
Poly(C)-binding Protein 2 Regulates the p53 Expression via Interactions with the 5′-Terminal Region of p53 mRNA

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
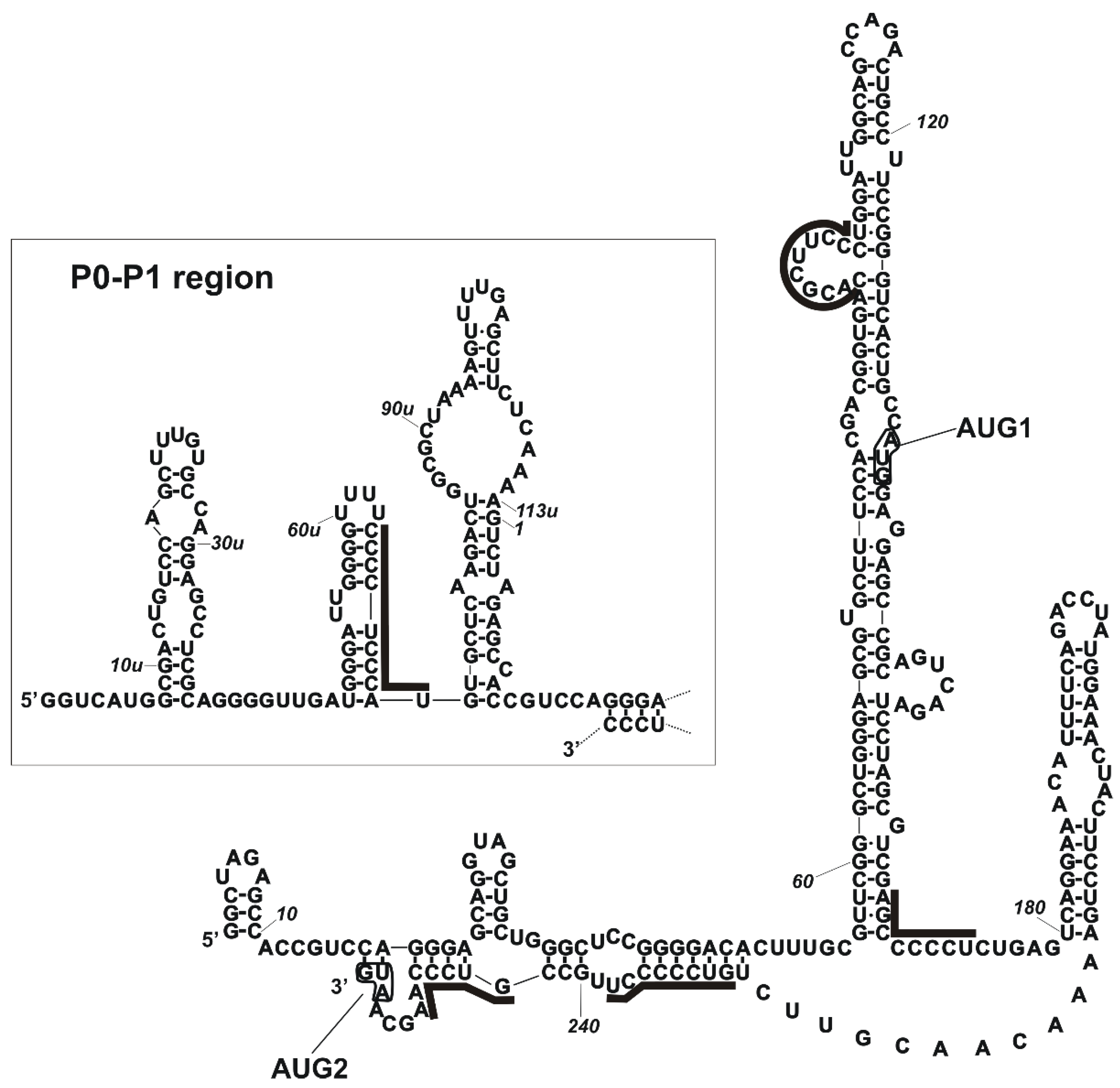
2.1. PCBP2 Binds Directly to the 5′-Terminal Region of p53 mRNA In Vitro
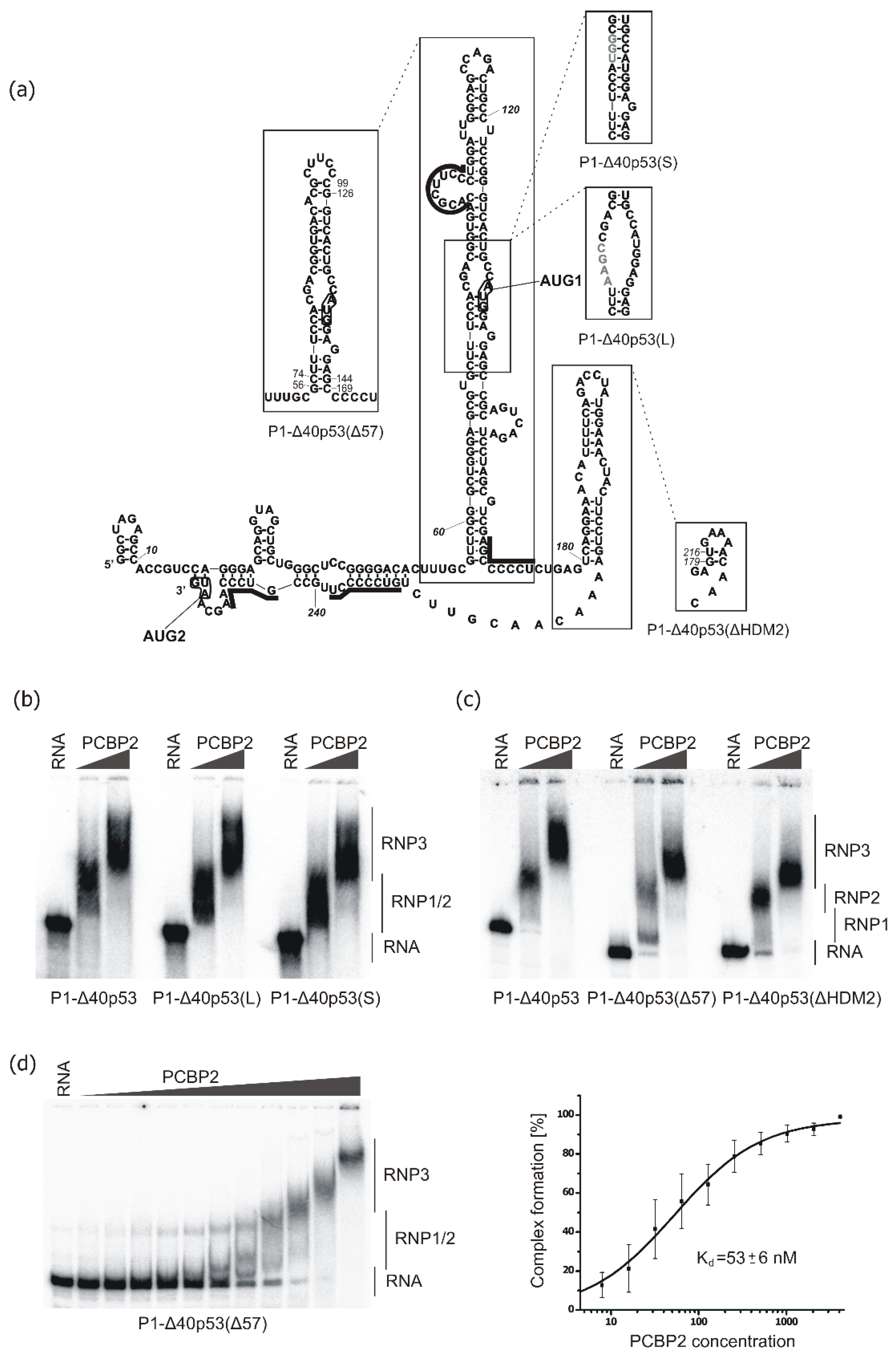
2.2. Binding of PCBP2 to Selected Structural Variants of the 5′-Terminal Region of p53 mRNA
2.3. Revealing PCBP2 Binding Sites within the 5′-Terminal Region of p53 mRNA by Structural Probing
2.4. Major PCBP2 Binding Site Revealed by Small Angle X-ray Scattering (SAXS)
2.5. Biological Meaning of PCBP2 Interactions with the 5′-Terminal Region of p53 mRNA
2.6. Is PCBP2 a Potential New ITAF?
3. Materials and Methods
3.1. Cell Culture and Stress Induction
3.2. siRNA and Plasmid Transfection
3.3. RNA Isolation, RT-PCR and Quantitative PCR (qPCR)
3.4. Western Blot
3.5. Protein Overexpression and Purification
3.6. RNA Transcription In Vitro and RNA Purification
3.7. Electrophoretic Mobility Shift Assay (EMSA)
3.8. Pb2+-Induced Cleavage and Primer Extension
3.9. Small Angle X-ray Scattering (SAXS)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aylon, Y.; Oren, M. The Paradox of p53: What, How, and Why? Cold Spring Harb. Perspect. Med. 2016, 6, a026328. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhang, C.; Hu, W.; Feng, Z. Tumor suppressor p53 and metabolism. J. Mol. Cell Biol. 2019, 11, 284–292. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, T.; Nakagawara, A. Role of p53 in Cell Death and Human Cancers. Cancers 2011, 3, 994–1013. [Google Scholar] [CrossRef]
- Kastenhuber, E.R.; Lowe, S.W. Putting p53 in Context. Cell 2017, 170, 1062–1078. [Google Scholar] [CrossRef] [Green Version]
- Haronikova, L.; Olivares-Illana, V.; Wang, L.; Karakostis, K.; Chen, S.; Fahraeus, R. The p53 mRNA: An integral part of the cellular stress response. Nucleic Acids Res. 2019, 47, 3257–3271. [Google Scholar] [CrossRef] [Green Version]
- Swiatkowska, A.; Dutkiewicz, M.; Machtel, P.; Janecki, D.M.; Kabacinska, M.; Zydowicz-Machtel, P.; Ciesiolka, J. Regulation of the p53 expression profile by hnRNP K under stress conditions. RNA Biol. 2020, 17, 1402–1415. [Google Scholar] [CrossRef]
- Galban, S.; Martindale, J.L.; Mazan-Mamczarz, K.; Lopez de Silanes, I.; Fan, J.; Wang, W.; Decker, J.; Gorospe, M. Influence of the RNA-binding protein HuR in pVHL-regulated p53 expression in renal carcinoma cells. Mol. Cell. Biol. 2003, 23, 7083–7095. [Google Scholar] [CrossRef] [Green Version]
- Burns, D.M.; Richter, J.D. CPEB regulation of human cellular senescence, energy metabolism, and p53 mRNA translation. Genes Dev. 2008, 22, 3449–3460. [Google Scholar] [CrossRef] [Green Version]
- Swiatkowska, A.; Dutkiewicz, M.; Zydowicz-Machtel, P.; Szpotkowska, J.; Janecki, D.M.; Ciesiolka, J. Translational Control in p53 Expression: The Role of 5′-Terminal Region of p53 mRNA. Int. J. Mol. Sci. 2019, 20, 5382. [Google Scholar] [CrossRef] [Green Version]
- Rahman-Roblick, R.; Roblick, U.J.; Hellman, U.; Conrotto, P.; Liu, T.; Becker, S.; Hirschberg, D.; Jornvall, H.; Auer, G.; Wiman, K.G. p53 targets identified by protein expression profiling. Proc. Natl. Acad. Sci. USA 2007, 104, 5401–5406. [Google Scholar] [CrossRef] [Green Version]
- Moumen, A.; Masterson, P.; O’Connor, M.J.; Jackson, S.P. hnRNP K: An HDM2 target and transcriptional coactivator of p53 in response to DNA damage. Cell 2005, 123, 1065–1078. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.; Kim, J.H.; Li, X.; Paek, K.Y.; Ha, S.H.; Ryu, S.H.; Wimmer, E.; Jang, S.K. Identification of cellular proteins enhancing activities of internal ribosomal entry sites by competition with oligodeoxynucleotides. Nucleic Acids Res. 2004, 32, 1308–1317. [Google Scholar] [CrossRef] [Green Version]
- Makeyev, A.V.; Liebhaber, S.A. The poly(C)-binding proteins: A multiplicity of functions and a search for mechanisms. RNA 2002, 8, 265–278. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.; Fenn, S.; Tjhen, R.; James, T.L. Structure of a construct of a human poly(C)-binding protein containing the first and second KH domains reveals insights into its regulatory mechanisms. J. Biol. Chem. 2008, 283, 28757–28766. [Google Scholar] [CrossRef] [Green Version]
- Yoga, Y.M.; Traore, D.A.; Sidiqi, M.; Szeto, C.; Pendini, N.R.; Barker, A.; Leedman, P.J.; Wilce, J.A.; Wilce, M.C. Contribution of the first K-homology domain of poly(C)-binding protein 1 to its affinity and specificity for C-rich oligonucleotides. Nucleic Acids Res. 2012, 40, 5101–5114. [Google Scholar] [CrossRef] [Green Version]
- Paziewska, A.; Wyrwicz, L.S.; Bujnicki, J.M.; Bomsztyk, K.; Ostrowski, J. Cooperative binding of the hnRNP K three KH domains to mRNA targets. FEBS Lett. 2004, 577, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Holcik, M.; Liebhaber, S.A. Four highly stable eukaryotic mRNAs assemble 3′ untranslated region RNA-protein complexes sharing cis and trans components. Proc. Natl. Acad. Sci. USA 1997, 94, 2410–2414. [Google Scholar] [CrossRef] [Green Version]
- Rizzotto, D.; Zaccara, S.; Rossi, A.; Dassi, E.; Inga, A. Translation control can shape TP53-dependent cell fate. Mol. Cell. Oncol. 2020, 7, 1767483. [Google Scholar] [CrossRef]
- Rizzotto, D.; Zaccara, S.; Rossi, A.; Galbraith, M.D.; Andrysik, Z.; Pandey, A.; Sullivan, K.D.; Quattrone, A.; Espinosa, J.M.; Dassi, E.; et al. Nutlin-Induced Apoptosis Is Specified by a Translation Program Regulated by PCBP2 and DHX30. Cell Rep. 2020, 30, 4355–4369.e6. [Google Scholar] [CrossRef]
- Blyn, L.B.; Swiderek, K.M.; Richards, O.; Stahl, D.C.; Semler, B.L.; Ehrenfeld, E. Poly(rC) binding protein 2 binds to stem-loop IV of the poliovirus RNA 5′ noncoding region: Identification by automated liquid chromatography-tandem mass spectrometry. Proc. Natl. Acad. Sci. USA 1996, 93, 11115–11120. [Google Scholar] [CrossRef] [Green Version]
- Blyn, L.B.; Towner, J.S.; Semler, B.L.; Ehrenfeld, E. Requirement of poly(rC) binding protein 2 for translation of poliovirus RNA. J. Virol. 1997, 71, 6243–6246. [Google Scholar] [CrossRef] [Green Version]
- Parsley, T.B.; Towner, J.S.; Blyn, L.B.; Ehrenfeld, E.; Semler, B.L. Poly (rC) binding protein 2 forms a ternary complex with the 5′-terminal sequences of poliovirus RNA and the viral 3CD proteinase. RNA 1997, 3, 1124–1134. [Google Scholar]
- Lin, X.; Yang, B.; Liu, W.; Tan, X.; Wu, F.; Hu, P.; Jiang, T.; Bao, Z.; Yuan, J.; Qiang, B.; et al. Interplay between PCBP2 and miRNA modulates ARHGDIA expression and function in glioma migration and invasion. Oncotarget 2016, 7, 19483–19498. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Liu, J.; Chen, C.; Zhang, W.; Qian, R.; Chen, X.; Lu, H.; Ge, J.; Zhao, C.; Zhang, D.; et al. PCBP2 Modulates Neural Apoptosis and Astrocyte Proliferation After Spinal Cord Injury. Neurochem. Res. 2016, 41, 2401–2414. [Google Scholar] [CrossRef]
- Ren, C.; Zhang, J.; Yan, W.; Zhang, Y.; Chen, X. RNA-binding Protein PCBP2 Regulates p73 Expression and p73-dependent Antioxidant Defense. J. Biol. Chem. 2016, 291, 9629–9637. [Google Scholar] [CrossRef] [Green Version]
- Thisted, T.; Lyakhov, D.L.; Liebhaber, S.A. Optimized RNA targets of two closely related triple KH domain proteins, heterogeneous nuclear ribonucleoprotein K and alphaCP-2KL, suggest Distinct modes of RNA recognition. J. Biol. Chem. 2001, 276, 17484–17496. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, D.; Freese, P.; Alexis, M.S.; Su, A.; Hochman, M.; Palden, T.; Bazile, C.; Lambert, N.J.; Van Nostrand, E.L.; Pratt, G.A.; et al. Sequence, Structure, and Context Preferences of Human RNA Binding Proteins. Mol. Cell 2018, 70, 854–867.e9. [Google Scholar] [CrossRef] [Green Version]
- Strudwick, S.; Carastro, L.M.; Stagg, T.; Lazarus, P. Differential transcription-coupled translational inhibition of human p53 expression: A potentially important mechanism of regulating p53 expression in normal versus tumor tissue. Mol. Cancer Res. 2003, 1, 463–474. [Google Scholar]
- Gorska, A.; Blaszczyk, L.; Dutkiewicz, M.; Ciesiolka, J. Length variants of the 5′ untranslated region of p53 mRNA and their impact on the efficiency of translation initiation of p53 and its N-truncated isoform DeltaNp53. RNA Biol. 2013, 10, 1726–1740. [Google Scholar] [CrossRef] [Green Version]
- Zydowicz-Machtel, P.; Swiatkowska, A.; Popenda, L.; Gorska, A.; Ciesiolka, J. Variants of the 5′-terminal region of p53 mRNA influence the ribosomal scanning and translation efficiency. Sci. Rep. 2018, 8, 1533. [Google Scholar] [CrossRef] [Green Version]
- Swiatkowska, A.; Zydowicz, P.; Sroka, J.; Ciesiolka, J. The role of the 5′ terminal region of p53 mRNA in the p53 gene expression. Acta Biochim. Pol. 2016, 63, 645–651. [Google Scholar] [CrossRef]
- Gamarnik, A.V.; Andino, R. Interactions of viral protein 3CD and poly(rC) binding protein with the 5′ untranslated region of the poliovirus genome. J. Virol. 2000, 74, 2219–2226. [Google Scholar] [CrossRef] [Green Version]
- Chkheidze, A.N.; Lyakhov, D.L.; Makeyev, A.V.; Morales, J.; Kong, J.; Liebhaber, S.A. Assembly of the alpha-globin mRNA stability complex reflects binary interaction between the pyrimidine-rich 3′ untranslated region determinant and poly(C) binding protein alphaCP. Mol. Cell. Biol. 1999, 19, 4572–4581. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.R.; Mitchell, S.A.; Spriggs, K.A.; Ostrowski, J.; Bomsztyk, K.; Ostarek, D.; Willis, A.E. Members of the poly (rC) binding protein family stimulate the activity of the c-myc internal ribosome entry segment in vitro and in vivo. Oncogene 2003, 22, 8012–8020. [Google Scholar] [CrossRef] [Green Version]
- Beckham, S.A.; Matak, M.Y.; Belousoff, M.J.; Venugopal, H.; Shah, N.; Vankadari, N.; Elmlund, H.; Nguyen, J.H.C.; Semler, B.L.; Wilce, M.C.J.; et al. Structure of the PCBP2/stem-loop IV complex underlying translation initiation mediated by the poliovirus type I IRES. Nucleic Acids Res. 2020, 48, 8006–8021. [Google Scholar] [CrossRef]
- Ciesiolka, J.; Lorenz, S.; Erdmann, V.A. Structural-Analysis of 3 Prokaryotic 5s Ribosomal-Rna Species and Selected 5s Ribosomal-Rna Ribosomal-Protein Complexes by Means of Pb(Ii)-Induced Hydrolysis. Eur. J. Biochem. 1992, 204, 575–581. [Google Scholar] [CrossRef]
- Kirsebom, L.A.; Ciesiolka, J. Pb(II)-induced cleavage of RNA. In Handbook of RNA Biochemistry; Hartmann, R.K., Schon, A., Westhof, E., Eds.; Wiley VCH GmbH& Co.: Wenheim, Germany, 2005; pp. 214–228. [Google Scholar]
- Blaszczyk, L.; Ciesiolka, J. Secondary structure and the role in translation initiation of the 5′-terminal region of p53 mRNA. Biochemistry 2011, 50, 7080–7092. [Google Scholar] [CrossRef]
- Gorska, A.; Swiatkowska, A.; Dutkiewicz, M.; Ciesiolka, J. Modulation of p53 expression using antisense oligonucleotides complementary to the 5′-terminal region of p53 mRNA in vitro and in the living cells. PLoS ONE 2013, 8, e78863. [Google Scholar] [CrossRef] [Green Version]
- Szpotkowska, J.; Swiatkowska, A.; Ciesiolka, J. Length and secondary structure of the 5′ non-coding regions of mouse p53 mRNA transcripts—mouse as a model organism for p53 gene expression studies. RNA Biol. 2019, 16, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Manszewski, T.; Szpotkowski, K.; Jaskolski, M. Crystallographic and SAXS studies of S-adenosyl-l-homocysteine hydrolase from Bradyrhizobium elkanii. IUCrJ 2017, 4, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Bierwagen, P.; Szpotkowski, K.; Jaskolski, M.; Urbanowicz, A. Borrelia outer surface protein C is capable of human fibrinogen binding. FEBS J. 2019, 286, 2415–2428. [Google Scholar] [CrossRef] [PubMed]
- Gudanis, D.; Popenda, L.; Szpotkowski, K.; Kierzek, R.; Gdaniec, Z. Structural characterization of a dimer of RNA duplexes composed of 8-bromoguanosine modified CGG trinucleotide repeats: A novel architecture of RNA quadruplexes. Nucleic Acids Res. 2016, 44, 2409–2416. [Google Scholar] [CrossRef] [Green Version]
- Urbanowicz, A.; Lewandowski, D.; Szpotkowski, K.; Figlerowicz, M. Tick receptor for outer surface protein A from Ixodes ricinus—the first intrinsically disordered protein involved in vector-microbe recognition. Sci. Rep. 2016, 6, 25205. [Google Scholar] [CrossRef] [Green Version]
- Dejgaard, K.; Leffers, H. Characterisation of the nucleic-acid-binding activity of KH domains. Different properties of different domains. Eur. J. Biochem. 1996, 241, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Leffers, H.; Dejgaard, K.; Celis, J.E. Characterisation of two major cellular poly(rC)-binding human proteins, each containing three K-homologous (KH) domains. Eur. J. Biochem. 1995, 230, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Perera, R.; Daijogo, S.; Walter, B.L.; Nguyen, J.H.; Semler, B.L. Cellular protein modification by poliovirus: The two faces of poly(rC)-binding protein. J. Virol. 2007, 81, 8919–8932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnova, V.V.; Shestakova, E.D.; Bikmetov, D.V.; Chugunova, A.A.; Osterman, I.A.; Serebryakova, M.V.; Sergeeva, O.V.; Zatsepin, T.S.; Shatsky, I.N.; Terenin, I.M. eIF4G2 balances its own mRNA translation via a PCBP2-based feedback loop. RNA 2019, 25, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Saldana-Meyer, R.; Recillas-Targa, F. Transcriptional and epigenetic regulation of the p53 tumor suppressor gene. Epigenetics 2011, 6, 1068–1077. [Google Scholar] [CrossRef] [Green Version]
- Marques-Ramos, A.; Candeias, M.M.; Menezes, J.; Lacerda, R.; Willcocks, M.; Teixeira, A.; Locker, N.; Romao, L. Cap-independent translation ensures mTOR expression and function upon protein synthesis inhibition. RNA 2017, 23, 1712–1728. [Google Scholar] [CrossRef] [Green Version]
- Krzesniak, M.; Zajkowicz, A.; Matuszczyk, I.; Rusin, M. Rapamycin prevents strong phosphorylation of p53 on serine 46 and attenuates activation of the p53 pathway in A549 lung cancer cells exposed to actinomycin D. Mech. Ageing Dev. 2014, 139, 11–21. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Z. IRES-mediated cap-independent translation, a path leading to hidden proteome. J. Mol. Cell Biol. 2019, 11, 911–919. [Google Scholar] [CrossRef] [Green Version]
- Ji, B.; Harris, B.R.; Liu, Y.; Deng, Y.; Gradilone, S.A.; Cleary, M.P.; Liu, J.; Yang, D.Q. Targeting IRES-Mediated p53 Synthesis for Cancer Diagnosis and Therapeutics. Int. J. Mol. Sci. 2017, 18, 93. [Google Scholar] [CrossRef] [Green Version]
- Stoneley, M.; Willis, A.E. Cellular internal ribosome entry segments: Structures, trans-acting factors and regulation of gene expression. Oncogene 2004, 23, 3200–3207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grover, R.; Ray, P.S.; Das, S. Polypyrimidine tract binding protein regulates IRES-mediated translation of p53 isoforms. Cell Cycle 2008, 7, 2189–2198. [Google Scholar] [CrossRef] [Green Version]
- Sharathchandra, A.; Lal, R.; Khan, D.; Das, S. Annexin A2 and PSF proteins interact with p53 IRES and regulate translation of p53 mRNA. RNA Biol. 2012, 9, 1429–1439. [Google Scholar] [CrossRef] [Green Version]
- Weingarten-Gabbay, S.; Khan, D.; Liberman, N.; Yoffe, Y.; Bialik, S.; Das, S.; Oren, M.; Kimchi, A. The translation initiation factor DAP5 promotes IRES-driven translation of p53 mRNA. Oncogene 2014, 33, 611–618. [Google Scholar] [CrossRef]
- Halaby, M.J.; Li, Y.; Harris, B.R.; Jiang, S.; Miskimins, W.K.; Cleary, M.P.; Yang, D.Q. Translational Control Protein 80 Stimulates IRES-Mediated Translation of p53 mRNA in Response to DNA Damage. BioMed Res. Int. 2015, 2015, 708158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Guo, K.; Kastan, M.B. Interactions of nucleolin and ribosomal protein L26 (RPL26) in translational control of human p53 mRNA. J. Biol. Chem. 2012, 287, 16467–16476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Kastan, M.B. 5′-3′-UTR interactions regulate p53 mRNA translation and provide a target for modulating p53 induction after DNA damage. Genes Dev. 2010, 24, 2146–2156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takagi, M.; Absalon, M.J.; McLure, K.G.; Kastan, M.B. Regulation of p53 translation and induction after DNA damage by ribosomal protein L26 and nucleolin. Cell 2005, 123, 49–63. [Google Scholar] [CrossRef] [Green Version]
- Swiatkowska, A.; Zydowicz, P.; Gorska, A.; Suchacka, J.; Dutkiewicz, M.; Ciesiolka, J. The Role of Structural Elements of the 5′-Terminal Region of p53 mRNA in Translation under Stress Conditions Assayed by the Antisense Oligonucleotide Approach. PLoS ONE 2015, 10, e0141676. [Google Scholar] [CrossRef]
- Zydowicz-Machtel, P.; Dutkiewicz, M.; Swiatkowska, A.; Gurda-Wozna, D.; Ciesiolka, J. Translation of human Delta133p53 mRNA and its targeting by antisense oligonucleotides complementary to the 5′-terminal region of this mRNA. PLoS ONE 2021, 16, e0256938. [Google Scholar] [CrossRef] [PubMed]
- Konarev, P.V.; Volkov, V.V.; Sokolova, A.V.; Koch, M.H.J.; Svergun, D.I. PRIMUS: A Windows PC-based system for small-angle scattering data analysis. J. Appl. Crystallogr. 2003, 36, 1277–1282. [Google Scholar] [CrossRef]
- Putnam, C.D.; Hammel, M.; Hura, G.L.; Tainer, J.A. X-ray solution scattering (SAXS) combined with crystallography and computation: Defining accurate macromolecular structures, conformations and assemblies in solution. Q. Rev. Biophys. 2007, 40, 191–285. [Google Scholar] [CrossRef] [PubMed]
- Antczak, M.; Popenda, M.; Zok, T.; Sarzynska, J.; Ratajczak, T.; Tomczyk, K.; Adamiak, R.W.; Szachniuk, M. New functionality of RNAComposer: An application to shape the axis of miR160 precursor structure. Acta Biochim. Pol. 2016, 63, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Popenda, M.; Szachniuk, M.; Antczak, M.; Purzycka, K.J.; Lukasiak, P.; Bartol, N.; Blazewicz, J.; Adamiak, R.W. Automated 3D structure composition for large RNAs. Nucleic Acids Res. 2012, 40, e112. [Google Scholar] [CrossRef] [PubMed]
- Schwede, T.; Kopp, J.; Guex, N.; Peitsch, M.C. SWISS-MODEL: An automated protein homology-modeling server. Nucleic Acids Res. 2003, 31, 3381–3385. [Google Scholar] [CrossRef] [Green Version]
- Svergun, D.; Barberato, C.; Koch, M.H.J. CRYSOL—A program to evaluate x-ray solution scattering of biological macromolecules from atomic coordinates. J. Appl. Crystallogr. 1995, 28, 768–773. [Google Scholar] [CrossRef]
- Bedard, K.M.; Walter, B.L.; Semler, B.L. Multimerization of poly(rC) binding protein 2 is required for translation initiation mediated by a viral IRES. RNA 2004, 10, 1266–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Paek, K.Y.; Choi, K.; Kim, T.D.; Hahm, B.; Kim, K.T.; Jang, S.K. Heterogeneous nuclear ribonucleoprotein C modulates translation of c-myc mRNA in a cell cycle phase-dependent manner. Mol. Cell. Biol. 2003, 23, 708–720. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; Xin, Z.; Zhao, Z.; Bao, W.; Lin, X.; Yin, B.; Zhao, J.; Yuan, J.; Qiang, B.; Peng, X. RNA-binding protein PCBP2 modulates glioma growth by regulating FHL3. J. Clin. Investig. 2013, 123, 2103–2118. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Kim, I.; Choi, K.; Choi, J.H.; Kim, E.; Lee, H.Y.; Park, J.; Kim Yoon, S. Poly(rC) binding protein 2 acts as a negative regulator of IRES-mediated translation of Hr mRNA. Exp. Mol. Med. 2018, 50, e441. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line Type | MCF-7 | HepG2 | HT-29 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RNA (construct) | P1-Δ40p53 | P0-Δ40p53 | P1-Δ40p53 | P0-Δ40p53 | P1-Δ40p53 | P0-Δ40p53 | ||||||
| Stress factor | - | Dox | - | Dox | - | Dox | - | Dox | - | Dox | - | Dox |
| Poly(C) Binding protein 2 (PCBP2) | ||||||||||||
| Peptide numbers (sequence matches) | 10 | 11 | 11 | 11 | 5 | 4 | 7 | 3 | 8 | 8 | 8 | 6 |
| Sequence coverage | 34.0 | 46.6 | 31.8 | 47.1 | 31.8 | 14.3 | 34.8 | 16.2 | 37.5 | 33.4 | 39.7 | 24.9 |
| Sample | Complex mRNA p53-PCBP2 | mRNA p53 | PCBP2 |
|---|---|---|---|
| Data collection | |||
| Instrument | P12, PETRA III | ||
| s range (nm−1) | 0.0088-5.0 | ||
| Wavelength (Å) | 1.24 | ||
| Temperature | 15 °C | ||
| Structural parameters | |||
| I(0) (from p(r)) (arbitrary units) | 0.4970 | 0.1472 | 0.4071 |
| Rg (from p(r)) (nm) | 6.77 | 6.37 | 4.01 |
| I(0) (from Guinier) (arbitrary units) | 0.52057 | 0.15571 | 0.3902 |
| Rg (from Gunier) (nm) | 6.79 | 6.53 | 3.98 |
| Rg (theoretical) (nm) | 6.013 | 6.47 | 3.3 |
| Dmax (nm) | 20 | 19.00 | 12.5 |
| Porod volume estimate (nm3) | 9125 | 7352 | 3002 |
| Dry volume calculated from the model (nm3) | 8893 | 7112 | 2962 |
| Molecular mass determination | |||
| Contrast (Δρ × 1010 cm−2) | 3.047 | ||
| Experimental molecular weight (Da) | 120,890 | 81,870 | 38,901 |
| Theoretical molecular weight (Da) | 120,260 | 81,680 | 38,580 |
| Software used | |||
| Primary data reduction | PRIMUS | ||
| Data processing | PRIMUS | ||
| Quaternary structure modelling | SASREF | ||
| Computation of model intensities | CRYSOL | ||
| 3D graphics representation | PyMOL | ||
| Abbreviations: Rg—Radius of gyration, Dmax—maximum dimension | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janecki, D.M.; Swiatkowska, A.; Szpotkowska, J.; Urbanowicz, A.; Kabacińska, M.; Szpotkowski, K.; Ciesiołka, J. Poly(C)-binding Protein 2 Regulates the p53 Expression via Interactions with the 5′-Terminal Region of p53 mRNA. Int. J. Mol. Sci. 2021, 22, 13306. https://doi.org/10.3390/ijms222413306
Janecki DM, Swiatkowska A, Szpotkowska J, Urbanowicz A, Kabacińska M, Szpotkowski K, Ciesiołka J. Poly(C)-binding Protein 2 Regulates the p53 Expression via Interactions with the 5′-Terminal Region of p53 mRNA. International Journal of Molecular Sciences. 2021; 22(24):13306. https://doi.org/10.3390/ijms222413306
Chicago/Turabian StyleJanecki, Damian M., Agata Swiatkowska, Joanna Szpotkowska, Anna Urbanowicz, Martyna Kabacińska, Kamil Szpotkowski, and Jerzy Ciesiołka. 2021. "Poly(C)-binding Protein 2 Regulates the p53 Expression via Interactions with the 5′-Terminal Region of p53 mRNA" International Journal of Molecular Sciences 22, no. 24: 13306. https://doi.org/10.3390/ijms222413306
APA StyleJanecki, D. M., Swiatkowska, A., Szpotkowska, J., Urbanowicz, A., Kabacińska, M., Szpotkowski, K., & Ciesiołka, J. (2021). Poly(C)-binding Protein 2 Regulates the p53 Expression via Interactions with the 5′-Terminal Region of p53 mRNA. International Journal of Molecular Sciences, 22(24), 13306. https://doi.org/10.3390/ijms222413306