
Time to Classify Tumours of the Stomach and the Kidneys According to Cell of Origin



Abstract
:1. Introduction
2. Tumour Biology
3. Cell of Origin of Tumours
4. Neuroendocrine Cells
5. Tumorigenesis Is Due to Genetic Changes
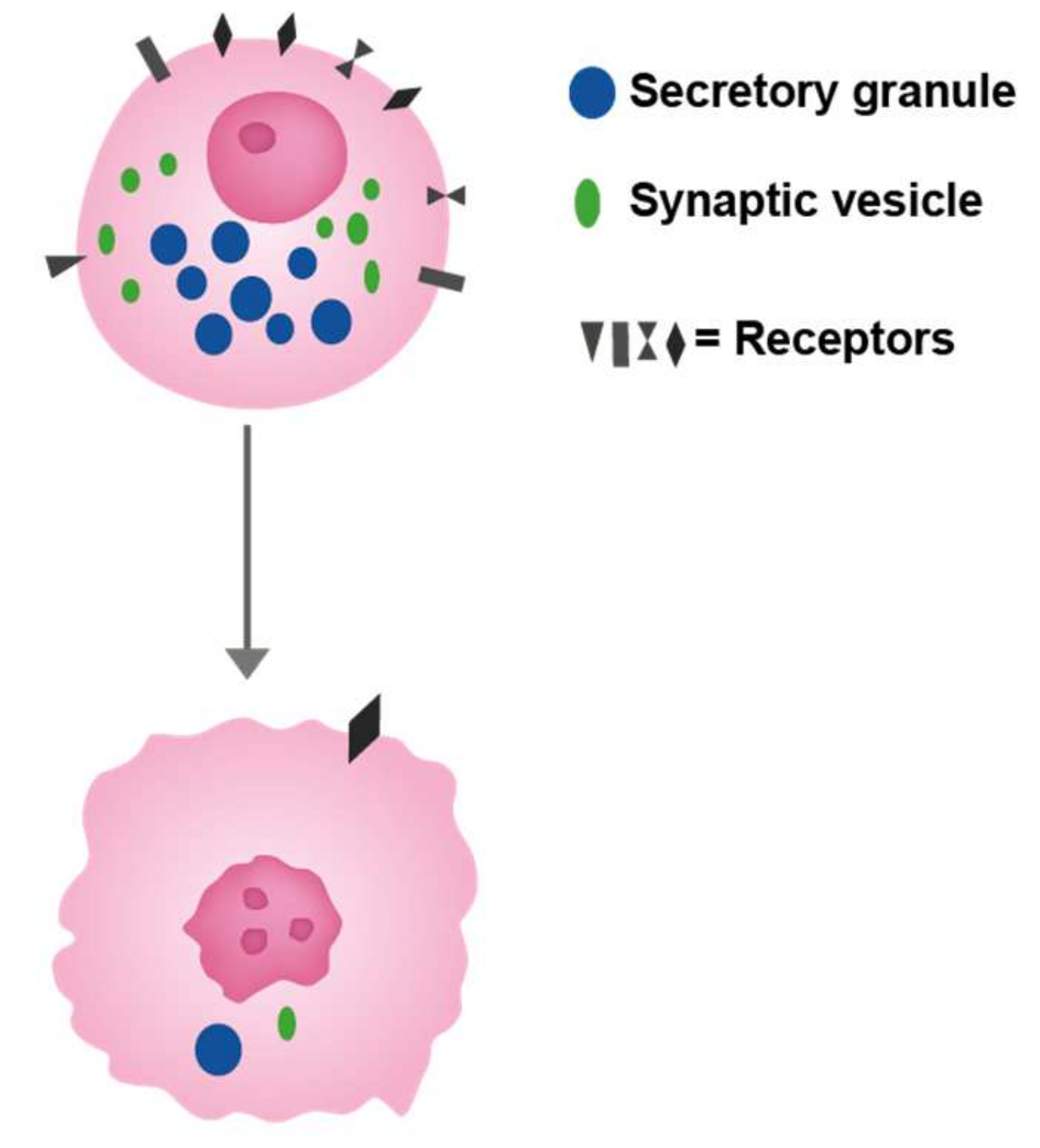
6. Specificity of Cellular Markers
7. Diagnostic Consequences of Improved Methods in Pathology
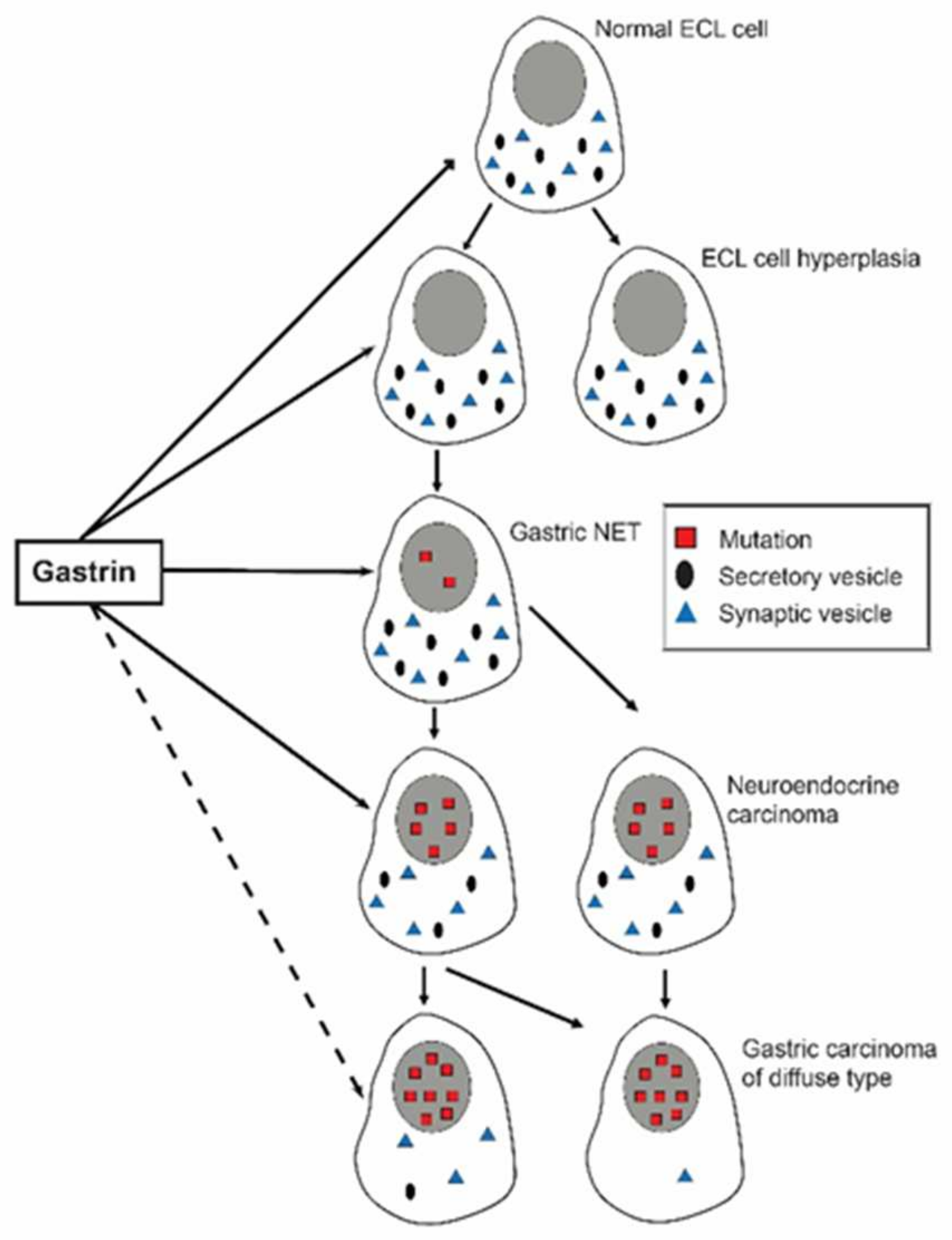
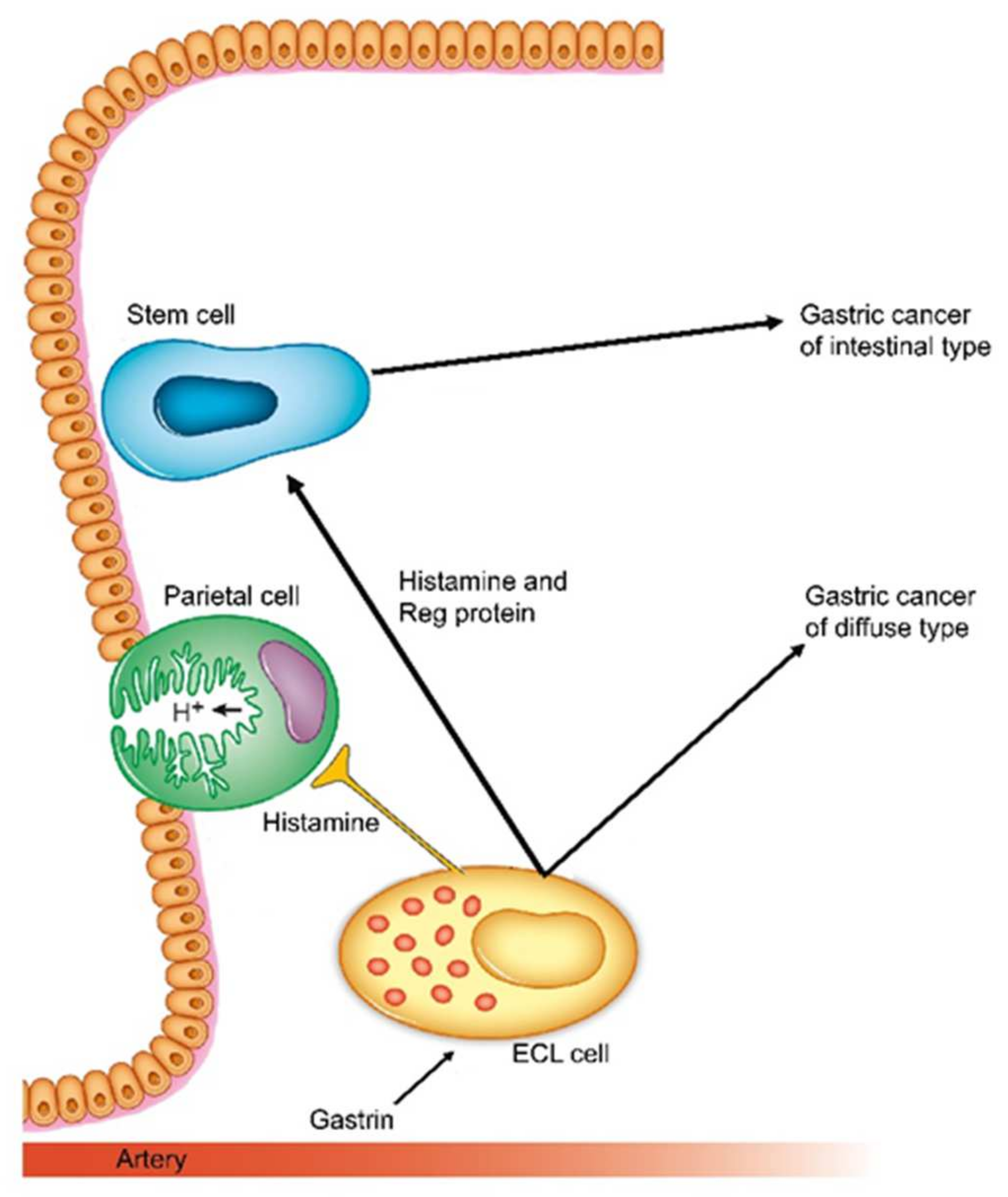
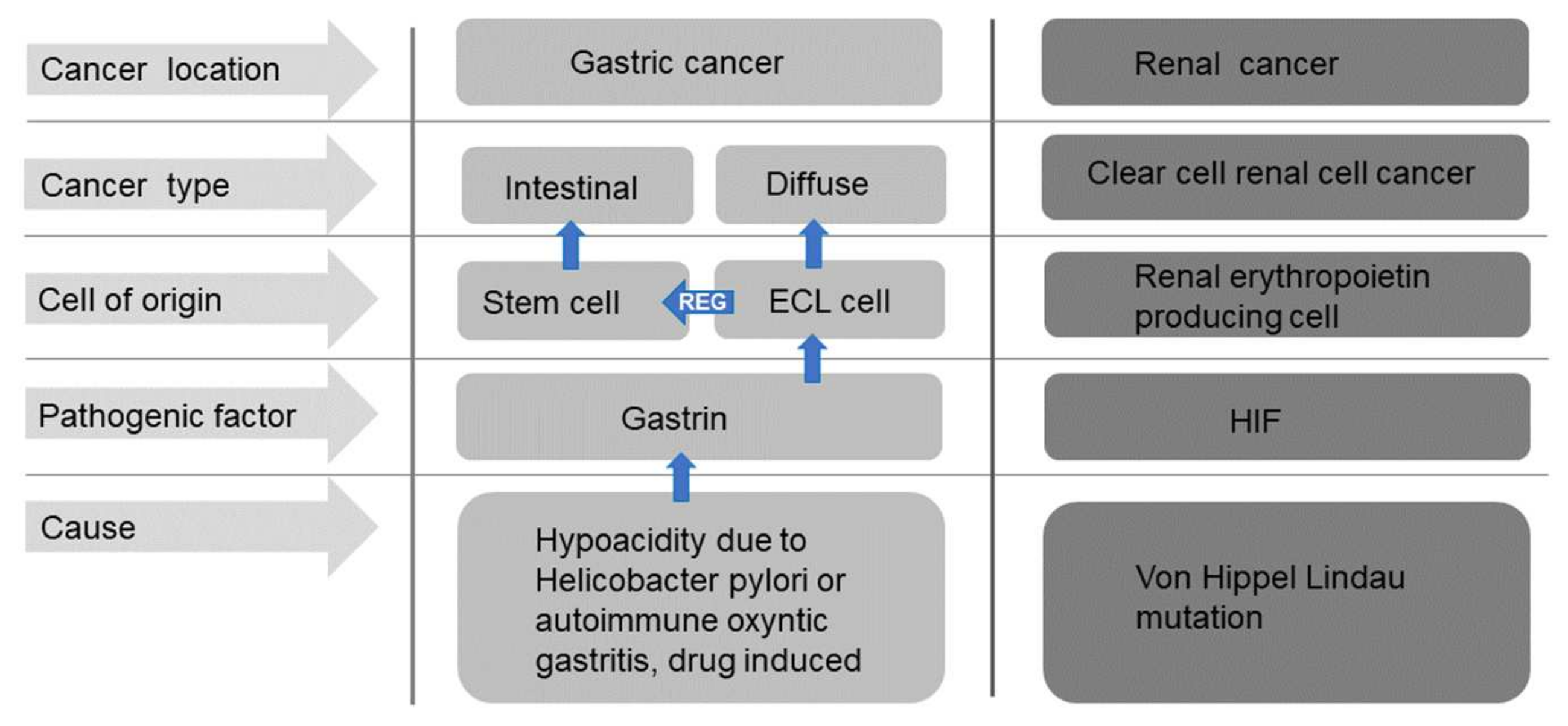
8. Gastric Cancer
9. Renal Cancer
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bullough, W.S. Mitotic and functional homeostasis: A speculative review. Cancer Res. 1965, 25, 1683–1727. [Google Scholar]
- Drozdov, I.; Kidd, M.; Gustafsson, B.I.; Svejda, B.; Joseph, R.; Pfragner, R.; Modlin, I.M. Autoregulatory effects of serotonin on proliferation and signaling pathways in lung and small intestine neuroendocrine tumor cell lines. Cancer 2009, 115, 4934–4945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sporn, M.B.; Roberts, A.B. Autocrine growth factors and cancer. Nature 1985, 313, 745–747. [Google Scholar] [CrossRef] [PubMed]
- Sandvik, A.K.; Lewin, M.J.; Waldum, H.L. Histamine release in the isolated vascularly perfused stomach of the rat: Regulation by autoreceptors. Br. J. Pharmacol. 1989, 96, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Hakanson, R.; Vallgren, S.; Ekelund, M.; Rehfeld, J.F.; Sundler, F. The vagus exerts trophic control of the stomach in the rat. Gastroenterology 1984, 86, 28–32. [Google Scholar] [CrossRef]
- Waldum, H.; Mjønes, P. Towards understanding of gastric cancer based upon physiological role of gastrin and ECL cells. Cancers 2020, 12, 3477. [Google Scholar] [CrossRef]
- Tielemans, Y.; Chen, D.; Sundler, F.; Håkanson, R.; Willems, G. Reversibility of the cell kinetic changes induced by omeprazole in the rat oxyntic mucosa: An autoradiographic study using tritiated thymidine. Scand. J. Gastroenterol. 1992, 27, 155–160. [Google Scholar] [CrossRef]
- D’Adda, T.; Annibale, B.; Delle Fave, G.; Bordi, C. Oxyntic endocrine cells of hypergastrinaemic patients. Differential response to antrectomy or octreotide. Gut 1996, 38, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Karnes, W.E., Jr.; Walsh, J.H. The gastrin hypothesis. Implications for antisecretory drug selection. J. Clin. Gastroenterol. 1990, 12 (Suppl. S2), S7–S12. [Google Scholar] [CrossRef] [PubMed]
- Peghini, P.L.; Annibale, B.; Azzoni, C.; Milione, M.; Corleto, V.D.; Gibril, F.; Venzon, D.J.; Delle Fave, G.; Bordi, C.; Jensen, R.T. Effect of chronic hypergastrinemia on human enterochromaffin-like cells: Insights from patients with sporadic gastrinomas. Gastroenterology 2002, 123, 68–85. [Google Scholar] [CrossRef]
- Lee, L.; Ramos-Alvarez, I.; Ito, T.; Jensen, R.T. Insights into effects/risks of chronic hypergastrinemia and lifelong PPI treatment in man based on studies of patients with Zollinger-Ellison syndrome. Int. J. Mol. Sci. 2019, 20, 5128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Axelson, J.; Ekelund, M.; Håkanson, R.; Sundler, F. Gastrin and the vagus interact in the trophic control of the rat oxyntic mucosa. Regul. Pept. 1988, 22, 237–243. [Google Scholar] [CrossRef]
- Waldum, H.L.; Haugen, O.A.; Brenna, E. Do neuroendocrine cells, particularly the D-cell, play a role in the development of gastric stump cancer? Cancer Detect. Prev. 1994, 18, 431–436. [Google Scholar]
- Waisberg, J.; de Matos, L.L.; do Amaral Antonio Mader, A.M.; Pezzolo, S.; Eher, E.M.; Capelozzi, V.L.; Speranzini, M.B. Neuroendocrine gastric carcinoma expressing somatostatin: A highly malignant, rare tumor. World J. Gastroenterol. 2006, 12, 3944–3947. [Google Scholar] [CrossRef]
- Jianu, C.S.; Fossmark, R.; Viset, T.; Qvigstad, G.; Sordal, O.; Marvik, R.; Waldum, H.L. Gastric carcinoids after long-term use of a proton pump inhibitor. Aliment. Pharmacol. Ther. 2012, 36, 644–649. [Google Scholar] [CrossRef]
- Procter Gamble Company/Astra Zeneca LP. Omeprazole magnesium tablets. NDA New Drug Appl. 2000, 21, 229. [Google Scholar]
- Nordqvist, S.R.; Fidler, W.J., Jr.; Woodruff, J.M.; Lewis, J.L., Jr. Clear cell adenocarcinoma of the cervix and vagina. A clinicopathologic study of 21 cases with and without a history of maternal ingestion of estrogens. Cancer 1976, 37, 858–871. [Google Scholar] [CrossRef]
- Calvete, O.; Reyes, J.; Zuniga, S.; Paumard-Hernandez, B.; Fernandez, V.; Bujanda, L.; Rodriguez-Pinilla, M.S.; Palacios, J.; Heine-Suner, D.; Banka, S.; et al. Exome sequencing identifies ATP4A gene as responsible of an atypical familial type I gastric neuroendocrine tumour. Hum. Mol. Genet. 2015, 24, 2914–2922. [Google Scholar] [CrossRef] [PubMed]
- Waldum, H.L.; Fossmark, R. Types of gastric carcinomas. Int. J. Mol. Sci. 2018, 19, 4109. [Google Scholar] [CrossRef] [Green Version]
- Mjones, P.G.; Nordrum, I.S.; Qvigstad, G.; Sordal, O.; Rian, L.L.; Waldum, H.L. Expression of erythropoietin and neuroendocrine markers in clear cell renal cell carcinoma. APMIS 2017, 125, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Choueiri, T.K.; Kaelin, W.G., Jr. Targeting the HIF2-VEGF axis in renal cell carcinoma. Nat. Med. 2020, 26, 1519–1530. [Google Scholar] [CrossRef]
- Mills, J.C.; Sansom, O.J. Reserve stem cells: Differentiated cells reprogram to fuel repair, metaplasia, and neoplasia in the adult gastrointestinal tract. Sci. Signal. 2015, 8, 385-re8. [Google Scholar] [CrossRef] [Green Version]
- Sell, S. On the stem cell origin of cancer. Am. J. Pathol. 2010, 176, 2584–2594. [Google Scholar] [CrossRef]
- Tielemans, Y.; Willems, G.; Sundler, F.; Hakanson, R. Self-replication of enterochromaffin-like cells in the mouse stomach. Digestion 1990, 45, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Barrett, P.; Hobbs, R.C.; Coates, P.J.; Risdon, R.A.; Wright, N.A.; Hall, P.A. Endocrine cells of the human gastrointestinal tract have no proliferative capacity. Histochem. J. 1995, 27, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Ooi, A.; Mai, M.; Ogino, T.; Ueda, H.; Kitamura, T.; Takahashi, Y.; Kawahara, E.; Nakanishi, I. Endocrine differentiation of gastric adenocarcinoma. The prevalence as evaluated by immunoreactive chromogranin A and its biologic significance. Cancer 1988, 62, 1096–1104. [Google Scholar] [CrossRef]
- Francis, J.M.; Kiezun, A.; Ramos, A.H.; Serra, S.; Pedamallu, C.S.; Qian, Z.R.; Banck, M.S.; Kanwar, R.; Kulkarni, A.A.; Karpathakis, A.; et al. Somatic mutation of CDKN1B in small intestine neuroendocrine tumors. Nat. Genet. 2013, 45, 1483–1486. [Google Scholar] [CrossRef] [PubMed]
- Carpizo, D.R.; Harris, C.R. Genetic drivers of ileal neuroendocrine tumors. Cancers 2021, 13, 5070. [Google Scholar] [CrossRef]
- Waldum, H.L.; Ringnes, E.; Nordbo, H.; Sordal, O.; Nordrum, I.S.; Hauso, O. The normal neuroendocrine cells of the upper gastrointestinal tract lack E-cadherin. Scand. J. Gastroenterol. 2014, 49, 974–978. [Google Scholar] [CrossRef]
- Solcia, E.; Sessa, F.; Rindi, G.; Villani, L.; Riva, C.; Buffa, R.; Capella, C. Classification and histogenesis of gastroenteropancreatic endocrine tumours. Eur. J. Clin. Investig. 1990, 20 (Suppl. S1), S72–S81. [Google Scholar] [CrossRef]
- Unger, R.H. Somatostatinoma. N. Engl. J. Med. 1977, 296, 998–1000. [Google Scholar] [CrossRef] [PubMed]
- Falkmer, U.G.; Gustafsson, T.; Wenzel, R.; Wierup, N.; Sundler, F.; Kulkarni, H.; Baum, R.P.; Falkmer, S.E. Malignant presacral ghrelinoma with long-standing hyperghrelinaemia. Ups. J. Med. Sci. 2015, 120, 299–304. [Google Scholar] [CrossRef]
- Kidd, M.; Modlin, I.M.; Gustafsson, B.I.; Drozdov, I.; Hauso, O.; Pfragner, R. Luminal regulation of normal and neoplastic human EC cell serotonin release is mediated by bile salts, amines, tastants, and olfactants. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G260–G272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, Y.; Ishihara, S.; Kadowaki, Y.; Fukui, H.; Chiba, T. Reg protein is a unique growth factor of gastric mucosal cells. J. Gastroenterol. 2004, 39, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Laskaratos, F.M.; Rombouts, K.; Caplin, M.; Toumpanakis, C.; Thirlwell, C.; Mandair, D. Neuroendocrine tumors and fibrosis: An unsolved mystery? Cancer 2017, 123, 4770–4790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bösch, F.; Bruewer, K.; D’Anastasi, M.; Ilhan, H.; Knoesel, T.; Pratschke, S.; Thomas, M.; Rentsch, M.; Guba, M.; Werner, J.; et al. Neuroendocrine tumors of the small intestine causing a desmoplastic reaction of the mesentery are a more aggressive cohort. Surgery 2018, 164, 1093–1099. [Google Scholar] [CrossRef]
- Liu, N.; Sun, S.; Wang, P.; Sun, Y.; Hu, Q.; Wang, X. The mechanism of secretion and metabolism of gut-derived 5-hydroxytryptamine. Int. J. Mol. Sci. 2021, 22, 7931. [Google Scholar] [CrossRef] [PubMed]
- Zeinali, F.; Fossmark, R.; Hauso, Ø.; Wiseth, R.; Hjertner, Ø.; Waldum, H.L. Serotonin in blood: Assessment of its origin by concomitant determination of β-thromboglobulin (platelets) and chromogranin A (enterochromaffin cells). Scand. J. Clin. Lab. Investig. 2013, 73, 148–153. [Google Scholar] [CrossRef]
- Jin, C.; Sharma, A.N.; Thevakumar, B.; Majid, M.; Al Chalaby, S.; Takahashi, N.; Tanious, A.; Arockiam, A.D.; Beri, N.; Amsterdam, E.A. Carcinoid heart disease: Pathophysiology, pathology, clinical manifestations, and management. Cardiology 2021, 146, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, B.I.; Tømmerås, K.; Nordrum, I.; Loennechen, J.P.; Brunsvik, A.; Solligård, E.; Fossmark, R.; Bakke, I.; Syversen, U.; Waldum, H. Long-term serotonin administration induces heart valve disease in rats. Circulation 2005, 111, 1517–1522. [Google Scholar] [CrossRef] [PubMed]
- Samadder, N.J.; Baffy, N.; Giridhar, K.V.; Couch, F.J.; Riegert-Johnson, D. Hereditary cancer syndromes—A primer on diagnosis and management, part 2: Gastrointestinal cancer syndromes. Mayo Clin. Proc. 2019, 94, 1099–1116. [Google Scholar] [CrossRef] [Green Version]
- Samadder, N.J.; Giridhar, K.V.; Baffy, N.; Riegert-Johnson, D.; Couch, F.J. Hereditary cancer syndromes—A primer on diagnosis and management: Part 1: Breast-ovarian cancer syndromes. Mayo Clin. Proc. 2019, 94, 1084–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldum, H.L.; Brenna, E.; Sandvik, A.K. Relationship of ECL cells and gastric neoplasia. Yale J. Biol. Med. 1998, 71, 325–335. [Google Scholar]
- O’Connor, D.T. Chromogranin: Widespread immunoreactivity in polypeptide hormone producing tissues and in serum. Regul. Pept. 1983, 6, 263–280. [Google Scholar] [CrossRef]
- Waldum, H.L.; Arnestad, J.S.; Brenna, E.; Eide, I.; Syversen, U.; Sandvik, A.K. Marked increase in gastric acid secretory capacity after omeprazole treatment. Gut 1996, 39, 649–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syversen, U.; Ramstad, H.; Gamme, K.; Qvigstad, G.; Falkmer, S.; Waldum, H.L. Clinical significance of elevated serum chromogranin A levels. Scand. J. Gastroenterol. 2004, 39, 969–973. [Google Scholar] [CrossRef]
- Bosman, E.T.; Carneiro, F.; Hruban, R.H.; Theise, N.D. WHO Classification of Tumours of the Digestive System, 4th ed.; World Health Organization: Geneva, Switzerland, 2010; 417p. [Google Scholar]
- Qvigstad, G.; Qvigstad, T.; Westre, B.; Sandvik, A.K.; Brenna, E.; Waldum, H.L. Neuroendocrine differentiation in gastric adenocarcinomas associated with severe hypergastrinemia and/or pernicious anemia. APMIS 2002, 110, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Qvigstad, G.; Falkmer, S.; Westre, B.; Waldum, H.L. Clinical and histopathological tumour progression in ECL cell carcinoids (ECLomas). APMIS 1999, 107, 1085–1092. [Google Scholar] [CrossRef]
- Waldum, H.L.; Sordal, O.F. Classification of epithelial malignant tumors—The differentiation between adenocarcinomas and neuroendocrine carcinomas: Why rely on nonspecific histochemistry and dismiss specific methods like immunohistochemistry and in situ hybridization? Appl. Immunohistochem. Mol. Morphol. AIMM 2016, 24, 309–312. [Google Scholar] [CrossRef]
- Lauren, P. The two histological main types of gastric carcinoma: Diffuse and so-called intestinal-type carcinoma. An attemt at a histo-clinical classification. APMIS 1965, 64, 31–49. [Google Scholar] [CrossRef]
- Karsten, F.H. The chemistry of Schiff’s reagent. Int. Rev. Cytol. 1960, 10, 1–100. [Google Scholar]
- Waldum, H.L.; Haugen, O.A.; Isaksen, C.; Mecsei, R.; Sandvik, A.K. Enterochromaffin-like tumour cells in the diffuse but not the intestinal type of gastric carcinomas. Eur. J. Gastroenterol. Hepatol. 1991, 3, 245–249. [Google Scholar] [CrossRef]
- Waldum, H.L.; Aase, S.; Kvetnoi, I.; Brenna, E.; Sandvik, A.K.; Syversen, U.; Johnsen, G.; Vatten, L.; Polak, J.M. Neuroendocrine differentiation in human gastric carcinoma. Cancer 1998, 83, 435–444. [Google Scholar] [CrossRef]
- Bakkelund, K.; Fossmark, R.; Nordrum, I.; Waldum, H. Signet ring cells in gastric carcinomas are derived from neuroendocrine cells. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2006, 54, 615–621. [Google Scholar] [CrossRef] [Green Version]
- Mjones, P.; Nordrum, I.S.; Sordal, O.; Sagatun, L.; Fossmark, R.; Sandvik, A.; Waldum, H.L. Expression of the cholecystokinin-B receptor in neoplastic gastric cells. Horm. Cancer 2018, 9, 40–54. [Google Scholar] [CrossRef] [Green Version]
- Sordal, O.; Qvigstad, G.; Nordrum, I.S.; Sandvik, A.K.; Gustafsson, B.I.; Waldum, H. The PAS positive material in gastric cancer cells of signet ring type is not mucin. Exp. Mol. Pathol. 2014, 96, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Martinsen, T.C.; Skogaker, N.E.; Fossmark, R.; Nordrum, I.S.; Sandvik, A.K.; Bendheim, M.O.; Bakkelund, K.E.; Waldum, H.L. Neuroendocrine cells in diffuse gastric carcinomas: An ultrastructural study with immunogold labeling of chromogranin A. Appl. Immunohistochem. Mol. Morphol. AIMM 2010, 18, 62–68. [Google Scholar] [CrossRef]
- Saper, C.B. A guide to the perplexed on the specificity of antibodies. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2009, 57, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Creutzfeldt, W.; Arnold, R.; Creutzfeldt, C.; Deuticke, U.; Frerichs, H.; Track, N.S. Biochemical and morphological investigations of 30 human insulinomas. Correlation between the tumour content of insulin and proinsulin-like components and the histological and ultrastructural appearance. Diabetologia 1973, 9, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Sørdal, Ø.; Qvigstad, G.; Nordrum, I.S.; Gustafsson, B.; Waldum, H.L. In situ hybridization in human and rodent tissue by the use of a new and simplified method. Appl. Immunohistochem. Mol. Morphol. AIMM 2013, 21, 185–189. [Google Scholar] [CrossRef]
- Roe, C.J.; Siddiqui, M.T.; Lawson, D.; Cohen, C. RNA In situ hybridization for Epstein-Barr virus and cytomegalovirus: Comparison with in- situ hybridization and immunohistochemistry. Appl. Immunohistochem. Mol. Morphol. AIMM 2019, 27, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Massoth, L.R.; Desai, N.; Szabolcs, A.; Harris, C.K.; Neyaz, A.; Crotty, R.; Chebib, I.; Rivera, M.N.; Sholl, L.M.; Stone, J.R.; et al. Comparison of RNA in situ hybridization and immunohistochemistry techniques for the detection and localization of SARS-CoV-2 in human tissues. Am. J. Surg. Pathol. 2021, 45, 14–24. [Google Scholar] [CrossRef]
- Mjønes, P.; Sagatun, L.; Nordrum, I.S.; Waldum, H.L. Neuron-specific enolase as an immunohistochemical marker is better than its reputation. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2017, 65, 687–703. [Google Scholar] [CrossRef] [PubMed]
- Sainz de Aja, J.; Dost, A.F.M.; Kim, C.F. Alveolar progenitor cells and the origin of lung cancer. J. Intern. Med. 2021, 289, 629–635. [Google Scholar] [CrossRef]
- Ferone, G.; Lee, M.C.; Sage, J.; Berns, A. Cells of origin of lung cancers: Lessons from mouse studies. Genes Dev. 2020, 34, 1017–1032. [Google Scholar] [CrossRef]
- Huang, Y.H.; Klingbeil, O.; He, X.Y.; Wu, X.S.; Arun, G.; Lu, B.; Somerville, T.D.D.; Milazzo, J.P.; Wilkinson, J.E.; Demerdash, O.E.; et al. POU2F3 is a master regulator of a tuft cell-like variant of small cell lung cancer. Genes Dev. 2018, 32, 915–928. [Google Scholar] [CrossRef] [PubMed]
- Bertuccio, P.; Chatenoud, L.; Levi, F.; Praud, D.; Ferlay, J.; Negri, E.; Malvezzi, M.; La Vecchia, C. Recent patterns in gastric cancer: A global overview. Int. J. Cancer 2009, 125, 666–673. [Google Scholar] [CrossRef]
- Ferro, A.; Peleteiro, B.; Malvezzi, M.; Bosetti, C.; Bertuccio, P.; Levi, F.; Negri, E.; La Vecchia, C.; Lunet, N. Worldwide trends in gastric cancer mortality (1980–2011), with predictions to 2015, and incidence by subtype. Eur. J. Cancer 2014, 50, 1330–1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, C.S.; Mayer, R.J. Gastric carcinoma. N. Engl. J. Med. 1995, 333, 32–41. [Google Scholar] [CrossRef]
- Siurala, M.; Seppala, K. Atrophic gastritis as a possible precursor of gastric carcinoma and pernicious anemia. Results of follow-up examinations. Acta Med. Scand. 1960, 166, 455–474. [Google Scholar] [CrossRef]
- Marshall, B.J.; Warren, J.R. Unidentified curved bacilli in the stomach of patients with gastritis and peptic ulceration. Lancet 1984, 1, 1311–1315. [Google Scholar] [CrossRef]
- Parsonnet, J.; Friedman, G.D.; Vandersteen, D.P.; Chang, Y.; Vogelman, J.H.; Orentreich, N.; Sibley, R.K. Helicobacter pylori infection and the risk of gastric carcinoma. N. Engl. J. Med. 1991, 325, 1127–1131. [Google Scholar] [CrossRef] [PubMed]
- Uemura, N.; Okamoto, S.; Yamamoto, S.; Matsumura, N.; Yamaguchi, S.; Yamakido, M.; Taniyama, K.; Sasaki, N.; Schlemper, R.J. Helicobacter pylori infection and the development of gastric cancer. N. Engl. J. Med. 2001, 345, 784–789. [Google Scholar] [CrossRef] [PubMed]
- Waldum, H.L.; Hauso, O.; Sordal, O.F.; Fossmark, R. Gastrin may mediate the carcinogenic effect of helicobacter pylori infection of the stomach. Dig. Dis. Sci. 2015, 60, 1522–1527. [Google Scholar] [CrossRef] [PubMed]
- Take, S.; Mizuno, M.; Ishiki, K.; Kusumoto, C.; Imada, T.; Hamada, F.; Yoshida, T.; Yokota, K.; Mitsuhashi, T.; Okada, H. Risk of gastric cancer in the second decade of follow-up after helicobacter pylori eradication. J. Gastroenterol. 2020, 55, 281–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buruk, F.; Berberoglu, U.; Pak, I.; Aksaz, E.; Celen, O. Gastric cancer and helicobacter pylori infection. Br. J. Surg. 1993, 80, 378–379. [Google Scholar] [CrossRef]
- Waldum, H.L.; Sordal, O.F.; Mjones, P.G. The enterochromaffin-like [ECL] cell-central in gastric physiology and pathology. Int. J. Mol. Sci. 2019, 20, 2444. [Google Scholar] [CrossRef] [Green Version]
- Fukui, H.; Kinoshita, Y.; Maekawa, T.; Okada, A.; Waki, S.; Hassan, S.; Okamoto, H.; Chiba, T. Regenerating gene protein may mediate gastric mucosal proliferation induced by hypergastrinemia in rats. Gastroenterology 1998, 115, 1483–1493. [Google Scholar] [CrossRef] [Green Version]
- Waldum, H.; Mjønes, P.G. Correct identification of cell of origin may explain many aspects of cancer: The role of neuroendocrine cells as exemplified from the stomach. Int. J. Mol. Sci. 2020, 21, 5751. [Google Scholar] [CrossRef] [PubMed]
- Boyce, M.; Moore, A.R.; Sagatun, L.; Parsons, B.N.; Varro, A.; Campbell, F.; Fossmark, R.; Waldum, H.L.; Pritchard, D.M. Netazepide, a gastrin/cholecystokinin-2 receptor antagonist, can eradicate gastric neuroendocrine tumours in patients with autoimmune chronic atrophic gastritis. Br. J. Clin. Pharmacol. 2017, 83, 466–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fossmark, R.; Sordal, O.; Jianu, C.S.; Qvigstad, G.; Nordrum, I.S.; Boyce, M.; Waldum, H.L. Treatment of gastric carcinoids type 1 with the gastrin receptor antagonist netazepide (YF476) results in regression of tumours and normalisation of serum chromogranin A. Aliment. Pharmacol. Ther. 2012, 36, 1067–1075. [Google Scholar] [CrossRef]
- Mancini, V.; Battaglia, M.; Ditonno, P.; Palazzo, S.; Lastilla, G.; Montironi, R.; Bettocchi, C.; Cavalcanti, E.; Ranieri, E.; Selvaggi, F.P. Current insights in renal cell cancer pathology. Urol. Oncol. 2008, 26, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Schoener, B.; Borger, J. Erythropoietin. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Nolan, K.A.; Wenger, R.H. Source and microenvironmental regulation of erythropoietin in the kidney. Curr. Opin. Nephrol. Hypertens. 2018, 27, 277–282. [Google Scholar] [CrossRef]
- Gerl, K.; Nolan, K.A.; Karger, C.; Fuchs, M.; Wenger, R.H.; Stolt, C.C.; Willam, C.; Kurtz, A.; Kurt, B. Erythropoietin production by PDGFR-β(+) cells. Pflügers Arch. 2016, 468, 1479–1487. [Google Scholar] [CrossRef]
- Buemi, M.; Cavallaro, E.; Floccari, F.; Sturiale, A.; Aloisi, C.; Trimarchi, M.; Corica, F.; Frisina, N. The pleiotropic effects of erythropoietin in the central nervous system. J. Neuropathol. Exp. Neurol. 2003, 62, 228–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Wang, Y.; Tong, D.; Liu, G.; Yuan, W.; Zhang, J.; Ye, J.; Zhang, Y.; Yuan, G.; Feng, Q.; et al. A somatic HIF2α mutation-induced multiple and recurrent pheochromocytoma/paraganglioma with polycythemia: Clinical study with literature review. Endocr. Pathol. 2017, 28, 75–82. [Google Scholar] [CrossRef]
- Chen, W.; Hill, H.; Christie, A.; Kim, M.S.; Holloman, E.; Pavia-Jimenez, A.; Homayoun, F.; Ma, Y.; Patel, N.; Yell, P.; et al. Targeting renal cell carcinoma with a HIF-2 antagonist. Nature 2016, 539, 112–117. [Google Scholar] [CrossRef] [Green Version]
- Voit, R.A.; Sankaran, V.G. Stabilizing HIF to ameliorate anemia. Cell 2020, 180, 6. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peculiarities with Neuroendocrine Tumours | These Peculiarities May Be Explained by Properties of the Cell of Origin |
|---|---|
| Despite an apparent benign phenotype, metastasize early | They are spread among other cells reflecting low adherence. The enterochromaffin like (ECL) cell in the stomach has been shown not to express E-cadherin. Low adherence facilitates spread. |
| Grow slowly. Cytotoxic drugs mainly without any effect on tumour growth | NE cells do divide, but very slowly |
| Tumour cells look very similar to the normal cell of origin. Low mutation rate. Immunotherapy does not seem promising | |
| Produce signal substances which may give symptoms if reaching the circulation in sufficient concentration and having an easily recognized effect | Produce signal substances |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Waldum, H.; Mjønes, P. Time to Classify Tumours of the Stomach and the Kidneys According to Cell of Origin. Int. J. Mol. Sci. 2021, 22, 13386. https://doi.org/10.3390/ijms222413386
Waldum H, Mjønes P. Time to Classify Tumours of the Stomach and the Kidneys According to Cell of Origin. International Journal of Molecular Sciences. 2021; 22(24):13386. https://doi.org/10.3390/ijms222413386
Chicago/Turabian StyleWaldum, Helge, and Patricia Mjønes. 2021. "Time to Classify Tumours of the Stomach and the Kidneys According to Cell of Origin" International Journal of Molecular Sciences 22, no. 24: 13386. https://doi.org/10.3390/ijms222413386
APA StyleWaldum, H., & Mjønes, P. (2021). Time to Classify Tumours of the Stomach and the Kidneys According to Cell of Origin. International Journal of Molecular Sciences, 22(24), 13386. https://doi.org/10.3390/ijms222413386