
Role of the Cholinergic Anti-Inflammatory Reflex in Central Nervous System Diseases

 , ,
, ,
Abstract
:1. Introduction
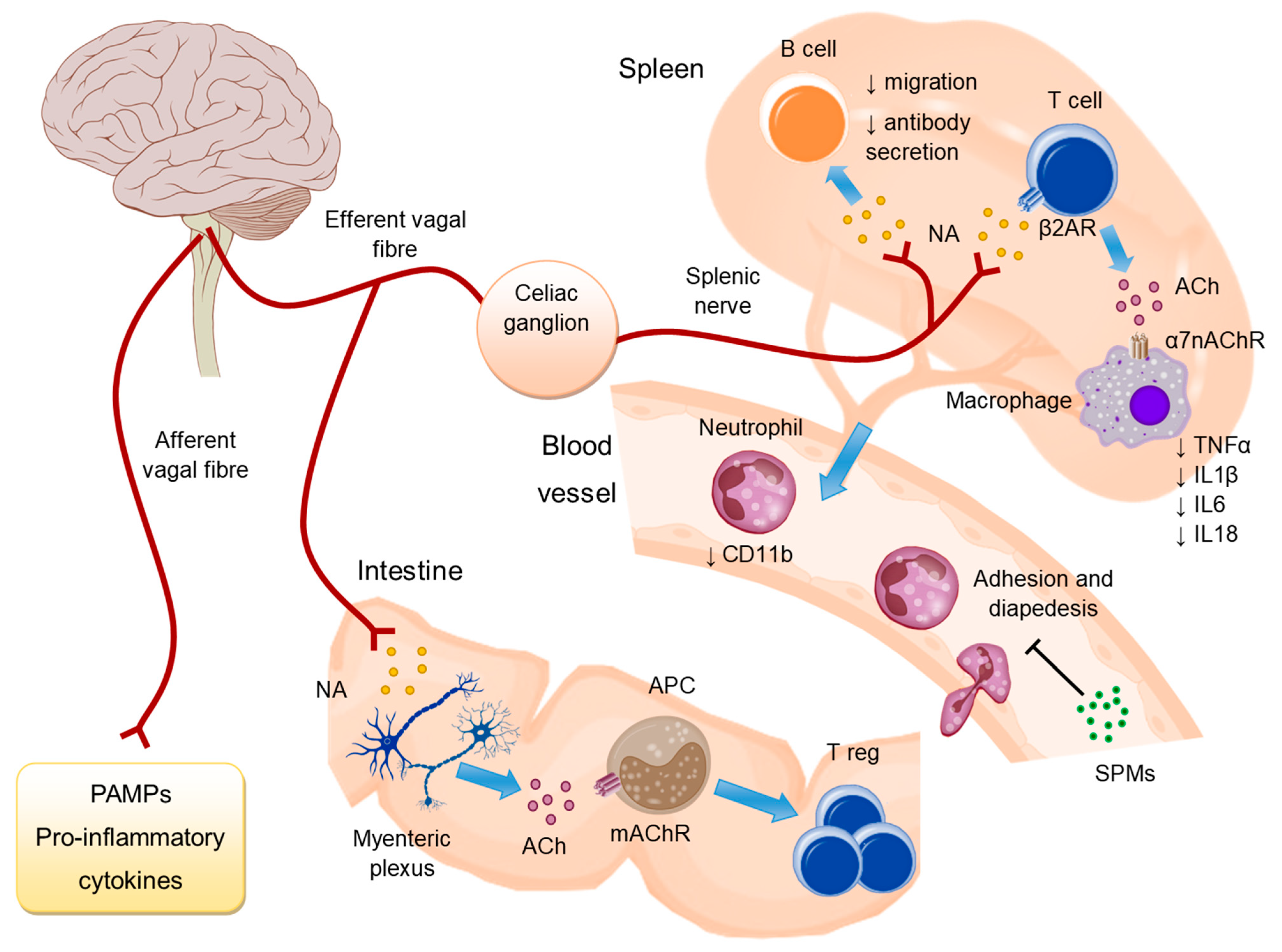
2. Inflammatory Reflex: Cholinergic Anti-Inflammatory Pathway
3. Clinical Applications of Inflammatory Reflex Modulation
4. Role of the Anti-Inflammatory Reflex and Main Advances in Chemical Modulation with α7 nAChR Agonists and VNS in Central Nervous System Diseases
4.1. Experimental Autoimmune Encephalomyelitis and Multiple Sclerosis
4.2. Alzheimer’s Disease
4.3. Parkinson’s Disease
4.4. Epilepsy
4.5. Depression
4.6. Migraine
4.7. Schizophrenia
5. Blockade of the Anti-Inflammatory Cholinergic Pathway Mediated by MicroRNAs
6. Discussion
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gidron, Y. Neuroimmunomodulation BT—Encyclopedia of Behavioral Medicine; Gellman, M.D., Turner, J.R., Eds.; Springer: New York, NY, USA, 2013; pp. 1321–1322. ISBN 978-1-4419-1005-9. [Google Scholar]
- Kamimura, D.; Tanaka, Y.; Hasebe, R.; Murakami, M. Bidirectional communication between neural and immune systems. Int. Immunol. 2020, 32, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Scanzano, A.; Cosentino, M. Adrenergic regulation of innate immunity: A review. Front. Pharmacol. 2015, 6, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoli, M.; Pucci, S.; Vilella, A.; Gotti, C. Neuronal and Extraneuronal Nicotinic Acetylcholine Receptors. Curr. Neuropharmacol. 2018, 16, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Corsi-Zuelli, F.M.D.G.; Brognara, F.; Quirino, G.F.D.S.; Hiroki, C.H.; Fais, R.S.; Del-Ben, C.M.; Ulloa, L.; Salgado, H.C.; Kanashiro, A.; Loureiro, C.M. Neuroimmune Interactions in Schizophrenia: Focus on Vagus Nerve Stimulation and Activation of the Alpha-7 Nicotinic Acetylcholine Receptor. Front. Immunol. 2017, 8, 618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlov, V.A.; Chavan, S.S.; Tracey, K.J. Molecular and Functional Neuroscience in Immunity. Annu. Rev. Immunol. 2018, 36, 783–812. [Google Scholar] [CrossRef]
- Lee, H.; Lee, S.; Cho, I.-H.; Lee, S.J. Toll-Like Receptors: Sensor Molecules for Detecting Damage to the Nervous System. Curr. Protein Pept. Sci. 2013, 14, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-J.; Liu, T.; Chen, G.; Wang, B.; Yu, X.-L.; Yin, C.; Ji, R.-R. TLR signaling adaptor protein MyD88 in primary sensory neurons contributes to persistent inflammatory and neuropathic pain and neuroinflammation. Sci. Rep. 2016, 6, 28188. [Google Scholar] [CrossRef] [Green Version]
- Kaushal, V.; Dye, R.; Pakavathkumar, P.; Foveau, B.; Flores, J.C.; Hyman, B.T.; Ghetti, B.; Koller, B.H.; Leblanc, A.C. Neuronal NLRP1 inflammasome activation of Caspase-1 coordinately regulates inflammatory interleukin-1-beta production and axonal degeneration-associated Caspase-6 activation. Cell Death Differ. 2015, 22, 1676–1686. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, W.; Zhang, L.; Dai, P.; Wang, K.; Hui, H.; Rao, W.; Peng, C.; Yang, J.; Yan, Z.; et al. Oxygen-glucose deprivation inducing B1 RNA inhibits neuronal cells metabolic activity by NLRP3 and associated proinflammatory cytokines production. Neurosci. Lett. 2015, 588, 147–153. [Google Scholar] [CrossRef]
- Wilhelm, I.; Nyúl-Tóth, Á.; Kozma, M.; Farkas, A.E.; Krizbai, I.A. Role of pattern recognition receptors of the neurovascular unit in inflamm-aging. Am. J. Physiol. Circ. Physiol. 2017, 313, H1000–H1012. [Google Scholar] [CrossRef] [Green Version]
- Trettel, F.; Di Castro, M.A.; Limatola, C. Chemokines: Key Molecules that Orchestrate Communication among Neurons, Microglia and Astrocytes to Preserve Brain Function. Neuroscience 2020, 439, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.E.S.; Goodkey, K.; Footz, T.; Voronova, A. Regulation of CNS precursor function by neuronal chemokines. Neurosci. Lett. 2020, 715, 134533. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, B.E.; Silverman, H.A.; Robbiati, S.; Gunasekaran, M.K.; Tsaava, T.; Battinelli, E.; Stiegler, A.; Bouton, C.E.; Chavan, S.S.; Tracey, K.J.; et al. Cytokine-specific Neurograms in the Sensory Vagus Nerve. Bioelectron. Med. 2016, 3, 7–17. [Google Scholar] [CrossRef]
- Bernik, T.R.; Friedman, S.G.; Ochani, M.; DiRaimo, R.; Ulloa, L.; Yang, H.; Sudan, S.; Czura, C.J.; Ivanova, S.M.; Tracey, K.J. Pharmacological Stimulation of the Cholinergic Antiinflammatory Pathway. J. Exp. Med. 2002, 195, 781–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borovikova, L.V.; Ivanova, S.; Zhang, M.; Yang, H.; Botchkina, G.I.; Watkins, L.R.; Wang, H.; Abumrad, N.; Eaton, J.W.; Tracey, K.J. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature 2000, 405, 458–462. [Google Scholar] [CrossRef]
- Tracey, K.J. The inflammatory reflex. Nature 2002, 420, 853–859. [Google Scholar] [CrossRef]
- Vida, G.; Peña, G.; Deitch, E.A.; Ulloa, L. α7-Cholinergic Receptor Mediates Vagal Induction of Splenic Norepinephrine. J. Immunol. 2011, 186, 4340–4346. [Google Scholar] [CrossRef] [Green Version]
- Rosas-Ballina, M.; Olofsson, P.S.; Ochani, M.; Valdés-Ferrer, S.I.; Levine, Y.A.; Reardon, C.; Tusche, M.W.; Pavlov, V.A.; Andersson, U.; Chavan, S.; et al. Acetylcholine-Synthesizing T Cells Relay Neural Signals in a Vagus Nerve Circuit. Science 2011, 334, 98–101. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Yu, M.; Ochani, M.; Amella, C.A.; Tanovic, M.; Susarla, S.; Li, J.H.; Wang, H.; Yang, H.; Ulloa, L.; et al. Nicotinic acetylcholine receptor α7 subunit is an essential regulator of inflammation. Nature 2003, 421, 384–388. [Google Scholar] [CrossRef]
- Pavlov, V.A.; Ochani, M.; Gallowitsch-Puerta, M.; Ochani, K.; Huston, J.M.; Czura, C.J.; Al-Abed, Y.; Tracey, K.J. Central muscarinic cholinergic regulation of the systemic inflammatory response during endotoxemia. Proc. Natl. Acad. Sci. USA 2006, 103, 5219–5223. [Google Scholar] [CrossRef] [Green Version]
- Guarini, S.; Altavilla, D.; Cainazzo, M.-M.; Giuliani, D.; Bigiani, A.; Marini, H.; Squadrito, G.; Minutoli, L.; Bertolini, A.; Marini, R.; et al. Efferent Vagal Fibre Stimulation Blunts Nuclear Factor-κB Activation and Protects Against Hypovolemic Hemorrhagic Shock. Circulation 2003, 107, 1189–1194. [Google Scholar] [CrossRef] [Green Version]
- De Jonge, W.J.; Van Der Zanden, E.P.; The, F.O.; Bijlsma, M.F.; Van Westerloo, D.J.; Bennink, R.J.; Berthoud, H.-R.; Uematsu, S.; Akira, S.; van den Wijngaard, R.M.; et al. Stimulation of the vagus nerve attenuates macrophage activation by activating the Jak2-STAT3 signaling pathway. Nat. Immunol. 2005, 6, 844–851. [Google Scholar] [CrossRef]
- Lu, B.; Kwan, K.; Levine, Y.A.; Olofsson, P.S.; Yang, H.; Li, J.; Joshi, S.; Wang, H.; Andersson, U.; Chavan, S.S.; et al. α7 Nicotinic Acetylcholine Receptor Signaling Inhibits Inflammasome Activation by Preventing Mitochondrial DNA Release. Mol. Med. 2014, 20, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Mina-Osorio, P.; Rosas-Ballina, M.; Valdes-Ferrer, S.I.; Al-Abed, Y.; Tracey, K.J.; Diamond, B. Neural Signaling in the Spleen Controls B-Cell Responses to Blood-Borne Antigen. Mol. Med. 2012, 18, 618–627. [Google Scholar] [CrossRef]
- Carnevale, D.; Perrotta, M.; Pallante, F.; Fardella, V.; Iacobucci, R.; Fardella, S.; Carnevale, L.; Carnevale, R.; De Lucia, M.; Cifelli, G.; et al. A cholinergic-sympathetic pathway primes immunity in hypertension and mediates brain-to-spleen communication. Nat. Commun. 2016, 7, 13035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, R.W.; Varma, S.; Peng-Nemeroff, T.; Sherry, B.; Balakhaneh, D.; Huston, J.; Tracey, K.J.; Al-Abed, Y.; Metz, C.N. Cholinergic stimulation blocks endothelial cell activation and leukocyte recruitment during inflammation. J. Exp. Med. 2005, 201, 1113–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huston, J.M.; Rosas-Ballina, M.; Xue, X.; Dowling, O.; Ochani, K.; Ochani, M.; Yeboah, M.M.; Chatterjee, P.K.; Tracey, K.J.; Metz, C.N. Cholinergic Neural Signals to the Spleen Down-Regulate Leukocyte Trafficking via CD11b. J. Immunol. 2009, 183, 552–559. [Google Scholar] [CrossRef] [Green Version]
- Dalli, J.; Colas, R.A.; Arnardottir, H.; Serhan, C.N. Vagal Regulation of Group 3 Innate Lymphoid Cells and the Immunoresolvent PCTR1 Controls Infection Resolution. Immunity 2017, 46, 92–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirakaj, V.; Dalli, J.; Granja, T.; Rosenberger, P.; Serhan, C.N. Vagus nerve controls resolution and pro-resolving mediators of inflammation. J. Exp. Med. 2014, 211, 1037–1048. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N.; Chiang, N.; Dalli, J. The resolution code of acute inflammation: Novel pro-resolving lipid mediators in resolution. Semin. Immunol. 2015, 27, 200–215. [Google Scholar] [CrossRef] [Green Version]
- Teratani, T.; Mikami, Y.; Nakamoto, N.; Suzuki, T.; Harada, Y.; Okabayashi, K.; Hagihara, Y.; Taniki, N.; Kohno, K.; Shibata, S.; et al. The liver–brain–gut neural arc maintains the Treg cell niche in the gut. Nature 2020, 585, 591–596. [Google Scholar] [CrossRef]
- Lehner, K.R.; Silverman, H.A.; Addorisio, M.E.; Roy, A.; Al-Onaizi, M.A.; Levine, Y.; Olofsson, P.S.; Chavan, S.S.; Gros, R.; Nathanson, N.M.; et al. Forebrain Cholinergic Signaling Regulates Innate Immune Responses and Inflammation. Front. Immunol. 2019, 10, 585. [Google Scholar] [CrossRef] [Green Version]
- Uchida, M.; Yamamoto, R.; Matsuyama, S.; Murakami, K.; Hasebe, R.; Hojyo, S.; Tanaka, Y.; Murakami, M. Gateway reflexes, neuronal circuits that regulate the gateways for autoreactive T cells in organs that have blood barriers. Int. Immunol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Sinniger, V.; Pellissier, S.; Fauvelle, F.; Trocmé, C.; Hoffmann, D.; Vercueil, L.; Cracowski, J.L.; David, O.; Bonaz, B. A 12-month pilot study outcomes of vagus nerve stimulation in Crohn’s disease. Neurogastroenterol. Motil. 2020, 32, e13911. [Google Scholar] [CrossRef]
- Koopman, F.A.; Chavan, S.S.; Miljko, S.; Grazio, S.; Sokolovic, S.; Schuurman, P.R.; Mehta, A.D.; Levine, Y.A.; Faltys, M.; Zitnik, R.; et al. Vagus nerve stimulation inhibits cytokine production and attenuates disease severity in rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 2016, 113, 8284–8289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastitskaya, S.; Thompson, N.; Holder, D. Selective Vagus Nerve Stimulation as a Therapeutic Approach for the Treatment of ARDS: A Rationale for Neuro-Immunomodulation in COVID-19 Disease. Front. Neurosci. 2021, 15, 667036. [Google Scholar] [CrossRef] [PubMed]
- Aranow, C.; Atish-Fregoso, Y.; Lesser, M.; Mackay, M.; Anderson, E.; Chavan, S.; Zanos, T.P.; Datta-Chaudhuri, T.; Bouton, C.; Tracey, K.J.; et al. Transcutaneous auricular vagus nerve stimulation reduces pain and fatigue in patients with systemic lupus erythematosus: A randomised, double-blind, sham-controlled pilot trial. Ann. Rheum. Dis. 2021, 80, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Tsaava, T.; Datta-Chaudhuri, T.; Addorisio, M.E.; Masi, E.B.; Silverman, H.A.; Newman, J.E.; Imperato, G.H.; Bouton, C.; Tracey, K.J.; Chavan, S.S.; et al. Specific vagus nerve stimulation parameters alter serum cytokine levels in the absence of inflammation. Bioelectron. Med. 2020, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caravaca, A.S.; Centa, M.; Gallina, A.L.; Tarnawski, L.; Olofsson, P.S. Neural reflex control of vascular inflammation. Bioelectron. Med. 2020, 6, 1–8. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, M.; Yu, X.; Zang, W. Pharmacological Modulation of Vagal Nerve Activity in Cardiovascular Diseases. Neurosci. Bull. 2019, 35, 156–166. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Wang, S.; Li, X.; Yu, L.; Yang, H.; Liu, Q.; Tang, J.; Zhou, S. Non-invasive Autonomic Neuromodulation Is Opening New Landscapes for Cardiovascular Diseases. Front. Physiol. 2020, 11, 550578. [Google Scholar] [CrossRef]
- Ingegnoli, F.; Buoli, M.; Antonucci, F.; Coletto, L.A.; Esposito, C.M.; Caporali, R. The Link Between Autonomic Nervous System and Rheumatoid Arthritis: From Bench to Bedside. Front. Med. 2020, 7, 589079. [Google Scholar] [CrossRef] [PubMed]
- Machado-Alba, J.E.; Calvo-Torres, L.F.; Gaviria-Mendoza, A.; Mejía-Vélez, C.A. Prescription profile of pyridostigmine use in a population of patients with myasthenia gravis. Muscle Nerve 2017, 56, 1041–1046. [Google Scholar] [CrossRef] [PubMed]
- Choque, P.N.B.; Vieira, R.P.; Ulloa, L.; Grabulosa, C.; Irigoyen, M.C.; De Angelis, K.; De Oliveira, A.P.L.; Tracey, K.J.; Pavlov, V.A.; Consolim-Colombo, F.M. The Cholinergic Drug Pyridostigmine Alleviates Inflammation During LPS-Induced Acute Respiratory Distress Syndrome. Front. Pharmacol. 2021, 12, 624895. [Google Scholar] [CrossRef]
- Fragoso-Saavedra, S.; Iruegas-Nunez, D.A.; Quintero-Villegas, A.; García-González, H.B.; Nuñez, I.; Carbajal-Morelos, S.L.; Audelo-Cruz, B.M.; Arias-Martínez, S.; Caro-Vega, Y.; Calva, J.J.; et al. A parallel-group, multicenter randomized, double-blinded, placebo-controlled, phase 2/3, clinical trial to test the efficacy of pyridostigmine bromide at low doses to reduce mortality or invasive mechanical ventilation in adults with severe SARS-CoV-2 infection: The Pyridostigmine In Severe COvid-19 (PISCO) trial protocol. BMC Infect. Dis. 2020, 20, 1–7. [Google Scholar] [CrossRef]
- Aubé, B.; Lévesque, S.A.; Paré, A.; Chamma, É.; Kébir, H.; Gorina, R.; Lécuyer, M.-A.; Alvarez, J.I.; De Koninck, Y.; Engelhardt, B.; et al. Neutrophils Mediate Blood–Spinal Cord Barrier Disruption in Demyelinating Neuroinflammatory Diseases. J. Immunol. 2014, 193, 2438–2454. [Google Scholar] [CrossRef] [Green Version]
- Wesselingh, R.; Butzkueven, H.; Buzzard, K.; Tarlinton, D.; O’Brien, T.J.; Monif, M. Innate Immunity in the Central Nervous System: A Missing Piece of the Autoimmune Encephalitis Puzzle? Front. Immunol. 2019, 10, 2066. [Google Scholar] [CrossRef]
- Ponomarev, E.D.; Shriver, L.P.; Maresz, K.; Dittel, B.N. Microglial cell activation and proliferation precedes the onset of CNS autoimmunity. J. Neurosci. Res. 2005, 81, 374–389. [Google Scholar] [CrossRef] [PubMed]
- Ifergan, I.; Kebir, H.; Bernard, M.; Wosik, K.; Dodelet-Devillers, A.; Cayrol, R.; Arbour, N.; Prat, A. The blood-brain barrier induces differentiation of migrating monocytes into Th17-polarizing dendritic cells. Brain 2008, 131, 785–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brambilla, R. The contribution of astrocytes to the neuroinflammatory response in multiple sclerosis and experimental autoimmune encephalomyelitis. Acta Neuropathol. 2019, 137, 757–783. [Google Scholar] [CrossRef]
- Goverman, J. Autoimmune T cell responses in the central nervous system. Nat. Rev. Immunol. 2009, 9, 393–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nizri, E.; Hamra-Amitay, Y.; Sicsic, C.; Lavon, I.; Brenner, T. Anti-inflammatory properties of cholinergic up-regulation: A new role for acetylcholinesterase inhibitors. Neuropharmacology 2006, 50, 540–547. [Google Scholar] [CrossRef]
- Di Pinto, G.; Di Bari, M.; Martin-Alvarez, R.; Sperduti, S.; Serrano-Acedo, S.; Gatta, V.; Tata, A.M.; Mengod, G. Comparative study of the expression of cholinergic system components in the CNS of experimental autoimmune encephalomyelitis mice: Acutevsremitting phase. Eur. J. Neurosci. 2018, 48, 2165–2181. [Google Scholar] [CrossRef] [PubMed]
- Reale, M.; Costantini, E.; Di Nicola, M.; D’Angelo, C.; Franchi, S.; D’Aurora, M.; Di Bari, M.; Orlando, V.; Galizia, S.; Ruggieri, S.; et al. Butyrylcholinesterase and Acetylcholinesterase polymorphisms in Multiple Sclerosis patients: Implication in peripheral inflammation. Sci. Rep. 2018, 8, 1319. [Google Scholar] [CrossRef]
- Di Bari, M.; Reale, M.; Di Nicola, M.; Orlando, V.; Galizia, S.; Porfilio, I.; Costantini, E.; D’Angelo, C.; Ruggieri, S.; Biagioni, S.; et al. Dysregulated Homeostasis of Acetylcholine Levels in Immune Cells of RR-Multiple Sclerosis Patients. Int. J. Mol. Sci. 2016, 17, 2009. [Google Scholar] [CrossRef] [Green Version]
- Gatta, V.; Mengod, G.; Reale, M.; Tata, A.M. Possible Correlation between Cholinergic System Alterations and Neuro/Inflammation in Multiple Sclerosis. Biomedicines 2020, 8, 153. [Google Scholar] [CrossRef] [PubMed]
- Habek, M. Immune and autonomic nervous system interactions in multiple sclerosis: Clinical implications. Clin. Auton. Res. 2019, 29, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Mizrachi, T.; Marsha, O.; Brusin, K.; Ben-David, Y.; Thakur, G.A.; Vaknin-Dembinsky, A.; Treinin, M.; Brenner, T. Suppression of neuroinflammation by an allosteric agonist and positive allosteric modulator of the α7 nicotinic acetylcholine receptor GAT107. J. Neuroinflamm. 2021, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Li, D.; Han, R.; Zhang, C.; Jin, W.-N.; Wood, K.; Liu, Q.; Shi, F.-D.; Hao, J. Acetylcholine-producing NK cells attenuate CNS inflammation via modulation of infiltrating monocytes/macrophages. Proc. Natl. Acad. Sci. USA 2017, 114, E6202–E6211. [Google Scholar] [CrossRef] [Green Version]
- Rothbard, J.B.; Kurnellas, M.P.; Ousman, S.S.; Brownell, S.; Rothbard, J.J.; Steinman, L. Small Heat Shock Proteins, Amyloid Fibrils, and Nicotine Stimulate a Common Immune Suppressive Pathway with Implications for Future Therapies. Cold Spring Harb. Perspect. Med. 2019, 9, a034223. [Google Scholar] [CrossRef] [PubMed]
- Rothbard, J.B.; Rothbard, J.J.; Soares, L.; Fathman, C.G.; Steinman, L. Identification of a common immune regulatory pathway induced by small heat shock proteins, amyloid fibrils, and nicotine. Proc. Natl. Acad. Sci. USA 2018, 115, 7081–7086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurnellas, M.P.; Ghosn, E.E.B.; Schartner, J.M.; Baker, J.; Rothbard, J.J.; Negrin, R.S.; Herzenberg, L.A.; Fathman, C.G.; Steinman, L.; Rothbard, J.B. Amyloid fibrils activate B-1a lymphocytes to ameliorate inflammatory brain disease. Proc. Natl. Acad. Sci. USA 2015, 112, 15016–15023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; St-Pierre, S.; Roy, P.; Morley, B.J.; Hao, J.; Simard, A.R. Infiltration of CCR2+Ly6Chigh Proinflammatory Monocytes and Neutrophils into the Central Nervous System Is Modulated by Nicotinic Acetylcholine Receptors in a Model of Multiple Sclerosis. J. Immunol. 2016, 196, 2095–2108. [Google Scholar] [CrossRef] [Green Version]
- Ke, P.; Shao, B.-Z.; Xu, Z.-Q.; Chen, X.-W.; Wei, W.; Liu, C. Activating α7 nicotinic acetylcholine receptor inhibits NLRP3 inflammasome through regulation of β-arrestin-1. CNS Neurosci. Ther. 2017, 23, 875–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reale, M.; Di Bari, M.; DI Nicola, M.; D’Angelo, C.; De Angelis, F.; Velluto, L.; Tata, A.M. Nicotinic receptor activation negatively modulates pro-inflammatory cytokine production in multiple sclerosis patients. Int. Immunopharmacol. 2015, 29, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Godin, J.-R.; Roy, P.; Quadri, M.; Bagdas, D.; Toma, W.; Narendrula-Kotha, R.; Kishta, O.A.; Damaj, M.I.; Horenstein, N.A.; Papke, R.L.; et al. A silent agonist of α7 nicotinic acetylcholine receptors modulates inflammation ex vivo and attenuates EAE. Brain Behav. Immun. 2020, 87, 286–300. [Google Scholar] [CrossRef]
- Nicolussi, E.-M.; Huck, S.; Lassmann, H.; Bradl, M. The cholinergic anti-inflammatory system limits T cell infiltration into the neurodegenerative CNS, but cannot counteract complex CNS inflammation. Neurobiol. Dis. 2009, 35, 24–31. [Google Scholar] [CrossRef]
- Zhou, L.; Lin, X.; Ma, X.; Liu, Y.; Ma, L.; Chen, Z.; Chen, H.; Si, L.; Chen, X. Acetylcholine regulates the development of experimental autoimmune encephalomyelitis via the CD4+ cells proliferation and differentiation. Int. J. Neurosci. 2020, 130, 788–803. [Google Scholar] [CrossRef]
- Alzheimer’s Association. Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2021, 17, 327–406. [Google Scholar] [CrossRef]
- Kokras, N.; Stamouli, E.; Sotiropoulos, I.; Katirtzoglou, E.A.; Siarkos, K.T.; Dalagiorgou, G.; Alexandraki, K.I.; Coulocheri, S.; Piperi, C.; Politis, A.M. Acetyl Cholinesterase Inhibitors and Cell-Derived Peripheral Inflammatory Cytokines in Early Stages of Alzheimer’s Disease. J. Clin. Psychopharmacol. 2018, 38, 138–143. [Google Scholar] [CrossRef]
- Sung, P.-S.; Lin, P.-Y.; Liu, C.-H.; Su, H.-C.; Tsai, K.-J. Neuroinflammation and Neurogenesis in Alzheimer’s Disease and Potential Therapeutic Approaches. Int. J. Mol. Sci. 2020, 21, 701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ADAPT-FS Research Group. Follow-up evaluation of cognitive function in the randomized Alzheimer’s Disease Anti-inflammatory Prevention Trial and its Follow-up Study. Alzheimer’s Dement. 2015, 11, 216–225.e1. [Google Scholar] [CrossRef] [Green Version]
- Butchart, J.; Brook, L.; Hopkins, V.; Teeling, J.; Püntener, U.; Culliford, D.; Sharples, R.; Sharif, S.; McFarlane, B.; Raybould, R.; et al. Etanercept in Alzheimer disease: A randomized, placebo-controlled, double-blind, phase 2 trial. Neurology 2015, 84, 2161–2168. [Google Scholar] [CrossRef] [Green Version]
- Meyer, P.-F.; Tremblay-Mercier, J.; Leoutsakos, J.; Madjar, C.; Lafaille-Maignan, M.-É.; Savard, M.; Rosa-Neto, P.; Poirier, J.; Etienne, P.; Breitner, J.; et al. Intrepad: A randomized trial of naproxen to slow progress of presymptomatic Alzheimer disease. Neurology 2019, 92, e2070–e2080. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Sarasúa, S.; Fernández-Pérez, I.; Espinosa-Fernández, V.; Sánchez-Pérez, A.M.; Ledesma, J.C. Can We Treat Neuroinflammation in Alzheimer’s Disease? Int. J. Mol. Sci. 2020, 21, 8751. [Google Scholar] [CrossRef] [PubMed]
- Winek, K.; Soreq, H.; Meisel, A. Regulators of cholinergic signaling in disorders of the central nervous system. J. Neurochem. 2021, 158, 1425–1438. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.-G.; Qian, Y.-H. Alpha 7 nicotinic acetylcholine receptor and its effects on Alzheimer’s disease. Neuropeptides 2019, 73, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Athar, T.; Al Balushi, K.; Khan, S.A. Recent advances on drug development and emerging therapeutic agents for Alzheimer’s disease. Mol. Biol. Rep. 2021, 1–17. [Google Scholar] [CrossRef]
- Reale, M.; Iarlori, C.; Gambi, F.; Feliciani, C.; Isabella, L.; Gambi, D. The acetylcholinesterase inhibitor, Donepezil, regulates a Th2 bias in Alzheimer’s disease patients. Neuropharmacology 2006, 50, 606–613. [Google Scholar] [CrossRef]
- Richardson, C.; Gard, P.R.; Klugman, A.; Isaac, M.; Tabet, N. Blood pro-inflammatory cytokines in Alzheimer’s disease in relation to the use of acetylcholinesterase inhibitors. Int. J. Geriatr. Psychiatry 2013, 28, 1312–1317. [Google Scholar] [CrossRef] [PubMed]
- Alhowail, A. Molecular insights into the benefits of nicotine on memory and cognition (Review). Mol. Med. Rep. 2021, 23, 1–6. [Google Scholar] [CrossRef]
- Srivareerat, M.; Tran, T.T.; Salim, S.; Aleisa, A.M.; Alkadhi, K.A. Chronic nicotine restores normal Aβ levels and prevents short-term memory and E-LTP impairment in Aβ rat model of Alzheimer’s disease. Neurobiol. Aging 2011, 32, 834–844. [Google Scholar] [CrossRef]
- Majdi, A.; Kamari, F.; Sadigh-Eteghad, S.; Gjedde, A. Molecular Insights Into Memory-Enhancing Metabolites of Nicotine in Brain: A Systematic Review. Front. Neurosci. 2019, 12, 1002. [Google Scholar] [CrossRef] [PubMed]
- Deardorff, W.J.; Shobassy, A.; Grossberg, G.T. Safety and clinical effects of EVP-6124 in subjects with Alzheimer’s disease currently or previously receiving an acetylcholinesterase inhibitor medication. Expert Rev. Neurother. 2015, 15, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.B.; Smith, D.C.; Hassert, D.L.; Browning, R.A.; Naritoku, D.K.; Jensen, R.A. Posttraining Electrical Stimulation of Vagal Afferents with Concomitant Vagal Efferent Inactivation Enhances Memory Storage Processes in the Rat. Neurobiol. Learn. Mem. 1998, 70, 364–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.; Fuchino, Y.; Miyamoto, D.; Nomura, H.; Matsuki, N. Vagus nerve stimulation enhances perforant path-CA3 synaptic transmission via the activation of β-adrenergic receptors and the locus coeruleus. Int. J. Neuropsychopharmacol. 2012, 15, 523–530. [Google Scholar] [CrossRef] [Green Version]
- Broncel, A.; Bocian, R.; Kłos-Wojtczak, P.; Konopacki, J. Medial septal cholinergic mediation of hippocampal theta rhythm induced by vagal nerve stimulation. PLoS ONE 2018, 13, e0206532. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarczyk, R.; Tejera, D.; Simon, B.J.; Heneka, M.T. Microglia modulation through external vagus nerve stimulation in a murine model of Alzheimer’s disease. J. Neurochem. 2017, 146, 76–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, L.; Lu, K.; Chen, X.; Huang, J.-Y.; Zhang, B.-P.; Zhang, H. Auricular vagus nerve stimulation protects against postoperative cognitive dysfunction by attenuating neuroinflammation and neurodegeneration in aged rats. Neurosci. Lett. 2019, 703, 104–110. [Google Scholar] [CrossRef]
- Clark, K.B.; Naritoku, D.K.; Smith, D.C.; Browning, R.A.; Jensen, R.A. Enhanced recognition memory following vagus nerve stimulation in human subjects. Nat. Neurosci. 1999, 2, 94–98. [Google Scholar] [CrossRef]
- Sjogren, M.J.C.; Hellstrom, P.T.O.; Jonsson, M.A.G.; Runnerstam, M.; Silander, H.C.-S.; Ben-Menachem, E. Cognition-Enhancing Effect of Vagus Nerve Stimulation in Patients with Alzheimer’s Disease: A pilot study. J. Clin. Psychiatry 2002, 63, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Merrill, C.A.; Jonsson, M.A.G.; Minthon, L.; Ejnell, H.; Silander, H.C.-S.; Blennow, K.; Karlsson, M.; Nordlund, A.; Rolstad, S.; Warkentin, S.; et al. Vagus Nerve Stimulation in Patients With Alzheimer’s Disease: Additional Follow-Up Results of a Pilot Study through 1 Year. J. Clin. Psychiatry 2006, 67, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, H.I.L.; Riphagen, J.M.; Razat, C.M.; Wiese, S.; Sack, A.T. Transcutaneous vagus nerve stimulation boosts associative memory in older individuals. Neurobiol. Aging 2015, 36, 1860–1867. [Google Scholar] [CrossRef] [PubMed]
- Barone, L.; Colicchio, G.; Policicchio, D.; Di Clemente, F.; Di Monaco, A.; Meglio, M.; Lanza, G.A.; Crea, F. Effect of Vagal Nerve Stimulation on Systemic Inflammation and Cardiac Autonomic Function in Patients with Refractory Epilepsy. Neuroimmunomodulation 2007, 14, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Aalbers, M.W.; Klinkenberg, S.; Rijkers, K.; Verschuure, P.; Kessels, A.; Aldenkamp, A.; Vles, J.; Majoie, M. The Effects of Vagus Nerve Stimulation on Pro- and Anti-Inflammatory Cytokines in Children with Refractory Epilepsy: An Exploratory Study. Neuroimmunomodulation 2012, 19, 352–358. [Google Scholar] [CrossRef]
- Rush, A.J.; George, M.S.; Sackeim, H.A.; Marangell, L.B.; Husain, M.M.; Giller, C.; Nahas, Z.; Haines, S.; Simpson, R.K.J.; Goodman, R. Vagus nerve stimulation (VNS) for treatment-resistant depressions: A multicenter study. Biol. Psychiatry 2000, 47, 276–286. [Google Scholar] [CrossRef]
- Chaudhry, S.R.; Lendvai, I.S.; Muhammad, S.; Westhofen, P.; Kruppenbacher, J.; Scheef, L.; Boecker, H.; Scheele, D.; Hurlemann, R.; Kinfe, T.M. Inter-ictal assay of peripheral circulating inflammatory mediators in migraine patients under adjunctive cervical non-invasive vagus nerve stimulation (nVNS): A proof-of-concept study. Brain Stimul. 2019, 12, 643–651. [Google Scholar] [CrossRef]
- Mondal, B.; Choudhury, S.; Banerjee, R.; Roy, A.; Chatterjee, K.; Basu, P.; Singh, R.; Halder, S.; Shubham, S.; Baker, S.N.; et al. Non-invasive vagus nerve stimulation improves clinical and molecular biomarkers of Parkinson’s disease in patients with freezing of gait. NPJ Park. Dis. 2021, 7, 1–11. [Google Scholar] [CrossRef]
- Liu, C. Targeting the cholinergic system in Parkinson’s disease. Acta Pharmacol. Sin. 2020, 41, 453–463. [Google Scholar] [CrossRef]
- Kaur, K.; Gill, J.S.; Bansal, P.K.; Deshmukh, R. Neuroinflammation—A major cause for striatal dopaminergic degeneration in Parkinson’s disease. J. Neurol. Sci. 2017, 381, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, E.C.; Standaert, D.G. Ten Unsolved Questions About Neuroinflammation in Parkinson’s Disease. Mov. Disord. 2021, 36, 16–24. [Google Scholar] [CrossRef]
- Pajares, M.; Rojo, A.I.; Manda, G.; Boscá, L.; Cuadrado, A. Inflammation in Parkinson’s Disease: Mechanisms and Therapeutic Implications. Cells 2020, 9, 1687. [Google Scholar] [CrossRef]
- Lu, J.Y.D.; Su, P.; Barber, J.E.M.; Nash, J.E.; Le, A.D.; Liu, F.; Wong, A.H.C. The neuroprotective effect of nicotine in Parkinson’s disease models is associated with inhibiting PARP-1 and caspase-3 cleavage. PeerJ 2017, 5, e3933. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Yang, B.; Tao, T.; Zhang, J.; Liu, Y.; Hu, J.; Fan, Y.; Zhang, G. Activation of α7-nAChRs protects SH-SY5Y cells from 1-methyl-4-phenylpyridinium-induced apoptotic cell death via ERK/p53 signaling pathway. J. Cell. Physiol. 2019, 234, 18480–18491. [Google Scholar] [CrossRef] [PubMed]
- Marogianni, C.; Sokratous, M.; Dardiotis, E.; Hadjigeorgiou, G.M.; Bogdanos, D.; Xiromerisiou, G. Neurodegeneration and Inflammation—An Interesting Interplay in Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 8421. [Google Scholar] [CrossRef] [PubMed]
- Barreto, G.E.; Eiarkov, A.; Moran, V.E. Beneficial effects of nicotine, cotinine and its metabolites as potential agents for Parkinson’s disease. Front. Aging Neurosci. 2015, 6, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganguly, U.; Singh, S.; Pal, S.; Prasad, S.; Agrawal, B.K.; Saini, R.V.; Chakrabarti, S. Alpha-Synuclein as a Biomarker of Parkinson’s Disease: Good, but Not Good Enough. Front. Aging Neurosci. 2021, 13, 702639. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, Y.; Li, Y.; Xu, S.; Tao, T.; Hua, Y.; Zhang, J.; Fan, Y. Activation of α7-nAChRs Promotes the Clearance of α-Synuclein and Protects Against Apoptotic Cell Death Induced by Exogenous α-Synuclein Fibrils. Front. Cell Dev. Biol. 2021, 9, 637319. [Google Scholar] [CrossRef] [PubMed]
- Stuckenholz, V.; Bacher, M.; Balzer-Geldsetzer, M.; Alvarez-Fischer, D.; Oertel, W.H.; Dodel, R.C.; Noelker, C. The α7 nAChR Agonist PNU-282987 Reduces Inflammation and MPTP-Induced Nigral Dopaminergic Cell Loss in Mice. J. Park. Dis. 2013, 3, 161–172. [Google Scholar] [CrossRef]
- Vetel, S.; Foucault-Fruchard, L.; Tronel, C.; Buron, F.; Vergote, J.; Bodard, S.; Routier, S.; Sérrière, S.; Chalon, S. Neuroprotective and anti-inflammatory effects of a therapy combining agonists of nicotinic α7 and σ1 receptors in a rat model of Parkinson’s disease. Neural Regen. Res. 2021, 16, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Farrand, A.Q.; Helke, K.L.; Gregory, R.A.; Gooz, M.; Hinson, V.K.; Boger, H.A. Vagus nerve stimulation improves locomotion and neuronal populations in a model of Parkinson’s disease. Brain Stimul. 2017, 10, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Cao, Z.; Ma, H.; Wang, G.; Wang, X.; Wang, Z.; Yang, Y.; Zhao, H.; Liu, G.; Li, L.; et al. Auricular Vagus Nerve Stimulation Exerts Antiinflammatory Effects and Immune Regulatory Function in a 6-OHDA Model of Parkinson’s Disease. Neurochem. Res. 2018, 43, 2155–2164. [Google Scholar] [CrossRef] [PubMed]
- Kin, I.; Sasaki, T.; Yasuhara, T.; Kameda, M.; Agari, T.; Okazaki, M.; Hosomoto, K.; Okazaki, Y.; Yabuno, S.; Kawauchi, S.; et al. Vagus Nerve Stimulation with Mild Stimulation Intensity Exerts Anti-Inflammatory and Neuroprotective Effects in Parkinson’s Disease Model Rats. Biomed. 2021, 9, 789. [Google Scholar] [CrossRef]
- Mondal, B.; Choudhury, S.; Simon, B.; Baker, M.R.; Kumar, H. Noninvasive vagus nerve stimulation improves gait and reduces freezing of gait in Parkinson’s disease. Mov. Disord. 2019, 34, 917–918. [Google Scholar] [CrossRef] [PubMed]
- Tanimura, A.; Du, Y.; Kondapalli, J.; Wokosin, D.L.; Surmeier, D.J. Cholinergic Interneurons Amplify Thalamostriatal Excitation of Striatal Indirect Pathway Neurons in Parkinson’s Disease Models. Neuron 2019, 101, 444–458.e6. [Google Scholar] [CrossRef] [Green Version]
- Pracucci, E.; Pillai, V.; Lamers, D.; Parra, R.; Landi, S. Neuroinflammation: A Signature or a Cause of Epilepsy? Int. J. Mol. Sci. 2021, 22, 6981. [Google Scholar] [CrossRef]
- Vezzani, A.; Balosso, S.; Ravizza, T. Neuroinflammatory pathways as treatment targets and biomarkers in epilepsy. Nat. Rev. Neurol. 2019, 15, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, I.E.; Berkovic, S.; Capovilla, G.; Connolly, M.B.; French, J.; Guilhoto, L.; Hirsch, E.; Jain, S.; Mathern, G.W.; Moshe, S.l.; et al. ILAE classification of the epilepsies: Position paper of the ILAE Commission for Classification and Terminology. Epilepsia 2017, 58, 512–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, I.; Mlsna, L.M.; Yoon, S.; Le, B.; Yu, S.; Xu, D.; Koh, S. A postnatal peak in microglial development in the mouse hippocampus is correlated with heightened sensitivity to seizure triggers. Brain Behav. 2015, 5, e00403. [Google Scholar] [CrossRef]
- Maroso, M.; Balosso, S.; Ravizza, T.; Liu, J.; Aronica, E.; Iyer, A.M.; Rossetti, C.; Molteni, M.; Casalgrandi, M.; Manfredi, A.A.; et al. Toll-like receptor 4 and high-mobility group box-1 are involved in ictogenesis and can be targeted to reduce seizures. Nat. Med. 2010, 16, 413–419. [Google Scholar] [CrossRef] [Green Version]
- Bonaz, B.; Picq, C.; Sinniger, V.; Mayol, J.-F.; Clarençon, D. Vagus nerve stimulation: From epilepsy to the cholinergic anti-inflammatory pathway. Neurogastroenterol. Motil. 2013, 25, 208–221. [Google Scholar] [CrossRef]
- Wang, Y.; Tan, B.; Wang, Y.; Chen, Z. Cholinergic Signaling, Neural Excitability, and Epilepsy. Molecules 2021, 26, 2258. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Liu, D.-G.; Ye, X.-M. Nicotinic Acetylcholine Receptor α7 Subunit Is an Essential Regulator of Seizure Susceptibility. Front. Neurol. 2021, 12, 656752. [Google Scholar] [CrossRef] [PubMed]
- Meller, S.; Brandt, C.; Theilmann, W.; Klein, J.; Löscher, W. Commonalities and differences in extracellular levels of hippocampal acetylcholine and amino acid neurotransmitters during status epilepticus and subsequent epileptogenesis in two rat models of temporal lobe epilepsy. Brain Res. 2019, 1712, 109–123. [Google Scholar] [CrossRef]
- Gnatek, Y.; Zimmerman, G.; Goll, Y.; Najami, N.; Soreq, H.; Friedman, A. Acetylcholinesterase loosens the brain’s cholinergic anti-inflammatory response and promotes epileptogenesis. Front. Mol. Neurosci. 2012, 5, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, N.K.; Kaur, S.; Goel, R.K. Exploring the ameliorative role of α7 neuronal nicotinic acetylcholine receptor modulation in epilepsy and associated comorbidities in post-PTZ-kindled mice. Epilepsy Behav. 2020, 103, 106862. [Google Scholar] [CrossRef] [PubMed]
- Feuerbach, D.; Lingenhoehl, K.; Olpe, H.-R.; Vassout, A.; Gentsch, C.; Chaperon, F.; Nozulak, J.; Enz, A.; Bilbe, G.; McAllister, K.; et al. The selective nicotinic acetylcholine receptor α7 agonist JN403 is active in animal models of cognition, sensory gating, epilepsy and pain. Neuropharmacology 2009, 56, 254–263. [Google Scholar] [CrossRef]
- Zhang, Y.; Cao, S.-X.; Sun, P.; He, H.-Y.; Yang, C.-H.; Chen, X.-J.; Shen, C.-J.; Wang, X.-D.; Chen, Z.; Berg, D.K.; et al. Loss of MeCP2 in cholinergic neurons causes part of RTT-like phenotypes via α7 receptor in hippocampus. Cell Res. 2016, 26, 728–742. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.-J.; Zhang, T.-Y.; Liu, H.-T.; Huang, Z.-X.; Teng, J.-M.; Deng, J.-X.; Zhong, J.-G.; Qian, X.; Sheng, X.-W.; Ding, J.-Q.; et al. Cytisine Exerts an Anti-Epileptic Effect via α7nAChRs in a Rat Model of Temporal Lobe Epilepsy. Front. Pharmacol. 2021, 12, 706225. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Carbonell, L.; Faulkner, H.; Higgins, S.; Koutroumanidis, M.; Leschziner, G. Vagus nerve stimulation for drug-resistant epilepsy. Pract. Neurol. 2020, 20, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Majoie, H.J.M.; Rijkers, K.; Berfelo, M.W.; Hulsman, J.A.R.J.; Myint, A.; Schwarz, M.; Vles, J.S.H. Vagus Nerve Stimulation in Refractory Epilepsy: Effects on Pro- and Anti-Inflammatory Cytokines in Peripheral Blood. Neuroimmunomodulation 2011, 18, 52–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boswijk, E.; Franssen, R.; Vijgen, G.H.E.J.; Wierts, R.; Van Der Pol, J.A.J.; Mingels, A.M.A.; Cornips, E.M.J.; Majoie, M.H.J.M.; van Marken Lichtenbelt, W.D.; Mottaghy, F.M.; et al. Short-term discontinuation of vagal nerve stimulation alters 18F-FDG blood pool activity: An exploratory interventional study in epilepsy patients. EJNMMI Res. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Voineskos, D.; Daskalakis, Z.J.; Blumberger, D.M. Management of Treatment-Resistant Depression: Challenges and Strategies. Neuropsychiatr. Dis. Treat. 2020, 16, 221–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köhler, C.A.; Freitas, T.H.; Maes, M.; De Andrade, N.Q.; Liu, C.S.; Fernandes, B.S.; Stubbs, B.; Solmi, M.; Veronese, N.; Herrmann, N.; et al. Peripheral cytokine and chemokine alterations in depression: A meta-analysis of 82 studies. Acta Psychiatr. Scand. 2017, 135, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Dudek, K.A.; Dion-Albert, L.; Lebel, M.; LeClair, K.; Labrecque, S.; Tuck, E.; Ferrer-Pérez, C.; Golden, S.A.; Tamminga, C.; Turecki, G.; et al. Molecular adaptations of the blood–brain barrier promote stress resilience vs. depression. Proc. Natl. Acad. Sci. USA 2020, 117, 3326–3336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ancelin, M.-L.; Scali, J.; Norton, J.; Ritchie, K.; Dupuy, A.M.; Chaudieu, I.; Ryan, J. Heterogeneity in HPA axis dysregulation and serotonergic vulnerability to depression. Psychoneuroendocrinology 2017, 77, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-C.; Yao, W.; Ren, Q.; Yang, C.; Dong, C.; Ma, M.; Wu, J.; Hashimoto, K. Depression-like phenotype by deletion of α7 nicotinic acetylcholine receptor: Role of BDNF-TrkB in nucleus accumbens. Sci. Rep. 2016, 6, 36705. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Liu, X.; Chang, D.; Zhang, X.; Lian, H.; Du, X.; Gao, L. Low-Dose Ketamine Improves LPS-Induced Depression-like Behavior in Rats by Activating Cholinergic Anti-inflammatory Pathways. ACS Chem. Neurosci. 2020, 11, 752–762. [Google Scholar] [CrossRef]
- Moriguchi, S.; Inagaki, R.; Yi, L.; Shibata, M.; Sakagami, H.; Fukunaga, K. Nicotine Rescues Depressive-like Behaviors via α7-type Nicotinic Acetylcholine Receptor Activation in CaMKIV Null Mice. Mol. Neurobiol. 2020, 57, 4929–4940. [Google Scholar] [CrossRef]
- Alzarea, S.; Rahman, S. Alpha-7 nicotinic receptor allosteric modulator PNU120596 prevents lipopolysaccharide-induced anxiety, cognitive deficit and depression-like behaviors in mice. Behav. Brain Res. 2019, 366, 19–28. [Google Scholar] [CrossRef]
- Mineur, Y.S.; Fote, G.M.; Blakeman, S.; Cahuzac, E.L.M.; Newbold, S.A.; Picciotto, M.R. Multiple Nicotinic Acetylcholine Receptor Subtypes in the Mouse Amygdala Regulate Affective Behaviors and Response to Social Stress. Neuropsychopharmacology 2016, 41, 1579–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mineur, Y.S.; Mose, T.N.; Blakeman, S.; Picciotto, M.R. Hippocampal α7 nicotinic ACh receptors contribute to modulation of depression-like behaviour in C57BL/6J mice. Br. J. Pharmacol. 2018, 175, 1903–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mineur, Y.S.; Picciotto, M.R. Nicotine receptors and depression: Revisiting and revising the cholinergic hypothesis. Trends Pharmacol. Sci. 2010, 31, 580–586. [Google Scholar] [CrossRef] [Green Version]
- Pomara, N.; Bruno, D.; Plaska, C.R.; Pillai, A.; Ramos-Cejudo, J.; Osorio, R.; Imbimbo, B.P.; Heslegrave, A.; Zetterberg, H.; Blennow, K. Evidence of upregulation of the cholinergic anti-inflammatory pathway in late-life depression. J. Affect. Disord. 2021, 286, 275–281. [Google Scholar] [CrossRef]
- Zhao, D.; Xu, X.; Pan, L.; Zhu, W.; Fu, X.; Guo, L.; Lu, Q.; Wang, J. Pharmacologic activation of cholinergic alpha7 nicotinic receptors mitigates depressive-like behavior in a mouse model of chronic stress. J. Neuroinflamm. 2017, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.S.; Koth, A.P.; Parfitt, G.M.; Cordeiro, M.F.; Peixoto, C.S.; Soubhia, A.; Moreira, F.P.; Wiener, C.D.; Oses, J.P.; Kaszubowski, E.; et al. Enhanced cholinergic-tone during the stress induce a depressive-like state in mice. Behav. Brain Res. 2018, 347, 17–25. [Google Scholar] [CrossRef]
- Belovicova, K.; Bogi, E.; Csatlosova, K.; Dubovicky, M. Animal tests for anxiety-like and depression-like behavior in rats. Interdiscip. Toxicol. 2017, 10, 40–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castagné, V.; Moser, P.; Roux, S.; Porsolt, R.D. Rodent Models of Depression: Forced Swim and Tail Suspension Behavioral Despair Tests in Rats and Mice. Curr. Protoc. Pharmacol. 2010, 49, 5.8.1–5.8.14. [Google Scholar] [CrossRef] [PubMed]
- López-Rubalcava, C. Strain Differences in the Behavioral Effects of Antidepressant Drugs in the Rat Forced Swimming Test. Neuropsychopharmacology 2000, 22, 191–199. [Google Scholar] [CrossRef]
- Baranowska, U.; Wiśniewska, R.J. The α7-nACh nicotinic receptor and its role in memory and selected diseases of the central nervous system. Postepy Hig. I Med. Dosw. 2017, 71, 633–648. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.-Q.; Zhang, W.-J.; Su, D.-F.; Zhang, G.-Q.; Miao, C.-Y. Cellular responses and functions of α7 nicotinic acetylcholine receptor activation in the brain: A narrative review. Ann. Transl. Med. 2021, 9, 509. [Google Scholar] [CrossRef] [PubMed]
- Jiang, A.; Su, P.; Li, S.; Wong, A.H.C.; Liu, F. Disrupting the α7nAChR–NR2A protein complex exerts antidepressant-like effects. Mol. Brain 2021, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Conway, C.R.; Kumar, A.; Xiong, W.; Bunker, M.; Aaronson, S.T.; Rush, A.J. Chronic Vagus Nerve Stimulation Significantly Improves Quality of Life in Treatment-Resistant Major Depression. J. Clin. Psychiatry 2018, 79. [Google Scholar] [CrossRef] [Green Version]
- Cristancho, P.; Cristancho, M.A.; Baltuch, G.H.; Thase, M.E.; O’Reardon, J.P. Effectiveness and Safety of Vagus Nerve Stimulation for Severe Treatment-Resistant Major Depression in Clinical Practice After FDA Approval: Outcomes at 1 year. J. Clin. Psychiatry 2011, 72, 1376–1382. [Google Scholar] [CrossRef] [PubMed]
- Trottier-Duclos, F.; Jodoin, V.D.; Fournier-Gosselin, M.-P.; Richer, F.; Desjardins, N.; Tieu, S.; Lespérance, P. A 6-Year Follow-up Study of Vagus Nerve Stimulation Effect on Quality of Life in Treatment-Resistant Depression: A Pilot Study. J. ECT 2018, 34, e58–e60. [Google Scholar] [CrossRef] [PubMed]
- Moeller, S.; Wang, R.; Aydin, M.; Lam, A.P.; Sitter, A.; Grüter, J.; Philipsen, A.; Müller, H.H.O. Rapid titration protocol—Experiences with a dynamic novel titration regime for vagus nerve stimulation in a group of depressive patients. J. Clin. Neurosci. 2020, 74, 262–264. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.-H.; Yang, M.-H.; Zhang, G.-Z.; Wang, X.-X.; Li, B.; Li, M.; Woelfer, M.; Walter, M.; Wang, L. Neural networks and the anti-inflammatory effect of transcutaneous auricular vagus nerve stimulation in depression. J. Neuroinflamm. 2020, 17, 1–11. [Google Scholar] [CrossRef]
- Cavestro, C.; Ferrero, M.; Mandrino, S.; Di Tavi, M.; Rota, E. Novelty in Inflammation and Immunomodulation in Migraine. Curr. Pharm. Des. 2019, 25, 2919–2936. [Google Scholar] [CrossRef]
- Friedman, M.H. Local Inflammation as a Mediator of Migraine and Tension-Type Headache. Headache J. Head Face Pain 2004, 44, 767–771. [Google Scholar] [CrossRef]
- Perini, F.; D’Andrea, G.; Galloni, E.; Pignatelli, F.; Billo, G.; Alba, S.; Bussone, G.; Toso, V. Plasma Cytokine Levels in Migraineurs and Controls. Headache J. Head Face Pain 2005, 45, 926–931. [Google Scholar] [CrossRef]
- Liu, Q.; Liu, C.; Jiang, L.; Li, M.; Long, T.; He, W.; Qin, G.; Chen, L.; Zhou, J. α7 Nicotinic acetylcholine receptor-mediated anti-inflammatory effect in a chronic migraine rat model via the attenuation of glial cell activation. J. Pain Res. 2018, 11, 1129–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beh, S.C.; Friedman, D.I. Acute vestibular migraine treatment with noninvasive vagus nerve stimulation. Neurology 2019, 93, e1715–e1719. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.-H.; Huang, Y.-C.; Huang, L.-T.; Chen, R.-M.; Chen, C. Cervical Noninvasive Vagus Nerve Stimulation for Migraine and Cluster Headache: A Systematic Review and Meta-Analysis. Neuromodulation Technol. Neural Interface 2020, 23, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Diener, H.-C.; Goadsby, P.J.; Ashina, M.; Al-Karagholi, M.A.-M.; Sinclair, A.; Mitsikostas, D.; Magis, D.; Pozo-Rosich, P.; Sieira, P.I.; Làinez, M.J.; et al. Non-invasive vagus nerve stimulation (nVNS) for the preventive treatment of episodic migraine: The multicentre, double-blind, randomised, sham-controlled PREMIUM trial. Cephalalgia 2019, 39, 1475–1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boström, A.; Scheele, D.; Stoffel-Wagner, B.; Hönig, F.; Chaudhry, S.R.; Muhammad, S.; Hurlemann, R.; Krauss, J.K.; Lendvai, I.S.; Chakravarthy, K.V.; et al. Saliva molecular inflammatory profiling in female migraine patients responsive to adjunctive cervical non-invasive vagus nerve stimulation: The MOXY Study. J. Transl. Med. 2019, 17, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fond, G.; Lançon, C.; Korchia, T.; Auquier, P.; Boyer, L. The Role of Inflammation in the Treatment of Schizophrenia. Front. Psychiatry 2020, 11, 160. [Google Scholar] [CrossRef] [Green Version]
- Jackson, A.J.; Miller, B.J. Meta-analysis of total and differential white blood cell counts in schizophrenia. Acta Psychiatr. Scand. 2020, 142, 18–26. [Google Scholar] [CrossRef]
- Miller, B.J.; Buckley, P.; Seabolt, W.; Mellor, A.L.; Kirkpatrick, B. Metaanálisis de los estudios publicados sobre alteraciones de los valores de las citocinas en la esquizofrenia: Efectos del estado clínico y de los antipsicóticos. Psiquiatr. Biológica 2011, 18, 127–136. [Google Scholar] [CrossRef]
- Momtazmanesh, S.; Zare-Shahabadi, A.; Rezaei, N. Cytokine Alterations in Schizophrenia: An Updated Review. Front. Psychiatry 2019, 10, 892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.K.; Miller, B.J. Meta-analysis of Cerebrospinal Fluid Cytokine and Tryptophan Catabolite Alterations in Psychiatric Patients: Comparisons between Schizophrenia, Bipolar Disorder, and Depression. Schizophr. Bull. 2018, 44, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Busse, S.; Busse, M.; Schiltz, K.; Bielau, H.; Gos, T.; Brisch, R.; Mawrin, C.; Schmitt, A.; Jordan, W.; Müller, U.J.; et al. Different distribution patterns of lymphocytes and microglia in the hippocampus of patients with residual versus paranoid schizophrenia: Further evidence for disease course-related immune alterations? Brain Behav. Immun. 2012, 26, 1273–1279. [Google Scholar] [CrossRef] [PubMed]
- Terry, A.V., Jr.; Callahan, P.M. α7 nicotinic acetylcholine receptors as therapeutic targets in schizophrenia: Update on animal and clinical studies and strategies for the future. Neuropharmacology 2020, 170, 108053. [Google Scholar] [CrossRef]
- Koukouli, F.; Rooy, M.; Tziotis, D.; Sailor, K.A.; O’Neill, H.C.; Levenga, J.; Witte, M.; Nilges, M.; Changeux, J.-P.; Hoeffer, C.A.; et al. Nicotine reverses hypofrontality in animal models of addiction and schizophrenia. Nat. Med. 2017, 23, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Freedman, R.; Olincy, A.; Buchanan, R.W.; Harris, J.G.; Gold, J.M.; Johnson, L.; Allensworth, D.; Guzman-Bonilla, A.; Clement, B.; Ball, M.P.; et al. Initial Phase 2 Trial of a Nicotinic Agonist in Schizophrenia. Am. J. Psychiatry 2008, 165, 1040–1047. [Google Scholar] [CrossRef] [Green Version]
- Tregellas, J.R.; Wylie, K.P. Alpha7 Nicotinic Receptors as Therapeutic Targets in Schizophrenia. Nicotine Tob. Res. 2019, 21, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Kem, W.R.; Olincy, A.; Johnson, L.; Harris, J.; Wagner, B.D.; Buchanan, R.W.; Christians, U.; Freedman, R. Pharmacokinetic Limitations on Effects of an Alpha7-Nicotinic Receptor Agonist in Schizophrenia: Randomized Trial with an Extended-Release Formulation. Neuropsychopharmacology 2018, 43, 583–589. [Google Scholar] [CrossRef] [Green Version]
- Perez, S.M.; Carreno, F.R.; Frazer, A.; Lodge, D.J. Vagal Nerve Stimulation Reverses Aberrant Dopamine System Function in the Methylazoxymethanol Acetate Rodent Model of Schizophrenia. J. Neurosci. 2014, 34, 9261–9267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, A.; Wolff-Menzler, C.; Pfeiffer, S.; Falkai, P.; Weidinger, E.; Jobst, A.; Hoell, I.; Malchow, B.; Yeganeh-Doost, P.; Strube, W.; et al. Transcutaneous noninvasive vagus nerve stimulation (tVNS) in the treatment of schizophrenia: A bicentric randomized controlled pilot study. Eur. Arch. Psychiatry Clin. Neurosci. 2015, 265, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Priyadarshini, M.; Arivarasu, N.A.; Shah, A.; Tabrez, S.; Priyamvada, S.; Aatif, M. MicroRNA: Novel modulators of the cholinergic anti-inflammatory pathway. Anti-Inflamm. Anti-Allergy Agents Med. Chem. 2013, 12, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; He, Z.; Wang, J. MicroRNA-124: A Key Player in Microglia-Mediated Inflammation in Neurological Diseases. Front. Cell. Neurosci. 2021, 15, 771898. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, H.; Nguyen, H.C.B.; Zhang, S.; Dill, B.D.; Molina, H.; Tavazoie, S.F. Modulated Expression of Specific tRNAs Drives Gene Expression and Cancer Progression. Cell 2016, 165, 1416–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reale, M.; Costantini, E. Cholinergic Modulation of the Immune System in Neuroinflammatory Diseases. Diseases 2021, 9, 29. [Google Scholar] [CrossRef]
- Enadorp, B.; Esoreq, H. Predicted overlapping microRNA regulators of acetylcholine packaging and degradation in neuroinflammation-related disorders. Front. Mol. Neurosci. 2014, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Shaked, I.; Meerson, A.; Wolf, Y.; Avni, R.; Greenberg, D.; Gilboa-Geffen, A.; Soreq, H. MicroRNA-132 Potentiates Cholinergic Anti-Inflammatory Signaling by Targeting Acetylcholinesterase. Immunity 2009, 31, 965–973. [Google Scholar] [CrossRef] [Green Version]
- Maharshak, N.; Shenhar-Tsarfaty, S.; Aroyo, N.; Orpaz, N.; Guberman, I.; Canaani, J.; Halpern, Z.; Dotan, I.; Berliner, S.; Soreq, H. MicroRNA-132 Modulates Cholinergic Signaling and Inflammation in Human Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2013, 19, 1346–1353. [Google Scholar] [CrossRef] [PubMed]
- Hamza, H.; Abdullah, A. MicroRNA-132 suppresses autoimmune encephalomyelitis by inducing cholinergic anti-inflammation: A new Ahr-based exploration. Eur. J. Immunol. 2013, 43, 2771–2782. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.-T.; Wang, J.; Lu, W.; Cao, Y.; Cai, W. Downregulated expression of microRNA-124 in pediatric intestinal failure patients modulates macrophages activation by inhibiting STAT3 and AChE. Cell Death Dis. 2016, 7, e2521. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, Q.; Gui, H.; Xu, D.-P.; Yang, Y.-L.; Su, D.-F.; Liu, X. MicroRNA-124 mediates the cholinergic anti-inflammatory action through inhibiting the production of pro-inflammatory cytokines. Cell Res. 2013, 23, 1270–1283. [Google Scholar] [CrossRef] [Green Version]
- Walgrave, H.; Zhou, L.; De Strooper, B.; Salta, E. The promise of microRNA-based therapies in Alzheimer’s disease: Challenges and perspectives. Mol. Neurodegener. 2021, 16, 1–16. [Google Scholar] [CrossRef]
- Winek, K.; Lobentanzer, S.; Nadorp, B.; Dubnov, S.; Dames, C.; Jagdmann, S.; Moshitzky, G.; Hotter, B.; Meisel, C.; Greenberg, D.S.; et al. Transfer RNA fragments replace microRNA regulators of the cholinergic poststroke immune blockade. Proc. Natl. Acad. Sci. USA 2020, 117, 32606–32616. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Disease | Range of Parameters | Approvals | Inflammatory Profile | Refs |
|---|---|---|---|---|
| Human Epilepsy | 0.75–1.75 mA, 30 Hz | FDA | no significant changes | [95,96] |
| Human Depression | 0.25 mA, 25 Hz | FDA | non evaluated | [97] |
| Human Migraine | 0.25 mA, 25 Hz | FDA | ↓IL-1β plasma levels | [98] |
| Parkinson’s disease | maximum output of 60 mA, 25 Hz | not approved | ↓ TNF- α | [99] |
| Alzheimer’s disease | 0.4–0.8 mA, 20 Hz | not approved | non evaluated | [92,93] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos-Martínez, I.E.; Rodríguez, M.C.; Cerbón, M.; Ramos-Martínez, J.C.; Ramos-Martínez, E.G. Role of the Cholinergic Anti-Inflammatory Reflex in Central Nervous System Diseases. Int. J. Mol. Sci. 2021, 22, 13427. https://doi.org/10.3390/ijms222413427
Ramos-Martínez IE, Rodríguez MC, Cerbón M, Ramos-Martínez JC, Ramos-Martínez EG. Role of the Cholinergic Anti-Inflammatory Reflex in Central Nervous System Diseases. International Journal of Molecular Sciences. 2021; 22(24):13427. https://doi.org/10.3390/ijms222413427
Chicago/Turabian StyleRamos-Martínez, Ivan Emmanuel, María Carmen Rodríguez, Marco Cerbón, Juan Carlos Ramos-Martínez, and Edgar Gustavo Ramos-Martínez. 2021. "Role of the Cholinergic Anti-Inflammatory Reflex in Central Nervous System Diseases" International Journal of Molecular Sciences 22, no. 24: 13427. https://doi.org/10.3390/ijms222413427
APA StyleRamos-Martínez, I. E., Rodríguez, M. C., Cerbón, M., Ramos-Martínez, J. C., & Ramos-Martínez, E. G. (2021). Role of the Cholinergic Anti-Inflammatory Reflex in Central Nervous System Diseases. International Journal of Molecular Sciences, 22(24), 13427. https://doi.org/10.3390/ijms222413427