproductive Function
3.3. BMPs Expression in Reproductive Tissue
3.4. BMPs in the Embryonic Development of the Female Reproductive Tract
3.5. BMPs in the Differentiation of hPSCs into 2D Tissues and 3D Organoids
4. Concluding Remarks and Future Directions
Funding
Conflicts of Interest
References
- Acton, Q.A. Issues in Biological and Life Sciences Research; Scholarly Media LLC: Atlanta, GA, USA, 2013; Available online: http://public.eblib.com/choice/PublicFullRecord.aspx?p=6536679 (accessed on 23 October 2021).
- Ali, I.H.A.; Brazil, D.P. Bone morphogenetic proteins and their antagonists: Current and emerging clinical uses: Targeting BMPs in human disease. Br. J. Pharmacol. 2014, 171, 3620–3632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aluganti Narasimhulu, C.; Singla, D.K. The Role of Bone Morphogenetic Protein 7 (BMP-7) in Inflammation in Heart Diseases. Cells 2020, 9, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alzamil, L.; Nikolakopoulou, K.; Turco, M.Y. Organoid systems to study the human female reproductive tract and pregnancy. Cell Death Differ. 2021, 28, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Atsuta, Y.; Takahashi, Y. Early formation of the Müllerian duct is regulated by sequential actions of BMP/Pax2- and FGF/Lim1 signaling. Development 2016, 143, 3549–3559. [Google Scholar] [CrossRef] [Green Version]
- Beppu, H.; Kawabata, M.; Hamamoto, T.; Chytil, A.; Minowa, O.; Noda, T.; Miyazono, K. BMP Type II Receptor Is Required for Gastrulation and Early Development of Mouse Embryos. Dev. Biol. 2000, 221, 249–258. [Google Scholar] [CrossRef] [Green Version]
- Bhattacherjee, A.; Rumi, M.A.K.; Staecker, H.; Smith, P.G. Bone Morphogenetic Protein 4 Mediates Estrogen-Regulated Sensory Axon Plasticity in the Adult Female Reproductive Tract. J. Neurosci. 2013, 33, 1050–1061. [Google Scholar] [CrossRef] [Green Version]
- Bier, E.; De Robertis, E.M. BMP gradients: A paradigm for morphogen-mediated developmental patterning. Science 2015, 348, aaa5838. [Google Scholar] [CrossRef] [Green Version]
- Blitz, I.L.; Cho, K.W.Y.; Chang, C. Twisted gastrulation loss-of-function analyses support its role as a BMP inhibitor during early Xenopus embryogenesis. Development 2003, 130, 4975–4988. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, A.; Behringer, R.R. Developmental genetics of the female reproductive tract in mammals. Nat. Rev. Genet. 2003, 4, 969–980. [Google Scholar] [CrossRef]
- Santana Gonzalez, L.; Rota, I.A.; Artibani, M.; Morotti, M.; Hu, Z.; Wietek, N.; Alsaadi, A.; Albukhari, A.; Sauka-Spengler, T.; Ahmed, A.A. Mechanistic Drivers of Müllerian Duct Development and Differentiation Into the Oviduct. Front. Cell Dev. Biol. 2021, 9, 605301. [Google Scholar] [CrossRef]
- Mullen, R.D.; Behringer, R.R. Molecular genetics of Müllerian duct formation, regression and differentiation. Sex. Dev. 2014, 8, 281–296. [Google Scholar] [CrossRef] [Green Version]
- Bragdon, B.; Moseychuk, O.; Saldanha, S.; King, D.; Julian, J.; Nohe, A. Bone Morphogenetic Proteins: A critical review. Cell. Signal. 2011, 23, 609–620. [Google Scholar] [CrossRef]
- Cai, Y. Revisiting old vaginal topics: Conversion of the Mllerian vagina and origin of the «sinus» vagina. Int. J. Dev. Biol. 2009, 53, 925–934. [Google Scholar] [CrossRef] [Green Version]
- Carreira, A.C.O.; Zambuzzi, W.F.; Rossi, M.C.; Filho, R.A.; Sogayar, M.C.; Granjeiro, J.M. Bone Morphogenetic Proteins. In Vitamins & Hormones; Elsevier: Amsterdam, The Netherlands, 2015; pp. 293–322. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0083672915000370 (accessed on 23 October 2021).
- Chan, S.W.; Rizwan, M.; Yim, E.K.F. Emerging Methods for Enhancing Pluripotent Stem Cell Expansion. Front. Cell Dev. Biol. 2020, 8, 70. [Google Scholar] [CrossRef]
- Settle, S.; Marker, P.; Gurley, K.; Sinha, A.; Thacker, A.; Wang, Y.; Higgins, K.; Cunha, G.; Kingsley, D.M. The BMP Family Member Gdf7 Is Required for Seminal Vesicle Growth, Branching Morphogenesis, and Cytodifferentiation. Dev. Biol. 2001, 234, 138–150. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Zhao, M.; Mundy, G.R. Bone Morphogenetic Proteins. Growth Factors 2004, 22, 233–241. [Google Scholar] [CrossRef]
- Chen, M.; Hsu, I.; Wolfe, A.; Radovick, S.; Huang, K.; Yu, S.; Chang, C.; Messing, E.M.; Yeh, S. Defects of Prostate Development and Reproductive System in the Estrogen Receptor-α Null Male Mice. Endocrinology 2009, 150, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Greenfeld, H.; Lin, J.; Mullins, M.C. The BMP signaling gradient is interpreted through concentration thresholds in dorsal–ventral axial patterning. PLoS Biol. 2021, 19, e3001059. [Google Scholar] [CrossRef]
- Cunha, G.R.; Robboy, S.J.; Kurita, T.; Isaacson, D.; Shen, J.; Cao, M.; Baskin, L.S. Development of the human female reproductive tract. Differentiation 2018, 103, 46–65. [Google Scholar] [CrossRef] [Green Version]
- De Vasconcelos da Cunha, E.; de Souza, G.B.; de Sousa Passos, J.R.; Silva, A.W.B.; Dau, A.M.; Saraiva, M.V.A.; Lobo, R.N.B.; Silva, J.R.V. Effects of bone morphogenetic protein 4 (BMP4) on in vitro development and survival of bovine preantral follicles enclosed in fragments ovarian tissue. Zygote 2017, 25, 256–264. [Google Scholar] [CrossRef]
- Demirhan, O. A homozygous BMPR1B mutation causes a new subtype of acromesomelic chondrodysplasia with genital anomalies. J. Med. Genet. 2005, 42, 314–317. [Google Scholar] [CrossRef]
- Lei, X.; Cui, K.; Cai, X.; Ren, Y.; Liu, Q.; Shi, D. Bone morphogenetic protein 1 is expressed in porcine ovarian follicles and promotes oocyte maturation and early embryonic development. J. Vet. Med. Sci. 2017, 79, 258–266. [Google Scholar] [CrossRef] [Green Version]
- You, J.; Wang, W.; Chang, H.-M.; Yi, Y.; Zhao, H.; Zhu, H.; Sun, Y.; Tang, M.; Wang, C.; Sang, Y.; et al. The BMP2 Signaling Axis Promotes Invasive Differentiation of Human Trophoblasts. Front. Cell Dev. Biol. 2021, 9, 607332. [Google Scholar] [CrossRef]
- Jaatinen, R.; Rosen, V.; Tuuri, T.; Ritvos, O. Identification of ovarian granulosa cells as a novel site of expression for bone morphogenetic protein-3 (BMP-3/osteogenin) and regulation of BMP-3 messenger ribonucleic acids by chorionic gonadotropin in cultured human granulosa-luteal cells. J. Clin. Endocrinol. Metab. 1996, 81, 3877–3882. [Google Scholar]
- Pierre, A.; Pisselet, C.; Dupont, J.; Bontoux, M.; Monget, P. Bone Morphogenetic Protein 5 Expression in the Rat Ovary: Biological Effects on Granulosa Cell Proliferation and Steroidogenesis1. Biol. Reprod. 2005, 73, 1102–1108. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, M.; Zhao, X.; Ren, X.; Shao, S.; Zou, M.; Zhang, L. Bone morphogenetic protein 6 expression in cumulus cells is negatively associated with oocyte maturation. Hum. Fertil. 2019, 24, 290–297. [Google Scholar] [CrossRef]
- Nagao, S.; Iwata, N.; Soejima, Y.; Takiguchi, T.; Aokage, T.; Kozato, Y.; Nakano, Y.; Nada, T.; Hasegawa, T.; Otsuka, F. Interaction of ovarian steroidogenesis and clock gene expression modulated by bone morphogenetic protein-7 in human granulosa cells. Endocr. J. 2019, 66, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.-J.; Wang, Y.-W.; Luo, C.-W. Human bone morphogenetic protein 8A promotes expansion and prevents apoptosis of cumulus cells in vitro. Mol. Cell. Endocrinol. 2021, 522, 111121. [Google Scholar] [CrossRef]
- Hosoya, T.; Otsuka, F.; Nakamura, E.; Terasaka, T.; Inagaki, K.; Tsukamoto-Yamauchi, N.; Hara, T.; Toma, K.; Komatsubara, M.; Makino, H. Regulatory role of BMP-9 in steroidogenesis by rat ovarian granulosa cells. J. Steroid Biochem. Mol. Biol. 2015, 147, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Zheng, W.; Li, L.; Huang, G.; Liu, Y.; Jiang, H.; Zhang, Y.; Tang, C. Loss of BMP-10 is correlated with poor survival in ovarian cancer. Pathol. Res. Pract. 2019, 215, 121–126. [Google Scholar] [CrossRef]
- McPherron, A.C.; Lawler, A.M.; Lee, S.-J. Regulation of anterior/posterior patterning of the axial skeleton by growth/differentiation factor 11. Nat. Genet. 1999, 22, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Ferrarini, E.; De Marco, G.; Orsolini, F.; Gianetti, E.; Benelli, E.; Fruzzetti, F.; Simoncini, T.; Agretti, P.; Tonacchera, M. Characterization of a novel mutation V136L in bone morphogenetic protein 15 identified in a woman affected by POI. J. Ovarian Res. 2021, 14, 85. [Google Scholar] [CrossRef] [PubMed]
- Granjeiro, J.M.; Oliveira, R.C.; Bustos-Valenzuela, J.C.; Sogayar, M.C.; Taga, R. Bone morphogenetic proteins: From structure to clinical use. Braz. J. Med. Biol. Res. 2005, 38, 1463–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grive, K.J.; Freiman, R.N. The developmental origins of the mammalian ovarian reserve. Development 2015, 142, 2554–2563. [Google Scholar] [CrossRef] [Green Version]
- Hardwick, J.C.; Kodach, L.L.; Offerhaus, G.J.; van den Brink, G.R. Bone morphogenetic protein signalling in colorectal cancer. Nat. Rev. Cancer 2008, 8, 806–812. [Google Scholar] [CrossRef]
- Hsu, D.R.; Economides, A.N.; Wang, X.; Eimon, P.M.; Harland, R.M. The Xenopus Dorsalizing Factor Gremlin Identifies a Novel Family of Secreted Proteins that Antagonize BMP Activities. Mol. Cell 1998, 1, 673–683. [Google Scholar] [CrossRef]
- Huang, C.-C.; Orvis, G.D.; Kwan, K.M.; Behringer, R.R. Lhx1 is required in Müllerian duct epithelium for uterine development. Dev. Biol. 2014, 389, 124–136. [Google Scholar] [CrossRef] [Green Version]
- James, R.G.; Schultheiss, T.M. Bmp signaling promotes intermediate mesoderm gene expression in a dose-dependent, cell-autonomous and translation-dependent manner. Dev. Biol. 2005, 288, 113–125. [Google Scholar] [CrossRef] [Green Version]
- Juengel, J.L.; McNatty, K.P. The role of proteins of the transforming growth factor-β superfamily in the intraovarian regulation of follicular development. Hum. Reprod. Update 2005, 11, 144–161. [Google Scholar] [CrossRef]
- Demiray, S.B.; Yilmaz, O.; Goker, E.N.T.; Tavmergen, E.; Calimlioglu, N.; Sezerman, U. Expression of the bone morphogenetic protein-2 (BMP2) in the human cumulus cells as a biomarker of oocytes and embryo quality. J. Hum. Reprod. Sci. 2017, 10, 194. [Google Scholar] [CrossRef]
- Luu, H.H.; Song, W.-X.; Luo, X.; Manning, D.; Luo, J.; Deng, Z.-L.; Sharff, K.A.; Montag, A.G.; Haydon, R.C.; He, T.-C. Distinct roles of bone morphogenetic proteins in osteogenic differentiation of mesenchymal stem cells. J. Orthop. Res. 2007, 25, 665–677. [Google Scholar] [CrossRef]
- Lochab, A.K.; Extavour, C.G. Bone Morphogenetic Protein (BMP) signaling in animal reproductive system development and function. Dev. Biol. 2017, 427, 258–269. [Google Scholar] [CrossRef]
- Massagué, J. TGF-β signal transduction. Annu. Rev. Biochem. 1998, 67, 753–791. [Google Scholar] [CrossRef]
- Bai, L.; Chang, H.; Zhang, L.; Zhu, Y.; Leung, P.C.K. BMP2 increases the production of BDNF through the upregulation of proBDNF and furin expression in human granulosa-lutein cells. FASEB J. 2020, 34, 16129–16143. [Google Scholar] [CrossRef]
- Ito, M.; Yoshino, O.; Ono, Y.; Yamaki-Ushijima, A.; Tanaka, T.; Shima, T. Bone morphogenetic protein-2 enhances gonadotropin-independent follicular development via sphingosine kinase 1. Am. J. Reprod. Immunol. 2021, 85, e13374. [Google Scholar] [CrossRef]
- Kobayashi, A.; Kwan, K.-M.; Carroll, T.J.; McMahon, A.P.; Mendelsohn, C.L.; Behringer, R.R. Distinct and sequential tissue-specific activities of the LIM-class homeobox gene Lim1 for tubular morphogenesis during kidney development. Development 2005, 132, 2809–2823. [Google Scholar] [CrossRef] [Green Version]
- Koch, L. BMPR-2 signalling is essential for maintenance of pregnancy. Nat. Rev. Endocrinol. 2013, 9, 380. [Google Scholar] [CrossRef]
- Kumar, S.; Punetha, M.; Jose, B.; Bharati, J.; Khanna, S.; Sonwane, A.; Green, J.A.; Whitworth, K.; Sarkar, M. Modulation of granulosa cell function via CRISPR-Cas fuelled editing of BMPR-IB gene in goats (Capra hircus). Sci. Rep. 2020, 10, 20446. [Google Scholar] [CrossRef]
- Lawson, K.A.; Dunn, N.R.; Roelen, B.A.J.; Zeinstra, L.M.; Davis, A.M.; Wright, C.V.E.; Korving, J.P.W.F.M.; Hogan, B.L.M. Bmp4 is required for the generation of primordial germ cells in the mouse embryo. Genes Dev. 1999, 13, 424–436. [Google Scholar] [CrossRef]
- Libretti, S.; Aeddula, N.R. Embryology, Genitourinary; StatPearls Publishing: Treasure Island, FL, USA, 2021. Available online: http://www.ncbi.nlm.nih.gov/books/NBK559309/ (accessed on 23 October 2021).
- Litwack, G. Bone morphogenic protein. Vitam. Horm. 2015, 99, 13–14. [Google Scholar]
- Dimitriou, R.; Giannoudis, P.V. Discovery and development of BMPs. Injury 2005, 36, S28–S33. [Google Scholar] [CrossRef]
- Pangas, S.A.; Li, X.; Umans, L.; Zwijsen, A.; Huylebroeck, D.; Gutierrez, C.; Wang, D.; Martin, J.F.; Jamin, S.P.; Behringer, R.R.; et al. Conditional Deletion of Smad1 and Smad5 in Somatic Cells of Male and Female Gonads Leads to Metastatic Tumor Development in Mice. Mol. Cell. Biol. 2008, 28, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Katagiri, T.; Watabe, T. Bone Morphogenetic Proteins. Cold Spring Harb. Perspect. Biol. 2016, 8, a021899. [Google Scholar] [CrossRef] [Green Version]
- Gomes, H.A.N.; Campos, L.B.; Praxedes, É.C.G.; Oliveira, M.F.; Pereira, A.F.; Silva, A.R.; Saraiva, M.V.A. BMP-15 activity on in vitro development of collared peccary (Pecari tajacu Linnaeus, 1758) preantral follicles. Reprod. Domest. Anim. 2020, 55, 958–964. [Google Scholar] [CrossRef]
- Galloway, S.M.; McNatty, K.P.; Cambridge, L.M.; Laitinen, M.P.E.; Juengel, J.L.; Jokiranta, T.S.; McLaren, R.J.; Luiro, K.; Dodds, K.G.; Montgomery, G.W.; et al. Mutations in an oocyte-derived growth factor gene (BMP15) cause increased ovulation rate and infertility in a dosage-sensitive manner. Nat. Genet. 2000, 25, 279–283. [Google Scholar] [CrossRef]
- Yan, C.; Wang, P.; DeMayo, J.; DeMayo, F.J.; Elvin, J.A.; Carino, C.; Prasad, S.V.; Skinner, S.S.; Dunbar, B.S.; Dube, J.L.; et al. Synergistic Roles of Bone Morphogenetic Protein 15 and Growth Differentiation Factor 9 in Ovarian Function. Mol. Endocrinol. 2001, 15, 854–866. [Google Scholar] [CrossRef]
- Luo, X.; Chang, H.; Yi, Y.; Leung, P.C.K.; Sun, Y. Bone morphogenetic protein 2 upregulates SERPINE2 expression through noncanonical SMAD2/3 and p38 MAPK signaling pathways in human granulosa-lutein cells. FASEB J. 2021, 35, e21845. [Google Scholar] [CrossRef]
- Renault, L.; Patiño, L.C.; Magnin, F.; Delemer, B.; Young, J.; Laissue, P.; Binart, N.; Beau, I. BMPR1A and BMPR1B Missense Mutations Cause Primary Ovarian Insufficiency. J. Clin. Endocrinol. Metab. 2020, 105, e1449-57. [Google Scholar] [CrossRef]
- Yi, S.E.; LaPolt, P.S.; Yoon, B.S.; Chen, J.Y.-C.; Lu, J.K.H.; Lyons, K.M. The type I BMP receptor BmprIB is essential for female reproductive function. Proc. Natl. Acad. Sci. USA 2001, 98, 7994–7999. [Google Scholar] [CrossRef] [Green Version]
- Souza, C.J.; MacDougall, C.; MacDougall, C.; Campbell, B.K.; McNeilly, A.S.; Baird, D.T. The Booroola (FecB) phenotype is associated with a mutation in the bone morphogenetic receptor type 1 B (BMPR1B) gene. J. Endocrinol. 2001, 169, R1–R6. [Google Scholar] [CrossRef] [Green Version]
- Tanwar, P.S.; McFarlane, J.R. Dynamic expression of bone morphogenetic protein 4 in reproductive organs of female mice. Reproduction 2011, 142, 573–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Ding, Y.; Liu, J.; Liu, Y.; Zhao, X.; Li, G.; Zhang, C.; Li, C.; Wang, Y.; Kalds, P.; et al. Highly efficient generation of sheep with a defined FecBB mutation via adenine base editing. Genet. Sel. Evol. 2020, 52, 35. [Google Scholar] [CrossRef] [PubMed]
- Tsevat, D.G.; Wiesenfeld, H.C.; Parks, C.; Peipert, J.F. Sexually transmitted diseases and infertility. Am. J. Obstet. Gynecol. 2017, 216, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Roy, S.K. Expression of Bone Morphogenetic Protein Receptor (BMPR) during Perinatal Ovary Development and Primordial Follicle Formation in the Hamster: Possible Regulation by FSH. Endocrinology 2009, 150, 1886–1896. [Google Scholar] [CrossRef] [Green Version]
- Urist, M.R. Bone: Formation by Autoinduction. Science 1965, 150, 893–899. [Google Scholar] [CrossRef]
- Vercellini, P.; Viganò, P.; Somigliana, E.; Fedele, L. Endometriosis: Pathogenesis and treatment. Nat. Rev. Endocrinol. 2014, 10, 261–275. [Google Scholar] [CrossRef]
- Nagashima, T.; Li, Q.; Clementi, C.; Lydon, J.P.; DeMayo, F.J.; Matzuk, M.M. BMPR2 is required for postimplantation uterine function and pregnancy maintenance. J. Clin. Investig. 2013, 123, 2539–2550. [Google Scholar] [CrossRef] [Green Version]
- Cha, J.; Sun, X.; Dey, S.K. Mechanisms of implantation: Strategies for successful pregnancy. Nat. Med. 2012, 18, 1754–1767. [Google Scholar] [CrossRef]
- Lee, K.Y.; Jeong, J.-W.; Wang, J.; Ma, L.; Martin, J.F.; Tsai, S.Y.; Lydon, J.P.; DeMayo, F.J. Bmp2 Is Critical for the Murine Uterine Decidual Response. Mol. Cell. Biol. 2007, 27, 5468–5478. [Google Scholar] [CrossRef] [Green Version]
- Monsivais, D.; Clementi, C.; Peng, J.; Fullerton, P.T.; Prunskaite-Hyyryläinen, R.; Vainio, S.J.; Matzuk, M.M. BMP7 Induces Uterine Receptivity and Blastocyst Attachment. Endocrinology 2017, 158, 979–992. [Google Scholar] [CrossRef]
- Clementi, C.; Tripurani, S.K.; Large, M.J.; Edson, M.A.; Creighton, C.J.; Hawkins, S.M.; Kovanci, E.; Kaartinen, V.; Lydon, J.P.; Pangas, S.A.; et al. Activin-Like Kinase 2 Functions in Peri-implantation Uterine Signaling in Mice and Humans. PLoS Genet. 2013, 9, e1003863. [Google Scholar] [CrossRef] [Green Version]
- Masse, J.; Watrin, T.; Laurent, A.; Deschamps, S.; Guerrier, D.; Pellerin, I. The developing female genital tract: From genetics to epigenetics. Int. J. Dev. Biol. 2009, 53, 411–424. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Pardali, E.; Sánchez-Duffhues, G.; ten Dijke, P. BMP signaling in vascular diseases. FEBS Lett. 2012, 586, 1993–2002. [Google Scholar] [CrossRef]
- Laronda, M.M.; Burdette, J.E.; Kim, J.; Woodruff, T.K. Recreating the female reproductive tract in vitro using iPSC technology in a linked microfluidics environment. Stem Cell Res. Ther. 2013, 4 (Suppl. 1), S13. [Google Scholar] [CrossRef] [Green Version]
- Mulsant, P.; Lecerf, F.; Fabre, S.; Schibler, L.; Monget, P.; Lanneluc, I.; Pisselet, C.; Riquet, J.; Monniaux, D.; Callebaut, I.; et al. Mutation in bone morphogenetic protein receptor-IB is associated with increased ovulation rate in Booroola Merino ewes. Proc. Natl. Acad. Sci. USA 2001, 98, 5104–5109. [Google Scholar] [CrossRef] [Green Version]
- Padgett, R.W.; Wozney, J.M.; Gelbart, W.M. Human BMP sequences can confer normal dorsal-ventral patterning in the Drosophila embryo. Proc. Natl. Acad. Sci. USA 1993, 90, 2905–2909. [Google Scholar] [CrossRef] [Green Version]
- Park, E.-S.; Woods, D.C.; Tilly, J.L. Bone morphogenetic protein 4 promotes mammalian oogonial stem cell differentiation via Smad1/5/8 signaling. Fertil. Steril. 2013, 100, 1468–1475. [Google Scholar] [CrossRef] [Green Version]
- Persani, L.; Rossetti, R.; Di Pasquale, E.; Cacciatore, C.; Fabre, S. The fundamental role of bone morphogenetic protein 15 in ovarian function and its involvement in female fertility disorders. Hum. Reprod. Update 2014, 20, 869–883. [Google Scholar] [CrossRef] [Green Version]
- Pesce, M.; Gioia Klinger, F.; De Felici, M. Derivation in culture of primordial germ cells from cells of the mouse epiblast: Phenotypic induction and growth control by Bmp4 signalling. Mech. Dev. 2002, 112, 15–24. [Google Scholar] [CrossRef]
- Richter, A.; Valdimarsdottir, L.; Hrafnkelsdottir, H.E.; Runarsson, J.F.; Omarsdottir, A.R.; Oostwaard, D.W.; Mummery, C.; Valdimarsdottir, G. BMP4 Promotes EMT and Mesodermal Commitment in Human Embryonic Stem Cells via SLUG and MSX2: Mesodermal Commitment in Embryonic Stem Cells. Stem Cells 2014, 32, 636–648. [Google Scholar] [CrossRef]
- Saitou, M.; Miyauchi, H. Gametogenesis from Pluripotent Stem Cells. Cell Stem Cell 2016, 18, 721–735. [Google Scholar] [CrossRef]
- Sanfins, A.; Rodrigues, P.; Albertini, D.F. GDF-9 and BMP-15 direct the follicle symphony. J. Assist. Reprod. Genet. 2018, 35, 1741–1750. [Google Scholar] [CrossRef]
- Heremans, R.; Jan, Z.; Timmerman, D.; Vankelecom, H. Organoids of the Female Reproductive Tract: Innovative Tools to Study Desired to Unwelcome Processes. Front. Cell Dev. Biol. 2021, 9, 661472. [Google Scholar] [CrossRef]
- Sepponen, K.; Lundin, K.; Knuus, K.; Vayrynen, P.; Raivio, T.; Tapanainen, J.S.; Tuuri, T. The Role of Sequential BMP Signaling in Directing Human Embryonic Stem Cells to Bipotential Gonadal Cells. J. Clin. Endocrinol. Metab. 2017, 102, 4303–4314. [Google Scholar] [CrossRef]
- Winnier, G.; Blessing, M.; Labosky, P.A.; Hogan, B.L. Bone morphogenetic protein-4 is required for mesoderm formation and patterning in the mouse. Genes Dev. 1995, 9, 2105–2116.89. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Li, J.; Tan, Z.; Wang, C.; Liu, T.; Chen, L.; Yong, J.; Jiang, W.; Sun, X.; Du, L.; et al. Short-term BMP-4 treatment initiates mesoderm induction in human embryonic stem cells. Blood 2008, 111, 1933–1941.90. [Google Scholar] [CrossRef] [Green Version]
- Yucer, N.; Holzapfel, M.; Jenkins Vogel, T.; Lenaeus, L.; Ornelas, L.; Laury, A.; Sareen, D.; Barrett, R.; Karlan, B.Y.; Svendsen, C.N. Directed Differentiation of Human Induced Pluripotent Stem Cells into Fallopian Tube Epithelium. Sci. Rep. 2017, 7, 10741. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| BMP Members | Function |
|---|---|
| BMP-1 | BMP-1 has a role in ovarian follicular development and early porcine embryo development [24]. |
| BMP-2 | BMP-2 plays a key function in placenta development [25]. |
| BMP-3 | BMP-3 is expressed in human granulosa cells (GCs) [26]. |
| BMP-4 | BMP-4 is an essential morphogen regulating vaginal development and is expressed in the early postnatal period [14]. |
| BMP-5 | BMP-5 is expressed by GCs in the rat ovary and exerts biological effects on proliferation and steroidogenesis of GCs in an autocrine manner [27]. |
| BMP-6 | BMP-6 expression is negatively associated with oocyte maturation in cumulus cells (CCs) [28]. |
| BMP-7 | BMP-7 modulates ovarian steroidogenesis in human GCs [29]. |
| BMP-8 | BMP-8 promotes expansion a prevents apoptosis of CCs in vitro [30]. |
| BMP-9 | BMP-9 is expressed in the ovary regulating steroidogenesis by GCs [31]. |
| BMP-10 | Low BMP-10 expression is correlated with poor progression of ovarian cancer [32]. |
| BMP-11 | BMP-11 knockout mice die shortly after birth [33]. |
| BMP-12, BMP-13, BMP-14 | No functions have been described in FRS. |
| BMP-15 | A novel BMP-15 mutation has been shown to provide additional understanding of the molecular basis of premature ovarian failure (POF) disease [34]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magro-Lopez, E.; Muñoz-Fernández, M.Á. The Role of BMP Signaling in Female Reproductive System Development and Function. Int. J. Mol. Sci. 2021, 22, 11927. https://doi.org/10.3390/ijms222111927
Magro-Lopez E, Muñoz-Fernández MÁ. The Role of BMP Signaling in Female Reproductive System Development and Function. International Journal of Molecular Sciences. 2021; 22(21):11927. https://doi.org/10.3390/ijms222111927
Chicago/Turabian StyleMagro-Lopez, Esmeralda, and María Ángeles Muñoz-Fernández. 2021. "The Role of BMP Signaling in Female Reproductive System Development and Function" International Journal of Molecular Sciences 22, no. 21: 11927. https://doi.org/10.3390/ijms222111927
APA StyleMagro-Lopez, E., & Muñoz-Fernández, M. Á. (2021). The Role of BMP Signaling in Female Reproductive System Development and Function. International Journal of Molecular Sciences, 22(21), 11927. https://doi.org/10.3390/ijms222111927