
The Lrp/AsnC-Type Regulator PA2577 Controls the EamA-like Transporter Gene PA2576 in Pseudomonas aeruginosa


Abstract
:1. Introduction
2. Results
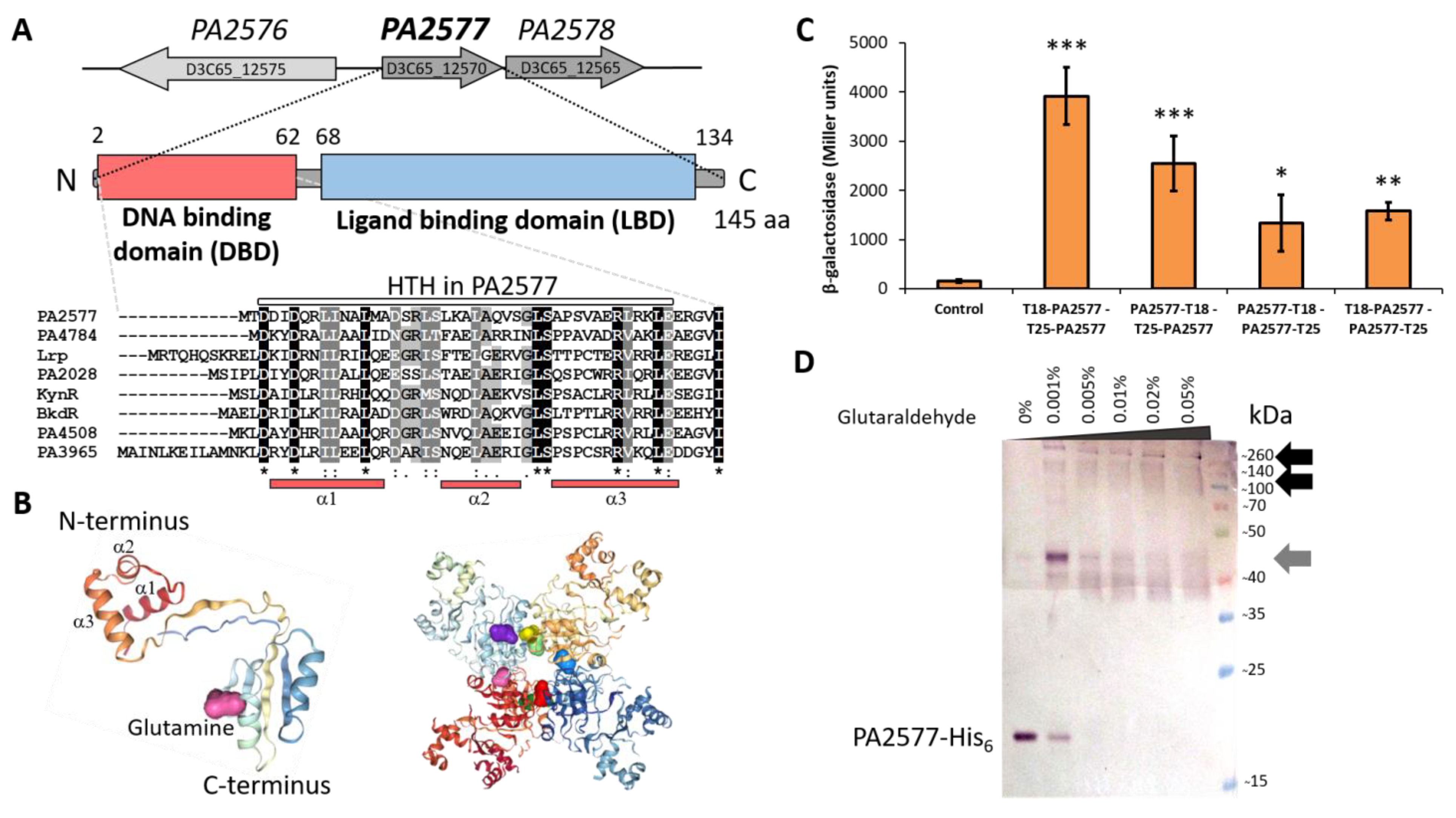
2.1. Genetic Organization of the P. aeruginosa PA2577 Region and Characteristics of PA2577
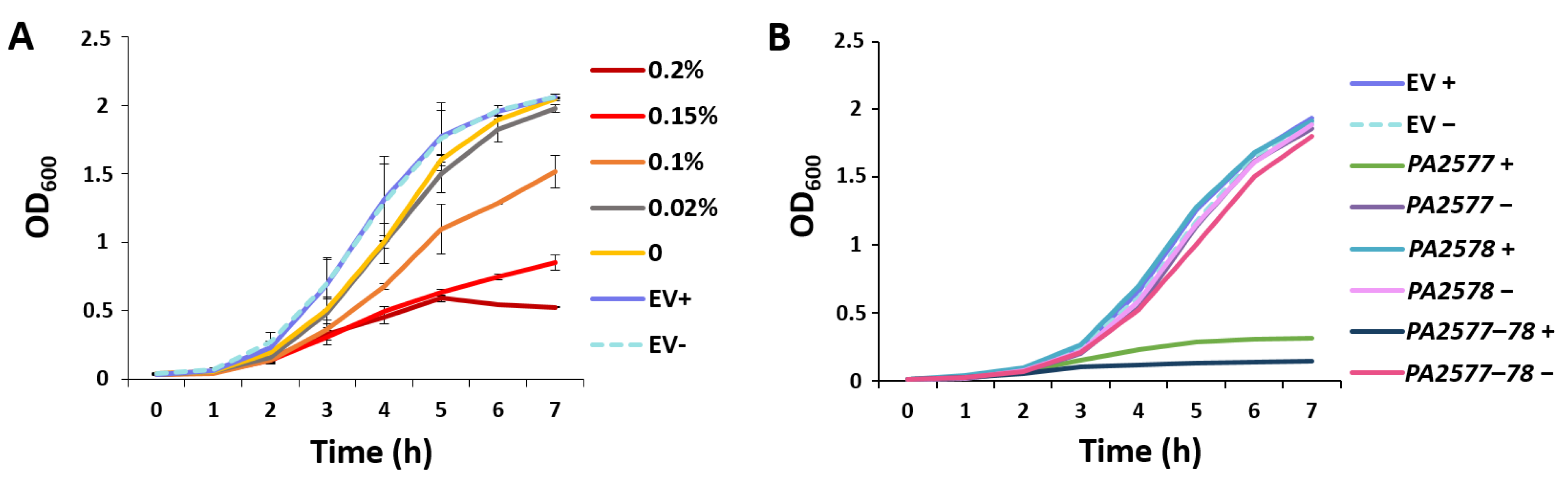
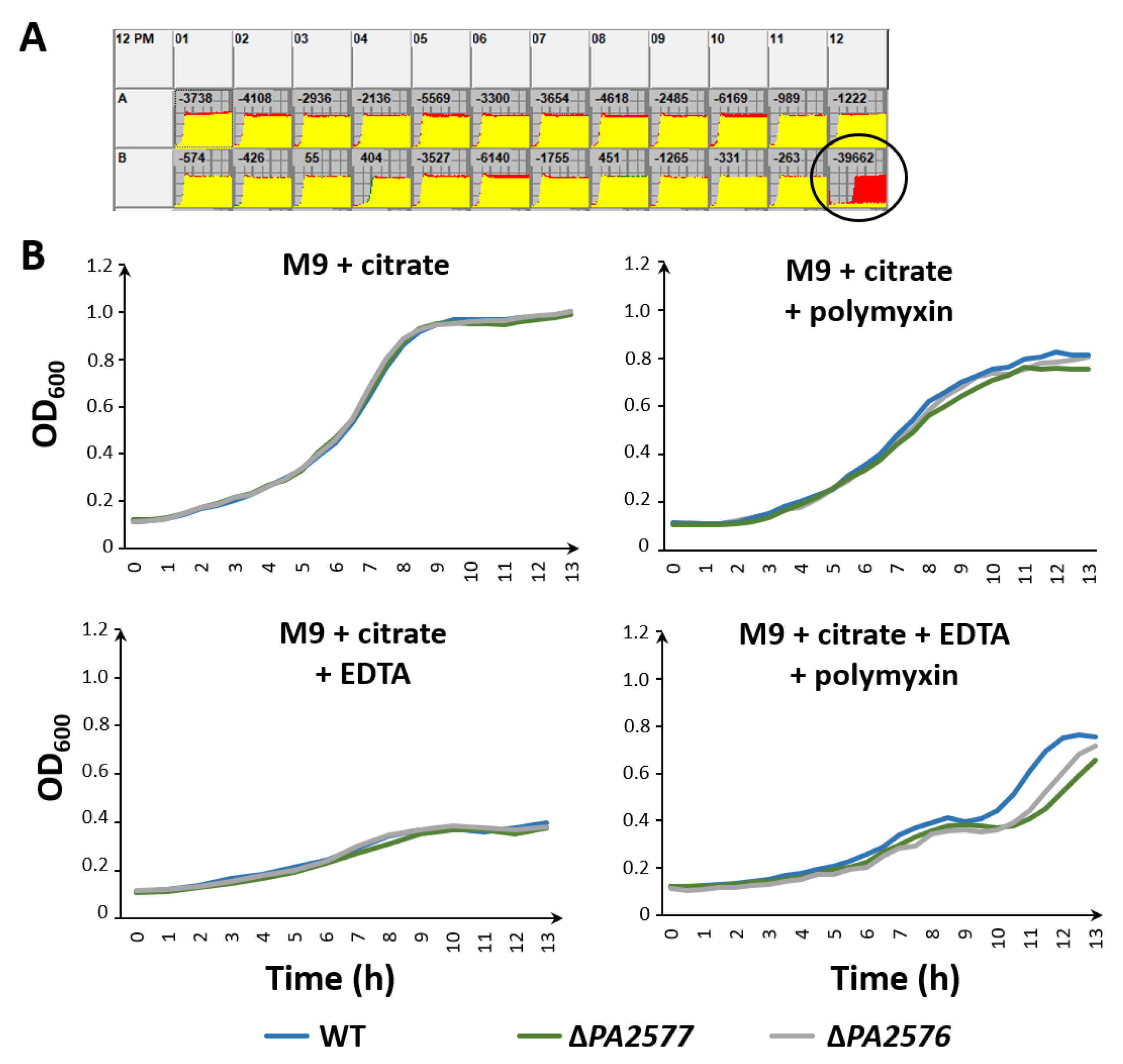
2.2. Effect of PA2577 Lack or Excess on Bacterial Growth
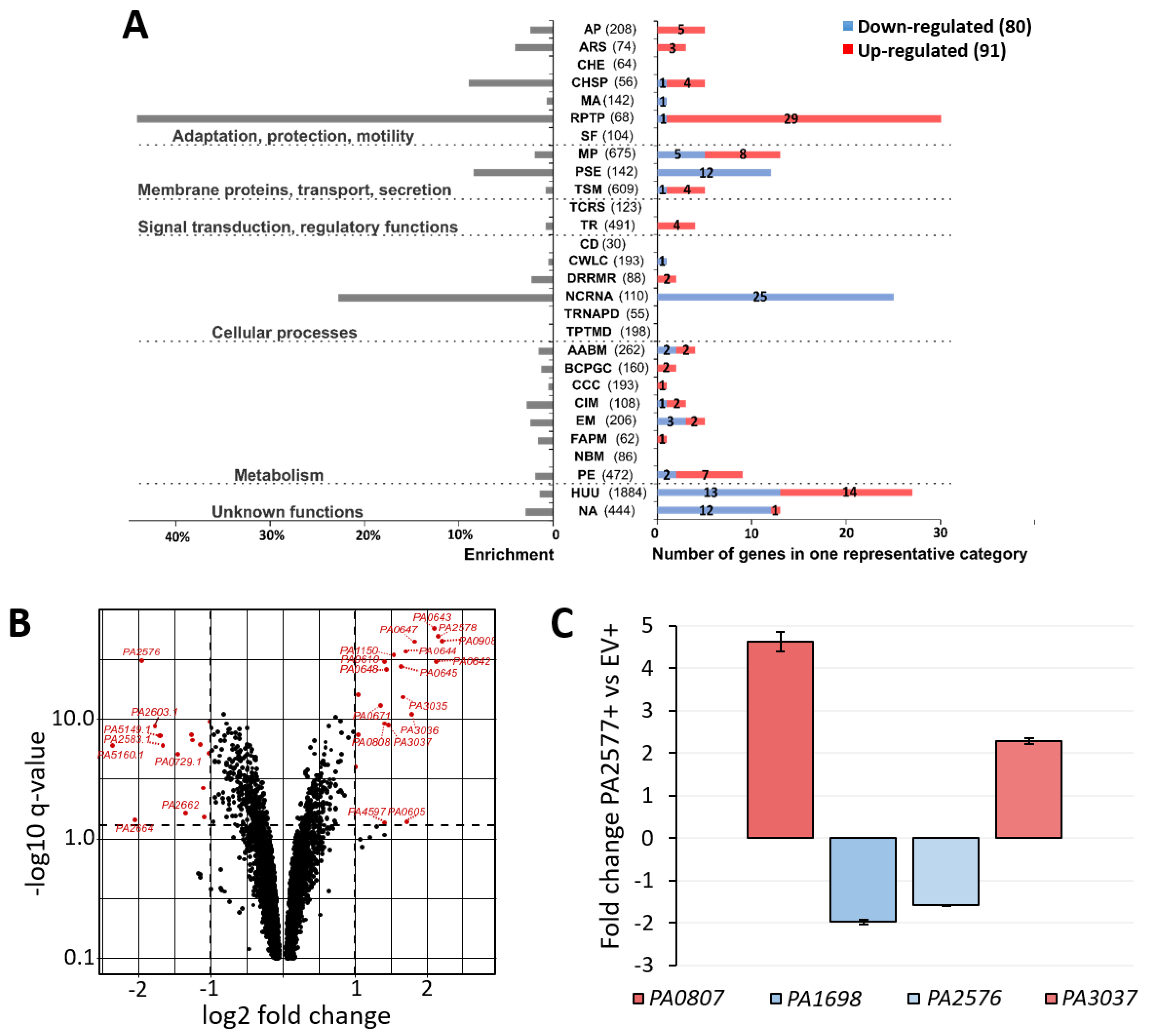
2.3. Transcriptomic Analysis of P. aeruginosa Cells Overexpressing PA2577
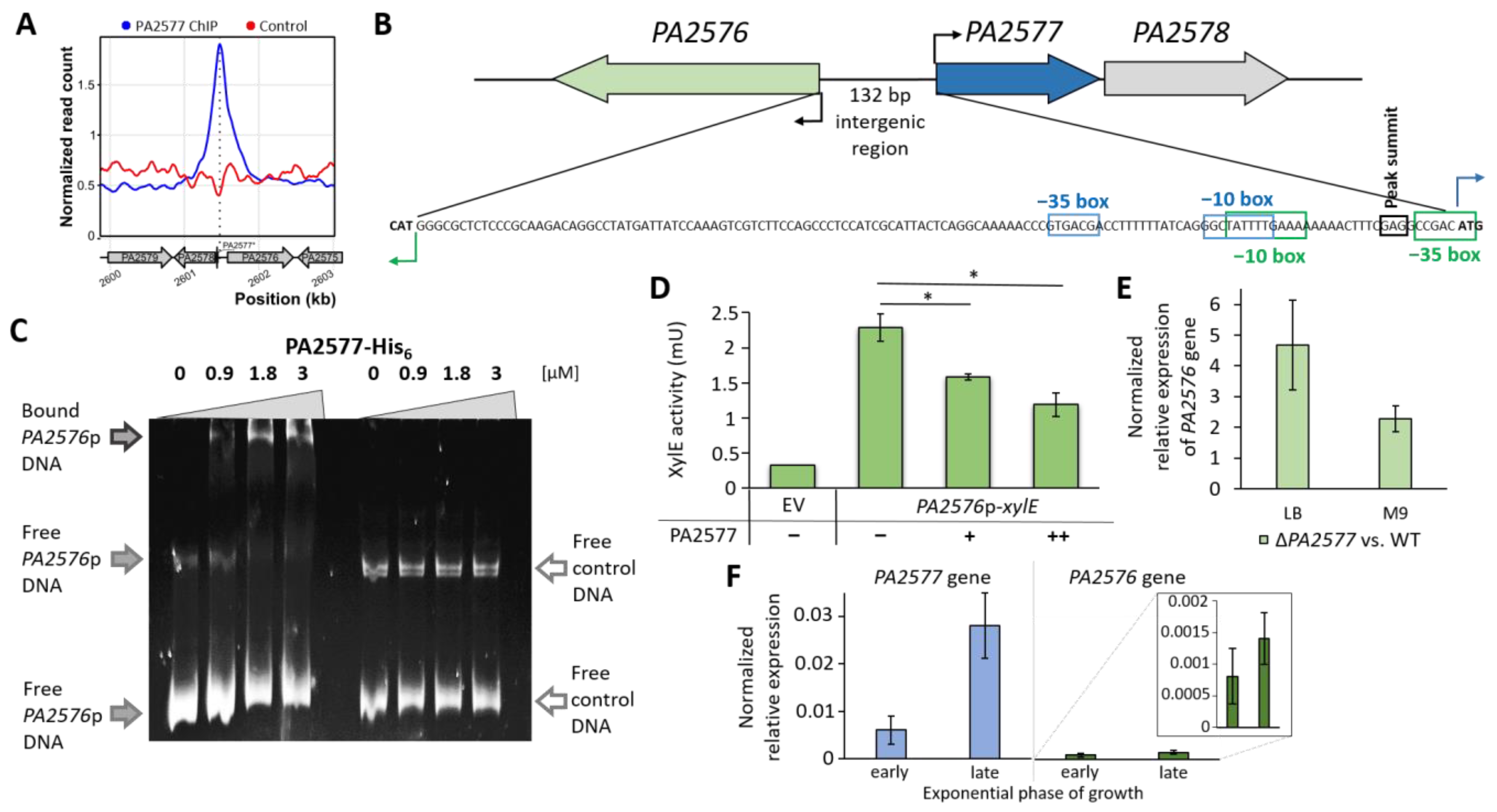
2.4. Identification of PA2577 Binding Sites in P. aeruginosa Genome
2.5. Regulatory Properties of PA2577
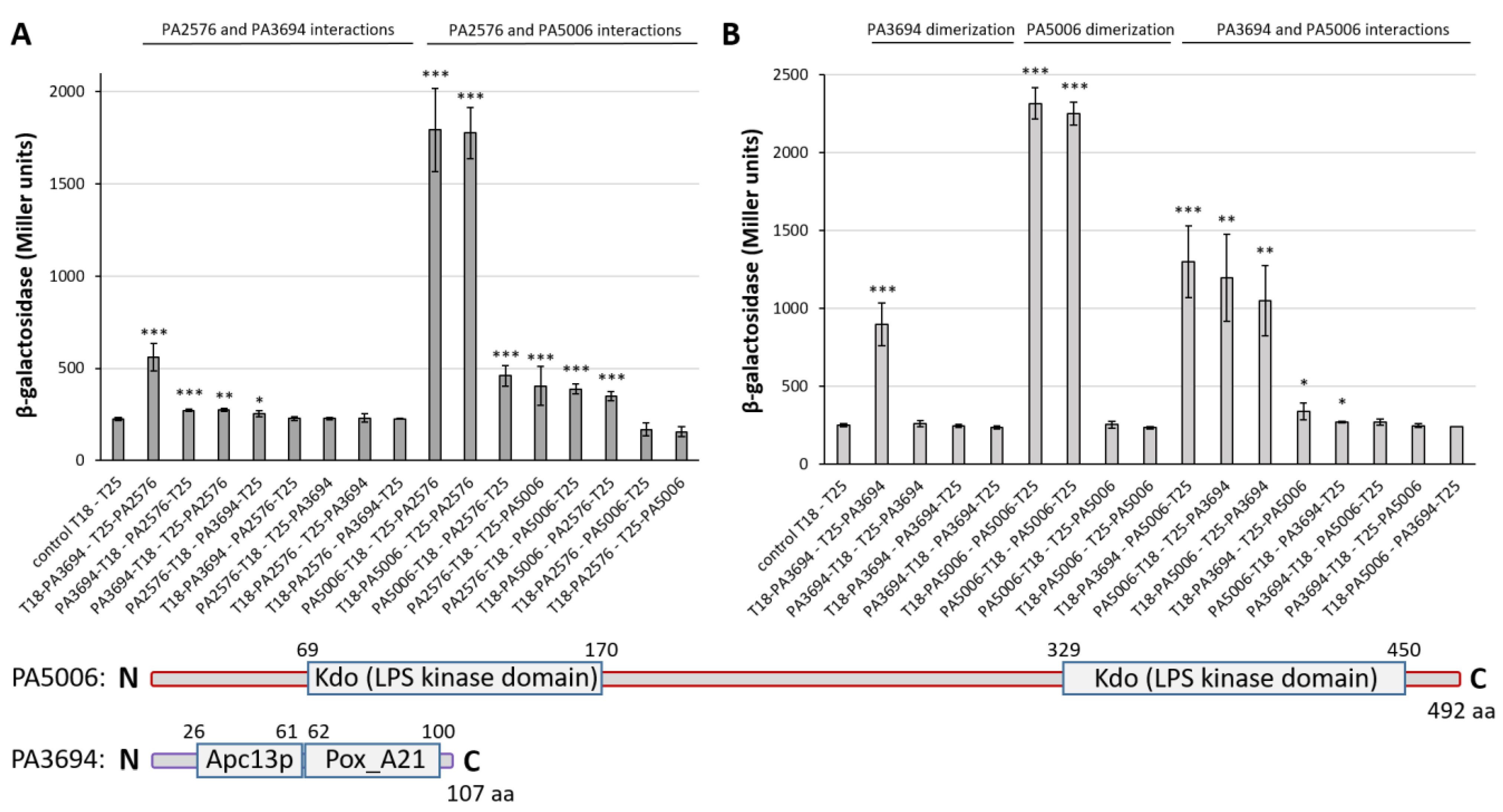
2.6. Characterization of PA2577 Target Gene PA2576
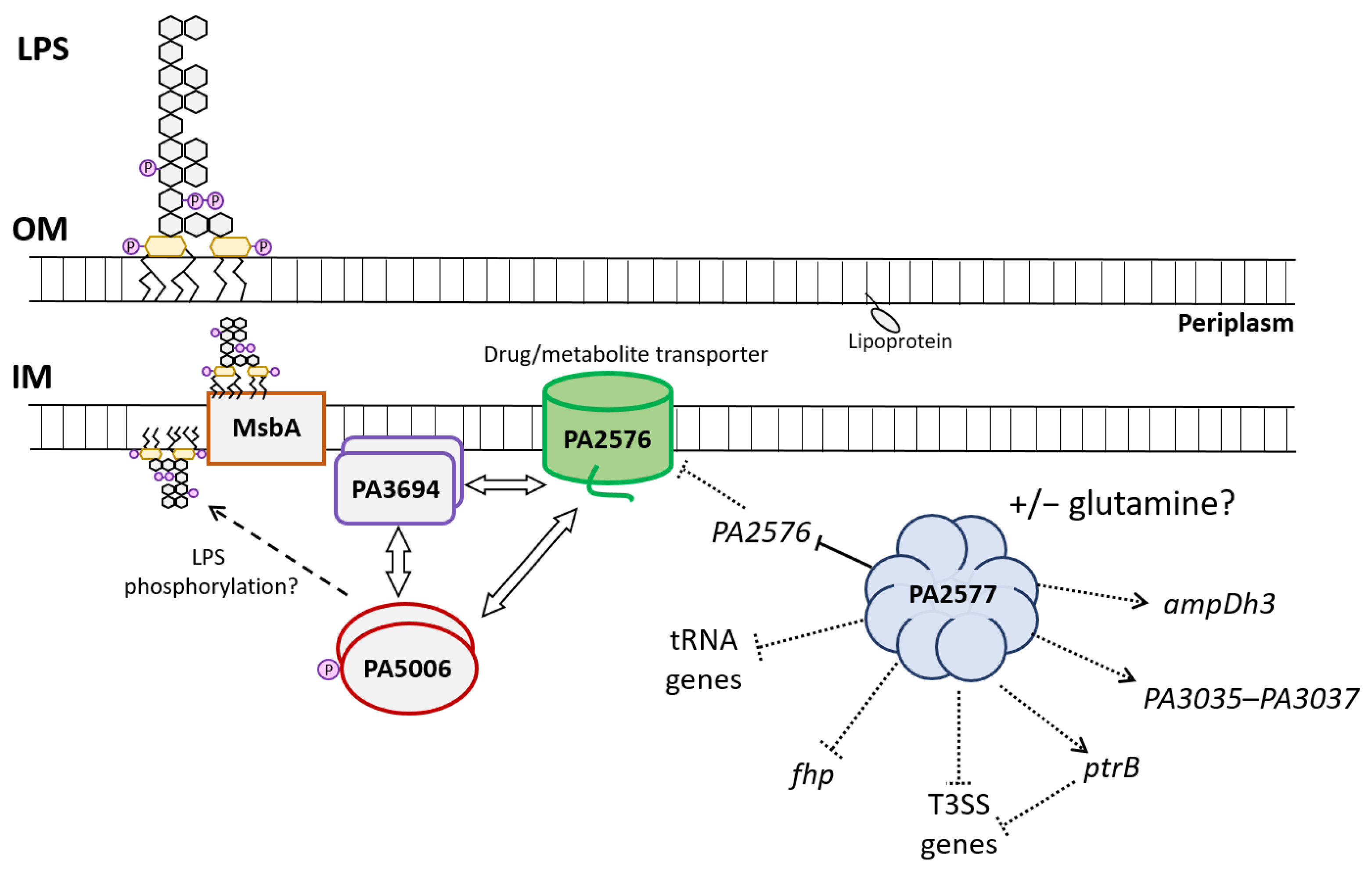
2.7. Characteristics of PA2577 Network and Its Role in P. aeruginosa
3. Discussion
4. Materials and Methods
4.1. Growth Conditions and Bacterial Strains and Plasmids
4.2. Growth Experiments
4.3. Analysis of Protein–Protein Interactions in the Bacterial Two-Hybrid System
4.4. Motility and Biofilm Formation Assays
4.5. RNA Isolation, RNA-Seq and RT-qPCR
4.6. Chromatin Immunoprecipitation with Sequencing
4.7. Protein Overproduction and Purification
4.8. Cross-Linking with Glutaraldehyde
4.9. In Vitro Protein–DNA Interactions
4.10. Regulatory Experiments and Promoter Activity Tests
4.11. Screening for Protein Partners
4.12. Phenotype Microarrays Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | Description | Reference |
| Escherichia coli | ||
| DH5α | F− Φ80lacZΔM15 Δ(lacZYA-argF) U169 recA1 endA1 hsdR17 (rK−, mK+) phoA supE44 λ thi-1 gyrA96 relA1 | [88] |
| S17-1 | pro ΔhsdR hsdM+ recA TpR SmR ΩRP4-Tc::Mu Kn::Tn7 | [89] |
| BL21 | F− ompT hsdSB (rB−mB−) gal dcm (λ DE3) | Novagen |
| BTH101 | F− cya-99 araD139 galE15 galK16 rpsL1 (StrR) hsdR2 mcrA1 mcrB1 | [37] |
| Pseudomonas aeruginosa | ||
| PAO1161 | PAO1161 RifR leu−, r−, m− | [30] |
| PAO1161 | PAO1161 RifR r−, m− | [90] |
| PAO1161 ΔPA2577 | PAO1161 RifR with deleted gene PA2577, allele exchange with the use of pMEB17 | This study |
| PAO1161 ΔPA2576 | PAO1161 RifR with deleted gene PA2576, allele exchange with the use of pMEB164 | This study |
| Plasmid Name | Description | Reference |
| pBBR1-MCS-1 | CmR, IncA/C broad-host-range cloning vector, lacZα–MCS, mob, T7p, T3p | [91] |
| pAMB9.37 | pBBR1-MCS-1 derivative with lacIQ-tacp, expression vector | [92] |
| pABB28.1 | pBBR1-MCS-1 derivative with lacIQ-tacp-flag, expression vector | [82] |
| pAKE600 | ApR, oriMB1, oriTRK2, sacB, suicide vector | [93] |
| pBGS18 | KmR, oriMB1, cloning vector | [94] |
| pCM132 | KmR, oriCColE1, oriVIncP, oriT, traJ’, trfA, broad-host-range vector, promoter-less lacZ reporter gene | [95] |
| pET28a(+) | KmR, oriMB1, T7p, lacO, His6-tag, T7 tag, expression vector | Novagen |
| pPTOI | KmR, oriVpSC101, promoter-less xylE cassette | [96] |
| pET28mod | KmR, oriMB1, T7p, lacO, His6-tag, modified to remove T7 tag | [97] |
| pKAB240 | ApR, oriMB1, pUC19 derivative with His6-mcs (MunI, HindIII, NotI, XhoI, BamHI)-flag | [82] |
| pKGB8 | CmR, broad-host-range expression vector, oriIncA/C, araC-araBADp | [39] |
| pUT18C | ApR; oriColE1, lacp–cyaT18–mcs | [37] |
| pUT18 | ApR; oriColE1, lacp–mcs–cyaT18 | [37] |
| pKT25 | KmR, orip15, lacp–cyaT25–mcs | [37] |
| pKNT25 | KmR; orip15, lacp–mcs–cyaT25 | [37] |
| pLKB4 | pUT18C with modified mcs | [98] |
| pKGB4 | pUT18 with modified mcs | [98] |
| pLKB2 | pKT25 with modified mcs | [98] |
| pKGB5 | pKNT25 with modified mcs | [98] |
| pMEB3 | pET28mod derivative, containing PA2577 gene inserted as EcoRI-SacI fragment amplified with the use of #1 and #2 primers to obtain His6-PA2577 fusion | This study |
| pMEB6 | pAKE600 derivative with upstream region of PA2577 gene inserted as EcoRI-HindIII fragment amplified with the use of #3 and #4 primers | This study |
| pMEB11 | pBGS18 derivative with downstream region of PA2577 gene inserted as HindIII-BamHI fragment amplified with the use of #5 and #6 primers | This study |
| pMEB12 | pKGB8 derivative, containing PA2577 gene under the control of BADp promoter; PA2577 gene from pMEB3 inserted as EcoRI-SacI fragment | This study |
| pMEB17 | pAKE600 derivative with fused upstream and downstream region of PA2577 gene; downstream region from pMEB11 inserted into pMEB6 as HindIII-BamHI creating up and down fusion linked with HindIII | This study |
| pMEB58 | pKGB4 derivative containing PA2577 gene without stop codon fused with T18; PA2577 amplified with the use of primers #1 and #18 inserted as EcoRI-KpnI fragment | This study |
| pMEB61 | pKGB5 derivative containing PA2577 gene without stop codon fused with T25; PA2577 amplified with the use of primers #1 and #18 inserted as EcoRI-KpnI fragment | This study |
| pMEB64 | pAMB9.37 derivative with inserted EcoRI-XhoI fragment containing PA2577 gene (cloned from pMEB3) under control of tacp promoter | This study |
| pMEB67 | pLKB2 derivative encoding T25-PA2577 translational fusion; PA2577 gene from pMEB64 inserted as EcoRI-KpnI fragment | This study |
| pMEB69 | pLKB4 derivative encoding T18-PA2577 translational fusion; PA2577 gene from pMEB64 inserted as EcoRI-KpnI fragment | This study |
| pMEB78 | pCM132 derivative with 192 bps fragment containing intergenic region upstream of PA2576 amplified with the use of #16 and #17 primers fused with lacZ to obtain transcriptional fusion | This study |
| pMEB79 | pKAB240 derivative with PA2577 gene fused with flag-tag; PA2577 gene without stop codon amplified with the use of #1 and #7 primers inserted as EcoRI-XhoI fragment | This study |
| pMEB83 | pAMB9.37 with modified mcs (EcoRI-PstI-SmaI-BglII-SalI inserted in the place of EcoRI-XhoI fragment) | This study |
| pMEB84 | pMEB83 with inserted EcoRI-SmaI fragment from pMEB79 containing PA2577-flag fusion | This study |
| pMEB105 | pET28mod derivative, containing PA2577 gene inserted as NcoI-XhoI fragment amplified with the use of #8 and #9 primers to obtain PA2577-His6 fusion | This study |
| pMEB121 | pKGB5 derivative containing PA2576 gene without stop codon fused with T25; PA2576 amplified with the use of primers #19 and #24 inserted as EcoRI-BamHI fragment | This study |
| pMEB122 | pLKB2 derivative encoding T25-PA2576 translational fusion; PA2576 gene inserted as EcoRI-BamHI fragment | This study |
| pMEB156 | pAKE600 derivative with upstream region of PA2576 gene inserted as EcoRI-HindIII fragment amplified with the use of #10 and #11 primers | This study |
| pMEB157 | pBGS18 derivative with downstream region of PA2576 gene inserted as HindIII-BamHI fragment amplified with the use of #12 and #13 primers | This study |
| pMEB164 | pAKE600 derivative with fused upstream and downstream region of PA2576 gene; downstream region from pMEB157 inserted into pMEB156 as HindIII-BamHI creating up and down fusion linked with HindIII | This study |
| pMEB185 | pAMB9.37 derivative with inserted EcoRI-KpnI fragment containing PA2578 gene amplified with the use of #25 and #26 under control of tacp promoter | This study |
| pMEB186 | pAMB9.37 derivative with inserted EcoRI-KpnI fragment containing PA2577–PA2578 operon amplified with the use of #1 and #26 under control of tacp promoter | This study |
| pMEB189 | pPTOI derivative with 192 bps fragment containing intergenic region upstream of PA2576 amplified with the use of #16 and #17 primers fused with xylE to obtain transcriptional fusion | This study |
| pMEB201 | pAMB9.37 derivative with inserted EcoRI-XhoI fragment containing PA2576 gene amplified with the use of #19 and #22 primers under control of tacp promoter | This study |
| pMEB239 | pLKB4 derivative encoding T18-PA3694 translational fusion; PA3694 gene inserted as EcoRI-SacI fragment | This study |
| pMEB240 | pLKB4 derivative encoding T18-PA5006 translational fusion; PA5006 gene inserted as BamHI-SacI fragment | This study |
| pMEB242 | pLKB2 derivative encoding T25-PA5006 translational fusion; PA5006 gene inserted as BamHI-SacI fragment | This study |
| pMEB246 | pKGB4 derivative containing PA5006 gene without stop codon fused with T18; PA5006 amplified with the use of primers #29 and #31 inserted as BamHI-SacI fragment | This study |
| pMEB247 | pKGB5 derivative containing PA5006 gene without stop codon fused with T25; PA5006 amplified with the use of primers #29 and #31 inserted as BamHI-SacI fragment | This study |
| pMEB248 | pKGB5 derivative containing PA3694 gene without stop codon fused with T25; PA3694 amplified with the use of primers #27 and #32 inserted as EcoRI-SacI fragment | This study |
| pMEB249 | pKGB4 derivative containing PA3694 gene without stop codon fused with T18; PA3694 amplified with the use of primers #27 and #32 inserted as EcoRI-SacI fragment | This study |
| pMEB250 | pLKB2 derivative encoding T25-PA3694 translational fusion; PA3694 gene inserted as EcoRI-SacI fragment | This study |
| Primers Used to Amplify DNA Fragments for Cloning | ||
|---|---|---|
| No. | Name | Sequence 5′-3′ |
| #1 | 2577ESF | GC GAATTC ATGACCGATGACATCGACC |
| #2 | 2577ESR | GC GAGCTC GAACCAGCGTTGCAGGATTG |
| #3 | 2577EHuF | GC GAATTC CCGAATACGCAGCGCCAGAAC |
| #4 | 2577EHuR | GC AAGCTT TCATCAATCGGTCATGTCGGCCTCG |
| #5 | 2577HBdF | GC AAGCTT TGAGCGAGGCACTCCGCATGC |
| #6 | 2577HBdR | GC GGATCC GCCGTTGCGGCGGTCGAACAG |
| #7 | 2577EX | CG CTCGAG GGTCATCGGGGGGAGCCG |
| #8 | 2577NXF | GC CCATGG ATGACCGATGACATCGACC |
| #9 | 2577NXR | CG CTCGAG ACCGGAACCGGATCCGGTCATCGGGGGGAGCCG |
| #10 | 2576EHuF | GC GAATTC CGTTCTTCCAGCTTGCGCAG |
| #11 | 2576EHuR | GCC AAGCTT TCA ATCGTT CAT GGGCGCTCTC |
| #12 | 2576HBdF | GCC AAGCTT TGA GGCGCGGGGGAAAGAAAAAG |
| #13 | 2576HBdR | GC GGATCC GTACTGTTCTTCGAAGACCAG |
| #14 | p2577EBF | GC GAATTC GCATGC CCAGCGCGGCACAGGCG |
| #15 | p2577EBR | GC GGATCC CATGTCGGCCTCGAAAG |
| #16 | p2576F | GC GAATTC GCATGC GGGACAGTCGCGAGTCAGC |
| #17 | p2576R | GC GGATCC GGCGCTCTCCCGCAAGACAG |
| #18 | 2577EK | CG GGTACC C GGTCATCGGGGGGAGCCG |
| #19 | 2576Ef | CG GAATTC ATGAACGATCCGATCCGTC |
| #20 | BADNsiIF | ACGGATGGCCTTTATGCATTTCTACAAACT |
| #21 | CM132RCy5 | Cy5-CTTCCACAGTAGTTCACCACC |
| #22 | 2576EXr | CG CTCGAG GAAAAGCCCCGCCTCGTCG |
| #23 | 2576EBrS | CG GGATCC GAAAAGCCCCGCCTCGTCG |
| #24 | 2576EBr-S | CG GGATCC C GCCGCGACGCACCGCCGGC |
| #25 | 2578EKf | CG GAATTC ATGCCCACACCCGGCACGG |
| #26 | 2578EKR | GC GGTACC GCTCCGTTCCAACCTGTAG |
| #27 | 3694ESf | CG GAATTC ATGAGCGACCGTAACGTAC |
| #28 | 3694ESr | AT GAGCTC AGACGGGTGATGTCTCCCT |
| #29 | 5006Bf | GC GGATCC ATGAGACTGGCCGAACTGC |
| #30 | 5006BSr | GA GAGCTC GGTGGTCGTCGCTTGTCGT |
| #31 | 5006BSr-S | AT GAGCTC C AGCCCCGGCGGCGTCGCCGA |
| #32 | 3694ESr-S | AT GAGCTC CTTTCACTTCCTGCGGGACT |
| Primers used in RT-qPCR analysis | ||
| #33 | rpsLF | CTCGGCACTGCGTAAGGTAT |
| #34 | rpsLR | TGTGCTCTTGCAGGTTGTGA |
| #35 | nadBF | CTACCTGGACATCAGCCACA |
| #36 | nadBR | GGTAATGTCGATGCCGAAGT |
| #37 | PA2576qF | CCTGGGAATCGTCCATACCG |
| #38 | PA2576qR | CGCGACCGGATAGATGAAGG |
| #39 | PA1698qF | GGAAAGCCAGCAGAAGCTC |
| #40 | PA1698qR | GGAGAAGCCCTCCAGGTAAG |
| #41 | PA3037qF | GACGAAAGCCTCGACCTGT |
| #42 | PA3037qR | GTCGAGGGTGACGAACAGAC |
| #43 | PA0807qF | CGACAACCTCAACGACACC |
| #44 | PA0807qR | GTAGTCGGGGAAGGTGAACA |
References
- Tani, T.H.; Khodursky, A.; Blumenthal, R.M.; Brown, P.O.; Matthews, R.G. Adaptation to Famine: A Family of Stationary-Phase Genes Revealed by Microarray Analysis. Proc. Natl. Acad. Sci. USA 2002, 99, 13471–13476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroner, G.M.; Wolfe, M.B.; Freddolino, P.L. Escherichia coli Lrp Regulates One-Third of the Genome via Direct, Cooperative, and Indirect Routes. J. Bacteriol. 2021, 201, e00411-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kölling, R.; Lother, H. AsnC: An Autogenously Regulated Activator of Asparagine Synthetase A Transcription in Escherichia coli. J. Bacteriol. 1985, 164, 310–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willins, D.A.; Ryan, C.W.; Platko, J.V.; Calvo, J.M. Characterization of Lrp, and Escherichia coli Regulatory Protein That Mediates a Global Response to Leucine. J. Biol. Chem. 1991, 266, 10768–10774. [Google Scholar] [CrossRef]
- Keuntje, B.; Masepohl, B.; Klipp, W. Expression of the putA Gene Encoding Proline Dehydrogenase from Rhodobacter capsulatus Is Independent of NtrC Regulation but Requires an Lrp-like Activator Protein. J. Bacteriol. 1995, 177, 6432–6439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madhusudhan, K.T.; Huang, N.; Sokatch, J.R. Characterization of BkdR-DNA Binding in the Expression of the bkd Operon of Pseudomonas putida. J. Bacteriol. 1995, 177, 636–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belitsky, B.R.; Gustafsson, M.C.; Sonenshein, A.L.; Von Wachenfeldt, C. An Lrp-like Gene of Bacillus subtilis Involved in Branched-Chain Amino Acid Transport. J. Bacteriol. 1997, 179, 5448–5457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, H.; Inagaki, K.; Eriguchi, S.I.; Tamura, T.; Esaki, N.; Soda, K.; Tanaka, H. Molecular Characterization of the mde Operon Involved in L-Methionine Catabolism of Pseudomonas putida. J. Bacteriol. 1997, 179, 3956–3962. [Google Scholar] [CrossRef] [Green Version]
- Friedberg, D.; Midkiff, M.; Calvo, J.M. Global versus Local Regulatory Roles for Lrp-Related Proteins: Haemophilus influenzae as a Case Study. J. Bacteriol. 2001, 183, 4004–4011. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, K.; Ishijima, S.A.; Clowney, L.; Koike, H.; Aramaki, H.; Tanaka, C.; Makino, K.; Suzuki, M. Feast/Famine Regulatory Proteins (FFRPs): Escherichia coli Lrp, AsnC and Related Archaeal Transcription Factors. FEMS Microbiol. Rev. 2006, 30, 89–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, K.; Winans, S.C. The putA Gene of Agrobacterium tumefaciens Is Transcriptionally Activated in Response to Proline by an Lrp-like Protein and Is Not Autoregulated. Mol. Microbiol. 1996, 22, 1025–1033. [Google Scholar] [CrossRef]
- Peekhaus, N.; Tolner, B.; Poolman, B.; Krämer, R. The Glutamate Uptake Regulatory Protein (Grp) of Zymomonas mobilis and Its Relation to the Global Regulator Lrp of Escherichia coli. J. Bacteriol. 1995, 177, 5140–5147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, H.; Ishijima, S.A.; Clowney, L.; Suzuki, M. The Archaeal Feast/Famine Regulatory Protein: Potential Roles of Its Assembly Forms for Regulating Transcription. Proc. Natl. Acad. Sci. USA 2004, 101, 2840–2845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaw, P.; Sedelnikova, S.E.; Muranova, T.; Wiese, S.; Ayora, S.; Alonso, J.C.; Brinkman, A.B.; Akerboom, J.; van der Oost, J.; Rafferty, J.B. Structural Insight into Gene Transcriptional Regulation and Effector Binding by the Lrp/AsnC Family. Nucleic Acids Res. 2006, 34, 1439–1449. [Google Scholar] [CrossRef] [Green Version]
- Leonard, P.M.; Smits, S.H.J.; Sedelnikova, S.E.; Brinkman, A.B.; de Vos, W.M.; van der Oost, J.; Rice, D.W.; Rafferty, J.B. Crystal Structure of the Lrp-like Transcriptional Regulator from the Archaeon Pyrococcus furiosus. EMBO J. 2001, 20, 990–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, M.C.M.; Gokulan, K.; Jacobs, W.R.; Ioerger, T.R.; Sacchettini, J.C. Crystal Structure of Mycobacterium tuberculosis LrpA, a Leucine-Responsive Global Regulator Associated with Starvation Response. Protein Sci. 2008, 17, 159–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, M.; Ishijima, S.A.; Suzuki, M. Interactions between the Archaeal Transcription Repressor FL11 and Its Coregulators Lysine and Arginine. Proteins 2009, 74, 520–525. [Google Scholar] [CrossRef] [PubMed]
- De los Rios, S.; Perona, J.J. Structure of the Escherichia coli Leucine-Responsive Regulatory Protein Lrp Reveals a Novel Octameric Assembly. J. Mol. Biol. 2007, 366, 1589–1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.; Zhen, J.; Li, Y.; Huang, Y.; Ai, X.; Li, Y.; Stojkoska, A.; Huang, X.; Ruan, C.; Li, J.; et al. Mycobacterium Lrp/AsnC Family Transcriptional Factor Modulates the Arginase Pathway as Both a Sensor and a Transcriptional Repressor. J. Genet. Genom. 2021, 48, 1020–1031. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Wang, H.; Xie, J. Regulatory and Pathogenesis Roles of Mycobacterium Lrp/AsnC Family Transcriptional Factors. J. Cell. Biochem. 2011, 112, 2655–2662. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, K.; Ishijima, S.A.; Koike, H.; Kurihara, C.; Shimowasa, A.; Kabasawa, M.; Kawashima, T.; Suzuki, M. Feast/Famine Regulation by Transcription Factor FL11 for the Survival of the Hyperthermophilic Archaeon Pyrococcus OT3. Structure 2007, 15, 1542–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashworth, J.; Plaisier, C.L.; Lo, F.Y.; Reiss, D.J.; Baliga, N.S. Inference of Expanded Lrp-like Feast/Famine Transcription Factor Targets in a Non-Model Organism Using Protein Structure-Based Prediction. PLoS ONE 2014, 9, e107863. [Google Scholar] [CrossRef] [PubMed]
- Bartosik, A.A.; Glabski, K.; Jecz, P.; Mikulska, S.; Fogtman, A.; Koblowska, M.; Jagura-Burdzy, G. Transcriptional Profiling of parA and parB Mutants in Actively Dividing Cells of an Opportunistic Human Pathogen Pseudomonas aeruginosa. PLoS ONE 2014, 9, e87276. [Google Scholar] [CrossRef]
- Galán-Vásquez, E.; Luna, B.; Martínez-Antonio, A. The Regulatory Network of Pseudomonas aeruginosa. Microb. Inform. Exp. 2011, 1, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramanian, D.; Schneper, L.; Kumari, H.; Mathee, K. A Dynamic and Intricate Regulatory Network Determines Pseudomonas aeruginosa Virulence. Nucleic Acids Res. 2013, 41, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.T.; Kwon, D.-H.; Hegazy, M.; Lu, C.-D. Transcriptome Analysis of Agmatine and Putrescine Catabolism in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2008, 190, 1966–1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.; Li, C.; Lu, C.-D. Regulation and Characterization of the DadRAX Locus for D-Amino Acid Catabolism in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2011, 193, 2107–2115. [Google Scholar] [CrossRef] [Green Version]
- Knoten, C.A.; Hudson, L.L.; Coleman, J.P.; Farrow, J.M.; Pesci, E.C. KynR, a Lrp/AsnC-Type Transcriptional Regulator, Directly Controls the Kynurenine Pathway in Pseudomonas aeruginosa. J. Bacteriol. 2011, 193, 6567–6575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lasocki, K.; Bartosik, A.A.; Mierzejewska, J.; Thomas, C.M.; Jagura-Burdzy, G. Deletion of the parA (soj) Homologue in Pseudomonas aeruginosa Causes ParB Instability and Affects Growth Rate, Chromosome Segregation, and Motility. J. Bacteriol. 2007, 189, 5762–5772. [Google Scholar] [CrossRef] [Green Version]
- Bartosik, A.A.; Mierzejewska, J.; Thomas, C.M.; Jagura-Burdzy, G. ParB Deficiency in Pseudomonas aeruginosa Destabilizes the Partner Protein ParA and Affects a Variety of Physiological Parameters. Microbiology 2009, 155, 1080–1092. [Google Scholar] [CrossRef] [Green Version]
- Brinkman, A.B.; Ettema, T.J.G.; Vos, W.M.D.; Oost, J.V.D. The Lrp Family of Transcriptional Regulators. Mol. Microbiol. 2003, 48, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Winsor, G.L.; Griffiths, E.J.; Lo, R.; Dhillon, B.K.; Shay, J.A.; Brinkman, F.S.L. Enhanced Annotations and Features for Comparing Thousands of Pseudomonas Genomes in the Pseudomonas Genome Database. Nucleic Acids Res. 2016, 44, D646–D653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirdita, M.; Ovchinnikov, S.; Steinegger, M. ColabFold-Making Protein Folding Accessible to All. bioRxiv 2021. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology Modelling of Protein Structures and Complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Guex, N.; Peitsch, M.C.; Schwede, T. Automated Comparative Protein Structure Modeling with SWISS-MODEL and Swiss-PdbViewer: A Historical Perspective. Electrophoresis 2009, 30, S162–S173. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Tao, H.; He, J.; Huang, S.-Y. The HDOCK Server for Integrated Protein–Protein Docking. Nat. Protoc. 2020, 15, 1829–1852. [Google Scholar] [CrossRef]
- Karimova, G.; Pidoux, J.; Ullmann, A.; Ladant, D. A Bacterial Two-Hybrid System Based on a Reconstituted Signal Transduction Pathway. Proc. Natl. Acad. Sci. USA 1998, 95, 5752–5756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkman, A.B.; Dahlke, I.; Tuininga, J.E.; Lammers, T.; Dumay, V.; de Heus, E.; Lebbink, J.H.; Thomm, M.; de Vos, W.M.; van Der Oost, J. An Lrp-like Transcriptional Regulator from the Archaeon Pyrococcus furiosus Is Negatively Autoregulated. J. Biol. Chem. 2000, 275, 38160–38169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawalek, A.; Glabski, K.; Bartosik, A.A.; Fogtman, A.; Jagura-Burdzy, G. Increased ParB Level Affects Expression of Stress Response, Adaptation and Virulence Operons and Potentiates Repression of Promoters Adjacent to the High Affinity Binding Sites parS3 and parS4 in Pseudomonas aeruginosa. PLoS ONE 2017, 12, e0181726. [Google Scholar] [CrossRef] [Green Version]
- Juan, C.; Moyá, B.; Pérez, J.L.; Oliver, A. Stepwise Upregulation of the Pseudomonas aeruginosa Chromosomal Cephalosporinase Conferring High-Level Beta-Lactam Resistance Involves Three AmpD Homologues. Antimicrob. Agents Chemother. 2006, 50, 1780–1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindberg, F.; Lindquist, S.; Normark, S. Inactivation of the AmpD Gene Causes Semiconstitutive Overproduction of the Inducible Citrobacter freundii Beta-Lactamase. J. Bacteriol. 1987, 169, 1923–1928. [Google Scholar] [CrossRef] [Green Version]
- Bonamore, A.; Boffi, A. Flavohemoglobin: Structure and Reactivity. IUBMB Life 2008, 60, 19–28. [Google Scholar] [CrossRef]
- Salamov, V.S.A.; Solovyevand, A. Automatic Annotation of Microbial Genomes and Metagenomic Sequences. In Metagenomics and Its Applications in Agriculture, Biomedicine and Environmental Studies; Li, R.W., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2011; pp. 61–78. [Google Scholar]
- Karimova, G.; Gauliard, E.; Davi, M.; Ouellette, S.P.; Ladant, D. Protein-Protein Interaction: Bacterial Two-Hybrid. In Bacterial Protein Secretion Systems; Humana Press: New York, NY, USA, 2017; pp. 159–176. [Google Scholar] [CrossRef]
- Mitura, M.; Lewicka, E.; Godziszewska, J.; Adamczyk, M.; Jagura-Burdzy, G. Alpha-Helical Protein KfrC Acts as a Switch between the Lateral and Vertical Modes of Dissemination of Broad-Host-Range RA3 Plasmid from IncU (IncP-6) Incompatibility Group. Int. J. Mol. Sci. 2021, 22, 4880. [Google Scholar] [CrossRef]
- Kusiak, M.B. Molecular and Structural Analysis of parB Gene of Pseudomonas aeruginosa. Ph.D. Thesis, Department of Microbial Biochemistry IBB PAS, Warsaw, Poland, 2010. [Google Scholar]
- Delucia, A.M.; Six, D.A.; Caughlan, R.E.; Gee, P.; Hunt, I.; Lam, J.S.; Dean, C.R. Lipopolysaccharide (LPS) Inner-Core Phosphates Are Required for Complete LPS Synthesis and Transport to the Outer Membrane in Pseudomonas aeruginosa PAO1. mBio 2011, 2, e00142-11. [Google Scholar] [CrossRef] [Green Version]
- Perumal, D.; Sakharkar, K.R.; Tang, T.H.; Chow, V.T.K.; Lim, C.S.; Samal, A.; Sugiura, N.; Sakharkar, M.K. Cloning and Targeted Disruption of Two Lipopolysaccharide Biosynthesis Genes, kdsA and waaG, of Pseudomonas aeruginosa PAO1 by Site-Directed Mutagenesis. J. Mol. Microbiol. Biotechnol. 2010, 19, 169–179. [Google Scholar] [CrossRef]
- De Kievit, T.R.; Lam, J.S. Isolation and Characterization of Two Genes, waaC (rfaC) and waaF (rfaF), Involved in Pseudomonas aeruginosa Serotype O5 Inner-Core Biosynthesis. J. Bacteriol. 1997, 179, 3451–3457. [Google Scholar] [CrossRef] [Green Version]
- Kumarevel, T.; Nakano, N.; Ponnuraj, K.; Gopinath, S.C.B.; Sakamoto, K.; Shinkai, A.; Kumar, P.K.R.; Yokoyama, S. Crystal Structure of Glutamine Receptor Protein from Sulfolobus tokodaii Strain 7 in Complex with Its Effector L-Glutamine: Implications of Effector Binding in Molecular Association and DNA Binding. Nucleic Acids Res. 2008, 36, 4808–4820. [Google Scholar] [CrossRef] [Green Version]
- Kudo, N.; Allen, M.D.; Koike, H.; Katsuya, Y.; Suzuki, M. Crystallization and Secondary-Structure Determination of a Protein of the Lrp/AsnC Family from a Hyperthermophilic Archaeon. Acta Crystallogr. Sect. D Biol. Crystallogr. 2001, 57, 469–471. [Google Scholar] [CrossRef]
- Marasco, R.; Varcamonti, M.; La Cara, F.; Ricca, E.; De Felice, M.; Sacco, M. In Vivo Footprinting Analysis of Lrp Binding to the ilvIH Promoter Region of Escherichia coli. J. Bacteriol. 1994, 176, 5197–5201. [Google Scholar] [CrossRef] [Green Version]
- Beloin, C.; Jeusset, J.; Revet, B.; Mirambeau, G.; Le Hégarat, F.; Le Cam, E. Contribution of DNA Conformation and Topology in Right-Handed DNA Wrapping by the Bacillus subtilis LrpC Protein. J. Biol. Chem. 2003, 278, 5333–5342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, A.; Shree, S.; Pandey, S.K.; Tripathi, R.P.; Ramachandran, R. Crystal Structure of Mycobacterium tuberculosis H37Rv AldR (Rv2779c), a Regulator of the ald Gene. J. Biol. Chem. 2016, 291, 11967–11980. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Calvo, J.M. Lrp, a Major Regulatory Protein in Escherichia coli, Bends DNA and Can Organize the Assembly of a Higher-Order Nucleoprotein Structure. EMBO J. 1993, 12, 2495–2501. [Google Scholar] [CrossRef]
- Suzuki, M. The DNA-Binding Specificity of Eubacterial and Archaeal FFRPs. Proc. Jpn. Acad. Ser. B 2003, 79B, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Sacco, M.; Ricca, E.; Lago, C.T.; De Felice, M.; Calvo, J.M. Organization of Lrp-Binding Sites Upstream of ilvIH in Salmonella typhimurium. Mol. Microbiol. 1993, 7, 883–891. [Google Scholar] [CrossRef]
- Wang, Q.; Calvo, J.M. Lrp, a Global Regulatory Protein of Escherichia coli, Binds Co-Operatively to Multiple Sites and Activates Transcription of ilvIH. J. Mol. Biol. 1993, 229, 306–318. [Google Scholar] [CrossRef]
- Nou, X.; Braaten, B.; Kaltenbach, L.; Low, D.A. Differential Binding of Lrp to Two Sets of Pap DNA Binding Sites Mediated by Pap I Regulates Pap Phase Variation in Escherichia coli. EMBO J. 1995, 14, 5785–5797. [Google Scholar] [CrossRef]
- Liu, C.F.; Brandt, G.S.; Hoang, Q.Q.; Naumova, N.; Lazarevic, V.; Hwang, E.S.; Dekker, J.; Glimcher, L.H.; Ringe, D.; Petsko, G.A. Crystal Structure of the DNA Binding Domain of the Transcription Factor T-Bet Suggests Simultaneous Recognition of Distant Genome Sites. Proc. Natl. Acad. Sci. USA 2016, 113, E6572–E6581. [Google Scholar] [CrossRef] [Green Version]
- Cirz, R.T.; O’Neill, B.M.; Hammond, J.A.; Head, S.R.; Romesberg, F.E. Defining the Pseudomonas aeruginosa SOS Response and Its Role in the Global Response to the Antibiotic Ciprofloxacin. J. Bacteriol. 2006, 188, 7101–7110. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Jin, S. PtrB of Pseudomonas aeruginosa Suppresses the Type III Secretion System under the Stress of DNA Damage. J. Bacteriol. 2005, 187, 6058–6068. [Google Scholar] [CrossRef] [Green Version]
- Rietsch, A.; Vallet-Gely, I.; Dove, S.L.; Mekalanos, J.J. ExsE, a Secreted Regulator of Type III Secretion Genes in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2005, 102, 8006–8011. [Google Scholar] [CrossRef] [Green Version]
- Allocati, N.; Federici, L.; Masulli, M.; Di Ilio, C. Glutathione Transferases in Bacteria. FEBS J. 2009, 276, 58–75. [Google Scholar] [CrossRef]
- De Wind, N.; de Jong, M.; Meijer, M.; Stuitje, A.R. Site-Directed Mutagenesis of the Escherichia coli Chromosome near OriC: Identification and Characterization of AsnC, a Regulatory Element in E. coli Asparagine Metabolism. Nucleic Acids Res. 1985, 13, 8797–8811. [Google Scholar] [CrossRef] [Green Version]
- Saier, M.H.; Tran, C.V.; Barabote, R.D. TCDB: The Transporter Classification Database for Membrane Transport Protein Analyses and Information. Nucleic Acids Res. 2006, 34, D181–D186. [Google Scholar] [CrossRef] [Green Version]
- Västermark, Å.; Almén, M.S.; Simmen, M.W.; Fredriksson, R.; Schiöth, H.B. Functional Specialization in Nucleotide Sugar Transporters Occurred through Differentiation of the Gene Cluster EamA (DUF6) before the Radiation of Viridiplantae. BMC Evol. Biol. 2011, 11, 123. [Google Scholar] [CrossRef] [Green Version]
- Franke, I.; Resch, A.; Daßler, T.; Maier, T.; Böck, A. YfiK from Escherichia coli Promotes Export of O-Acetylserine and Cysteine. J. Bacteriol. 2003, 185, 1161–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouanet, C.; Nasser, W. The PecM Protein of the Phytopathogenic Bacterium Erwinia chrysanthemi, Membrane Topology and Possible Involvement in the Efflux of the Blue Pigment Indigoidine. J. Mol. Microbiol. Biotechnol. 2001, 3, 309–318. [Google Scholar]
- Hadley, B.; Litfin, T.; Day, C.J.; Haselhorst, T.; Zhou, Y.; Tiralongo, J. Nucleotide Sugar Transporter SLC35 Family Structure and Function. Comput. Struct. Biotechnol. J. 2019, 17, 1123–1134. [Google Scholar] [CrossRef]
- Ghanei, H.; Abeyrathne, P.D.; Lam, J.S. Biochemical Characterization of MsbA from Pseudomonas aeruginosa. J. Biol. Chem. 2007, 282, 26939–26947. [Google Scholar] [CrossRef] [Green Version]
- Williams McMackin, E.A.; Djapgne, L.; Corley, J.M.; Yahr, T.L. Fitting Pieces into the Puzzle of Pseudomonas aeruginosa Type III Secretion System Gene Expression. J. Bacteriol. 2019, 201, e00209-19. [Google Scholar] [CrossRef] [Green Version]
- Diaz, M.; King, J.; Yahr, T. Intrinsic and Extrinsic Regulation of Type III Secretion Gene Expression in Pseudomonas aeruginosa. Front. Microbiol. 2011, 2, 89. [Google Scholar] [CrossRef] [Green Version]
- Augustin, D.K.; Song, Y.; Baek, M.S.; Sawa, Y.; Singh, G.; Taylor, B.; Rubio-Mills, A.; Flanagan, J.L.; Wiener-Kronish, J.P.; Lynch, S.V. Presence or Absence of Lipopolysaccharide O Antigens Affects Type III Secretion by Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 2203–2209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Irani, V.R.; Rowe, J.J. Enhancement of Transformation in Pseudomonas aeruginosa PAO1 by Mg2+ and Heat. BioTechniques 1997, 22, 54–56. [Google Scholar] [CrossRef]
- Miller, J.H. Experiments in Molecular Genetics; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1972; ISBN 978-0-87969-106-6. [Google Scholar]
- Rashid, M.H.; Kornberg, A. Inorganic Polyphosphate Is Needed for Swimming, Swarming, and Twitching Motilities of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2000, 97, 4885–4890. [Google Scholar] [CrossRef] [Green Version]
- Kotecka, K.; Kawalek, A.; Kobylecki, K.; Bartosik, A.A. The MarR-Type Regulator PA3458 Is Involved in Osmoadaptation Control in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2021, 22, 3982. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Kawalek, A.; Bartosik, A.A.; Glabski, K.; Jagura-Burdzy, G. Pseudomonas aeruginosa Partitioning Protein ParB Acts as a Nucleoid-Associated Protein Binding to Multiple Copies of a parS-Related Motif. Nucleic Acids Res. 2018, 46, 4592–4606. [Google Scholar] [CrossRef] [Green Version]
- Modrzejewska, M.; Kawalek, A.; Bartosik, A.A. The LysR-Type Transcriptional Regulator BsrA (PA2121) Controls Vital Metabolic Pathways in Pseudomonas aeruginosa. mSystems 2021, 6, e00015-21. [Google Scholar] [CrossRef] [PubMed]
- Jagura-Burdzy, G.; Thomas, C.M. Purification of KorA Protein from Broad Host Range Plasmid RK2: Definition of a Hierarchy of KorA Operators. J. Mol. Biol. 1995, 253, 39–50. [Google Scholar] [CrossRef]
- Zukowski, M.M.; Gaffney, D.F.; Speck, D.; Kauffmann, M.; Findeli, A.; Wisecup, A.; Lecocq, J.P. Chromogenic Identification of Genetic Regulatory Signals in Bacillus subtilis Based on Expression of a Cloned Pseudomonas Gene. Proc. Natl. Acad. Sci. USA 1983, 80, 1101–1105. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Pawłowska, J.; Aleksandrzak-Piekarczyk, T.; Banach, A.; Kiersztyn, B.; Muszewska, A.; Serewa, L.; Szatraj, K.; Wrzosek, M. Preliminary Studies on the Evolution of Carbon Assimilation Abilities within Mucorales. Fungal Biol. 2016, 120, 752–763. [Google Scholar] [CrossRef]
- Bochner, B.R.; Gadzinski, P.; Panomitros, E. Phenotype MicroArrays for High-Throughput Phenotypic Testing and Assay of Gene Function. Genome Res. 2001, 11, 1246–1255. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D. Studies on Transformation of Escherichia coli with Plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Simon, R.; Priefer, U.; Pühler, A. A Broad Host Range Mobilization System for in vivo Genetic Engineering: Transposon Mutagenesis in Gram Negative Bacteria. Bio/Technology 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Kawalek, A.; Kotecka, K.; Modrzejewska, M.; Gawor, J.; Jagura-Burdzy, G.; Bartosik, A.A. Genome Sequence of Pseudomonas aeruginosa PAO1161, a PAO1 Derivative with the ICEPae1161 Integrative and Conjugative Element. BMC Genom. 2020, 21, 14. [Google Scholar] [CrossRef]
- Kovach, M.E.; Phillips, R.W.; Elzer, P.H.; Roop, R.M.; Peterson, K.M. PBBR1MCS: A Broad-Host-Range Cloning Vector. BioTechniques 1994, 16, 800–802. [Google Scholar] [PubMed]
- Ludwiczak, M.; Dolowy, P.; Markowska, A.; Szarlak, J.; Kulinska, A.; Jagura-Burdzy, G. Global Transcriptional Regulator KorC Coordinates Expression of Three Backbone Modules of the Broad-Host-Range RA3 Plasmid from IncU Incompatibility Group. Plasmid 2013, 70, 131–145. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, A.K.; Hothersall, J.; Thomas, C.M. Quorum-Sensing-Dependent Regulation of Biosynthesis of the Polyketide Antibiotic Mupirocin in Pseudomonas fluorescens NCIMB 10586. Microbiol. Read. Engl. 2001, 147, 2127–2139. [Google Scholar] [CrossRef] [Green Version]
- Spratt, B.G.; Hedge, P.J.; te Heesen, S.; Edelman, A.; Broome-Smith, J.K. Kanamycin-Resistant Vectors That Are Analogues of Plasmids PUC8, PUC9, PEMBL8 and PEMBL9. Gene 1986, 41, 337–342. [Google Scholar] [CrossRef]
- Marx, C.J.; Lidstrom, M.E. Development of Improved Versatile Broad-Host-Range Vectors for Use in Methylotrophs and Other Gram-Negative Bacteria. Microbiol. Read. Engl. 2001, 147, 2065–2075. [Google Scholar] [CrossRef] [Green Version]
- Thorsted, P.B.; Shah, D.S.; Macartney, D.; Kostelidou, K.; Thomas, C.M. Conservation of the Genetic Switch between Replication and Transfer Genes of IncP Plasmids but Divergence of the Replication Functions Which Are Major Host-Range Determinants. Plasmid 1996, 36, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Lukaszewicz, M.; Kostelidou, K.; Bartosik, A.A.; Cooke, G.D.; Thomas, C.M.; Jagura-Burdzy, G. Functional Dissection of the ParB Homologue (KorB) from IncP-1 Plasmid RK2. Nucleic Acids Res. 2002, 30, 1046–1055. [Google Scholar] [CrossRef]
- Bartosik, A.A.; Glabski, K.; Jecz, P.; Lasocki, K.; Mikosa, M.; Plochocka, D.; Thomas, C.M.; Jagura-Burdzy, G. Dissection of the Region of Pseudomonas aeruginosa ParA That Is Important for Dimerization and Interactions with Its Partner ParB. Microbiol. Read. Engl. 2014, 160, 2406–2420. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Modrzejewska, M.; Kawalek, A.; Bartosik, A.A. The Lrp/AsnC-Type Regulator PA2577 Controls the EamA-like Transporter Gene PA2576 in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2021, 22, 13340. https://doi.org/10.3390/ijms222413340
Modrzejewska M, Kawalek A, Bartosik AA. The Lrp/AsnC-Type Regulator PA2577 Controls the EamA-like Transporter Gene PA2576 in Pseudomonas aeruginosa. International Journal of Molecular Sciences. 2021; 22(24):13340. https://doi.org/10.3390/ijms222413340
Chicago/Turabian StyleModrzejewska, Magdalena, Adam Kawalek, and Aneta Agnieszka Bartosik. 2021. "The Lrp/AsnC-Type Regulator PA2577 Controls the EamA-like Transporter Gene PA2576 in Pseudomonas aeruginosa" International Journal of Molecular Sciences 22, no. 24: 13340. https://doi.org/10.3390/ijms222413340
APA StyleModrzejewska, M., Kawalek, A., & Bartosik, A. A. (2021). The Lrp/AsnC-Type Regulator PA2577 Controls the EamA-like Transporter Gene PA2576 in Pseudomonas aeruginosa. International Journal of Molecular Sciences, 22(24), 13340. https://doi.org/10.3390/ijms222413340