
Using Knock-Out Mutants to Investigate the Adhesion of Staphylococcus aureus to Abiotic Surfaces

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
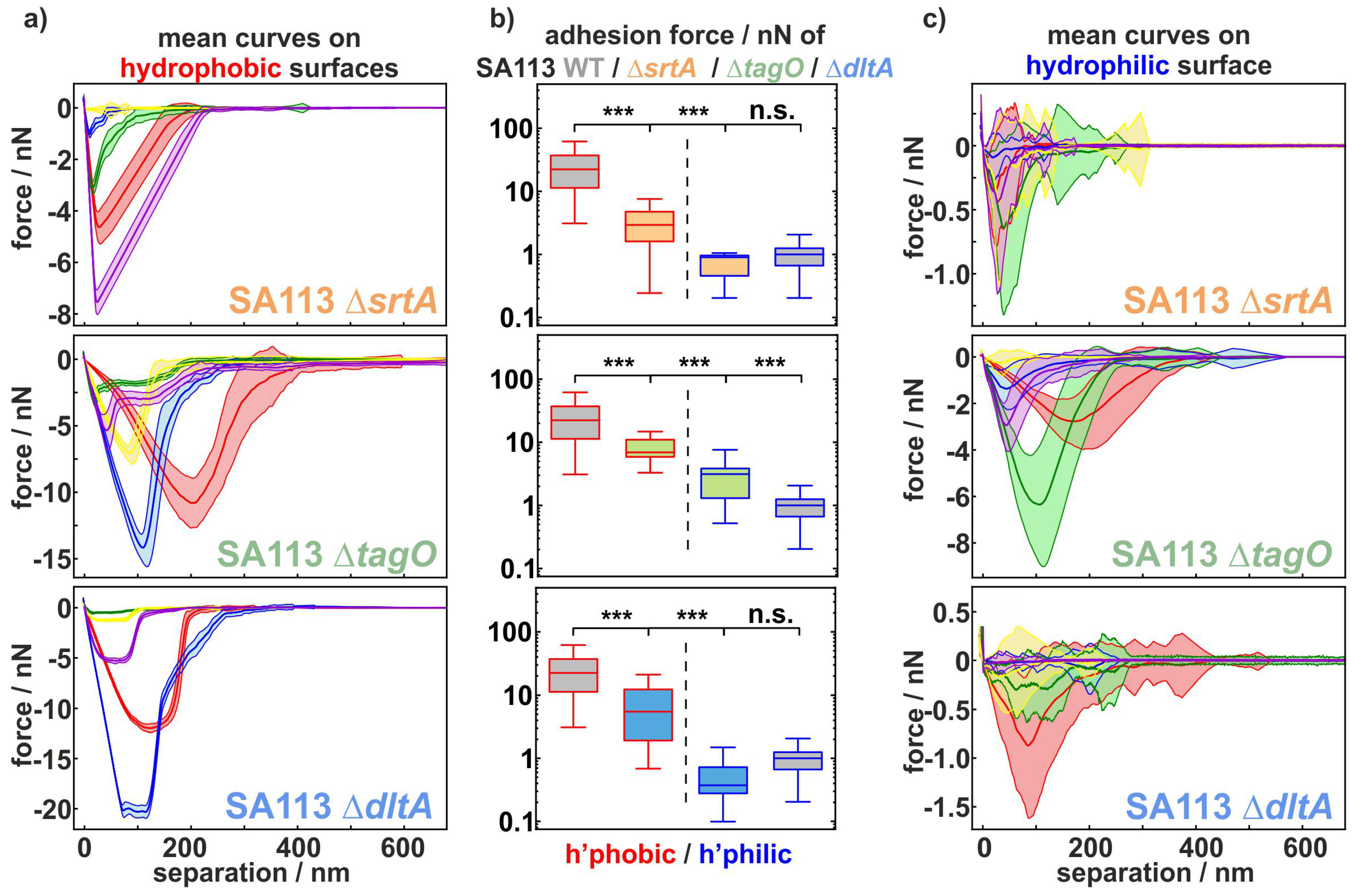
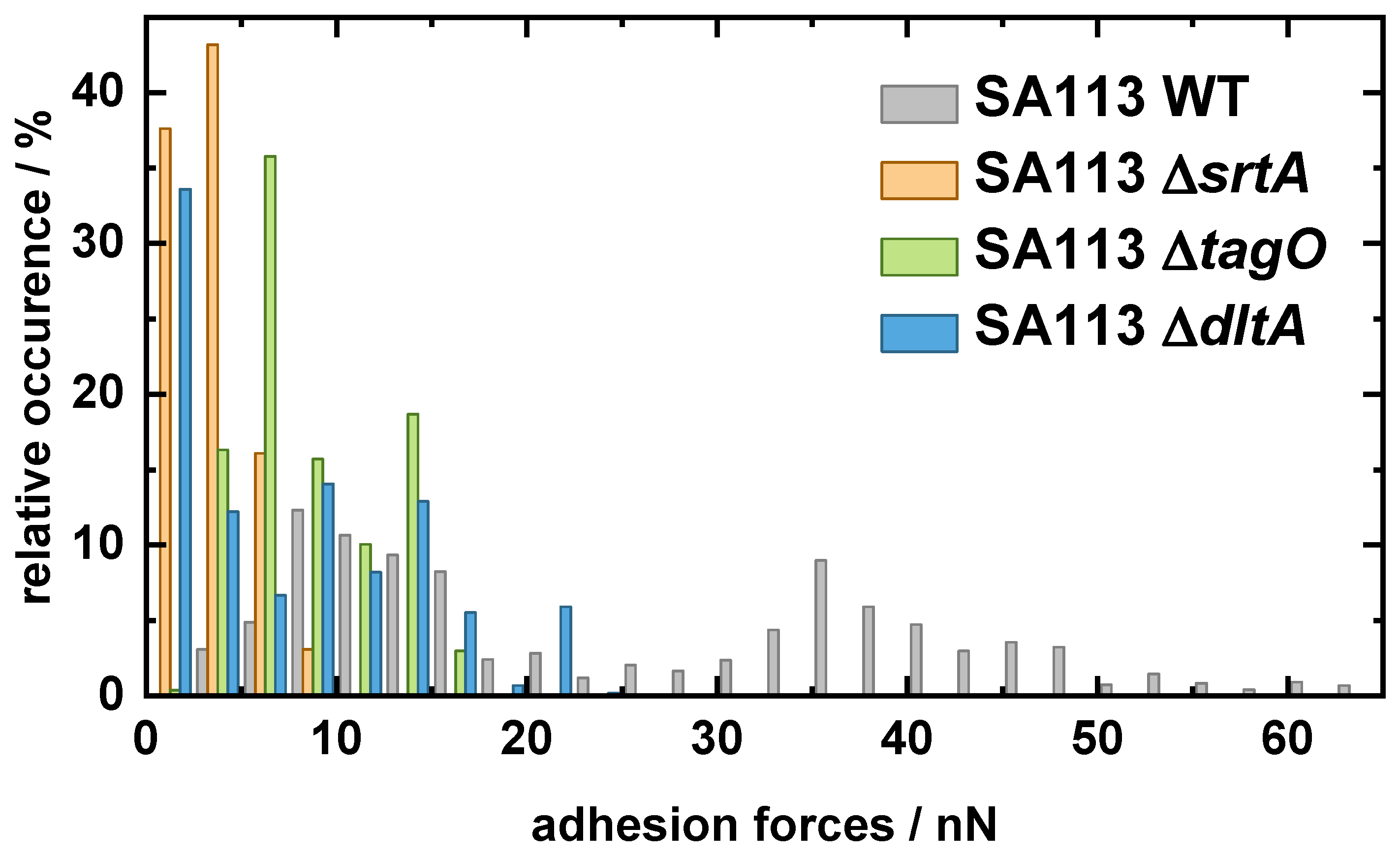
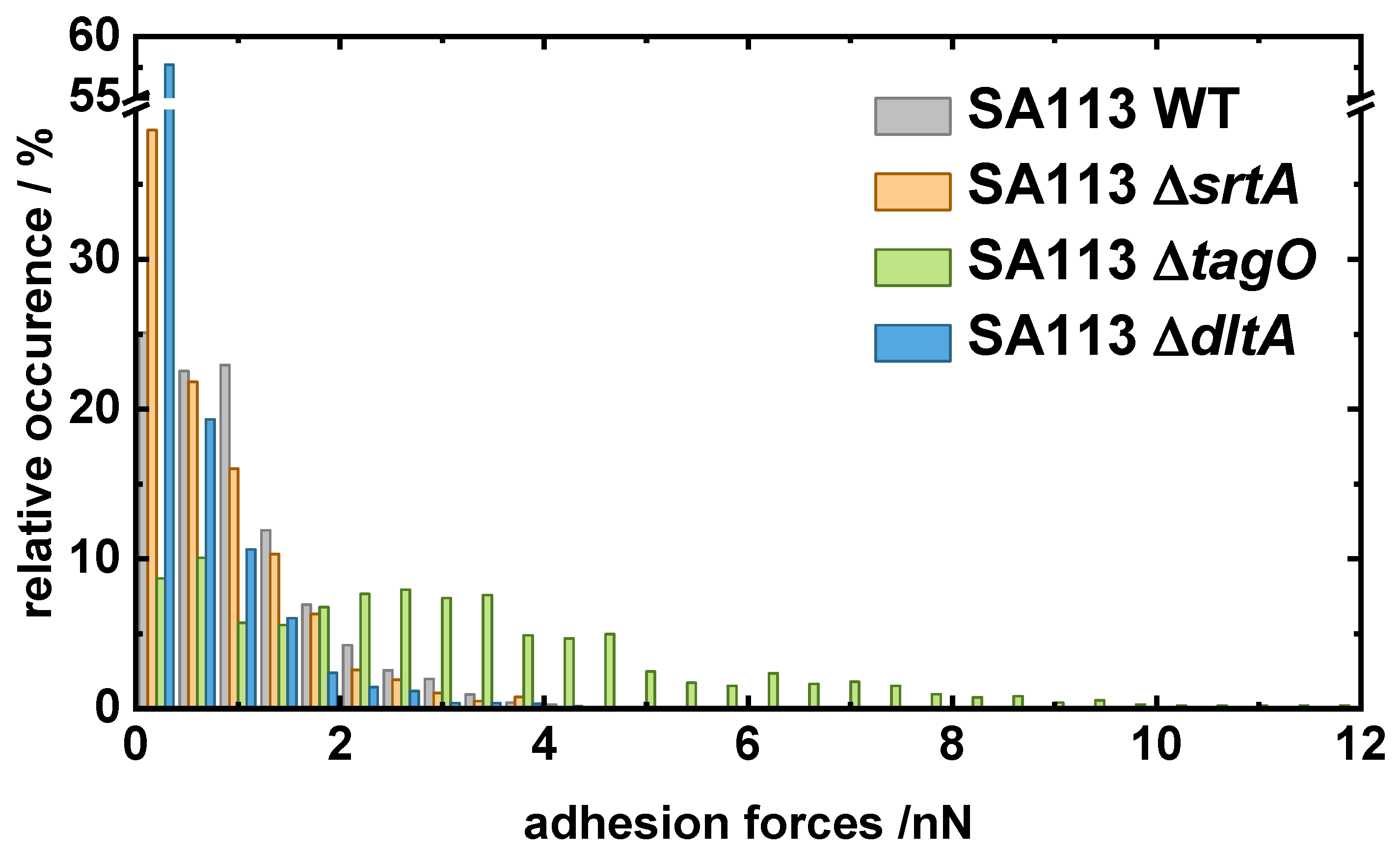
2.1. Comparing the Adhesion of Knock-Out Mutants to Hydrophobic and Hydrophilic Surfaces
- SA113 tagO: lacking the gene tagO encoding a glycosyltransferase that catalyzes the first committed step of wall teichoic acid (WTA) synthesis (but having lipoteichoic acids) [31].
- SA113 dltA: lacking the gene dltA encoding the D-alanine-D-alanyl carrier protein ligase catalyzing the first step in the D-alanylation of lipoteichoic acids (LTAs). As a consequence, the wall and lipoteichoic acids of this mutant lack D-alanine, leading to an increased negative surface charge of the cell wall [39].
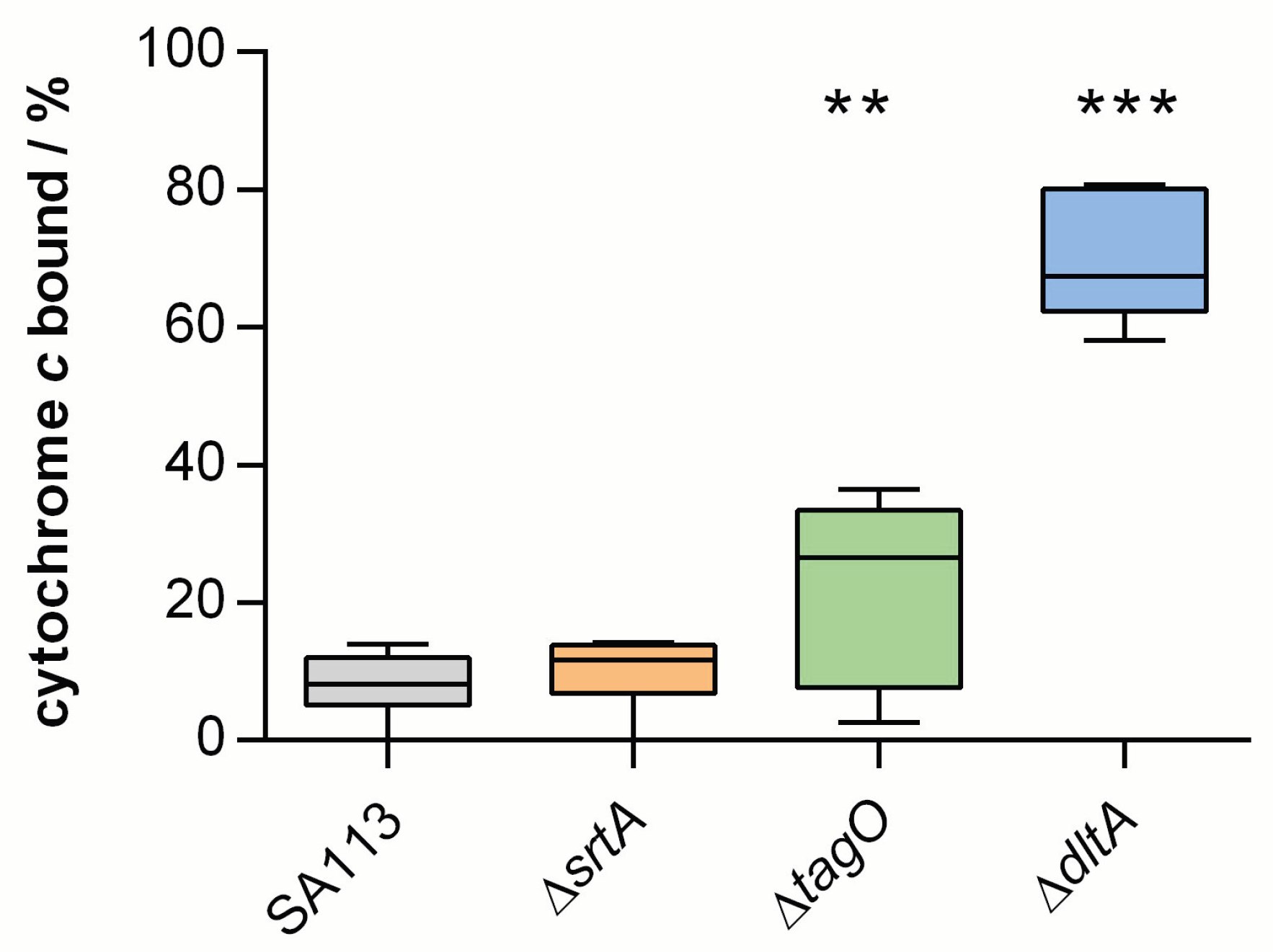
2.2. Comparing the Surface Charge of Knock-Out Mutants to the Parental Strain
2.3. Statistical Analysis of Mutants Adhesion to Hydrophobic Substrates
2.4. Statistical Analysis of Mutants Adhesion to Hydrophilic Substrates
3. Materials and Methods
3.1. Substrate Preparation
3.2. Bacterial Strains and Growth Conditions
3.3. Cytochrome c Binding Assay
3.4. Single-Cell Force Spectroscopy
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AFM | Atomic force microscopy |
| SCFS | Single-cell force spectroscopy |
| WTA | Wall teichoic acid |
| LTA | Lipoteichoic acid |
Appendix A. Comparison of All Cells
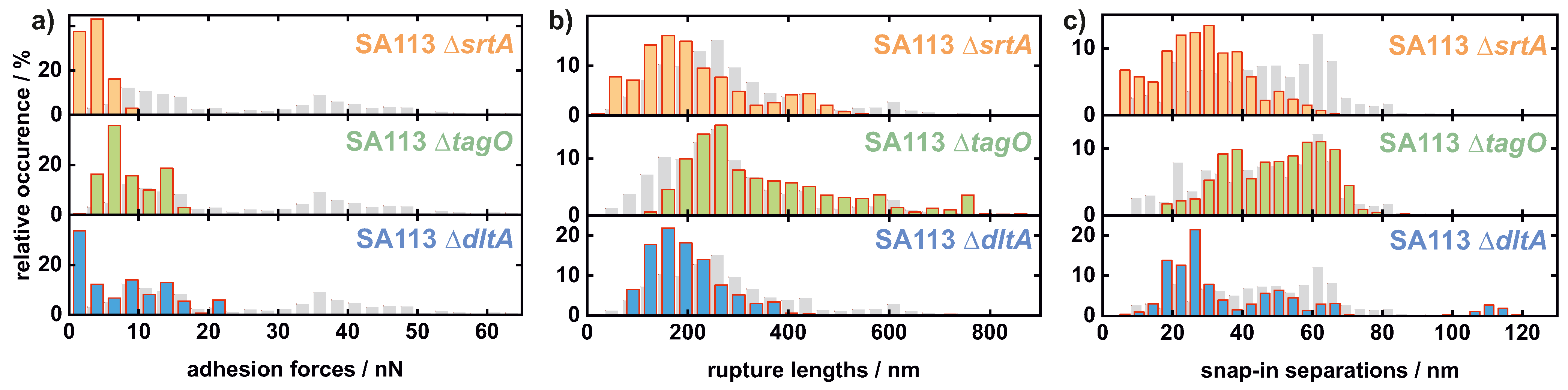
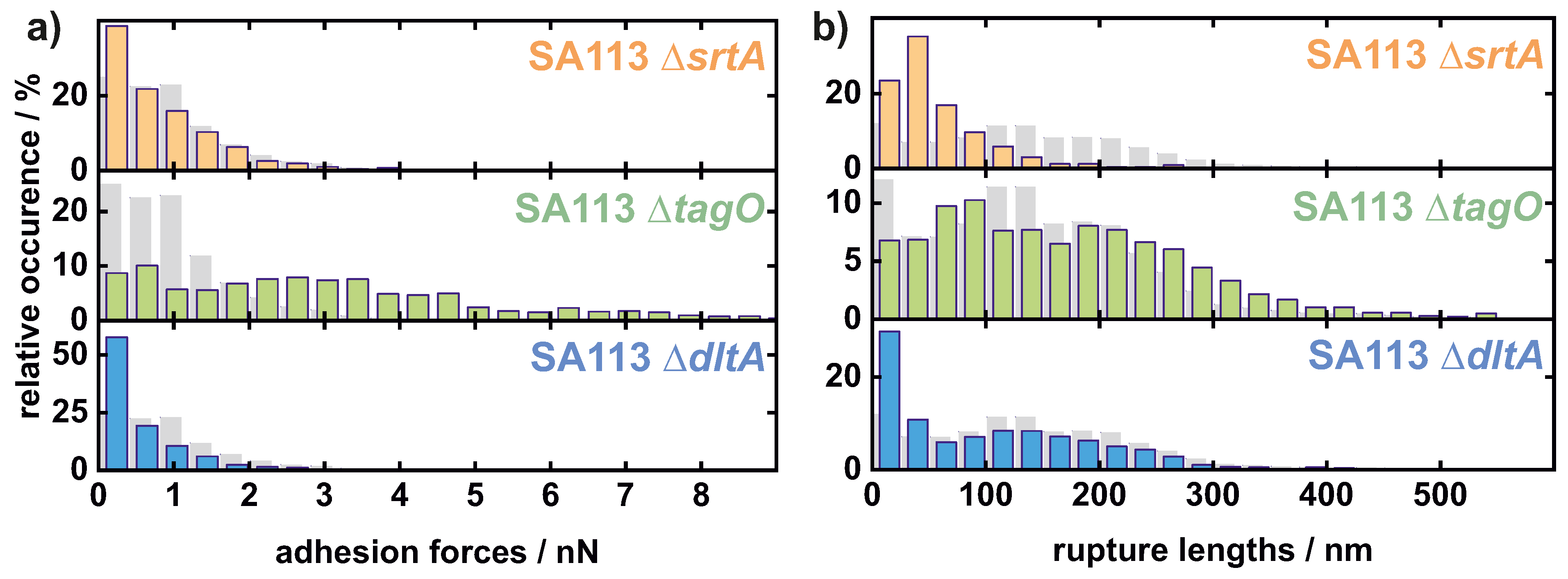
Appendix A.1. Comparison of All Cells Measured on Hydrophobic Surfaces

Appendix A.2. Comparison of All Cells Measured on Hydrophilic Surfaces

References
- Lowy, F.D. Staphylococcus aureus Infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the Natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Yongsunthon, R.; Fowler, V.G.; Lower, B.H.; Vellano, F.P.; Alexander, E.; Reller, L.B.; Corey, G.R.; Lower, S.K. Correlation between Fundamental Binding Forces and Clinical Prognosis of Staphylococcus aureus Infections of Medical Implants. Langmuir 2007, 23, 2289–2292. [Google Scholar] [CrossRef] [PubMed]
- Heilmann, C. Adhesion Mechanisms of Staphylococci. In Advances in Experimental Medicine and Biology; Springer: Dordrecht, The Netherlands, 2011; pp. 105–123. [Google Scholar] [CrossRef]
- Thewes, N.; Thewes, A.; Loskill, P.; Peisker, H.; Bischoff, M.; Herrmann, M.; Santen, L.; Jacobs, K. Stochastic binding of Staphylococcus aureus to hydrophobic surfaces. Soft Matter. 2015, 11, 8913–8919. [Google Scholar] [CrossRef]
- Gotz, F.; Peters, G. Colonization of Medical Devices by Coagulase-Negative Staphylococci. In Infections Associated with Indwelling Medical Devices; ASM Press: Washington, DC, USA, 2014; pp. 55–88. [Google Scholar] [CrossRef]
- Christensen, G.D.; Baldassarri, L.; Simpson, W.A. Methods for studying microbial colonization of plastics. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1995; pp. 477–500. [Google Scholar] [CrossRef]
- Chamis, A.L.; Peterson, G.E.; Cabell, C.H.; Corey, G.R.; Sorrentino, R.A.; Greenfield, R.A.; Ryan, T.; Reller, L.B.; Fowler, V.G. Staphylococcus aureus Bacteremia in PatientsWith Permanent Pacemakers or Implantable Cardioverter-Defibrillators. Circulation 2001, 104, 1029–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mylonakis, E.; Calderwood, S.B. Infective Endocarditis in Adults. N. Engl. J. Med. 2001, 345, 1318–1330. [Google Scholar] [CrossRef]
- Olsen, M.A.; Mayfield, J.; Lauryssen, C.; Polish, L.B.; Jones, M.; Vest, J.; Fraser, V.J. Risk factors for surgical site infection in spinal surgery. J. Neurosurg. Spine 2003, 98, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Darouiche, R.O. Treatment of Infections Associated with Surgical Implants. N. Engl. J. Med. 2004, 350, 1422–1429. [Google Scholar] [CrossRef]
- Arciola, C.R.; Campoccia, D.; Speziale, P.; Montanaro, L.; Costerton, J.W. Biofilm formation in Staphylococcus implant infections. A review of molecular mechanisms and implications for biofilm-resistant materials. Biomaterials 2012, 33, 5967–5982. [Google Scholar] [CrossRef]
- Razavi, M.; Shepard, D.S.; Suaya, J.A.; Stason, W.B. Postoperative Staphylococcus aureus Infections in Medicare Beneficiaries. PLoS ONE 2014, 9, e110133. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Elimelech, M. Bioinspired Single Bacterial Cell Force Spectroscopy. Langmuir 2009, 25, 9656–9659. [Google Scholar] [CrossRef]
- Thewes, N.; Loskill, P.; Jung, P.; Peisker, H.; Bischoff, M.; Herrmann, M.; Jacobs, K. Hydrophobic interaction governs unspecific adhesion of staphylococci: A single cell force spectroscopy study. Beilstein J. Nanotechnol. 2014, 5, 1501–1512. [Google Scholar] [CrossRef] [Green Version]
- Dufrene, Y.F. Sticky microbes: Forces in microbial cell adhesion. Trends Microbiol. 2015, 23, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Sullan, R.M.A.; Li, J.K.; Crowley, P.J.; Brady, L.J.; Dufrene, Y.F. Binding Forces of Streptococcus mutans P1 Adhesin. ACS Nano 2015, 9, 1448–1460. [Google Scholar] [CrossRef] [Green Version]
- Aguayo, S.; Donos, N.; Spratt, D.; Bozec, L. Single-bacterium nanomechanics in biomedicine: Unravelling the dynamics of bacterial cells. Nanotechnology 2015, 26, 062001. [Google Scholar] [CrossRef] [PubMed]
- Thewes, N.; Loskill, P.; Spengler, C.; Humbert, S.; Bischoff, M.; Jacobs, K. A detailed guideline for the fabrication of single bacterial probes used for atomic force spectroscopy. Eur. Phys. J. E 2015, 38, 140. [Google Scholar] [CrossRef]
- Mitchell, G.; Lamontagne, C.A.; Brouillette, E.; Grondin, G.; Talbot, B.G.; Grandbois, M.; Malouin, F. Staphylococcus aureus SigB activity promotes a strong fibronectin-bacterium interaction which may sustain host tissue colonization by small-colony variants isolated from cystic fibrosis patients. Mol. Microbiol. 2008, 70, 1540–1555. [Google Scholar] [CrossRef]
- Ovchinnikova, E.S.; Krom, B.P.; Busscher, H.J.; van der Mei, H.C. Evaluation of adhesion forces of Staphylococcus aureus along the length of Candida albicanshyphae. BMC Microbiol. 2012, 12, 281. [Google Scholar] [CrossRef] [Green Version]
- Herman-Bausier, P.; El-Kirat-Chatel, S.; Foster, T.J.; Geoghegan, J.A.; Dufrene, Y.F. Staphylococcus aureus Fibronectin-Binding Protein A Mediates Cell-Cell Adhesion through Low-Affinity Homophilic Bonds. mBio 2015, 6, e00413-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanzieleghem, T.; Herman-Bausier, P.; Dufrene, Y.F.; Mahillon, J. Staphylococcus epidermidis Affinity for Fibrinogen-Coated Surfaces Correlates with the Abundance of the SdrG Adhesin on the Cell Surface. Langmuir 2015, 31, 4713–4721. [Google Scholar] [CrossRef] [PubMed]
- Spengler, C.; Thewes, N.; Nolle, F.; Faidt, T.; Umanskaya, N.; Hannig, M.; Bischoff, M.; Jacobs, K. Enhanced adhesion of Streptococcus mutansto hydroxyapatite after exposure to saliva. J. Mol. Recognit. 2017, 30, e2615. [Google Scholar] [CrossRef] [PubMed]
- Viljoen, A.; Mignolet, J.; Viela, F.; Mathelie-Guinlet, M.; Dufrene, Y.F. How Microbes Use Force To Control Adhesion. J. Bacteriol. 2020, 202, e00125–20. [Google Scholar] [CrossRef] [PubMed]
- Sjollema, J.; van der Mei, H.C.; Hall, C.L.; Peterson, B.W.; de Vries, J.; Song, L.; Jong, E.D.d.; Busscher, H.J.; Swartjes, J.J.T.M. Detachment and successive re-attachment of multiple, reversibly-binding tethers result in irreversible bacterial adhesion to surfaces. Sci. Rep. 2017, 7, 4369. [Google Scholar] [CrossRef] [Green Version]
- Maikranz, E.; Spengler, C.; Thewes, N.; Thewes, A.; Nolle, F.; Jung, P.; Bischoff, M.; Santen, L.; Jacobs, K. Different binding mechanisms of Staphylococcus aureus to hydrophobic and hydrophilic surfaces. Nanoscale 2020, 12, 19267–19275. [Google Scholar] [CrossRef]
- Spengler, C.; Thewes, N.; Jung, P.; Bischoff, M.; Jacobs, K. Determination of the nano-scaled contact area of staphylococcal cells. Nanoscale 2017, 9, 10084–10093. [Google Scholar] [CrossRef]
- Spengler, C.; Glatz, B.A.; Maikranz, E.; Bischoff, M.; Klatt, M.A.; Santen, L.; Fery, A.; Jacobs, K. The adhesion capability of S. aureus cells is heterogeneously distributed over the cell envelope. bioRxiv 2021. [Google Scholar] [CrossRef]
- Weidenmaier, C.; Peschel, A.; Xiong, Y.Q.; Kristian, S.A.; Dietz, K.; Yeaman, M.R.; Bayer, A.S. Lack of Wall Teichoic Acids in Staphylococcus aureus Leads to Reduced Interactions with Endothelial Cells and to Attenuated Virulence in a Rabbit Model of Endocarditis. J. Infect. Dis. 2005, 191, 1771–1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira, L.M.F.; Steindorff, M.; Darisipudi, M.N.; Mrochen, D.M.; Trube, P.; Broker, B.M.; Bronstrup, M.; Tegge, W.; Holtfreter, S. Discovery of Staphylococcus aureus Adhesion Inhibitors by Automated Imaging and Their Characterization in a Mouse Model of Persistent Nasal Colonization. Microorganisms 2021, 9, 631. [Google Scholar] [CrossRef]
- Weidenmaier, C.; Kokai-Kun, J.F.; Kulauzovic, E.; Kohler, T.; Thumm, G.; Stoll, H.; Gotz, F.; Peschel, A. Differential roles of sortase-anchored surface proteins and wall teichoic acid in Staphylococcus aureus nasal colonization. Int. J. Med. Microbiol. 2008, 298, 505–513. [Google Scholar] [CrossRef]
- Kerdudou, S.; Laschke, M.W.; Sinha, B.; Preissner, K.T.; Menger, M.D.; Herrmann, M. Fibronectin binding proteins contribute to the adherence of Staphylococcus aureus to intact endothelium in vivo. Thromb. Haemost. 2006, 96, 183–189. [Google Scholar] [CrossRef]
- Corrigan, R.M.; Miajlovic, H.; Foster, T.J. Surface proteins that promote adherence of Staphylococcus aureus to human desquamated nasal epithelial cells. BMC Microbiol. 2009, 9, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergara-Irigaray, M.; Maira-Litran, T.; Merino, N.; Pier, G.B.; Penades, J.R.; Lasa, I. Wall teichoic acids are dispensable for anchoring the PNAG exopolysaccharide to the Staphylococcus aureus cell surface. Microbiology 2008, 154, 865–877. [Google Scholar] [CrossRef] [Green Version]
- Gross, M.; Cramton, S.E.; Gotz, F.; Peschel, A. Key role of teichoic acid net charge in Staphylococcus aureus colonization of artificial surfaces. Infect. Immun. 2001, 69, 3423. [Google Scholar] [CrossRef] [Green Version]
- Maxe, I.; Ryden, C.; Wadstrom, T.; Rubin, K. Specific attachment of Staphylococcus aureus to immobilized fibronectin. Infect. Immun. 1986, 54, 695–704. [Google Scholar] [CrossRef] [Green Version]
- Peschel, A.; Otto, M.; Jack, R.W.; Kalbacher, H.; Jung, G.; Gotz, F. Inactivation of the dlt Operon in Staphylococcus aureus Confers Sensitivity to Defensins, Protegrins, and Other Antimicrobial Peptides. J. Biol. Chem. 1999, 274, 8405–8410. [Google Scholar] [CrossRef] [Green Version]
- Bur, S.; Preissner, K.T.; Herrmann, M.; Bischoff, M. The Staphylococcus aureus Extracellular Adherence Protein Promotes Bacterial Internalization by Keratinocytes Independent of Fibronectin-Binding Proteins. J. Investig. Dermatol. 2013, 133, 2004–2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dramsi, S.; Magnet, S.; Davison, S.; Arthur, M. Covalent attachment of proteins to peptidoglycan. FEMS Microbiol. Rev. 2008, 32, 307–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuhaus, F.C.; Baddiley, J. A Continuum of Anionic Charge: Structures and Functions of d-Alanyl-Teichoic Acids in Gram-Positive Bacteria. Microbiol. Mol. Biol. Rev. 2003, 67, 686–723. [Google Scholar] [CrossRef] [Green Version]
- Bellion, M.; Santen, L.; Mantz, H.; Hahl, H.; Quinn, A.; Nagel, A.; Gilow, C.; Weitenberg, C.; Schmitt, Y.; Jacobs, K. Protein adsorption on tailored substrates: Long-range forces and conformational changes. J. Phys. Condens. Matter 2008, 20, 404226. [Google Scholar] [CrossRef]
- Schlag, M.; Biswas, R.; Krismer, B.; Kohler, T.; Zoll, S.; Yu, W.; Schwarz, H.; Peschel, A.; Gotz, F. Role of staphylococcal wall teichoic acid in targeting the major autolysin Atl. Mol. Microbiol. 2010, 75, 864–873. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.; Frankel, M.B.; Schneewind, O.; Missiakas, D. Release of protein A from the cell wall of Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2014, 111, 1574–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertsche, U.; Yang, S.J.; Kuehner, D.; Wanner, S.; Mishra, N.N.; Roth, T.; Nega, M.; Schneider, A.; Mayer, C.; Grau, T.; et al. Increased Cell Wall Teichoic Acid Production and D-alanylation Are Common Phenotypes among Daptomycin-Resistant Methicillin-Resistant Staphylococcus aureus (MRSA) Clinical Isolates. PLoS ONE 2013, 8, e67398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Wely, K.H.; Swaving, J.; Freudl, R.; Driessen, A.J. Translocation of proteins across the cell envelope of Gram-positive bacteria. FEMS Microbiol. Rev. 2001, 25, 437–454. [Google Scholar] [CrossRef]
- Sarvas, M.; Harwood, C.R.; Bron, S.; van Dijl, J.M. Post-translocational folding of secretory proteins in Gram-positive bacteria. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2004, 1694, 311–327. [Google Scholar] [CrossRef]
- Forster, B.M.; Marquis, H. Protein transport across the cell wall of monoderm Gram-positive bacteria. Mol. Microbiol. 2012, 84, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Hyyrylainen, H.L.; Vitikainen, M.; Thwaite, J.; Wu, H.; Sarvas, M.; Harwood, C.R.; Kontinen, V.P.; Stephenson, K. D-Alanine substitution of teichoic acids as a modulator of protein folding and stability at the cytoplasmic membrane/cell wall interface of Bacillus subtilis. J. Biol. Chem. 2000, 275, 26696–26703. [Google Scholar] [CrossRef]
- Nouaille, S.; Commissaire, J.; Gratadoux, J.J.; Ravn, P.; Bolotin, A.; Gruss, A.; Loir, Y.L.; Langella, P. Influence of Lipoteichoic Acid d -Alanylation on Protein Secretion in Lactococcus lactis as Revealed by Random Mutagenesis. Appl. Environ. Microbiol. 2004, 70, 1600–1607. [Google Scholar] [CrossRef] [Green Version]
- Davis, E.; Kennedy, D.; Halperin, S.A.; Lee, S.F. Role of the Cell Wall Microenvironment in Expression of a Heterologous SpaP-S1 Fusion Protein by Streptococcus gordonii. Appl. Environ. Microbiol. 2011, 77, 1660–1666. [Google Scholar] [CrossRef] [Green Version]
- Schneewind, O.; Missiakas, D. Lipoteichoic Acids, Phosphate-Containing Polymers in the Envelope of Gram-Positive Bacteria. J. Bacteriol. 2014, 196, 1133–1142. [Google Scholar] [CrossRef] [Green Version]
- Peschel, A.; Vuong, C.; Otto, M.; Gotz, F. The d-Alanine Residues of Staphylococcus aureus Teichoic Acids Alter the Susceptibility to Vancomycin and the Activity of Autolytic Enzymes. Antimicrob. Agents Chemother. 2000, 44, 2845–2847. [Google Scholar] [CrossRef] [Green Version]
- Biswas, R.; Voggu, L.; Simon, U.K.; Hentschel, P.; Thumm, G.; Gotz, F. Activity of the major staphylococcal autolysin Atl. FEMS Microbiol. Lett. 2006, 259, 260–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenbeis, J.; Saffarzadeh, M.; Peisker, H.; Jung, P.; Thewes, N.; Preissner, K.T.; Herrmann, M.; Molle, V.; Geisbrecht, B.V.; Jacobs, K.; et al. The Staphylococcus aureus Extracellular Adherence Protein Eap Is a DNA Binding Protein Capable of Blocking Neutrophil Extracellular Trap Formation. Front. Cell. Infect. Microbiol. 2018, 8, 235. [Google Scholar] [CrossRef]
- Li, B.; Logan, B.E. Bacterial adhesion to glass and metal-oxide surfaces. Colloids Surf. B Biointerfaces 2004, 36, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Beaussart, A.; El-Kirat-Chatel, S.; Herman, P.; Alsteens, D.; Mahillon, J.; Hols, P.; Dufrene, Y.F. Single-Cell Force Spectroscopy of Probiotic Bacteria. Biophys. J. 2013, 104, 1886–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glowalla, E.; Tosetti, B.; Kronke, M.; Krut, O. Proteomics-Based Identification of Anchorless Cell Wall Proteins as Vaccine Candidates against Staphylococcus aureus. Infect. Immun. 2009, 77, 2719–2729. [Google Scholar] [CrossRef] [Green Version]
- Hempel, K.; Pane-Farre, J.; Otto, A.; Sievers, S.; Hecker, M.; Becher, D. Quantitative Cell Surface Proteome Profiling for SigBDependent Protein Expression in the Human Pathogen Staphylococcus aureus via Biotinylation Approach. J. Proteome Res. 2010, 9, 1579–1590. [Google Scholar] [CrossRef] [PubMed]
- Solis, N.; Larsen, M.R.; Cordwell, S.J. Improved accuracy of cell surface shaving proteomics in Staphylococcus aureus using a false-positive control. Proteomics 2010, 10, 2037–2049. [Google Scholar] [CrossRef] [Green Version]
- Ventura, C.L.; Malachowa, N.; Hammer, C.H.; Nardone, G.A.; Robinson, M.A.; Kobayashi, S.D.; DeLeo, F.R. Identification of a Novel Staphylococcus aureus Two-Component Leukotoxin Using Cell Surface Proteomics. PLoS ONE 2010, 5, e11634. [Google Scholar] [CrossRef]
- Dreisbach, A.; Hempel, K.; Buist, G.; Hecker, M.; Becher, D.; van Dijl, J.M. Profiling the surfacome of Staphylococcus aureus. Proteomics 2010, 10, 3082–3096. [Google Scholar] [CrossRef]
- Dreisbach, A.; van Dijl, J.M.; Buist, G. The cell surface proteome of Staphylococcus aureus. Proteomics 2011, 11, 3154–3168. [Google Scholar] [CrossRef] [PubMed]
- Lessel, M.; Baumchen, O.; Klos, M.; Hahl, H.; Fetzer, R.; Paulus, M.; Seemann, R.; Jacobs, K. Self-assembled silane monolayers: An efficient step-by-step recipe for high-quality, low energy surfaces. Surf. Interface Anal. 2015, 47, 557–564. [Google Scholar] [CrossRef]
- Hamilton, A.; Popham, D.L.; Carl, D.J.; Lauth, X.; Nizet, V.; Jones, A.L. Penicillin-Binding Protein 1a Promotes Resistance of Group B Streptococcus to Antimicrobial Peptides. Infect. Immun. 2006, 74, 6179–6187. [Google Scholar] [CrossRef] [Green Version]
- Beaussart, A.; Herman, P.; El-Kirat-Chatel, S.; Lipke, P.N.; Kucharikova, S.; Dijck, P.V.; Dufrene, Y.F. Single-cell force spectroscopy of the medically important Staphylococcus epidermidis–Candida albicans interaction. Nanoscale 2013, 5, 10894. [Google Scholar] [CrossRef] [Green Version]
- Zeng, G.; Muller, T.; Meyer, R.L. Single-Cell Force Spectroscopy of Bacteria Enabled by Naturally Derived Proteins. Langmuir 2014, 30, 4019–4025. [Google Scholar] [CrossRef] [PubMed]
- Herman, P.; El-Kirat-Chatel, S.; Beaussart, A.; Geoghegan, J.A.; Foster, T.J.; Dufrene, Y.F. The binding force of the staphylococcal adhesin SdrG is remarkably strong. Mol. Microbiol. 2014, 93, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Herman, P.; El-Kirat-Chatel, S.; Beaussart, A.; Geoghegan, J.A.; Vanzieleghem, T.; Foster, T.J.; Hols, P.; Mahillon, J.; Dufrene, Y.F. Forces Driving the Attachment of Staphylococcus epidermidis to Fibrinogen-Coated Surfaces. Langmuir 2013, 29, 13018–13022. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spengler, C.; Nolle, F.; Thewes, N.; Wieland, B.; Jung, P.; Bischoff, M.; Jacobs, K. Using Knock-Out Mutants to Investigate the Adhesion of Staphylococcus aureus to Abiotic Surfaces. Int. J. Mol. Sci. 2021, 22, 11952. https://doi.org/10.3390/ijms222111952
Spengler C, Nolle F, Thewes N, Wieland B, Jung P, Bischoff M, Jacobs K. Using Knock-Out Mutants to Investigate the Adhesion of Staphylococcus aureus to Abiotic Surfaces. International Journal of Molecular Sciences. 2021; 22(21):11952. https://doi.org/10.3390/ijms222111952
Chicago/Turabian StyleSpengler, Christian, Friederike Nolle, Nicolas Thewes, Ben Wieland, Philipp Jung, Markus Bischoff, and Karin Jacobs. 2021. "Using Knock-Out Mutants to Investigate the Adhesion of Staphylococcus aureus to Abiotic Surfaces" International Journal of Molecular Sciences 22, no. 21: 11952. https://doi.org/10.3390/ijms222111952
APA StyleSpengler, C., Nolle, F., Thewes, N., Wieland, B., Jung, P., Bischoff, M., & Jacobs, K. (2021). Using Knock-Out Mutants to Investigate the Adhesion of Staphylococcus aureus to Abiotic Surfaces. International Journal of Molecular Sciences, 22(21), 11952. https://doi.org/10.3390/ijms222111952