
Gut Microbiota as a Mediator of Essential and Toxic Effects of Zinc in the Intestines and Other Tissues


 ,
,  ,
,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
- The impact of Zn on characteristics of gut microbiota in various organisms.
- Chicks
- Piglets
- Laboratory rodents
- Humans
- The influence of Zn on gut microbiota upon exposure to toxic and infectious agents.
- The potential role of Zn-induced microbiota in modulating systemic effects with a focus on extraintestinal diseases.
- Interactive effects of Zn and probiotics on gut microbiota.
2. Relationship between Zn Status and Gut Microbiota
2.1. Poultry
2.2. Pigs
2.3. Laboratory Rodents
2.4. Human
2.5. Summary

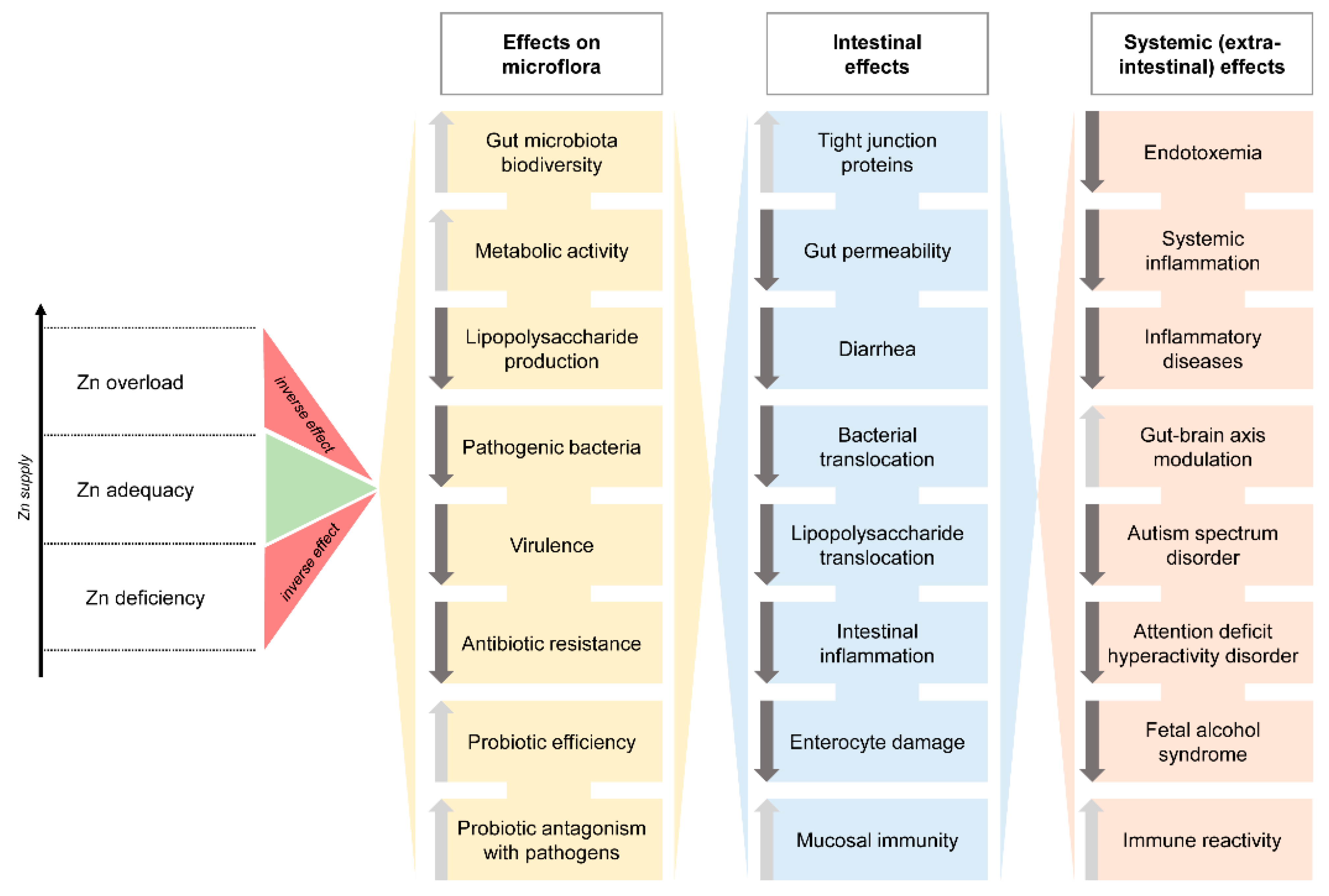{kind=link}
| Species | Zn Form | Dose | Microbiota Biodiversity | Reduced Taxa | Increased Taxa | Ref. |
|---|---|---|---|---|---|---|
| Broilers | Zn bacitracin | 50 ppm Zn | Increased | Lactobacillus Eubacterium | Clostridiales Faecalibacterium | [24] |
| Broilers | Zn hydroxychloride | 20–100 mg Zn/kg Zn | Decreased | Bacillus | Lactobacillus | [25] |
| Piglets | Zn oxide | 2250 mg Zn/kg | Decreased | Lactobacillus E. coli (at high doses) | E. coli (at low doses) | [21] |
| Piglets | Zn oxide | 3042 mg Zn/kg (high dose) | Increased | L. reuteri | Enterobacteriaceae W. cibaria W. confuse Leuconostoc citreum S. equinus | [33] |
| Piglets | Zn oxide NPs | 600–2000 mg Zn/kg | Decreased (ileum) Increased (cecum, colon) | Lactobacillus (ileum) Oscillospira, Prevotella (cecum, colon) | Streptococcus (ileum) Lactobacillus (cecum, colon) | [31] |
| Piglets | Coated nano ZnO | 0.100 g Zn/kg diet | Increased | R. flavefaciens | Lachnospiraceae | [34] |
| Mice | Zn chloride | 12–250 mg/kg b.w. | No effect | Lactobacillaceae Enterobacteriaceae | Clostridiacea | [42] |
| Mice | Zn sulfate | 100 Zn µg/d (high dose) | Increased | Pseudomonodales Enterobacteriacae Clostridiales Bacteroides Campylobacter | [45] |
3. Zn and Microbiota upon Exposure to Toxic and Infectious Agents
4. Extraintestinal Effects in Models of Human Diseases
5. Probiotics
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
References
- Maret, W. Zinc biochemistry: From a single zinc enzyme to a key element of life. Adv. Nutr. 2013, 4, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Skalny, A.V.; Aschner, M.; Tinkov, A.A. Zinc. Adv. Food Nutr. Res. 2021, 96, 251–310. [Google Scholar] [CrossRef] [PubMed]
- Chasapis, C.T.; Ntoupa, P.A.; Spiliopoulou, C.A.; Stefanidou, M.E. Recent aspects of the effects of zinc on human health. Arch. Toxicol. 2020, 94, 1443–1460. [Google Scholar] [CrossRef] [PubMed]
- Cerasi, M.; Ammendola, S.; Battistoni, A. Competition for zinc binding in the host-pathogen interaction. Front. Cell. Infect. Microbiol. 2013, 3, 108. [Google Scholar] [CrossRef]
- Liuzzi, J.P.; Lichten, L.A.; Rivera, S.; Blanchard, R.K.; Aydemir, T.B.; Knutson, M.D.; Ganz, T.; Cousins, R.J. Interleukin-6 regulates the zinc transporter Zip14 in liver and contributes to the hypozincemia of the acute-phase response. Proc. Natl. Acad. Sci. USA 2005, 102, 6843–6848. [Google Scholar] [CrossRef] [Green Version]
- Zackular, J.P.; Chazin, W.J.; Skaar, E.P. Nutritional immunity: S100 proteins at the host-pathogen interface. J. Biol. Chem. 2015, 290, 18991–18998. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.T.; Karim, M.M. Metallothionein: A Potential Link in the Regulation of Zinc in Nutritional Immunity. Biol. Trace Elem. Res. 2018, 182, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ammendola, S.; Pasquali, P.; Pistoia, C.; Petrucci, P.; Petrarca, P.; Rotilio, G.; Battistoni, A. High-affinity Zn2+ uptake system ZnuABC is required for bacterial zinc homeostasis in intracellular environments and contributes to the virulence of Salmonella enterica. Infect. Immun. 2007, 75, 5867–5876. [Google Scholar] [CrossRef] [Green Version]
- McDevitt, C.A.; Ogunniyi, A.D.; Valkov, E.; Lawrence, M.C.; Kobe, B.; McEwan, A.G.; Paton, J.C. A molecular mechanism for bacterial susceptibility to zinc. PLoS Pathog. 2011, 7, e1002357. [Google Scholar] [CrossRef] [Green Version]
- Raghupathi, K.R.; Koodali, R.T.; Manna, A.C. Size-dependent bacterial growth inhibition and mechanism of antibacterial activity of zinc oxide nanoparticles. Langmuir 2011, 27, 4020–4028. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Labrie, J.; Tremblay, Y.D.; Haine, D.; Mourez, M.; Jacques, M. Zinc as an agent for the prevention of biofilm formation by pathogenic bacteria. J. Appl. Microbiol. 2013, 115, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Shreiner, A.B.; Kao, J.Y.; Young, V.B. The gut microbiome in health and in disease. Curr. Opin. Gastroenterol. 2015, 31, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Hase, K. Gut microbiota-generated metabolites in animal health and disease. Nat. Chem. Biol. 2014, 10, 416–424. [Google Scholar] [CrossRef]
- Baquero, F.; Nombela, C. The microbiome as a human organ. Clin. Microbiol. Infect. 2012, 18, 2–4. [Google Scholar] [CrossRef] [Green Version]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Clarke, G.; Stilling, R.M.; Kennedy, P.J.; Stanton, C.; Cryan, J.F.; Dinan, T.G. Minireview: Gut microbiota: The neglected endocrine organ. Mol. Endocrinol. 2014, 28, 1221–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.; Yun, C.; Pang, Y.; Qiao, J. The impact of the gut microbiota on the reproductive and metabolic endocrine system. Gut Microbes 2021, 13, 1–21. [Google Scholar] [CrossRef]
- Du Toit, A. The gut microbiome and mental health. Nat. Rev. Microbiol. 2019, 17, 196. [Google Scholar] [CrossRef] [PubMed]
- Southon, S.; Gee, J.M.; Bayliss, C.E.; Wyatt, G.M.; Horn, N.; Johnson, I.T. Intestinal microflora, morphology and enzyme activity in zinc-deficient and Zn-supplemented rats. Br. J. Nutr. 1986, 55, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Koren, O.; Tako, E. Chronic Dietary Zinc Deficiency Alters Gut Microbiota Composition and Function. Proceedings 2020, 61, 16. [Google Scholar]
- Shen, J.; Chen, Y.; Wang, Z.; Zhou, A.; He, M.; Mao, L.; Zou, H.; Peng, Q.; Xue, B.; Wang, L.; et al. Coated zinc oxide improves intestinal immunity function and regulates microbiota composition in weaned piglets. Br. J. Nutr. 2014, 111, 2123–2134. [Google Scholar] [CrossRef]
- Zhang, T.; Zhu, X.; Guo, J.; Gu, A.Z.; Li, D.; Chen, J. Toxicity Assessment of Nano-ZnO Exposure on the Human Intestinal Microbiome, Metabolic Functions, and Resistome Using an In Vitro Colon Simulator. Environ. Sci. Technol. 2021, 55, 6884–6896. [Google Scholar] [CrossRef] [PubMed]
- Durrani, M.; Nazli, R.; Sher, N.; Abubakr, M.; Ali, J. Gut microbiome profile in zinc deficient infants using next generation sequencing. Khyber Med Univ. J. 2021, 13, 4–9. [Google Scholar]
- Li, D.; Achkar, J.P.; Haritunians, T.; Jacobs, J.P.; Hui, K.Y.; D’Amato, M.; Brand, S.; Radford-Smith, G.; Halfvarson, J.; Niess, J.H.; et al. A Pleiotropic Missense Variant in SLC39A8 Is Associated with Crohn’s Disease and Human Gut Microbiome Composition. Gastroenterology 2016, 151, 724–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collij, V.; Imhann, F.; Vich Vila, A.; Fu, J.; Dijkstra, G.; Festen, E.; Voskuil, M.D.; Daly, M.J.; Xavier, R.J.; Wijmenga, C.; et al. SLC39A8 missense variant is associated with Crohn’s disease but does not have a major impact on gut microbiome composition in healthy subjects. PLoS ONE 2019, 14, e0211328. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Zhao, W.J.; Zheng, W.J.; Mao, L.; Lian, H.Z.; Hu, X.; Hua, Z.C. Effects of Different Zinc Species on Cellar Zinc Distribution, Cell Cycle, Apoptosis and Viability in MDAMB231 Cells. Biol. Trace Elem. Res. 2016, 170, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Reed, S.; Neuman, H.; Moscovich, S.; Glahn, R.P.; Koren, O.; Tako, E. Chronic Zinc Deficiency Alters Chick Gut Microbiota Composition and Function. Nutrients 2015, 7, 9768–9784. [Google Scholar] [CrossRef] [PubMed]
- Reed, S.; Knez, M.; Uzan, A.; Stangoulis, J.; Glahn, R.P.; Koren, O.; Tako, E. Alterations in the Gut (Gallus gallus) Microbiota Following the Consumption of Zinc Biofortified Wheat (Triticum aestivum)-Based Diet. J. Agric. Food Chem. 2018, 66, 6291–6299. [Google Scholar] [CrossRef] [PubMed]
- Crisol-Martínez, E.; Stanley, D.; Geier, M.S.; Hughes, R.J.; Moore, R.J. Understanding the mechanisms of zinc bacitracin and avilamycin on animal production: Linking gut microbiota and growth performance in chickens. Appl. Microbiol. Biotechnol. 2017, 101, 4547. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.T.; Morgan, N.; Roberts, J.R.; Wu, S.B.; Swick, R.A.; Toghyani, M. Zinc hydroxychloride supplementation improves tibia bone development and intestinal health of broiler chickens. Poult. Sci. 2021, 100, 101254. [Google Scholar] [CrossRef]
- Fatholahi, A.; Khalaji, S.; Hosseini, F.; Abbasi, M. Nano-Bio zinc synthesized by Bacillus subtilis modulates broiler performance, intestinal morphology and expression of tight junction’s proteins. Livest. Sci. 2021, 251, 104660. [Google Scholar] [CrossRef]
- Gielda, L.M.; DiRita, V.J. Zinc competition among the intestinal microbiota. MBio 2012, 3, e00171-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhouma, M.; Fairbrother, J.M.; Beaudry, F.; Letellier, A. Post weanling diarrhea in pigs: Risk factors and non-colistin-based control strategies. Acta Vet. Scand. 2017, 59, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.H.; Xiao, K.; Song, J.; Luan, Z.S. Effects of zinc oxide supported on zeolite on growth performance, intestinal microflora and permeability, and cytokines expression of weaned pigs. Anim. Feed Sci. Technol. 2013, 181, 65–71. [Google Scholar] [CrossRef]
- Wang, W.; Van Noten, N.; Degroote, J.; Romeo, A.; Vermeir, P.; Michiels, J. Effect of zinc oxide sources and dosages on gut microbiota and integrity of weaned piglets. J. Anim. Physiol. Anim. Nutr. 2019, 103, 231–241. [Google Scholar] [CrossRef]
- Xia, T.; Lai, W.; Han, M.; Han, M.; Ma, X.; Zhang, L. Dietary ZnO nanoparticles alters intestinal microbiota and inflammation response in weaned piglets. Oncotarget 2017, 8, 64878–64891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starke, I.C.; Pieper, R.; Neumann, K.; Zentek, J.; Vahjen, W. The impact of high dietary zinc oxide on the development of the intestinal microbiota in weaned piglets. FEMS Microbiol. Ecol. 2014, 87, 416–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vahjen, W.; Pieper, R.; Zentek, J. Increased dietary zinc oxide changes the bacterial core and enterobacterial composition in the ileum of piglets. J. Anim. Sci. 2011, 89, 2430–2439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Bai, M.; Xu, K.; Zhou, J.; Zhang, X.; Yu, R.; Huang, R.; Yin, Y. Effects of different concentrations of coated nano zinc oxide material on fecal bacterial composition and intestinal barrier in weaned piglets. J. Sci. Food Agric. 2021, 101, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Caine, W.R.; Metzler-Zebeli, B.U.; McFall, M.; Miller, B.; Ward, T.L.; Kirkwood, R.N.; Mosenthin, R. Supplementation of diets for gestating sows with zinc amino acid complex and gastric intubation of suckling pigs with zinc-methionine on mineral status, intestinal morphology and bacterial translocation in lipopolysaccharide-challenged early weaned pigs. Res. Vet. Sci. 2009, 86, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Broom, L.J.; Miller, H.M.; Kerr, K.G.; Knapp, J.S. Effects of zinc oxide and Enterococcus faecium SF68 dietary supplementation on the performance, intestinal microbiota and immune status of weaned piglets. Res. Vet. Sci. 2006, 80, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Pieper, R.; Vahjen, W.; Neumann, K.; Van Kessel, A.G.; Zentek, J. Dose-dependent effects of dietary zinc oxide on bacterial communities and metabolic profiles in the ileum of weaned pigs. J. Anim. Physiol. Anim. Nutr. 2012, 96, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Urriola, P.E.; Luo, Z.H.; Rambo, Z.J.; Wilson, M.E.; Torrison, J.L.; Shurson, G.C.; Chen, C. Metabolomics revealed diurnal heat stress and zinc supplementation-induced changes in amino acid, lipid, and microbial metabolism. Physiol. Rep. 2016, 4, e12676. [Google Scholar] [CrossRef] [PubMed]
- Sauer, A.K.; Grabrucker, A.M. Zinc Deficiency during Pregnancy Leads to Altered Microbiome and Elevated Inflammatory Markers in Mice. Front. Neurosci. 2019, 13, 1295. [Google Scholar] [CrossRef]
- Kable, M.E.; Riazati, N.; Kirschke, C.P.; Zhao, J.; Tepaamorndech, S.; Huang, L. The Znt7-null mutation has sex dependent effects on the gut microbiota and goblet cell population in the mouse colon. PLoS ONE 2020, 15, e0239681. [Google Scholar] [CrossRef] [PubMed]
- Mayneris-Perxachs, J.; Bolick, D.T.; Leng, J.; Medlock, G.L.; Kolling, G.L.; Papin, J.A.; Swann, J.R.; Guerrant, R.L. Protein- and zinc-deficient diets modulate the murine microbiome and metabolic phenotype. Am. J. Clin. Nutr. 2016, 104, 1253–1262. [Google Scholar] [CrossRef]
- Foligné, B.; George, F.; Standaert, A.; Garat, A.; Poiret, S.; Peucelle, V.; Ferreira, S.; Sobry, H.; Muharram, G.; Lucau-Danila, A.; et al. High-dose dietary supplementation with zinc prevents gut inflammation: Investigation of the role of metallothioneins and beyond by transcriptomic and metagenomic studies. FASEB J. 2020, 34, 12615–12633. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.; Wong, C.; Bouranis, J.; Sharpton, T.; Ho, E. Zinc Status Elicits Age-Dependent Effects in the Gut Microbiome. Curr. Dev. Nutr. 2020, 4 (Suppl. S2), 1552. [Google Scholar] [CrossRef]
- Plum, L.M.; Rink, L.; Haase, H. The essential toxin: Impact of zinc on human health. Int. J. Environ. Res. Public Health 2010, 7, 1342–1365. [Google Scholar] [CrossRef] [Green Version]
- Podany, A.; Rauchut, J.; Wu, T.; Kawasawa, Y.I.; Wright, J.; Lamendella, R.; Soybel, D.I.; Kelleher, S.L. Excess Dietary Zinc Intake in Neonatal Mice Causes Oxidative Stress and Alters Intestinal Host-Microbe Interactions. Mol. Nutr. Food Res. 2019, 63, e1800947. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Cheng, K.; Zhou, L.; He, J.; Zheng, X.; Zhang, L.; Zhong, X.; Wang, T. Evaluation of Long-Term Toxicity of Oral Zinc Oxide Nanoparticles and Zinc Sulfate in Mice. Biol. Trace Elem. Res. 2017, 178, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Yoo, A.; Lin, M.; Mustapha, A. Zinc Oxide and Silver Nanoparticle Effects on Intestinal Bacteria. Materials 2021, 14, 2489. [Google Scholar] [CrossRef]
- Ishaq, S.L.; Page, C.M.; Yeoman, C.J.; Murphy, T.W.; Van Emon, M.L.; Stewart, W.C. Zinc AA supplementation alters yearling ram rumen bacterial communities but zinc sulfate supplementation does not. J. Anim. Sci. 2019, 97, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.-J.; Park, Y.-J.; Cho, J.H.; Song, M.-H.; Gu, B.-H.; Yun, W.; Lee, J.-H.; An, J.-S.; Kim, Y.-J.; Lee, J.-S.; et al. Changes in Diarrhea Score, Nutrient Digestibility, Zinc Utilization, Intestinal Immune Profiles, and Fecal Microbiome in Weaned Piglets by Different Forms of Zinc. Animals 2021, 11, 1356. [Google Scholar] [CrossRef]
- Wu, R.; Mei, X.; Wang, J.; Sun, W.; Xue, T.; Lin, C.; Xu, D. Zn(ii)-Curcumin supplementation alleviates gut dysbiosis and zinc dyshomeostasis during doxorubicin-induced cardiotoxicity in rats. Food Funct. 2019, 10, 5587–5604. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Mei, X.; Ye, Y.; Xue, T.; Wang, J.; Sun, W.; Lin, C.; Xue, R.; Zhang, J.; Xu, D. Zn(II)-curcumin solid dispersion impairs hepatocellular carcinoma growth and enhances chemotherapy by modulating gut microbiota-mediated zinc homeostasis. Pharmacol. Res. 2019, 150, 104454. [Google Scholar] [CrossRef] [PubMed]
- Gaulke, C.A.; Rolshoven, J.; Wong, C.P.; Hudson, L.G.; Ho, E.; Sharpton, T.J. Marginal Zinc Deficiency and Environmentally Relevant Concentrations of Arsenic Elicit Combined Effects on the Gut Microbiome. mSphere 2018, 3, e00521-18. [Google Scholar] [CrossRef] [Green Version]
- Sheng, Y.; Fan, F.; Jensen, O.; Zhong, Z.; Kan, B.; Wang, H.; Zhu, J. Dual Zinc Transporter Systems in Vibrio cholerae Promote Competitive Advantages over Gut Microbiome. Infect. Immun. 2015, 83, 3902–3908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, Y.; Lei, Z.; Yuan, J.; Yang, Y.; Guo, Y.; Zhang, B. Effect of zinc on growth performance, gut morphometry, and cecal microbial community in broilers challenged with Salmonella enterica serovar typhimurium. J. Microbiol. 2014, 52, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, P.; Saha, T.; Sheikh, I.A.; Chakraborty, S.; Aoun, J.; Chakrabarti, M.K.; Rajendran, V.M.; Ameen, N.A.; Dutta, S.; Hoque, K.M. Zinc ameliorates intestinal barrier dysfunctions in shigellosis by reinstating claudin-2 and -4 on the membranes. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G229–G246. [Google Scholar] [CrossRef]
- Zackular, J.P.; Skaar, E.P. The role of zinc and nutritional immunity in Clostridium difficile infection. Gut Microbes 2018, 9, 469–476. [Google Scholar] [CrossRef]
- Zackular, J.P.; Moore, J.L.; Jordan, A.T.; Juttukonda, L.J.; Noto, M.J.; Nicholson, M.R.; Crews, J.D.; Semler, M.W.; Zhang, Y.; Ware, L.B.; et al. Dietary zinc alters the microbiota and decreases resistance to Clostridium difficile infection. Nat. Med. 2016, 22, 1330–1334. [Google Scholar] [CrossRef]
- Parvataneni, S.; Dasari, A.R. Zinc Level and Its Role in Recurrent Clostridium difficile Infection: A Case Report and Literature Review. J. Investig. Med. High Impact Case Rep. 2020, 8, 2324709620941315. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.; Zhang, M.; Tian, S.; Wang, J.; Zhu, W. Chitosan-chelated zinc modulates cecal microbiota and attenuates inflammatory response in weaned rats challenged with Escherichia coli. J. Microbiol. 2020, 58, 780–792. [Google Scholar] [CrossRef] [PubMed]
- Pei, X.; Xiao, Z.; Liu, L.; Wang, G.; Tao, W.; Wang, M.; Zou, J.; Leng, D. Effects of dietary zinc oxide nanoparticles supplementation on growth performance, zinc status, intestinal morphology, microflora population, and immune response in weaned pigs. J. Sci. Food Agric. 2019, 99, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- Johanns, V.C.; Epping, L.; Semmler, T.; Ghazisaeedi, F.; Lübke-Becker, A.; Pfeifer, Y.; Eichhorn, I.; Merle, R.; Bethe, A.; Walther, B.; et al. High-Zinc Supplementation of Weaned Piglets Affects Frequencies of Virulence and Bacteriocin Associated Genes Among Intestinal Escherichia coli Populations. Front. Vet. Sci. 2020, 7, 614513. [Google Scholar] [CrossRef]
- Crane, J.K.; Naeher, T.M.; Shulgina, I.; Zhu, C.; Boedeker, E.C. Effect of zinc in enteropathogenic Escherichia coli infection. Infect. Immun. 2007, 75, 5974–5984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bücker, R.; Zakrzewski, S.S.; Wiegand, S.; Pieper, R.; Fromm, A.; Fromm, M.; Günzel, D.; Schulzke, J.D. Zinc prevents intestinal epithelial barrier dysfunction induced by alpha-hemolysin-producing Escherichia coli 536 infection in porcine colon. Vet. Microbiol. 2020, 243, 108632. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, Z.; Wang, P.; Yu, X.; Ding, H.; Wang, Z.; Feng, J. Effect of Long-Term and Short-Term Imbalanced Zn Manipulation on Gut Microbiota and Screening for Microbial Markers Sensitive to Zinc Status. Microbiol. Spectr. 2021, 9, e0048321. [Google Scholar] [CrossRef]
- Ciesinski, L.; Guenther, S.; Pieper, R.; Kalisch, M.; Bednorz, C.; Wieler, L.H. High dietary zinc feeding promotes persistence of multi-resistant E. coli in the swine gut. PLoS ONE 2018, 13, e0191660. [Google Scholar] [CrossRef] [PubMed]
- Gammoh, N.Z.; Rink, L. Zinc in Infection and Inflammation. Nutrients 2017, 9, 624. [Google Scholar] [CrossRef] [Green Version]
- Souffriau, J.; Timmermans, S.; Vanderhaeghen, T.; Wallaeys, C.; Van Looveren, K.; Aelbrecht, L.; Dewaele, S.; Vandewalle, J.; Goossens, E.; Verbanck, S.; et al. Zinc inhibits lethal inflammatory shock by preventing microbe-induced interferon signature in intestinal epithelium. EMBO Mol. Med. 2020, 12, e11917. [Google Scholar] [CrossRef] [PubMed]
- Straub, R.H.; Schradin, C. Chronic inflammatory systemic diseases: An evolutionary trade-off between acutely beneficial but chronically harmful programs. Evol. Med. Public Health 2016, 2016, 37–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, S.Y.; Tang, Q.Q. Altered intestinal microflora and barrier injury in severe acute pancreatitis can be changed by zinc. Int. J. Med. Sci. 2021, 18, 3050. [Google Scholar] [CrossRef]
- Lee, A.A.; Baker, J.R.; Wamsteker, E.J.; Saad, R.; DiMagno, M.J. Small Intestinal Bacterial Overgrowth Is Common in Chronic Pancreatitis and Associates With Diabetes, Chronic Pancreatitis Severity, Low Zinc Levels, and Opiate Use. Am. J. Gastroenterol. 2019, 114, 1163–1171. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.R.; Vaishnava, S. Zinc supplementation modulates T helper 17 cells via the gut microbiome. J. Immunol. 2019, 202, 191.13. [Google Scholar]
- Vela, G.; Stark, P.; Socha, M.; Sauer, A.K.; Hagmeyer, S.; Grabrucker, A.M. Zinc in gut-brain interaction in autism and neurological disorders. Neural. Plast. 2015, 972791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, G.C.; Jung, Y.; Lee, K.; Fourie, C.; Handley, K.M.; Montgomery, J.M.; Taylor, M.W. Effect of dietary zinc supplementation on the gastrointestinal microbiota and host gene expression in the Shank3B-/-mouse model of autism spectrum disorder. bioRxiv 2021. [Google Scholar] [CrossRef]
- Yu, R.; Ahmed, T.; Jiang, H.; Zhou, G.; Zhang, M.; Lv, L.; Li, B. Impact of Zinc Oxide Nanoparticles on the Composition of Gut Microbiota in Healthy and Autism Spectrum Disorder Children. Materials 2021, 14, 5488. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Yu, R.; Ahmed, T.; Jiang, H.; Zhang, M.; Lv, L.; Alhumaydhi, F.A.; Allemailem, K.S.; Li, B. Biosynthesis and Characterization of Zinc Oxide Nanoparticles and Their Impact on the Composition of Gut Microbiota in Healthy and Attention-Deficit Hyperactivity Disorder Children. Front. Microbiol. 2021, 12, 700707. [Google Scholar] [CrossRef]
- Skalny, A.V.; Skalnaya, M.G.; Grabeklis, A.R.; Skalnaya, A.A.; Tinkov, A.A. Zinc deficiency as a mediator of toxic effects of alcohol abuse. Eur. J. Nutr. 2018, 57, 2313–2322. [Google Scholar] [CrossRef]
- Zhong, W.; Wei, X.; Hao, L.; Lin, T.D.; Yue, R.; Sun, X.; Guo, W.; Dong, H.; Li, T.; Ahmadi, A.R.; et al. Paneth Cell Dysfunction Mediates Alcohol-related Steatohepatitis through Promoting Bacterial Translocation in Mice: Role of Zinc Deficiency. Hepatology 2020, 71, 1575–1591. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Cho, K.Y. Altered Gut Microbiota and Shift in Bacteroidetes between Young Obese and Normal-Weight Korean Children: A Cross-Sectional Observational Study. Biomed. Res. Int. 2020, 11, 1–9. [Google Scholar] [CrossRef]
- Crovesy, L.; Masterson, D.; Rosado, E.L. Profile of the gut microbiota of adults with obesity: A systematic review. Eur. J. Clin. Nutr. 2020, 74, 1251–1262. [Google Scholar] [CrossRef]
- Olechnowicz, J.; Tinkov, A.; Skalny, A.; Suliburska, J. Zinc status is associated with inflammation, oxidative stress, lipid, and glucose metabolism. J. Physiol. Sci. 2018, 68, 19–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Zhang, S.; Chen, C.; Jiang, X.; Qiu, J.; Qiu, Y.; Zhang, Y.; Wang, T.; Qin, X.; Zou, Z.; et al. Crosstalk of gut microbiota and serum/hippocampus metabolites in neurobehavioral impairments induced by zinc oxide nanoparticles. Nanoscale 2020, 12, 21429–21439. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Cheng, S.; Jiang, X.; Zhang, J.; Bai, L.; Qin, X.; Zou, Z.; Chen, C. Gut-brain communication in hyperfunction of 5-hydroxytryptamine induced by oral zinc oxide nanoparticles exposure in young mice. Food Chem. Toxicol. 2020, 135, 110906. [Google Scholar] [CrossRef]
- Malyar, R.M.; Li, H.; Liu, D.; Abdulrahim, Y.; Farid, R.A.; Gan, F.; Ali, W.; Enayatullah, H.; Banuree, S.; Huang, K.; et al. Selenium/Zinc-Enriched probiotics improve serum enzyme activity, antioxidant ability, inflammatory factors and related gene expression of Wistar rats inflated under heat stress. Life Sci. 2020, 248, 117464. [Google Scholar] [CrossRef]
- Kwon, J.Y.; Lee, S.H.; Jhun, J.; Choi, J.; Jung, K.; Cho, K.H.; Kim, S.J.; Yang, C.W.; Park, S.H.; Cho, M.L. The Combination of Probiotic Complex, Rosavin, and Zinc Improves Pain and Cartilage Destruction in an Osteoarthritis Rat Model. J. Med. Food 2018, 21, 364–371. [Google Scholar] [CrossRef]
- Shah, M.; Zaneb, H.; Masood, S.; Khan, R.U.; Ashraf, S.; Sikandar, A.; Rehman, H.; Rehman, H.U. Effect of Dietary Supplementation of Zinc and Multi-Microbe Probiotic on Growth Traits and Alteration of Intestinal Architecture in Broiler. Probiotics Antimicrob Proteins 2019, 11, 931–937. [Google Scholar] [CrossRef]
- Shah, M.; Zaneb, H.; Masood, S.; Khan, R.U.; Mobashar, M.; Khan, I.; Din, S.; Khan, M.S.; Rehman, H.U.; Tinelli, A. Single or Combined Applications of Zinc and Multi-strain Probiotic on Intestinal Histomorphology of Broilers Under Cyclic Heat Stress. Probiotics Antimicrob Proteins 2020, 12, 473–480. [Google Scholar] [CrossRef]
- Kelleher, S.L.; Casas, I.; Carbajal, N.; Lönnerdal, B. Supplementation of infant formula with the probiotic lactobacillus reuteri and zinc: Impact on enteric infection and nutrition in infant rhesus monkeys. J. Pediatr. Gastroenterol. Nutr. 2002, 35, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Bergillos-Meca, T.; Navarro-Alarcón, M.; Cabrera-Vique, C.; Artacho, R.; Olalla, M.; Giménez, R.; Moreno-Montoro, M.; Ruiz-Bravo, A.; Lasserrot, A.; Ruiz-López, M.D. The probiotic bacterial strain Lactobacillus fermentum D3 increases in vitro the bioavailability of Ca, P, and Zn in fermented goat milk. Biol. Trace Elem. Res. 2013, 151, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Mudroňová, D.; Gancarčíková, S.; Nemcová, R. Influence of Zinc Sulphate on the Probiotic Properties of Lactobacillus plantarum CCM 7102. Folia Vet. 2019, 63, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Šefcová, M.; Levkut, M.; Bobíková, K.; Karaffová, V.; Revajová, V.; Maruščáková, I.C.; Levkutová, M.; Ševčíková, Z.; Herich, R.; Levkut, M. Cytokine response after stimulation of culture cells by zinc and probiotic strain. Vitr. Cell. Dev. Biol. Anim. 2019, 55, 830–837. [Google Scholar] [CrossRef] [PubMed]
- Behnsen, J.; Liu, J.; Valeri, M.; Hoover, E.; Tjokrosurjo, J.; Montaldo, N.P.; Treacy-Abarca, S.; Garibay, O.; Gilston, B.A.; Edwards, R.A.; et al. Probiotic Escherichia coli Nissle 1917 Uses Zinc Transporters and the Siderophore Yersiniabactin to Acquire Zinc in the Inflamed Gut and Outcompete Salmonella Typhimurium. FASEB J. 2017, 31, 622.6. [Google Scholar]
- Salvatore, S.; Hauser, B.; Devreker, T.; Vieira, M.C.; Luini, C.; Arrigo, S.; Nespoli, L.; Vandenplas, Y. Probiotics and zinc in acute infectious gastroenteritis in children: Are they effective? Nutrition 2007, 23, 498–506. [Google Scholar] [CrossRef]
- Ahmadipour, S.; Mohsenzadeh, A.; Alimadadi, H.; Salehnia, M.; Fallahi, A. Treating Viral Diarrhea in Children by Probiotic and Zinc Supplements. Pediatr. Gastroenterol. Hepatol. Nutr. 2019, 22, 162–170. [Google Scholar] [CrossRef]
- Surono, I.S.; Martono, P.D.; Kameo, S.; Suradji, E.W.; Koyama, H. Effect of probiotic L. plantarum IS-10506 and zinc supplementation on humoral immune response and zinc status of Indonesian pre-school children. J. Trace Elem. Med. Biol. 2014, 28, 465–469. [Google Scholar] [CrossRef]
- Gonzales, A.D.; Reinert, J.P. Zinc and Probiotic Therapy for Management of Hepatic Encephalopathy. Sr. Care Pharm. 2020, 35, 171–175. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skalny, A.V.; Aschner, M.; Lei, X.G.; Gritsenko, V.A.; Santamaria, A.; Alekseenko, S.I.; Prakash, N.T.; Chang, J.-S.; Sizova, E.A.; Chao, J.C.J.; et al. Gut Microbiota as a Mediator of Essential and Toxic Effects of Zinc in the Intestines and Other Tissues. Int. J. Mol. Sci. 2021, 22, 13074. https://doi.org/10.3390/ijms222313074
Skalny AV, Aschner M, Lei XG, Gritsenko VA, Santamaria A, Alekseenko SI, Prakash NT, Chang J-S, Sizova EA, Chao JCJ, et al. Gut Microbiota as a Mediator of Essential and Toxic Effects of Zinc in the Intestines and Other Tissues. International Journal of Molecular Sciences. 2021; 22(23):13074. https://doi.org/10.3390/ijms222313074
Chicago/Turabian StyleSkalny, Anatoly V., Michael Aschner, Xin Gen Lei, Viktor A. Gritsenko, Abel Santamaria, Svetlana I. Alekseenko, Nagaraja Tejo Prakash, Jung-Su Chang, Elena A. Sizova, Jane C. J. Chao, and et al. 2021. "Gut Microbiota as a Mediator of Essential and Toxic Effects of Zinc in the Intestines and Other Tissues" International Journal of Molecular Sciences 22, no. 23: 13074. https://doi.org/10.3390/ijms222313074
APA StyleSkalny, A. V., Aschner, M., Lei, X. G., Gritsenko, V. A., Santamaria, A., Alekseenko, S. I., Prakash, N. T., Chang, J. -S., Sizova, E. A., Chao, J. C. J., Aaseth, J., & Tinkov, A. A. (2021). Gut Microbiota as a Mediator of Essential and Toxic Effects of Zinc in the Intestines and Other Tissues. International Journal of Molecular Sciences, 22(23), 13074. https://doi.org/10.3390/ijms222313074