
Fighting Cancer with Bacteria and Their Toxins



Abstract
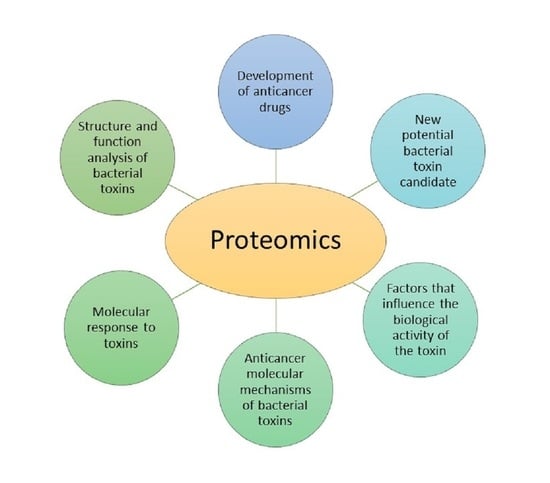
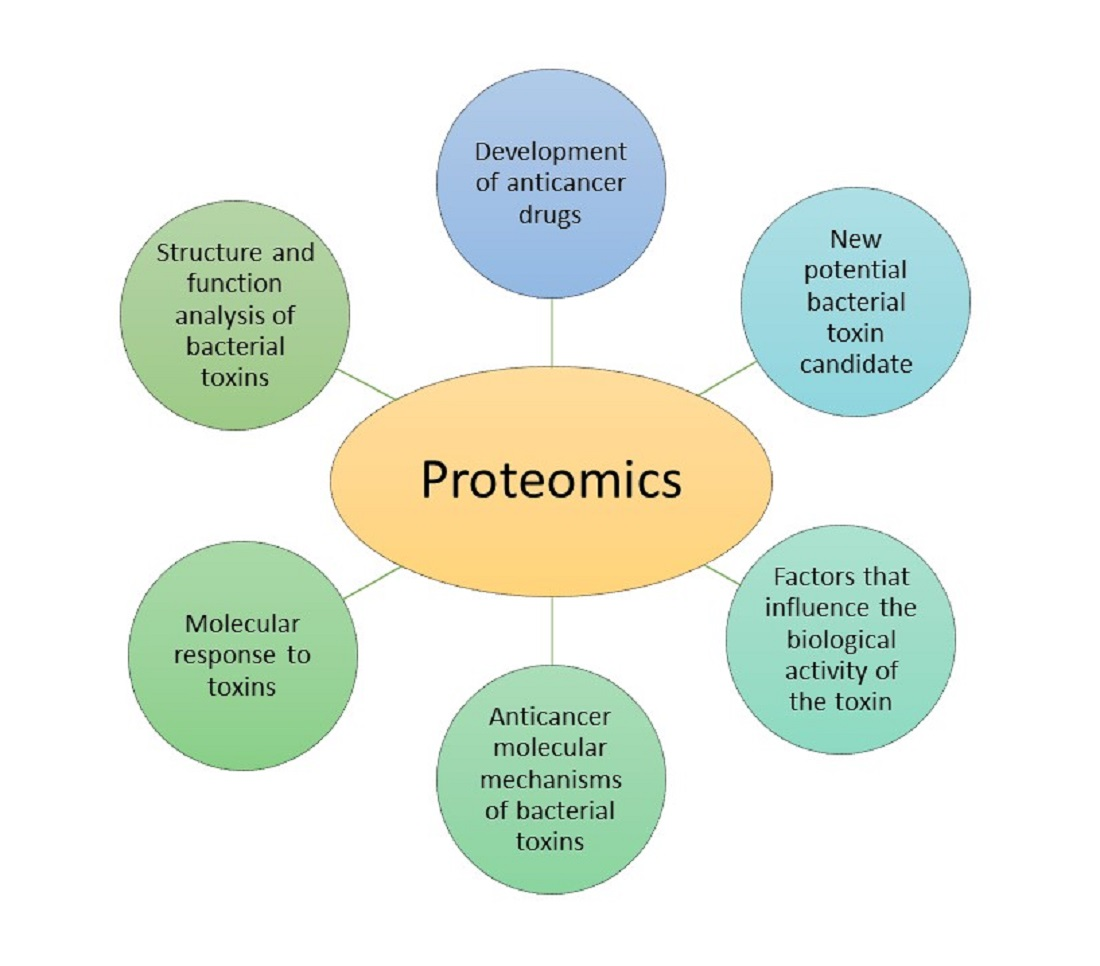
:
1. Introduction

{kind=link}
{kind=link}
{kind=link}
| Bacteria (Family/Genus/Species) | Type of Sample | Correlation or Mechanism | Reference |
|---|---|---|---|
| Granulicatella, Strepotoccocus, Abiotrophia | Buccal-oral, sputum | Significant difference in presence and distribution | [43] |
| Captocytophaga, Selenomonas, Veilonella | Saliva | Significant difference in presence and distribution | [44] |
| Veilonella, Megasphaera Actinomyces, Arthrobacter, Capnocytophaga Rothia, Streptococcus | Bronchoalveolar lavage | Significant difference in presence and distribution | [45] |
| Thermos, Ralstonia, Legionella | Lung cancer vs health lung tissue | Significant difference in presence and distribution | [46] |
| Streptococcus viridans, Granulicatella adiacens | Sputum | Significant difference in presence and distribution | [46] |
| Streptococcus intermedius, Mycobacterium tuberculosis | Lung tissue and bronchoscopy samples | Significant difference in presence and distribution | [6] |
| Streptococcus | Bronchial brushing | Significant difference in presence and distribution | [37] |
| Streptococcus, Veilonella | Lung cancer vs health lung tissue | Significant difference in presence and distribution | [39] |
| Streptococcus, Neisseria | Lung cancer vs health lung tissue | Significant difference in presence and distribution | [47] |
| Proctobacteria Firmicutes Bacteroides | Lung tissue | Significant difference in presence and distribution | [48] |
| Acidovorax, Klebsiella, Rhodoferax, Anaerococcus Cyanobacteria | Lung tissue | Significant difference in presence and distribution | [39,49] |
| Streptococcus, Prevotella | Lung tissue | Significant difference in presence and distribution | [50] |
| Sphingomonas, Blastomonas | Saliva | Significant difference in presence and distribution | [51] |
| Bradyrhizobium japanicum | Bronchial brushing | Significant difference in presence and distribution | [52] |
| Veillonella, Prevotella, Streptococcus | Lung cancer tissue | Upregulation of ERK and PI3K signaling pathways | [39] |
| Acidovorax, Klebsiella, Rhodoferax, Anaerococcus | Lung cancer vs health lung tissue | Significantly higher abundance of in SCC than in adenocarcinoma | [36] |
| Acidovorax | Lung cancer vs health lung tissue | Abundant in patients with TP53 mutation-positive SCCLC and smoking history | [36] |
| Bradyrhizobium japonicum, Acidovorax | Bronchial brushing | Significant difference in presence and distribution | [52] |
| Bacteroidaceae, Lachnospiraceae, Ruminococcaceae | Lung cancer vs health lung tissue | Significant difference in presence and distribution | [38] |
| Mycobacterium tuberculosis | Lung cancer vs health lung tissue | Chronic inflammation | [11] |
2. History of Using Bacteria in Cancer Therapy
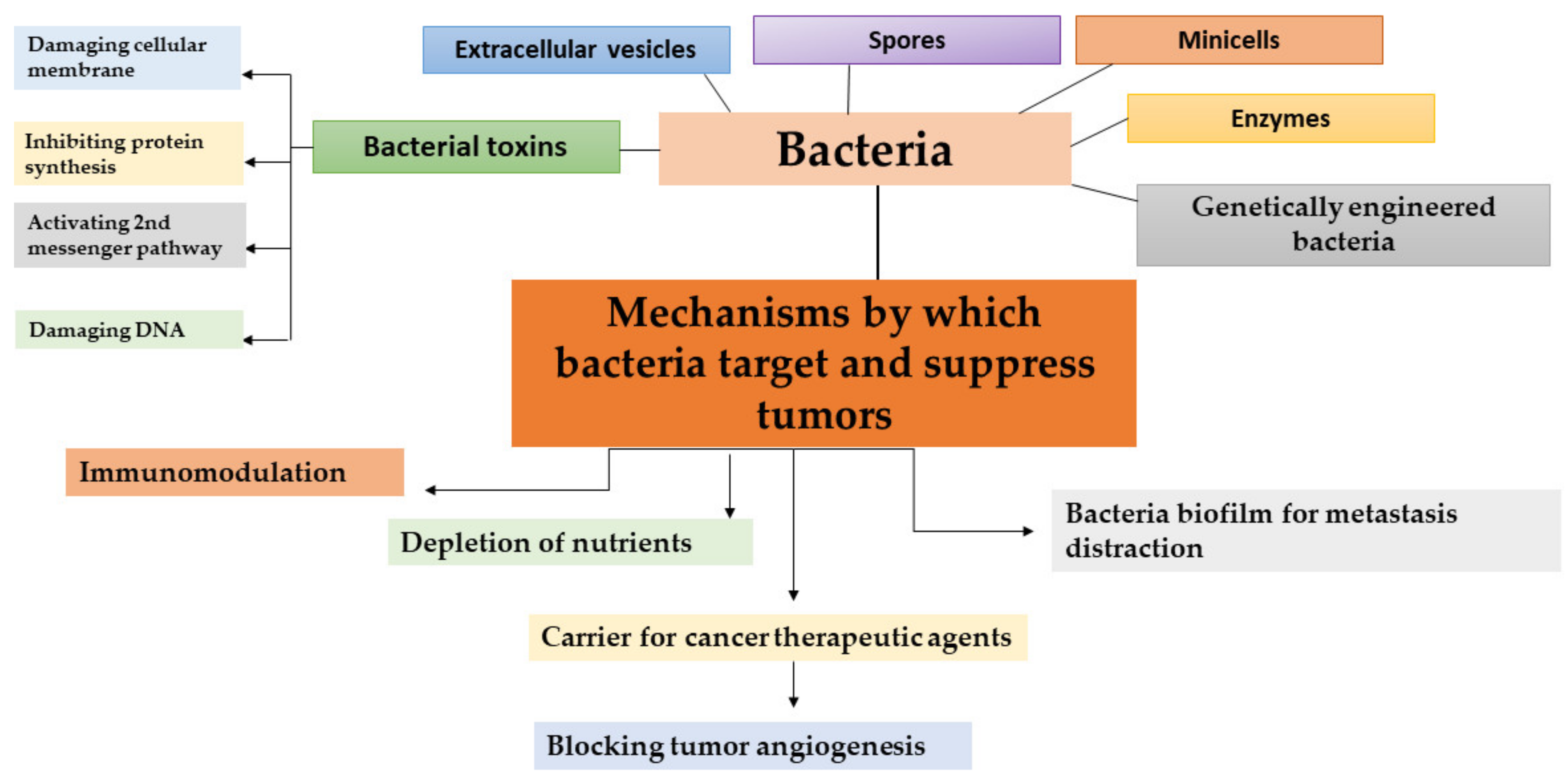
3. Molecular Mechanisms of Anticancer Bacterial Action
4. Bacterial Toxins for Cancer Therapy
4.1. Introduction to Bacterial Toxins
| Immunotoxin. | Antigen | Target | Toxin | Cancer type | Pipeline | Clinical Trial Identifier or Reference |
|---|---|---|---|---|---|---|
| Diphtheria Toxin Based | ||||||
| Denileukin Diftitox (DAB389IL2) | IL-2R | IL2 | DT (DAB389) | Melanoma, hematological | 2008. approved by FDA for CTCL | [82,83] |
| DAB8486IL2 | IL-2R | IL2 | DT (DAB486) | Hematological | Phase I/II | [84,85] |
| Tagraxofusp SL-401 (DT388-IL3) | IL-3R | Variant IL-3 | DT | Hematological | 2018 approved by FDA for BPDCN | [86] |
| Tf-CRM107 (transMID) | TfR | Transferrin | DT (CRM107) | CNS | Phase I, III | NCT 00088400 NCT 00083447 NCT 00052624 |
| DAB389EGF | EGFR | EGF | DT (DAB389) | EGFR positive cancers | Phase I/II | [87] |
| UCHT1 | CD3 | Murine anti-CD3-bis Fv | DT | Hematological | Phase I/II | NCT 00611208 NCT 01888081 |
| DT 2219ARL bispecific | CD19 and CD22 | Anti-CD22, Anti-CD 19 (sFv) | DT (DAB389) | Hematological | Phase I/II | NCT 00889408 NCT 02370160 |
| Pseudomonas Toxin Based | ||||||
| TP-38 | EGFR | TGFalpha | PE | CNS tumors | Phase II | NCT 00104091 NCT 00074334 |
| Moxetumomab pasudodotox | CD22 | Murine anti-CD22 d sFv fragment 2 | PE | Hematological | Phase I, II, III | NCT 02338050 NCT 01829711 NCT 02227108 NCT 01030536 |
| LMB-2 | CD25 | Anti-CD25scFv fragment | PE | Hematological, skin cancers | Phase II | NCT 00077922 NCT 00080536 NCT 00295958 NCT 00924170 NCT 00321555 NCT 00002765 |
| SS1P | Mesothelin | Murine antimesothelin dsFv fragmentr | PE (PE38) | Mesothelioma, Cervical, Head and Neck, Lung, ovarian cancers | Phase I/II | NCT 01362790 NCT 00024687 NCT 00006981 |
| Oportuzumabmonotox (VB4-8454) | EpCAM | Humanized anti-EpCAM scFv fragment | PE | Head and Neck, Squamous cell, bladder cancers | Phase II/III | NCT 00462488 NCT 00272181 NCT 02449239 |
4.2. Proteomic Analysis of Bacterial Toxins Targeting Cancer Cells
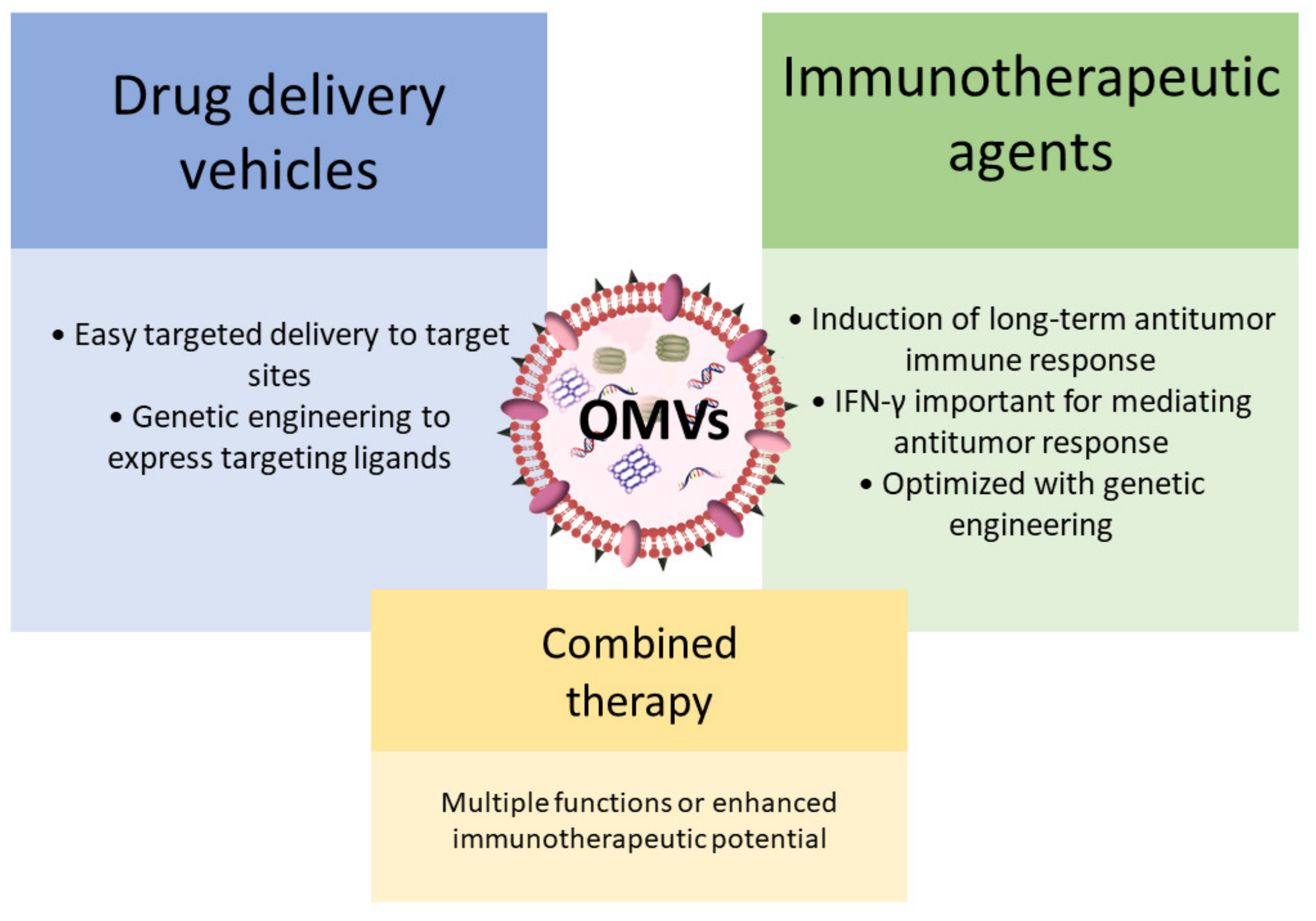
5. Bacterial Extracellular Vesicles as Model to Fight Cancer
6. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- International Agency for Research on Cancer. Cancer Incidence, Mortality and Prevalence Worldwide GLOBOCAN 2012. Available online: https://gco.iarc.fr/ (accessed on 27 September 2021).
- Malvezzi, M.; Carioli, G.; Bertuccio, P.; Boffetta, P.; Levi, F.; La Vecchia, C.; Negri, E. European cancer mortality predictions for the year 2017, with focus on lung cancer. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2017, 28, 1117–1123. [Google Scholar] [CrossRef]
- Novello, S.; Brahmer, J.R.; Stabile, L.P.; Siegfried, J.M. Gender-Related Differences in Lung Cancer. In IASLC Thoracic Oncology; Pass, H.I., Ball, D., Scagliotti, G.V., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 30–45. ISBN 9780323523578. [Google Scholar]
- Malhotra, J.; Malvezzi, M.; Negri, E.; La Vecchia, C.; Boffetta, P. Risk factors for lung cancer worldwide. Eur. Respir. J. 2016, 48, 889–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.-N.; Ma, Q.; Ge, Y.; Yi, C.-X.; Wei, L.-Q.; Tan, J.-C.; Chu, Q.; Li, J.-Q.; Zhang, P.; Wang, H. Microbiome dysbiosis in lung cancer: From composition to therapy. NPJ Precis. Oncol. 2020, 4, 33. [Google Scholar] [CrossRef] [PubMed]
- Cameron, S.J.S.; Lewis, K.E.; Huws, S.A.; Hegarty, M.J.; Lewis, P.D.; Pachebat, J.A.; Mur, L.A.J. A pilot study using metagenomic sequencing of the sputum microbiome suggests potential bacterial biomarkers for lung cancer. PLoS ONE 2017, 12, e0177062. [Google Scholar] [CrossRef] [Green Version]
- Gomes, S.; Cavadas, B.; Ferreira, J.; Marques, P.; Monteiro, C.; Sucena, M.; Sousa, C.; Vaz Rodrigues, L.; Teixeira, G.; Pinto, P.; et al. Profiling of lung microbiota discloses differences in adenocarcinoma and squamous cell carcinoma. Sci. Rep. 2019, 9, 12838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.; Su, X.; Yuan, M.; Zhang, S.; He, J.; Deng, Q.; Qiu, W.; Dong, H.; Cai, S. The characterization of lung microbiome in lung cancer patients with different clinicopathology. Am. J. Cancer Res. 2019, 9, 2047. [Google Scholar]
- Al-Hilu, S.A.; Al-Shujairi, W.H. Dual Role of Bacteria in Carcinoma: Stimulation and Inhibition. Int. J. Microbiol. 2020, 2020, 4639761. [Google Scholar] [CrossRef]
- De Martel, C.; Ferlay, J.; Franceschi, S.; Vignat, J.; Bray, F.; Forman, D.; Plummer, M. Global burden of cancers attributable to infections in 2008: A review and synthetic analysis. Lancet Oncol. 2012, 13, 607–615. [Google Scholar] [CrossRef]
- Shiels, M.; Albanes, D.; Virtamo, J.; Engels, E. Increased risk of lung cancer in men with tuberculosis in the alphatocopherol, beta-carotene cancer prevention study. Cancer Epidemiol. Biomark. Prev. 2011, 20, 672–678. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.K.; Tripathi, D.; Kulkarni, S.; Rajan, M.G.R. Mycobacterium tuberculosis H37Rv infected THP-1 cells induce epithelial mesenchymal transition (EMT) in lung adenocarcinoma epithelial cell line (A549). Cell. Immunol. 2016, 300, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Zhan, P.; Suo, L.; Qian, Q.; Shen, X.; Qiu, L.-X.; Yu, L.; Song, Y. Chlamydia pneumoniae infection and lung cancer risk: A meta-analysis. Eur. J. Cancer 2011, 47, 742–747. [Google Scholar] [CrossRef]
- Di Domenico, E.G.; Cavallo, I.; Pontone, M.; Toma, L.; Ensoli, F. Biofilm Producing Salmonella Typhi: Chronic Colonization and Development of Gallbladder Cancer. Int. J. Mol. Sci. 2017, 18, 1887. [Google Scholar] [CrossRef]
- Chattopadhyay, I.; Verma, M.; Panda, M. Role of Oral Microbiome Signatures in Diagnosis and Prognosis of Oral Cancer. Technol. Cancer Res. Treat. 2019, 18, 1533033819867354. [Google Scholar] [CrossRef] [Green Version]
- Perera, M.; Al-hebshi, N.N.; Speicher, D.J.; Perera, I.; Johnson, N.W. Emerging role of bacteria in oral carcinogenesis: A review with special reference to perio-Pathogenic bacteria. J. Oral Microbiol. 2016, 8, 32762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, J.A. Study Links Periodontal Disease Bacteria to Pancreatic Cancer Risk. JAMA 2016, 315, 2653–2654. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, M.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.; Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; et al. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-Cadherin/β-Catenin signaling via its FadA adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Wang, S.; Wang, H.; Tang, Y.-J.; Tang, Y.; Liang, X. Who is who in oral cancer? Exp. Cell Res. 2019, 384, 111634. [Google Scholar] [CrossRef] [PubMed]
- Chang, A.H.; Parsonnet, J. Role of bacteria in oncogenesis. Clin. Microbiol. Rev. 2010, 23, 837–857. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.-G.; Duan, G.-C.; Fan, Q.-T.; Chen, S.-Y. Role of Helicobacter pylori infection in pathogenesis of gastric carcinoma. World J. Gastrointest. Pathophysiol. 2016, 7, 97. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, V.; Memar, B.; Momtazi, A.A.; Sahebkar, A.; Gholamin, M.; Abbaszadegan, M.R. Cytokine networks and their association with Helicobacter pylori infection in gastric carcinoma. J. Cell. Physiol. 2018, 233, 2791–2803. [Google Scholar] [CrossRef]
- Vlăduţ, C.; Ciocîrlan, M.; Costache, R.S.; Jinga, M.; Balaban, V.D.; Costache, D.O.; Diculescu, M. Is mucosa-Associated lymphoid tissue lymphoma an infectious disease? Role of Helicobacter pylori and eradication antibiotic therapy (Review). Exp. Ther. Med. 2020, 20, 3546. [Google Scholar] [CrossRef]
- Bultman, S. Interplay between diet, gut microbiota, epigenetic events, and colorectal cancer. Mol. Nutr. Food Res. 2017, 61, 1500902. [Google Scholar] [CrossRef] [Green Version]
- Meng, C.; Bai, C.; Brown, T.D.; Hood, L.E.; Tian, Q. Human Gut Microbiota and Gastrointestinal Cancer. Genomics. Proteom. Bioinform. 2018, 16, 33–49. [Google Scholar] [CrossRef]
- Shang, F.-M.; Liu, H.-L. Fusobacterium nucleatum and colorectal cancer: A review. World J. Gastrointest. Oncol. 2018, 10, 71. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Guo, F.; Yu, Y.; Sun, T.; Ma, D.; Han, J.; Qian, Y.; Kryczek, I.; Sun, D.; Nagarsheth, N.; et al. Fusobacterium nucleatum Promotes Chemoresistance to Colorectal Cancer by Modulating Autophagy. Cell 2017, 170, 548. [Google Scholar] [CrossRef] [Green Version]
- Lucas, C.; Barnich, N.; Nguyen, H.T.T. Microbiota, Inflammation and Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 310. [Google Scholar] [CrossRef] [Green Version]
- Gagnière, J.; Raisch, J.; Veziant, J.; Barnich, N.; Bonnet, R.; Buc, E.; Bringer, M.-A.; Pezet, D.; Bonnet, M. Gut microbiota imbalance and colorectal cancer. World J. Gastroenterol. 2016, 22, 501. [Google Scholar] [CrossRef]
- Mesnard, B.; De Vroey, B.; Maunoury, V.; Lecuit, M. Immunoproliferative small intestinal disease associated with Campylobacter jejuni. Dig. Liver Dis. 2012, 44, 799–800. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Vuai, M.S.; Zhong, M. The role of bacteria in cancer therapy–Enemies in the past, but allies at present. Infect. Agent. Cancer 2018, 13, 9. [Google Scholar] [CrossRef]
- Ferreri, A.J.M.; Guidoboni, M.; Ponzoni, M.; De Conciliis, C.; Dell’Oro, S.; Fleischhauer, K.; Caggiari, L.; Lettini, A.A.; Dal Cin, E.; Ieri, R.; et al. Evidence for an Association Between Chlamydia psittaci and Ocular Adnexal Lymphomas. JNCI J. Natl. Cancer Inst. 2004, 96, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Abeni, C.; Rota, L.; Ogliosi, C.; Bertocchi, P.; Centurini, P.B.; Zaniboni, A. Correlation among Streptococcus bovis, endocarditis and septicemia in a patient with advanced colon cancer: A case report. J. Med. Case Reports 2013, 7, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Kovaleva, O.V.; Romashin, D.; Zborovskaya, I.B.; Davydov, M.M.; Shogenov, M.S.; Gratchev, A. Human lung microbiome on the way to cancer. Clin. Dev. Immunol. 2019, 2019, 1–6. [Google Scholar] [CrossRef]
- Jin, C.; Lagoudas, G.K.; Zhao, C.; Bullman, S.; Bhutkar, A.; Hu, B.; Ameh, S.; Sandel, D.; Liang, X.S.; Mazzilli, S.; et al. Commensal Microbiota Promote Lung Cancer Development via γδ T Cells. Cell 2019, 176, 998–1013.e16. [Google Scholar] [CrossRef] [Green Version]
- Greathouse, K.L.; White, J.R.; Vargas, A.J.; Bliskovsky, V.V.; Beck, J.A.; von Muhlinen, N.; Polley, E.C.; Bowman, E.D.; Khan, M.A.; Robles, A.I.; et al. Interaction between the microbiome and TP53 in human lung cancer. Genome Biol. 2018 191 2018, 19, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-X.; Tao, L.-L.; Zhang, J.; Zhu, Y.-G.; Zheng, Y.; Liu, D.; Zhou, M.; Ke, H.; Shi, M.-M.; Qu, J.-M. Difference of lower airway microbiome in bilateral protected specimen brush between lung cancer patients with unilateral lobar masses and control subjects. Int. J. Cancer 2018, 142, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Peters, B.A.; Hayes, R.B.; Goparaju, C.; Reid, C.; Pass, H.I.; Ahn, J. The microbiome in lung cancer tissue and recurrence-Free survival. Cancer Epidemiol. Biomarkers Prev. 2019, 28, 731. [Google Scholar] [CrossRef] [Green Version]
- Tsay, J.-C.J.; Wu, B.G.; Badri, M.H.; Clemente, J.C.; Shen, N.; Meyn, P.; Li, Y.; Yie, T.-A.; Lhakhang, T.; Olsen, E.; et al. Airway Microbiota Is Associated with Upregulation of the PI3K Pathway in Lung Cancer. Am. J. Respir. Crit. Care Med. 2018, 198, 1188. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Fang, Z.; Xue, Y.; Zhang, J.; Zhu, J.; Gao, R.; Yao, S.; Ye, Y.; Wang, S.; Lin, C.; et al. Specific gut microbiome signature predicts the early-stage lung cancer. Gut Microbes 2020, 11, 1030. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, A.K.; Gaydos, C.A.; Agreda, P.; Holden, J.P.; Chatterjee, N.; Goedert, J.J.; Caporaso, N.E.; Engels, E.A. Chlamydia pneumoniae Infection and Risk for Lung Cancer. Cancer Epidemiol. Prev. Biomarkers 2010, 19, 1498–1505. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; García Rodríguez, L.; Hernández-Díaz, S. Antibiotic use and the risk of lung cancer. Cancer Epidemiol. Biomarkers Prev. 2008, 17, 1308–1315. [Google Scholar] [CrossRef] [Green Version]
- Hosgood, H.D., III; Sapkota, A.R.; Rothman, N.; Rohan, T.; Hu, W.; Xu, J.; Vermeulen, R.; He, X.; White, J.R. The Potential Role of Lung Microbiota in Lung Cancer Attributed to Household Coal Burning Exposures. Environ. Mol. Mutagen. 2014, 55, 643. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Yang, M.; Liu, J.; Gao, R.; Hu, J.; Li, J.; Zhang, L.; Shi, Y.; Guo, H.; Cheng, J.; et al. Discovery and validation of potential bacterial biomarkers for lung cancer. Am. J. Cancer Res. 2015, 5, 3111. [Google Scholar]
- Lee, S.H.; Sung, J.Y.; Yong, D.; Chun, J.; Kim, S.Y.; Song, J.H.; Chung, K.S.; Kim, E.Y.; Jung, J.Y.; Kang, Y.A.; et al. Characterization of microbiome in bronchoalveolar lavage fluid of patients with lung cancer comparing with benign mass like lesions. Lung Cancer 2016, 102, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Gail, M.H.; Consonni, D.; Carugno, M.; Humphrys, M.; Pesatori, A.C.; Caporaso, N.E.; Goedert, J.J.; Ravel, J.; Landi, M.T. Characterizing human lung tissue microbiota and its relationship to epidemiological and clinical features. Genome Biol. 2016 171 2016, 17, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durack, J.; Lynch, S.V.; Nariya, S.; Bhakta, N.R.; Beigelman, A.; Castro, M.; Dyer, A.-M.; Israel, E.; Kraft, M.; Martin, R.J.; et al. Features of the bronchial bacterial microbiome associated with atopy, asthma and responsiveness to inhaled corticosteroid treatment. J. Allergy Clin. Immunol. 2017, 140, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Nuñez, M.; Millares, L.; Pomares, X.; Ferrari, R.; Pérez-Brocal, V.; Gallego, M.; Espasa, M.; Moya, A.; Monsó, E. Severity-Related Changes of Bronchial Microbiome in Chronic Obstructive Pulmonary Disease. J. Clin. Microbiol. 2014, 52, 4217. [Google Scholar] [CrossRef] [Green Version]
- Apopa, P.L.; Alley, L.; Penney, R.B.; Arnaoutakis, K.; Steliga, M.A.; Jeffus, S.; Bircan, E.; Gopalan, B.; Jin, J.; Patumcharoenpol, P.; et al. PARP1 Is Up-Regulated in Non-small Cell Lung Cancer Tissues in the Presence of the Cyanobacterial Toxin Microcystin. Front. Microbiol. 2018, 9, 1757. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; O’Brien, J.L.; Ajami, N.J.; Scheurer, M.E.; Amirian, E.S.; Armstrong, G.; Tsavachidis, S.; Thrift, A.P.; Jiao, L.; Wong, M.C.; et al. Lung tissue microbial profile in lung cancer is distinct from emphysema. Am. J. Cancer Res. 2018, 8, 1775. [Google Scholar]
- Yang, J.; Mu, X.; Wang, Y.; Zhu, D.; Zhang, J.; Liang, C.; Chen, B.; Wang, J.; Zhao, C.; Zuo, Z.; et al. Dysbiosis of the Salivary Microbiome Is Associated With Non-Smoking Female Lung Cancer and Correlated With Immunocytochemistry Markers. Front. Oncol. 2018, 8, 520. [Google Scholar] [CrossRef]
- Jin, J.; Gan, Y.; Liu, H.; Wang, Z.; Yuan, J.; Deng, T.; Zhou, Y.; Zhu, Y.; Zhu, H.; Yang, S.; et al. Diminishing microbiome richness and distinction in the lower respiratory tract of lung cancer patients: A multiple comparative study design with independent validation. Lung Cancer 2019, 136, 129–135. [Google Scholar] [CrossRef]
- Oelschlaeger, T.A. Bacteria as tumor therapeutics? Bioeng. Bugs 2010, 1, 146. [Google Scholar] [CrossRef] [Green Version]
- Weerakkody, L.R.; Witharana, C. The role of bacterial toxins and spores in cancer therapy. Life Sci. 2019, 235, 116839. [Google Scholar] [CrossRef]
- Patyar, S.; Joshi, R.; Byrav, D.P.; Prakash, A.; Medhi, B.; Das, B. Bacteria in cancer therapy: A novel experimental strategy. J. Biomed. Sci. 2010, 17, 21. [Google Scholar] [CrossRef] [Green Version]
- Kucerova, P.; Cervinkova, M. Spontaneous regression of tumour and the role of microbial infection–Possibilities for cancer treatment. Anticancer. Drugs 2016, 27, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef]
- Elinav, E.; Nowarski, R.; Thaiss, C.A.; Hu, B.; Jin, C.; Flavell, R.A. Inflammation-Induced cancer: Crosstalk between tumours, immune cells and microorganisms. Nat. Rev. Cancer 2013, 13, 759–771. [Google Scholar] [CrossRef]
- Dzutsev, A.; Badger, J.H.; Perez-Chanona, E.; Roy, S.; Salcedo, R.; Smith, C.K.; Trinchieri, G. Microbes and Cancer. Ann. Rev. Immunol. 2017, 35, 199–228. [Google Scholar] [CrossRef] [PubMed]
- Routy, B.; Le Chatelier, E.; Derosa, L.; Duong, C.P.M.; Alou, M.T.; Daillère, R.; Fluckiger, A.; Messaoudene, M.; Rauber, C.; Roberti, M.P.; et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science 2018, 359, 91–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Wang, S.; Zheng, B.; Qiu, X.; Wang, H.; Chen, L. Modulation of Gut Microbiota to Enhance Effect of Checkpoint Inhibitor Immunotherapy. Front. Immunol. 2021, 0, 2554. [Google Scholar] [CrossRef] [PubMed]
- Noci, V.L.; Guglielmetti, S.; Arioli, S.; Balsari, A.; Tagliabue, E.; Correspondence, L.S. Modulation of Pulmonary Microbiota by Antibiotic or Probiotic Aerosol Therapy: A Strategy to Promote Immunosurveillance against Lung Metastases. Cell Reports 2018, 24, 3528–3538. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; Kaur, S. Bacteriocins as Potential Anticancer Agents. Front. Pharmacol. 2015, 6, 272. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.H.; Dejea, C.M.; Edler, D.; Hoang, L.T.; Santidrian, A.F.; Felding, B.H.; Ivanisevic, J.; Cho, K.; Wick, E.C.; Hechenbleikner, E.M.; et al. Metabolism Links Bacterial Biofilms and Colon Carcinogenesis. Cell Metab. 2015, 21, 891–897. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Fu, G.-F.; Fan, Y.-R.; Liu, W.-H.; Liu, X.-J.; Wang, J.-J.; Xu, G.-X. Bifidobacterium adolescentis as a delivery system of endostatin for cancer gene therapy: Selective inhibitor of angiogenesis and hypoxic tumor growth. Cancer Gene Ther. 2003, 10, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.-F.; Zhu, L.-P.; Hu, B.; Fu, G.-F.; Zhang, H.-Y.; Wang, J.-J.; Xu, G.-X. A new expression plasmid in Bifidobacterium longum as a delivery system of endostatin for cancer gene therapy. Cancer Gene Ther. 2006, 14, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.K.; Liu, Q.; Liang, K.; Li, P.; Kong, Q. Bacteria-Derived minicells for cancer therapy. Cancer Lett. 2020, 491, 11–21. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Choy, H.E.; Lee, J.-H.; Kim, G.-J. Generation of Minicells from an Endotoxin-Free Gram-Positive Strain Corynebacterium glutamicum. J. Microbiol. Biotechnol. 2015, 25, 554–558. [Google Scholar] [CrossRef] [Green Version]
- Whittle, J.R.; Lickliter, J.D.; Gan, H.K.; Scott, A.M.; Simes, J.; Solomon, B.J.; MacDiarmid, J.A.; Brahmbhatt, H.; Rosenthal, M.A. First in human nanotechnology doxorubicin delivery system to target epidermal growth factor receptors in recurrent glioblastoma. J. Clin. Neurosci. 2015, 22, 1889–1894. [Google Scholar] [CrossRef] [PubMed]
- Sedighi, M.; Bialvaei, A.Z.; Hamblin, M.R.; Ohadi, E.; Asadi, A.; Halajzadeh, M.; Lohrasbi, V.; Mohammadzadeh, N.; Amiriani, T.; Krutova, M.; et al. Therapeutic bacteria to combat cancer; current advances, challenges, and opportunities. Cancer Med. 2019, 8, 3167. [Google Scholar] [CrossRef]
- Sharma, P.C.; Sharma, D.; Sharma, A.; Bhagat, M.; Ola, M.; Thakur, V.K.; Bhardwaj, J.K.; Goyal, R.K. Recent advances in microbial toxin-Related strategies to combat cancer. Semin. Cancer Biol. 2021, in press. [Google Scholar] [CrossRef]
- Fuchs, H.; Bachran, C. Targeted Tumor Therapies at a Glance. Curr. Drug Targets 2009, 10, 89–93. [Google Scholar] [CrossRef]
- Akbari, B.; Farajnia, S.; Khosroshahi, S.A.; Safari, F.; Yousefi, M.; Dariushnejad, H.; Rahbarnia, L. Immunotoxins in cancer therapy: Review and update. Int. Rev. Immunol. 2017, 36, 207–219. [Google Scholar] [CrossRef]
- Zahaf, N.-I.; Schmidt, G. Bacterial Toxins for Cancer Therapy. Toxins 2017, 9, 236. [Google Scholar] [CrossRef] [Green Version]
- Pahle, J.; Walther, W. Bacterial Toxins for Oncoleaking Suicidal Cancer Gene Therapy. Recent Results Cancer Res. 2016, 209, 95–110. [Google Scholar] [CrossRef]
- Hasenpusch, G.; Pfeifer, C.; Aneja, M.K.; Wagner, K.; Reinhardt, D.; Gilon, M.; Ohana, P.; Hochberg, A.; Rudolph, C. Aerosolized BC-819 Inhibits Primary but Not Secondary Lung Cancer Growth. PLoS ONE 2011, 6, e20760. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; McClane, B.A. Use of Clostridium perfringens enterotoxin and the enterotoxin receptor-Binding domain (C-CPE) for cancer treatment: Opportunities and challenges. J. Toxicol. 2012, 2012, 981626. [Google Scholar] [CrossRef] [Green Version]
- Zielinski, R.; Lyakhov, I.; Hassan, M.; Kuban, M.; Shafer-Weaver, K.; Gandjbakhche, A.; Capala, J. HER2-Affitoxin: A Potent Therapeutic Agent for the Treatment of HER2-Overexpressing Tumors. Clin. Cancer Res. 2011, 17, 5071–5081. [Google Scholar] [CrossRef] [Green Version]
- Hashimi, S.M.; Grant, B.; Alqurashi, N.; Alowaidi, F.; Wei, M.Q. EGF ligand fused to truncated Pseudomonas aeruginosa exotoxin A specifically targets and inhibits EGFR-Positive cancer cells. Oncol. Rep. 2018, 40, 2690–2697. [Google Scholar] [CrossRef] [Green Version]
- Gholami, N.; Cohan, R.A.; Razavi, A.; Bigdeli, R.; Dashbolaghi, A.; Asgary, V. Cytotoxic and apoptotic properties of a novel nano-Toxin formulation based on biologically synthesized silver nanoparticle loaded with recombinant truncated pseudomonas exotoxin A. J. Cell. Physiol. 2020, 235, 3711–3720. [Google Scholar] [CrossRef]
- Yang, W.S.; Park, S.-O.; Yoon, A.-R.; Yoo, J.Y.; Kim, M.K.; Yun, C.-O.; Kim, C.-W. Suicide cancer gene therapy using pore-forming toxin, streptolysin O. Mol. Cancer Ther. 2006, 5, 1610–1619. [Google Scholar] [CrossRef] [Green Version]
- Telang, S.; Rasku, M.A.; Clem, A.L.; Carter, K.; Klarer, A.C.; Badger, W.R.; Milam, R.A.; Rai, S.N.; Pan, J.; Gragg, H.; et al. Phase II trial of the regulatory T cell-Depleting agent, denileukin diftitox, in patients with unresectable stage IV melanoma. BMC Cancer 2011, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Talpur, R.; Duvic, M. Pilot Study of Denileukin Diftitox Alternate Dosing Regimen in Patients With Cutaneous Peripheral T-Cell Lymphomas. Clin. Lymphoma Myeloma Leuk. 2012, 12, 180–185. [Google Scholar] [CrossRef]
- LeMaistre, C.; Meneghetti, C.; Rosenblum, M.; Reuben, J.; Parker, K.; Shaw, J.; Deisseroth, A.; Woodworth, T.; Parkinson, D. Phase I Trial of an Interleukin-2 (IL-2) Fusion Toxin (DAB486IL-2) in Hematologic Malignancies Expressing the IL-2 Receptor. Blood 1992, 79, 2547–2554. [Google Scholar] [CrossRef] [Green Version]
- Platanias, L.C.; Ratain, M.J.; O’brien, S.; Larson, R.A.; Vardiman, J.W.; Shaw, J.P.; Williams, S.F.; Baron, J.M.; Parker, K.; Woodworth, T.G. Phase I Trial of a Genetically Engineered Interleukin-2 Fusion Toxin (DAB486IL-2) as a 6 Hour Intravenous Infusion in Patients with Hematologic Malignancies. Leuk. Lymphoma 2009, 14, 257–262. [Google Scholar] [CrossRef]
- Testa, U.; Pelosi, E.; Castelli, G. CD123 as a Therapeutic Target in the Treatment of Hematological Malignancies. Cancers 2019, 11, 1358. [Google Scholar] [CrossRef] [Green Version]
- Kreitman, R.J. Immunotoxins for targeted cancer therapy. AAPS J. 2006, 8, E532–E551. [Google Scholar] [CrossRef]
- Karpiński, T.M.; Adamczak, A. Anticancer activity of bacterial proteins and peptides. Pharmaceutics 2018, 10, 54. [Google Scholar] [CrossRef] [Green Version]
- Duport, C.; Alpha-Bazin, B.; Armengaud, J. Advanced proteomics as a powerful tool for studying toxins of human bacterial pathogens. Toxins 2019, 11, 576. [Google Scholar] [CrossRef] [Green Version]
- Kull, S.; Schulz, K.M.; née Strotmeier, J.W.; Kirchner, S.; Schreiber, T.; Bollenbach, A.; Dabrowski, P.W.; Nitsche, A.; Kalb, S.R.; Dorner, M.B.; et al. Isolation and Functional Characterization of the Novel Clostridium botulinum Neurotoxin A8 Subtype. PLoS ONE 2015, 10, e0116381. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.J.; Hill, K.K.; Xie, G.; Foley, B.T.; Williamson, C.H.D.; Foster, J.T.; Johnson, S.L.; Chertkov, O.; Teshima, H.; Gibbons, H.S.; et al. Genomic sequences of six botulinum neurotoxin-Producing strains representing three clostridial species illustrate the mobility and diversity of botulinum neurotoxin genes. Infect. Genet. Evol. 2015, 30, 102–113. [Google Scholar] [CrossRef] [Green Version]
- Hill, K.K.; Smith, T.J. Genetic Diversity Within Clostridium botulinum Serotypes, Botulinum Neurotoxin Gene Clusters and Toxin Subtypes. In Botulinum Neurotoxins. Current Topics in Microbiology and Immunology; Rummel, A., Binz, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 364, pp. 1–20. [Google Scholar]
- Josić, D.; Peršurić, Ž.; Rešetar, D.; Martinović, T.; Saftić, L.; Kraljević Pavelić, S. Use of Foodomics for Control of Food Processing and Assessing of Food Safety. Adv. Food Nutr. Res. 2017, 81, 187–229. [Google Scholar] [CrossRef]
- Kalb, S.R.; Boyer, A.E.; Barr, J.R. Mass spectrometric detection of bacterial protein toxins and their enzymatic activity. Toxins 2015, 7, 3497–3511. [Google Scholar] [CrossRef] [Green Version]
- Morineaux, V.; Mazuet, C.; Hilaire, D.; Enche, J.; Popoff, M.R. Characterization of botulinum neurotoxin type A subtypes by immunocapture enrichment and liquid chromatography–Tandem mass spectrometry. Anal. Bioanal. Chem. 2015, 407, 5559–5570. [Google Scholar] [CrossRef]
- Malet, J.K.; Impens, F.; Carvalho, F.; Hamon, M.A.; Cossart, P.; Ribet, D. Rapid remodeling of the host epithelial cell proteome by the listeriolysin O (LLO) pore-Forming toxin. Mol. Cell. Proteomics 2018, 17, 1627–1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Lin, S.; De Mel, N.; Parupudi, A.; Delmar, J.; Pandey, M.; Wang, X.; Wang, J. Deamidation in Moxetumomab Pasudotox Leading to Conformational Change and Immunotoxin Activity Loss. J. Pharm. Sci. 2020, 109, 2676–2683. [Google Scholar] [CrossRef]
- Bachran, C.; Gupta, P.K.; Bachran, S.; Leysath, C.E.; Hoover, B.; Fattah, R.J.; Leppla, S.H. Reductive Methylation and Mutation of an Anthrax Toxin Fusion Protein Modulates its Stability and Cytotoxicity. Sci. Reports 2014, 4, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Moss, D.L.; Park, H.-W.; Mettu, R.R.; Landry, S.J. Deimmunizing substitutions in Pseudomonas exotoxin domain III perturb antigen processing without eliminating T-cell epitopes. J. Biol. Chem. 2019, 294, 4667. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.-C.; Yu, W.-W.; Qi, Y.-J.; Xu, L.-F.; Wang, Z.-R.; Qiang, Y.-W.; Ma, F.; Ma, X.-L. Quantitative proteomic analysis reveals that Luks-PV exerts antitumor activity by regulating the key proteins and metabolic pathways in HepG2 cells. Anticancer. Drugs 2020, 31, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Srinivas, P.R.; Srivastava, S.; Hanash, S.; Wright, G.L. Proteomics in Early Detection of Cancer. Clin. Chem. 2001, 47, 1901–1911. [Google Scholar] [CrossRef] [Green Version]
- Amiri-Dashatan, N.; Koushki, M.; Abbaszadeh, H.A.; Rostami-Nejad, M.; Rezaei-Tavirani, M. Proteomics applications in health: Biomarker and drug discovery and food industry. Iran. J. Pharm. Res. 2018, 17, 1523–1536. [Google Scholar] [CrossRef] [PubMed]
- Begić, M.; Josić, D. Biofilm formation and extracellular microvesicles—The way of foodborne pathogens toward resistance. Electrophoresis 2020, 41, 1718–1739. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhou, H.; Yang, C.; Wu, Y.; Zhou, X.; Liu, H.; Wang, Y. Bacterial outer membrane vesicles as a platform for biomedical applications: An update. J. Control. Release 2020, 323, 253–268. [Google Scholar] [CrossRef]
- Kim, O.Y.; Park, H.T.; Dinh, N.T.H.; Choi, S.J.; Lee, J.; Kim, J.H.; Lee, S.-W.; Gho, Y.S. Bacterial outer membrane vesicles suppress tumor by interferon-γ-mediated antitumor response. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Chronopoulos, A.; Kalluri, R. Emerging role of bacterial extracellular vesicles in cancer. Oncogene 2020, 39, 6951–6960. [Google Scholar] [CrossRef]
- Palomino, R.A.Ñ.; Vanpouille, C.; Costantini, P.E.; Margolis, L. Microbiota–host communications: Bacterial extracellular vesicles as a common language. PLoS Pathog. 2021, 17, e1009508. [Google Scholar] [CrossRef]
- Toyofuku, M.; Nomura, N.; Eberl, L. Types and origins of bacterial membrane vesicles. Nat. Rev. Microbiol. 2019, 17, 13–24. [Google Scholar] [CrossRef]
- Lee, J.; Kim, O.; Gho, Y. Proteomic profiling of Gram-negative bacterial outer membrane vesicles: Current perspectives. Proteomics. Clin. Appl. 2016, 10, 897–909. [Google Scholar] [CrossRef]
- Hong, J.; Dauros-Singorenko, P.; Whitcombe, A.; Payne, L.; Blenkiron, C.; Phillips, A.; Swift, S. Analysis of the Escherichia coli extracellular vesicle proteome identifies markers of purity and culture conditions. J. Extracell. Vesicles 2019, 8, 1632099. [Google Scholar] [CrossRef] [Green Version]
- Gujrati, V.; Kim, S.; Kim, S.-H.; Min, J.J.; Choy, H.E.; Kim, S.C.; Jon, S. Bioengineered Bacterial Outer Membrane Vesicles as Cell-Specific Drug-Delivery Vehicles for Cancer Therapy. ACS Nano 2014, 8, 1525–1537. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.; Choi, S.; Jang, S.; Park, K.; Kim, S.; Choi, J.; Lim, J.; Lee, S.; Park, J.; Di Vizio, D.; et al. Bacterial protoplast-Derived nanovesicles as vaccine delivery system against bacterial infection. Nano Lett. 2015, 15, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fang, Z.; Li, R.; Huang, X.; Liu, Q. Design of Outer Membrane Vesicles as Cancer Vaccines: A New Toolkit for Cancer Therapy. Cancers 2019, 11, 1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuerban, K.; Gao, X.; Zhang, H.; Liu, J.; Dong, M.; Wu, L.; Ye, R.; Feng, M.; Ye, L. Doxorubicin-Loaded bacterial outer-membrane vesicles exert enhanced anti-tumor efficacy in non-Small-Cell lung cancer. Acta Pharm. Sin. B 2020, 10, 1534–1548. [Google Scholar] [CrossRef]
- Gnopo, Y.; Watkins, H.; Stevenson, T.; DeLisa, M.; Putnam, D. Designer outer membrane vesicles as immunomodulatory systems*Reprogramming bacteria for vaccine delivery. Adv. Drug Deliv. Rev. 2017, 114, 132–142. [Google Scholar] [CrossRef]
- Wang, S.; Huang, W.; Li, K.; Yao, Y.; Yang, X.; Bai, H.; Sun, W.; Liu, C.; Ma, Y. Engineered outer membrane vesicle is potent to elicit HPV16E7-specific cellular immunity in a mouse model of TC-1 graft tumor. Int. J. Nanomedicine 2017, 12, 6813–6825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhao, R.; Cheng, K.; Zhang, K.; Wang, Y.; Zhang, Y.; Li, Y.; Liu, G.; Xu, J.; Xu, J.; et al. Bacterial Outer Membrane Vesicles Presenting Programmed Death 1 for Improved Cancer Immunotherapy via Immune Activation and Checkpoint Inhibition. ACS Nano 2020, 14, 16698–16711. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Q.; Xu, J.; Deng, D.; Chao, T.; Li, J.; Zhang, R.; Peng, R.; Liu, Z. Bacteria-Derived membrane vesicles to advance targeted photothermal tumor ablation. Biomaterials 2021, 268, 120550. [Google Scholar] [CrossRef] [PubMed]


| Bacteria | Cancer | Reference |
|---|---|---|
| Mycobacterium tuberculosis | Lung cancer | [11,12] |
| Chlamidia pneumonia | Lung cancer | [13] |
| Salmonella enterica, subsp typhi | Gallbladder cancer | [14] |
| Prrphyromonas gingivalis | Oral cancer Pancreatic cancer | [15,16,17] |
| Fusobacterium nucleatum | Oral cancer | [15,16,18] |
| Treponema denticola | Oral cancer | [19] |
| Streptococcus anginosus | Oral cancer | [19] |
| Helicobacter pylori | Gastric cancer, Mucosa-associated lymphoid tissue lymphoma | [20,21,22,23] |
| Bacteroides fragilis | Colon cancer | [24,25] |
| Fusobacterium nucleatum | Colon cancer | [26,27,28] |
| E.coli | Colon cancer | [29] |
| Campylobacter jejuni | Small intestinal lymphoma | [30] |
| Citrobacter rodentium | Colon cancer | [31] |
| Chlamydia psittaci | Cervical cancer, ocular lymphoma | [32] |
| Citrobacter rodentium | Colorectal cancer | [31] |
| Streptococcus bovis | Colorectal neoplasia | [33] |
| Sample Type | Toxin | Purpose of the Study | Mass Spectrometry (MS) Technique Applied in the Study | Reference |
|---|---|---|---|---|
| Crude culture supernatants, biological and food samples artificially spiked with culture supernatant of each C. botulinum A subtype | BoNT | Characterization of BoNT type A subtypes | HPLC-ESI-IT HPLC-QqQ | [95] |
| Cell cultures incubated with LLO | LLO | Identification of host proteome alterations induced by the LLO | HPLC-LTQ Orbitrap RSLCnano-Q Exactive high-field hybrid Quadrupole-Orbitrap | [96] |
| Moxetumomab pasudotox | PE (PE38) | Examination of structural and biological impact of deamidation | Hydrogen-deuterium exchange MS | [97] |
| Anthrax toxin fusion protein | PE | Examination of modifications of the therapeutic fusion protein that were predicted to improve its potency on target cells | ESI-MS | [98] |
| Exotoxin A domain III | PE | Identification how deimmunizing mutations alter structure, processing and immunogenicity of PE-III | NanoLC-Orbitrap Fusion MS | [99] |
| Cell lines HepG2 treated with LukS-PV | PVL | Identification of differentially expressed proteins to better understand antitumor activity of LukS-PV | NanoUPLC-QTOF | [100] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trivanović, D.; Pavelić, K.; Peršurić, Ž. Fighting Cancer with Bacteria and Their Toxins. Int. J. Mol. Sci. 2021, 22, 12980. https://doi.org/10.3390/ijms222312980
Trivanović D, Pavelić K, Peršurić Ž. Fighting Cancer with Bacteria and Their Toxins. International Journal of Molecular Sciences. 2021; 22(23):12980. https://doi.org/10.3390/ijms222312980
Chicago/Turabian StyleTrivanović, Dragan, Krešimir Pavelić, and Željka Peršurić. 2021. "Fighting Cancer with Bacteria and Their Toxins" International Journal of Molecular Sciences 22, no. 23: 12980. https://doi.org/10.3390/ijms222312980
APA StyleTrivanović, D., Pavelić, K., & Peršurić, Ž. (2021). Fighting Cancer with Bacteria and Their Toxins. International Journal of Molecular Sciences, 22(23), 12980. https://doi.org/10.3390/ijms222312980