
Developmental Toxicity and Biotransformation of Two Anti-Epileptics in Zebrafish Embryos and Early Larvae

 ,
,  , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Exposure of Zebrafish Embryos and Larvae to CBZ and PHE—Gross Morphological Investigation
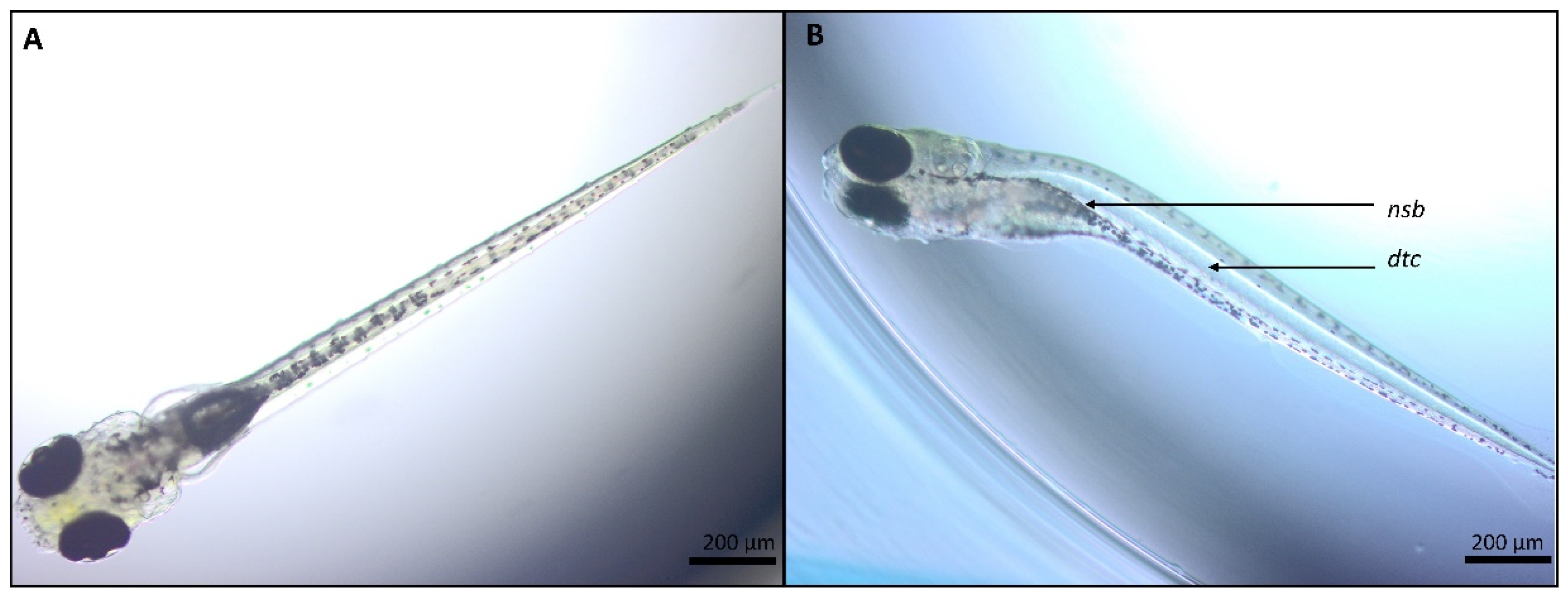
2.1.1. CBZ and Its Metabolite E-CBZ
2.1.2. Phenytoin and Its Metabolite HPPH
2.2. Exposure of Zebrafish Embryos and Larvae to CBZ and PHE—Analytical Investigation
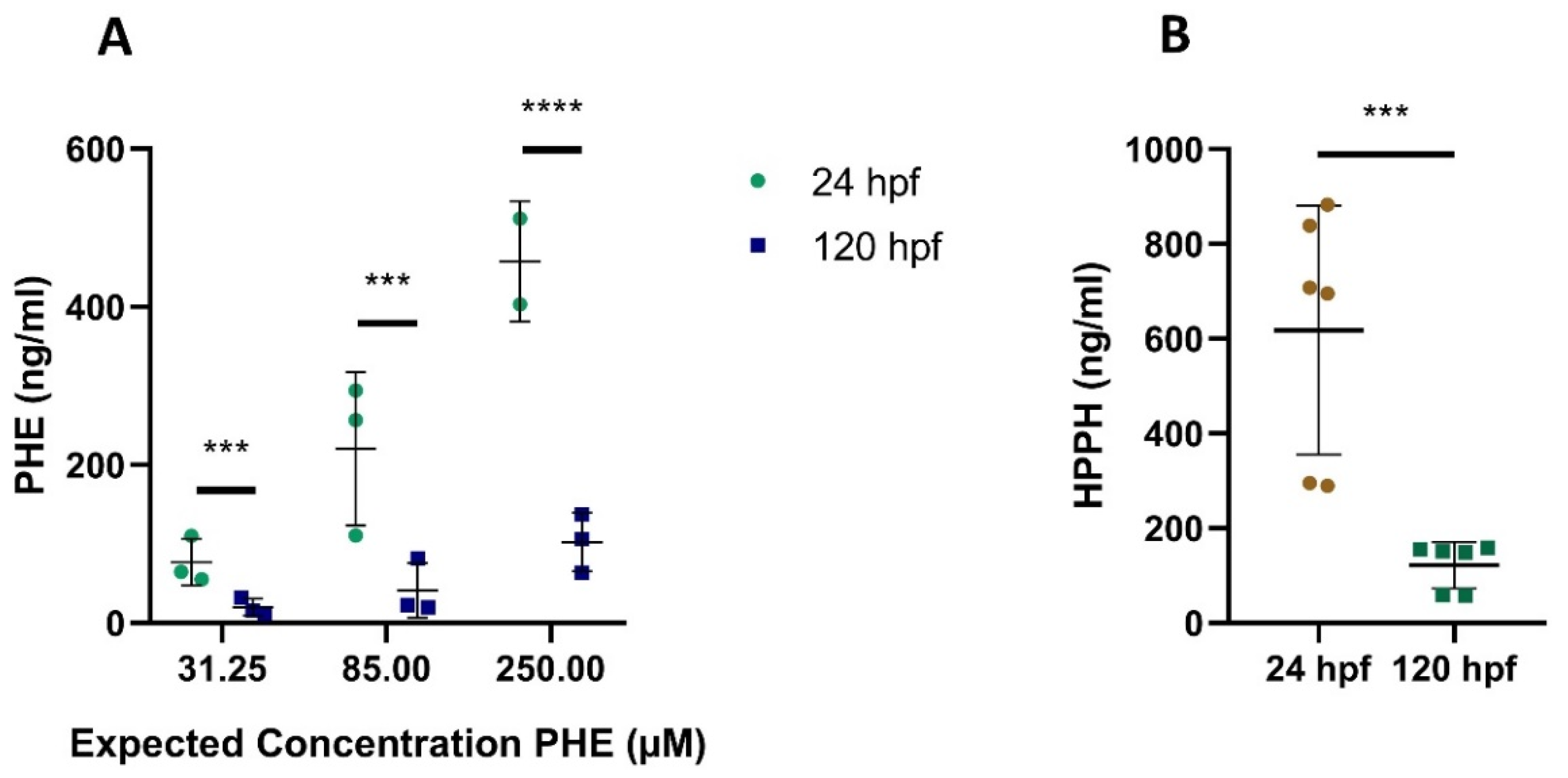
2.2.1. Quantification of CBZ and PHE, and Their Metabolites E-CBZ and HPPH, in Medium Samples after Exposure
2.2.2. Quantification of the Metabolites E-CBZ and HPPH in the Culture Medium after Exposure to the Parent Compounds
2.2.3. Quantification of the Parent Compounds CBZ and PHE, and Their Metabolites E-CBZ and HPPH, in Zebrafish Embryos/Larvae Homogenate Extracts
Quantification of CBZ and PHE, and Their Metabolites E-CBZ and HPPH, in Zebrafish Embryos/Larvae Homogenate Extracts after Direct Exposure
Quantification of the Metabolite, E-CBZ and HPPH, in Embryos and Larvae after Exposure to CBZ and PHE
2.3. In-Vitro Metabolism of CBZ and PHE: A Cross-Species Comparison
2.3.1. Quantification of E-CBZ after Microsomal Incubation with CBZ
2.3.2. Quantification of HPPH after Microsomal Incubation with PHE
3. Discussion
4. Materials and Methods
4.1. Adult Zebrafish Housing and Egg Production
4.2. Zebrafish Embryo and Larva Exposure to CBZ, E-CBZ, PHE and HPPH
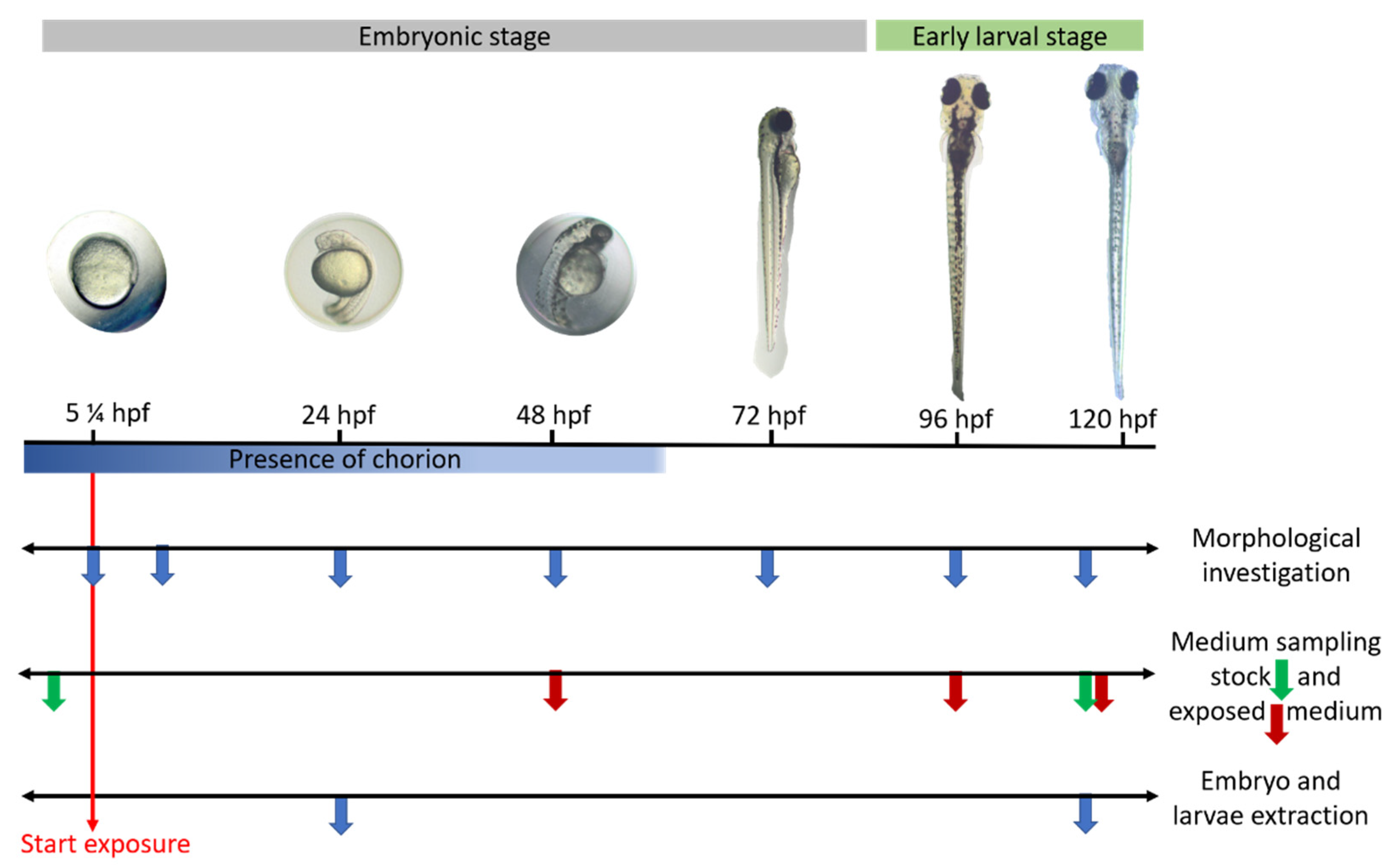
4.2.1. Test Conditions: Zebrafish Embryos and Larvae Exposure
4.2.2. Morphological Evaluation of the Embryos/Larvae over Time
4.2.3. Extraction of the CBZ and PHE, and the Metabolites, E-CBZ and HPPH, in Embryos and Larvae
4.3. In-Vitro Drug Metabolism of CBZ and PHE in Adult Zebrafish, Rat, Rabbit and Human Tissue
4.3.1. Adult Zebrafish Liver Tissue Collection and Microsomal Protein Preparation
4.3.2. Microsomal Incubation with CBZ and PHE
4.4. Analytical Investigation
4.4.1. Quantification of the Parent Compounds, CBZ and PHE, in Zebrafish Embryo Culture Medium and Zebrafish Embryos/Larvae Extracts
4.4.2. Quantification of the Metabolites, E-CBZ and HPPH in Zebrafish Embryo Culture Medium, Zebrafish Embryos/Larvae Extracts and Microsomal Reaction Samples
4.5. Calculated and Statistical Data
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACN | acetonitrile |
| AOP | adverse outcome pathway |
| CBZ | carbamazepine |
| CYP | cytochrome P450 |
| DMSO | dimethyl sulfoxide |
| E-CBZ | carbamazepine-10,11-epoxide |
| ECVAM | European Centre for the Validation of Alternative Methods |
| EDTA | ethylenediaminetetraacetic acid |
| EMA | European Medicines Agency |
| EMS | embryo solution |
| H | hours |
| Hpf | hours post fertilization |
| H2O | water |
| HPLC | high-performance liquid chromatography |
| HPPH | 5-(4-hydroxyphenyl)-5-phenylhydantoin |
| IS | internal standard |
| L | liters |
| LAMO | lamotrigine |
| LC-MS | liquid chromatography-mass spectrometry |
| LLOQ | lower limit of quantification |
| mDarT | metabolic Danio rerio test |
| Min | minutes |
| mL | millilitres |
| mZEDTA | metabolic zebrafish embryo developmental toxicity assay |
| PHE | phenytoin |
| µL | microliters |
| Xg | time gravity |
| % | percentage |
References
- Willhite, C.C.; Mirkes, P.E. Developmental Toxicology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 2, ISBN 9780123864543. [Google Scholar]
- EMA ICH S5 (R3) guideline on reproductive toxicology: Detection of Toxicity to Reproduction for Human Pharmaceuticals Step 5. Eur. Med. Agency Comm. Med. Prod. Hum. Use 2020, 5.
- Brannen, K.C.; Panzica-Kelly, J.M.; Danberry, T.L.; Augustine-Rauch, K.A. Development of a zebrafish embryo teratogenicity assay and quantitative prediction model. Birth Defects Res. Part B Dev. Reprod. Toxicol. 2010, 89, 66–77. [Google Scholar] [CrossRef]
- Horzmann, K.A.; Freeman, J.L. Making waves: New developments in toxicology with the zebrafish. Toxicol. Sci. 2018, 163, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Teraoka, H.; Dong, W.; Hiraga, T. Zebrafish as a novel experimental model for developmental toxicology. Congenit. Anom. 2003, 43, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Parng, C.; Seng, W.L.; Semino, C.; McGrath, P. Zebrafish: A preclinical model for drug screening. Assay Drug Dev. Technol. 2002, 1, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Cassar, S.; Adatto, I.; Freeman, J.L.; Gamse, J.T.; Lawrence, C.; Muriana, A.; Peterson, R.T.; Van Cruchten, S.; Zon, L.I.; Chicago, N.; et al. Use of Zebrafish in Drug Discovery Toxicology. Chem. Res. Toxicol. 2020, 33, 95–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, S.; Marigliani, B.; Akgün-Ölmez, S.G.; Ireland, D.; Cruz, R.; Busquet, F.; Flick, B.; Lalu, M.; Ghandakly, E.C.; de Vries, R.B.M.; et al. A systematic review to compare chemical hazard predictions of the zebrafish embryotoxicity test with mammalian prenatal developmental toxicity. Toxicol. Sci. 2021, 183, 14–35. [Google Scholar] [CrossRef] [PubMed]
- Augustine-Rauch, K.; Zhang, C.X.; Panzica-Kelly, J.M. In vitro developmental toxicology assays: A review of the state of the science of rodent and zebrafish whole embryo culture and embryonic stem cell assays. Birth Defects Res. Part C Embryo Today Rev. 2010, 90, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Ball, J.; Hillegass, J.M.; Flood, A.; Zhang, C.X.; Panzica-Kelly, J.; Cao, J.; Coburn, A.; Enright, B.P.; Tornesi, M.B.; Hetheridge, M.; et al. Inter-laboratory assessment of a harmonized zebrafish developmental toxicology assay—Progress report on phase I. Reprod. Toxicol. 2012, 33, 155–164. [Google Scholar] [CrossRef]
- Selderslaghs, I.W.T.; Blust, R.; Witters, H.E. Feasibility study of the zebrafish assay as an alternative method to screen for developmental toxicity and embryotoxicity using a training set of 27 compounds. Reprod. Toxicol. 2012, 33, 142–154. [Google Scholar] [CrossRef]
- Coecke, S.; Ahr, H.; Blaauboer, B.J.; Bremer, S.; Casati, S.; Castell, J.; Combes, R.; Corvi, R.; Crespi, C.L.; Cunningham, M.L.; et al. Metabolism: A bottleneck in in vitro toxicological test development. ATLA Altern. Lab. Anim. 2006, 34, 49–84. [Google Scholar] [CrossRef]
- Piersma, A.H.; Ezendam, J.; Luijten, M.; Muller, J.J.A.; Rorije, E.; Van Der Ven, L.T.M.; Van Benthem, J. A critical appraisal of the process of regulatory implementation of novel in vivo and in vitro methods for chemical hazard and risk assessment. Crit. Rev. Toxicol. 2014, 44, 876–894. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.A.; Hyland, R.; Jones, B.C.; Smith, D.A.; Hurst, S.; Goosen, T.C.; Peterkin, V.; Koup, J.R.; Ball, S.E. Drug-drug interactions for UDP-glucuronosyltransferase substrates: A pharmacokinetic explanation for typically observed low exposure (AUC 1/AUC) ratios. Drug Metab. Dispos. 2004, 32, 1201–1208. [Google Scholar] [CrossRef] [Green Version]
- Fred Peter, G. Cytochrome p450 and chemical toxicology. Chem. Res. Toxicol. 2008, 21, 70–83. [Google Scholar] [CrossRef]
- Strähle, U.; Scholz, S.; Geisler, R.; Greiner, P.; Hollert, H.; Rastegar, S.; Schumacher, A.; Selderslaghs, I.; Weiss, C.; Witters, H.; et al. Zebrafish embryos as an alternative to animal experiments—A commentary on the definition of the onset of protected life stages in animal welfare regulations. Reprod. Toxicol. 2012, 33, 128–132. [Google Scholar] [CrossRef]
- Field, H.A.; Ober, E.A.; Roeser, T.; Stainier, D.Y.R. Formation of the digestive system in zebrafish I. Liver morphogenesis. Dev. Biol. 2003, 253, 279–290. [Google Scholar] [CrossRef] [Green Version]
- Verbueken, E.; Alsop, D.; Saad, M.; Pype, C.; Van Peer, E.; Casteleyn, C.; Van Ginneken, C.; Wilson, J.; Van Cruchten, S.J. In Vitro Biotransformation of Two Human CYP3A Probe Substrates and Their Inhibition during Early Zebrafish Development. Int. J. Mol. Sci. 2017, 18, 217. [Google Scholar] [CrossRef] [Green Version]
- Verbueken, E.; Bars, C.; Ball, J.S.; Periz-stanacev, J.; Marei, W.F.A.; Tochwin, A.; Gabriëls, I.J.; Michiels, E.D.G.; Stinckens, E.; Vergauwen, L.; et al. From mRNA Expression of Drug Disposition Genes to In Vivo Assessment of CYP-Mediated Biotransformation during Zebrafish Embryonic and Larval Development. Int. J. Mol. Sci. 2018, 19, 3976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otte, J.C.; Schmidt, A.D.; Hollert, H.; Braunbeck, T. Spatio-temporal development of CYP1 activity in early life-stages of zebrafish (Danio rerio). Aquat. Toxicol. 2010, 100, 38–50. [Google Scholar] [CrossRef]
- Bräunig, J.; Schiwy, S.; Broedel, O.; Müller, Y.; Frohme, M.; Hollert, H.; Keiter, S.H. Time-dependent expression and activity of cytochrome P450 1s in early life-stages of the zebrafish (Danio rerio). Environ. Sci. Pollut. Res. 2015, 22, 16319–16328. [Google Scholar] [CrossRef]
- Oziolor, E.M.; Carey, A.N.; Matson, C.W. A non-destructive BFCOD assay for in vivo measurement of cytochrome P450 3A (CYP3A) enzyme activity in fish embryos and larvae. Ecotoxicology 2017, 26, 809–819. [Google Scholar] [CrossRef] [PubMed]
- Kais, B.; Schiwy, S.; Hollert, H.; Keiter, S.H.; Braunbeck, T. In vivo EROD assays with the zebrafish (Danio rerio) as rapid screening tools for the detection of dioxin-like activity. Sci. Total Environ. 2017, 590–591, 269–280. [Google Scholar] [CrossRef]
- Otte, J.C.; Schultz, B.; Fruth, D.; Fabian, E.; Van Ravenzwaay, B.; Salinas, E.R. Intrinsic Xenobiotic Metabolizing Enzyme Activities in Early Life Stages of Zebrafish ( Danio rerio ). Toxicol. Sci. 2017, 159, 86–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaji, T.; Yamashita, N.; Umeda, H.; Zhang, S. Cytochrome P450 Expression and Chemical Metabolic Activity before Full Liver Development in Zebrafish. Pharmaceuticals 2020, 13, 456. [Google Scholar] [CrossRef] [PubMed]
- Loerracher, A.K.; Braunbeck, T. Inducibility of cytochrome P450-mediated 7-methoxycoumarin-O-demethylase activity in zebrafish (Danio rerio) embryos. Aquat. Toxicol. 2020, 225, 105540. [Google Scholar] [CrossRef] [PubMed]
- Busquet, F.; Nagel, R.; Von Landenberg, F.; Mueller, S.O.; Huebler, N.; Broschard, T.H. Development of a New Screening Assay to Identify Proteratogenic Substances using Zebrafish Danio rerio Embryo Combined with an Exogenous Mammalian Metabolic Activation System ( m Dar T ) Franc. Toxicol. Sci. 2008, 104, 177–188. [Google Scholar] [CrossRef] [Green Version]
- Pype, C.; Verbueken, E.; Saad, M.A.; Casteleyn, C.R.; Van Ginneken, C.J.; Knapen, D.; Van Cruchten, S.J. Incubation at 32.5 °C and above causes malformations in the zebrafish embryo. Reprod. Toxicol. 2015, 56, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Pype, C.; Verbueken, E.; Saad, M.A.; Bars, C.; Van Ginneken, C.; Knapen, D.; Van Cruchten, S.J. Antioxidants reduce reactive oxygen species but not embryotoxicity in the metabolic Danio rerio test (mDarT). Reprod. Toxicol. 2017, 72, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Hoyberghs, J.; Bars, C.; Pype, C.; Foubert, K.; Ayuso, M.; Van Ginneken, C.; Ball, J.; Van Cruchten, S.J. Refinement of the zebrafish embryo developmental toxicity assay. MethodsX 2020, 7, 101087. [Google Scholar] [CrossRef]
- Weigt, S.; Huebler, N.; Braunbeck, T.; von Landenberg, F.; Broschard, T.H. Zebrafish teratogenicity test with metabolic activation (mDarT): Effects of phase I activation of acetaminophen on zebrafish Danio rerio embryos. Toxicology 2010, 275, 36–49. [Google Scholar] [CrossRef]
- Šuláková, A.; Nykodemová, J.; Palivec, P.; Jurok, R.; Rimpelová, S.; Leonhardt, T.; Šíchová, K.; Páleníček, T.; Kuchař, M. 25CN-NBOMe Metabolites in Rat Urine, Human Liver Microsomes and C. elegans—Structure Determination and Synthesis of the Most Abundant Metabolites. Metabolites 2021, 11, 212. [Google Scholar] [CrossRef] [PubMed]
- Boos, J.; Küpker, F.; Blaschke, G.; Jürgens, H. Trofosfamide metabolism in different species-ifosfamide is the predominant metabolite. Cancer Chemother. Pharmacol. 1993, 33, 71–76. [Google Scholar] [CrossRef]
- Nau, H. Species differences in pharmacokinetics and drug teratogenesis. Environ. Health Perspect. 1986, Volume 70, 113–129. [Google Scholar] [CrossRef]
- Weigt, S.; Huebler, N.; Strecker, R.; Braunbeck, T.; Broschard, T.H. Zebrafish (Danio rerio) embryos as a model for testing proteratogens. Toxicology 2011, 281, 25–36. [Google Scholar] [CrossRef]
- Le Fol, V.; Brion, F.; Hillenweck, A.; Perdu, E.; Bruel, S.; Aït-Aïssa, S.; Cravedi, J.P.; Zalko, D. Comparison of the in vivo biotransformation of two emerging estrogenic contaminants, BP2 and BPS, in zebrafish embryos and adults. Int. J. Mol. Sci. 2017, 18, 704. [Google Scholar] [CrossRef] [PubMed]
- Kislyuk, S.; Kroonen, J.; Adams, E.; Augustijns, P.; de Witte, P.; Cabooter, D. Development of a sensitive and quantitative UHPLC-MS/MS method to study the whole-body uptake of pharmaceuticals in zebrafish. Talanta 2017, 174, 780–788. [Google Scholar] [CrossRef]
- Brox, S.; Seiwert, B.; Haase, N.; Küster, E.; Reemtsma, T. Metabolism of clofibric acid in zebrafish embryos (Danio rerio) as determined by liquid chromatography-high resolution-mass spectrometry. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2016, 185–186, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Brox, S.; Ritter, A.P.; Küster, E.; Reemtsma, T. A quantitative HPLC-MS/MS method for studying internal concentrations and toxicokinetics of 34 polar analytes in zebrafish (Danio rerio) embryos. Anal. Bioanal. Chem. 2014, 406, 4831–4840. [Google Scholar] [CrossRef]
- Kerr, B.R.; KenCneth, K.E.; Wurden, C.J.; Klein, C.M.; Kroetz, D.L.; Gonzalez, F.J.; Levy, R.H. Human liver carbamazepine metabolism—role of CYP3A4 and CYP2C8 in 10,11-epoxide formation. Biochem. Pharmacol. 1994, 47, 1969–1979. [Google Scholar] [CrossRef]
- Giessmann, T.; May, K.; Modess, C.; Wegner, D.; Hecker, U.; Zschiesche, M.; Dazert, P.; Grube, M.; Kroemer, H.K.; Siegmund, W. Carbamazepine regulates intestinal P-glycoprotein and multidrug resistance protein MRP2 and influences disposition of talinolol in humans. Clin. Pharmacol. Ther. 2004, 76, 192–200. [Google Scholar] [CrossRef]
- Yamashita, H.; Kazawa, T.; Minatogawa, Y.; Ebisawa, T.; Yamauchi, T. Time-course of hepatic cytochrome P450 subfamily induction by chronic carbamazepine treatment in rats. Int. J. Neuropsychopharmacol. 2002, 5, 47–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tateishi, T.; Asoh, M.; Nakura, H.; Watanabe, M.; Tanaka, M.; Kumai, T. Carbamazepine induces multiple cytochrome P450 subfamilies in rats. Chem. Biol. Interact. 1999, 117, 257–268. [Google Scholar] [CrossRef]
- Mesdjian, E.; Sérée, E.; Charvet, B.; Mirrione, A.; Bourgarel-Rey, V.; Desobry, A.; Barra, Y. Metabolism of carbamazepine by CYP3A6: A model for in vitro drug interactions studies. Life Sci. 1999, 64, 827–835. [Google Scholar] [CrossRef]
- Chang, W.; Hung, S.; Carleton, B.; Chung, W. An update on CYP2C9 polymorphisms and phenytoin metabolism: Implications for adverse effects. Expert Opin. Drug Metab. Toxicol. 2020, 16, 723–734. [Google Scholar] [CrossRef]
- Edeki, T.I.; Brase, D.A. Phenytoin Disposition and Toxicity: Role of Pharmacogenetic and Interethnic Factors. DRUG Metab. Rev. 1995, 27, 449–469. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.-G.; Park, J.-Y.; Kim, M.-J.; Shon, J.-H.; Yoon, Y.-R.; Cha, I.-J.; Lee, S.-S.; Oh, S.-W.; Kim, S.-W.; Flockhart, D.A. Inhibitory Effects of Tricyclic Antidepressants (TCAs) on Human Cytochrome P450 Enzymes in Vitro: Mechanism of Drug Interaction between TCAs and Phenytoin. Drug Metab. Dispos. 2002, 30, 1102–1107. [Google Scholar] [CrossRef] [Green Version]
- Ghosal, A.; Sadrieh, N.; Reik, L.; Levin, W.; Thomas, P.E. Induction of the male-specific cytochrome P450 3A2 in female rats by phenytoin. Arch. Biochem. Biophys. 1996, 332, 153–162. [Google Scholar] [CrossRef]
- Saitoh, T.; Kokue, E.; Shimoda, M. The impact of acute phase response on the plasma clearance of antipyrine, theophylline, phenytoin and nifedipine in rabbits. J. Vet. Pharmacol. Ther. 2000, 23, 153–158. [Google Scholar] [CrossRef]
- Doecke, C.J.; Sansom, L.N.; McManus, M.E. Phenytoin 4-hydroxylation by rabbit liver P4502C3 and identification of orthologs in human liver microsomes. Biochem. Biophys. Res. Commun. 1990, 166, 860–866. [Google Scholar] [CrossRef]
- Martinez, C.S.; Feas, D.A.; Siri, M.; Igartúa, D.E.; Chiaramoni, N.S.; Alonso, S.V.; Prieto, M.J. In vivo study of teratogenic and anticonvulsant e ff ects of antiepileptics drugs in zebra fi sh embryo and larvae. Neurotoxicol. Teratol. 2018, 66, 17–24. [Google Scholar] [CrossRef]
- Zhou, S.; Chen, Q.; Di, C.; Shao, Y.; Hollert, H.; Seiler, T. Behavioral profile alterations in zebrafish larvae exposed to environmentally relevant concentrations of eight priority pharmaceuticals. Sci. Total Environ. 2019, 664, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Pruvot, B.; Quiroz, Y.; Voncken, A.; Jeanray, N.; Piot, A.; Martial, J.A.; Muller, M. A panel of biological tests reveals developmental effects of pharmaceutical pollutants on late stage zebrafish embryos. Reprod. Toxicol. 2012, 34, 568–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Den Brandhof, E.; Montforts, M. Fish embryo toxicity of carbamazepine, diclofenac and metoprolol. Ecotoxicol. Environ. Saf. 2010, 73, 1862–1866. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Santiago-schübel, B.; Xiao, H.; Thiele, B.; Zhu, Z.; Qiu, Y.; Hollert, H.; Küppers, S. An efficient laboratory workflow for environmental risk assessment of organic chemicals. Chemosphere 2015, 131, 34–40. [Google Scholar] [CrossRef]
- Pohl, J.; Golovko, O.; Carlsson, G.; Orn, S.; Schmitz, M.; Ahi, E.P. Chemosphere Gene co-expression network analysis reveals mechanisms underlying ozone-induced carbamazepine toxicity in zebra fi sh (Danio rerio) embryos. Chemosphere J. 2021, 276, 130282. [Google Scholar] [CrossRef] [PubMed]
- Kelsh, R.N.; Schmid, B.; Eisen, J.S. Genetic Analysis of Melanophore Development in Zebrafish Embryos. Dev. Biol. 2000, 293, 277–293. [Google Scholar] [CrossRef]
- Pohl, J.; Golovko, O.; Carlsson, G.; Eriksson, J.; Glynn, A.; Stefan, O.; Weiss, J. Carbamazepine Ozonation Byproducts: Toxicity in Zebra fi sh (Danio rerio) Embryos and Chemical Stability. Environ. Sci. Technol. 2020, 54, 2913–2921. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; van Mil, H.G.J.; Richardson, M.K. Large-Scale assessment of the zebrafish embryo as a possible predictive model in toxicity testing. PLoS ONE 2011, 6, e21076. [Google Scholar] [CrossRef] [Green Version]
- Kamali, F.; Bali, D.; McLaughlin, W.; Seymour, R. Phenytoin metabolism to 5-(4-hydroxyphenyl)-5-phenylhydantoin (HPPH) in man, cat and rat in vitro and in vivo and suceptibility to phenytoin-induced gingival overgrowth. J. Periodontal Res. 1999, 34, 145–153. [Google Scholar] [CrossRef]
- Yamanaka, H.; Nakajima, M.; Hara, Y.; Katoh, M.; Tachibana, O.; Yamashita, J.; Yokoi, T. Urinary excretion of phenytoin metabolites, 5-(4’-hydroxyphenyl)-5-phenylhydantoin and its O-glucuronide in humans and analysis of genetic polymorphisms of UDP-glucuronosyltransferases. Drug Metab. Pharmacokinet. 2005, 20, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Brox, S.; Ritter, A.P.; Küster, E.; Reemtsma, T. Influence of the perivitelline space on the quantification of internal concentrations of chemicals in eggs of zebrafish embryos (Danio rerio). Aquat. Toxicol. 2014, 157, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Halbach, K.; Ulrich, N.; Goss, K.U.; Seiwert, B.; Wagner, S.; Scholz, S.; Luckenbach, T.; Bauer, C.; Schweiger, N.; Reemtsma, T. Yolk sac of zebrafish embryos as backpack for chemicals? Environ. Sci. Technol. 2020, 54, 10159–10169. [Google Scholar] [CrossRef] [PubMed]
- Pohl, J.; Ahrens, L.; Carlsson, G.; Golovko, O.; Norrgren, L.; Weiss, J.; Orn, S. Embryotoxicity of ozonated diclofenac, carbamazepine, and oxazepam in zebrafish (Danio rerio). Chemosphere 2019, 225, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Muniz, M.S.; Halbach, K.; Alves Araruna, I.C.; Martins, R.X.; Seiwert, B.; Lechtenfeld, O.; Reemtsma, T.; Farias, D. Moxidectin toxicity to zebrafish embryos: Bioaccumulation and biomarker responses. Environ. Pollut. 2021, 283, 117096. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ji, C.; Yan, L.; Lu, M.; Lu, C.; Zhao, M. The identification of the metabolites of chlorothalonil in zebrafish (Danio rerio) and their embryo toxicity and endocrine effects at environmentally relevant levels. Environ. Pollut. 2016, 218, 8–15. [Google Scholar] [CrossRef]
- Namdari, R.; Jones, K.; Chuang, S.S.; Van Cruchten, S.; Dincer, Z.; Downes, N.; Mikkelsen, L.F.; Harding, J.; Jäckel, S.; Jacobsen, B.; et al. Species selection for nonclinical safety assessment of drug candidates: Examples of current industry practice. Regul. Toxicol. Pharmacol. 2021, 126, 105029. [Google Scholar] [CrossRef]
- Chng, H.T.; Ho, H.K.; Yap, C.W.; Lam, S.H.; Chan, E.C.Y. An investigation of the bioactivation potential and metabolism profile of zebrafish versus human. J. Biomol. Screen. 2012, 17, 974–986. [Google Scholar] [CrossRef] [Green Version]
- Jones, H.S.; Trollope, H.T.; Hutchinson, T.H.; Panter, G.H.; Chipman, J.K. Metabolism of ibuprofen in zebrafish larvae. Xenobiotica 2012, 42, 1069–1075. [Google Scholar] [CrossRef]
- Alderton, W.; Berghmans, S.; Butler, P.; Chassaing, H.; Fleming, A.; Golder, Z.; Richards, F.; Gardner, I. Accumulation and metabolism of drugs and CYP probe substrates in zebrafish larvae. Xenobiotica 2010, 40, 547–557. [Google Scholar] [CrossRef]
- Saad, M.; Matheeussen, A.; Bijttebier, S.; Verbueken, E.; Pype, C.; Casteleyn, C.; Van Ginneken, C.; Apers, S.; Maes, L.; Cos, P.; et al. In vitro CYP-mediated drug metabolism in the zebrafish (embryo) using human reference compounds. Toxicol. Vitr. 2017, 42, 329–336. [Google Scholar] [CrossRef]
- Saad, M.; Bijttebier, S.; Matheeussen, A.; Verbueken, E.; Pype, C.; Casteleyn, C.; Van Ginneken, C.; Maes, L.; Cos, P.; Van Cruchten, S.J. Data in Brief UPLC / MS MS data of testosterone metabolites in human and zebra fi sh liver microsomes and whole zebra fi sh larval microsomes. Data Br. 2018, 16, 644–648. [Google Scholar] [CrossRef]
- Borges, A.C.; Pereira, N.; Franco, M.; Vale, L.; Pereira, M.; Cunha, M.V.; Amaro, A.; Albuquerque, T.; Rebelo, M. Implementation of a Zebrafish Health Program in a Research Facility: A 4-Year Retrospective Study. Zebrafish 2016, 13, S115–S125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Hamm, J.T.; Ceger, P.; Allen, D.; Stout, M.; Maull, E.A.; Baker, G.; Zmarowski, A.; Padilla, S.; Perkins, E.; Planchart, A.; et al. Characterizing sources of variability in zebrafish embryo screening protocols. ALTEX 2019, 36, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Nagel, R. DarT: The embryo test with the Zebrafish Danio rerio--a general model in ecotoxicology and toxicology. ALTEX Altern. Tierexperimenten 2002, 19 (Suppl. S1), 38–48. [Google Scholar]
- Hill, J.R. In Vitro Drug Metabolism Using Liver Microsomes. Curr. Protoc. Pharmacol. 2003, 7.8, 1–11. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Deviation Tail | Oedema | Malformation Head | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 120 hpf Experiment CBZ/E-CBZ | # | Coagulation | No Hatching | Curve | Tissue Deviation | Head | Pericardium | Yolk | Deviation Shape Head | Deviation Mouth | Deviation Eye | Deviation Pigmentation | Swim Bladder Not Inflated |
| EMS | 1 | 2/20 | 2/20 | 2/20 | 1/20 | ||||||||
| 2 | |||||||||||||
| EMS 0.5% DMSO | 1 | 1/20 | 1/20 | 1/20 | |||||||||
| 2 | 1/20 | 1/20 | 2/20 | ||||||||||
| EMS 1% DMSO | 1 | 1/20 | 2/20 | ||||||||||
| 2 | 1/20 | 1/20 | |||||||||||
| CBZ 31.25 µM | 1 | 8/20 ** | |||||||||||
| 2 | 2/20 | ||||||||||||
| CBZ 85 µM | 1 | 1/20 | 7/20 ** | ||||||||||
| 2 | 2/20 | 1/18 | 1/18 | 3/18 | 2/18 | 1/18 | 2/18 | ||||||
| CBZ 250 µM | 1 | 5/20 * | 2/20 | 5/20 * | 3/20 | 4/20 | 5/20 * | 19/20 **** | 16/20 **** | ||||
| 2 | 1/20 | 2/20 | 4/20 | 3/20 | 3/20 | 3/20 | 8/20 ** | 18/20 **** | 11/20 ** | ||||
| E-CBZ 250 µM | 1 | 1/20 | 1/20 | 1/20 | 1/20 | 3/20 | |||||||
| 2 | 1/20 | 1/20 | 1/20 | 6/20 * | |||||||||
| Deviation Tail | Oedema | Malformation Head | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 120 hpf Experiment PHE/HPPH | # | No Hatching | Elbow | Curve | Tissue Deviation | Head | Pericardium | Yolk | Deviation Shape Head | Deviation Eye | Deviation Pigmentation | Swim Bladder Not Inflated |
| EMS | 1 | 1/20 | 1/20 | |||||||||
| 2 | ||||||||||||
| EMS 0.5% DMSO | 1 | 2/20 | 2/20 | 1/20 | 1/20 | 1/20 | ||||||
| 2 | 1/20 | 1/20 | 1/20 | 1/20 | 2/20 | |||||||
| PHE 31.25 µM | 1 | 2/20 | 2/20 | 4/20 | ||||||||
| 2 | 1/20 | 4/20 | 2/20 | |||||||||
| PHE 85 µM | 1 | 3/20 | 2/20 | 1/20 | 4/20 | 2/20 | 2/20 | 6/20 | ||||
| 2 | 1/20 | 4/20 | 2/20 | 2/20 | 3/20 | 3/20 | 2/20 | 13/20 *** | ||||
| PHE 250 µM | 1 | 1/20 | 7/20 * | 1/20 | 1/20 | 1/20 | 1/20 | 2/20 | 3/20 | 3/20 | 16/20 **** | |
| 2 | 1/20 | 2/20 | 5/20 | 1/20 | 1/20 | 1/20 | 3/20 | 4/20 | 7/20 * | 15/20 **** | ||
| HPPH 250 µM | 1 | 2/20 | 1/20 | 2/20 | 5/20 | |||||||
| 2 | 1/20 | 6/20 | ||||||||||
| Nominal [X] | 31.25 µM | 85 µM | 250 µM | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Time Points | 0 hpf | 48 hpf | 96 hpf | 120 hpf | 0 hpf | 48 hpf | 96 hpf | 120 hpf | 0 hpf | 48 hpf | 96 hpf | 120 hpf | |
| Compound | CBZ | CBZ | CBZ | E-CBZ | CBZ | ||||||||
| Stock | 95.6 ± 2.0% | 101.6 ± 7.7% | 81.7 ± 18.7% | 90.4 ± 6.6% | 92.1 ± 5.3% | 98.4 ± 4.6% | |||||||
| Culture medium | 95.4 ± 16.2% | 101.2 ± 21.7% | 109.6 ± 5.4% | 110.0 ± 4.8% | 116.8 ± 4.7% | 113.9 ± 4.5% | 113.4 ± 14.1% | 114.2 ± 2.6% | 117.3 ± 9.3% | 111.5 ± 2.6% | |||
| Nominal [X] | 31.25 µM | 85 µM | 250 µM | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Time Points | 0 hpf | 48 hpf | 96 hpf | 120 hpf | 0 hpf | 48 hpf | 96 hpf | 120 hpf | 0 hpf | 48 hpf | 96 hpf | 120 hpf | |
| Compound | PHE | PHE | PHE | HPPH | PHE | ||||||||
| Stock | 53.3 ± 23.3% | 62.01 ± 5.25% | 86.4 ± 47.2% | 64.5 ± 1.2% | 24.7 ± 3.4% | 30.6 ± 7.9% | |||||||
| Culture medium | 80 ± 4.8% | 84.5 ± 8.4% | 81.2 ± 11.1% | 79.1 ± 19.6% | 64.8 ± 8.7% | 64.2 ± 10.7% | 33.2 ± 6.6% | 43.3 ± 8.4% | 89.9 ± 15.5% | 33.4 ± 1.2% | |||
| LC-MS System | Compound | Quantifier Transition | Qualifier Transition |
|---|---|---|---|
| ACQUITY UPLC-triple quadrupole system | CBZ | 237 → 194 | 237 → 165 |
| PHE | 253 → 182 | 253 → 104 | |
| LAMO | 256 → 211 | 256 → 108 | |
| Shimadzu UPLC-MS-8050 triple quadrupole system | E-CBZ | 253.1 → 210.2 | 253.1 → 180.3 |
| HPPH | 268.9 → 120.1 | 268.9 → 104.2 | |
| LAMO | 255.9 → 109.1 | 255.9 → 211.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bars, C.; Hoyberghs, J.; Valenzuela, A.; Buyssens, L.; Ayuso, M.; Van Ginneken, C.; Labro, A.J.; Foubert, K.; Van Cruchten, S.J. Developmental Toxicity and Biotransformation of Two Anti-Epileptics in Zebrafish Embryos and Early Larvae. Int. J. Mol. Sci. 2021, 22, 12696. https://doi.org/10.3390/ijms222312696
Bars C, Hoyberghs J, Valenzuela A, Buyssens L, Ayuso M, Van Ginneken C, Labro AJ, Foubert K, Van Cruchten SJ. Developmental Toxicity and Biotransformation of Two Anti-Epileptics in Zebrafish Embryos and Early Larvae. International Journal of Molecular Sciences. 2021; 22(23):12696. https://doi.org/10.3390/ijms222312696
Chicago/Turabian StyleBars, Chloé, Jente Hoyberghs, Allan Valenzuela, Laura Buyssens, Miriam Ayuso, Chris Van Ginneken, Alain J. Labro, Kenn Foubert, and Steven J. Van Cruchten. 2021. "Developmental Toxicity and Biotransformation of Two Anti-Epileptics in Zebrafish Embryos and Early Larvae" International Journal of Molecular Sciences 22, no. 23: 12696. https://doi.org/10.3390/ijms222312696
APA StyleBars, C., Hoyberghs, J., Valenzuela, A., Buyssens, L., Ayuso, M., Van Ginneken, C., Labro, A. J., Foubert, K., & Van Cruchten, S. J. (2021). Developmental Toxicity and Biotransformation of Two Anti-Epileptics in Zebrafish Embryos and Early Larvae. International Journal of Molecular Sciences, 22(23), 12696. https://doi.org/10.3390/ijms222312696