
Akt Signaling Pathway Is Activated in the Minor Salivary Glands of Patients with Primary Sjögren’s Syndrome

 , ,
, ,
Abstract
:1. Introduction
2. Results
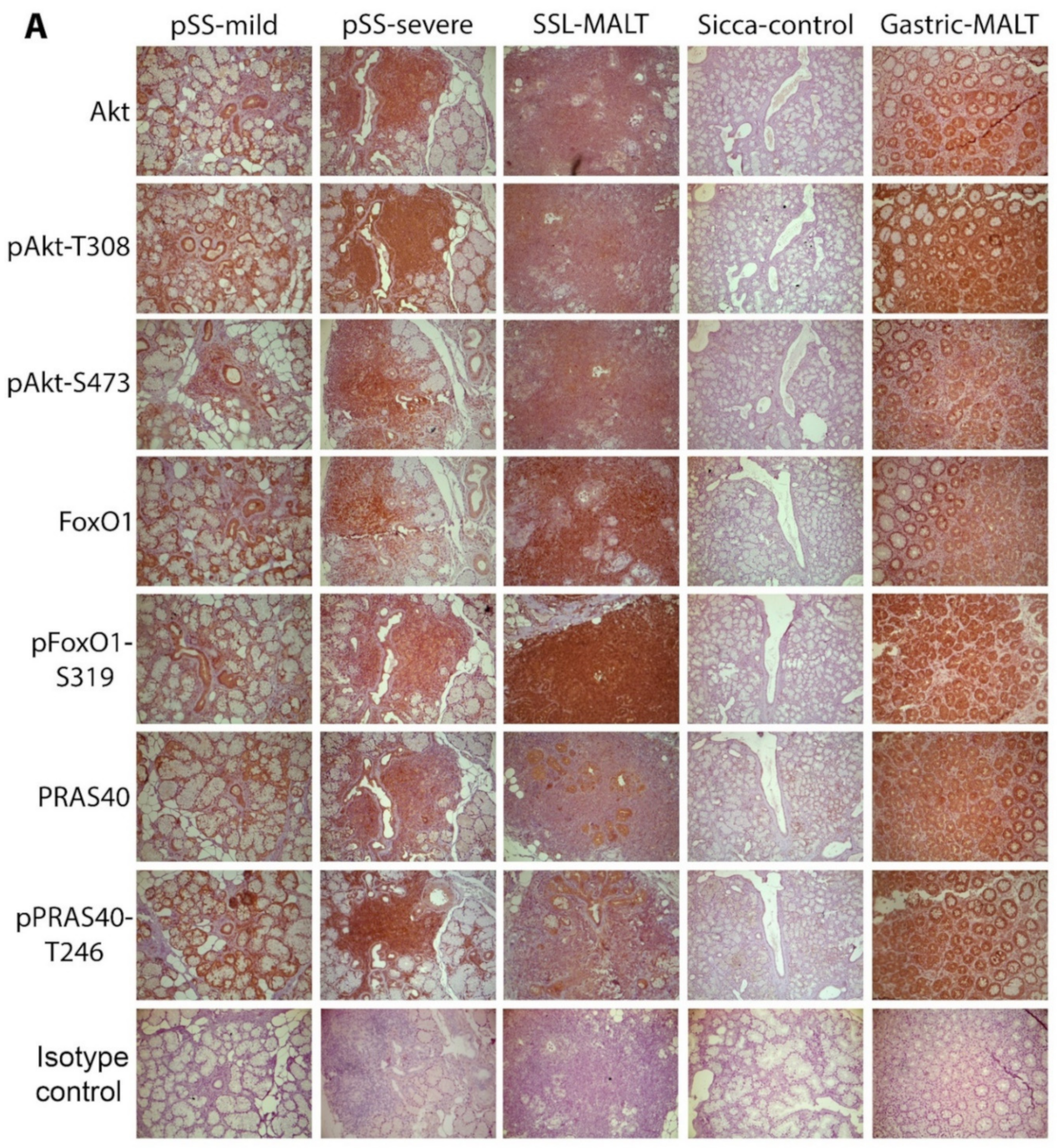
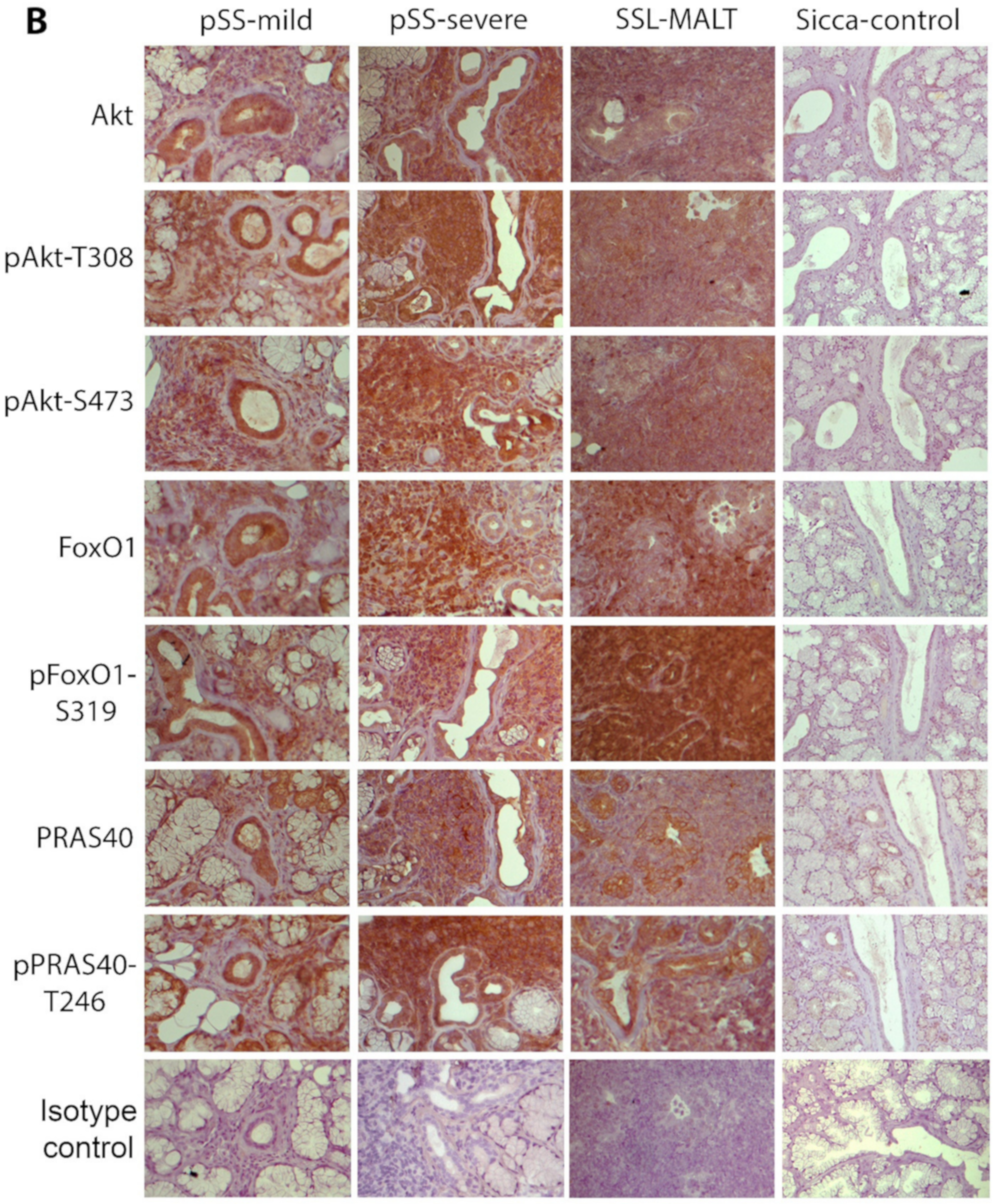
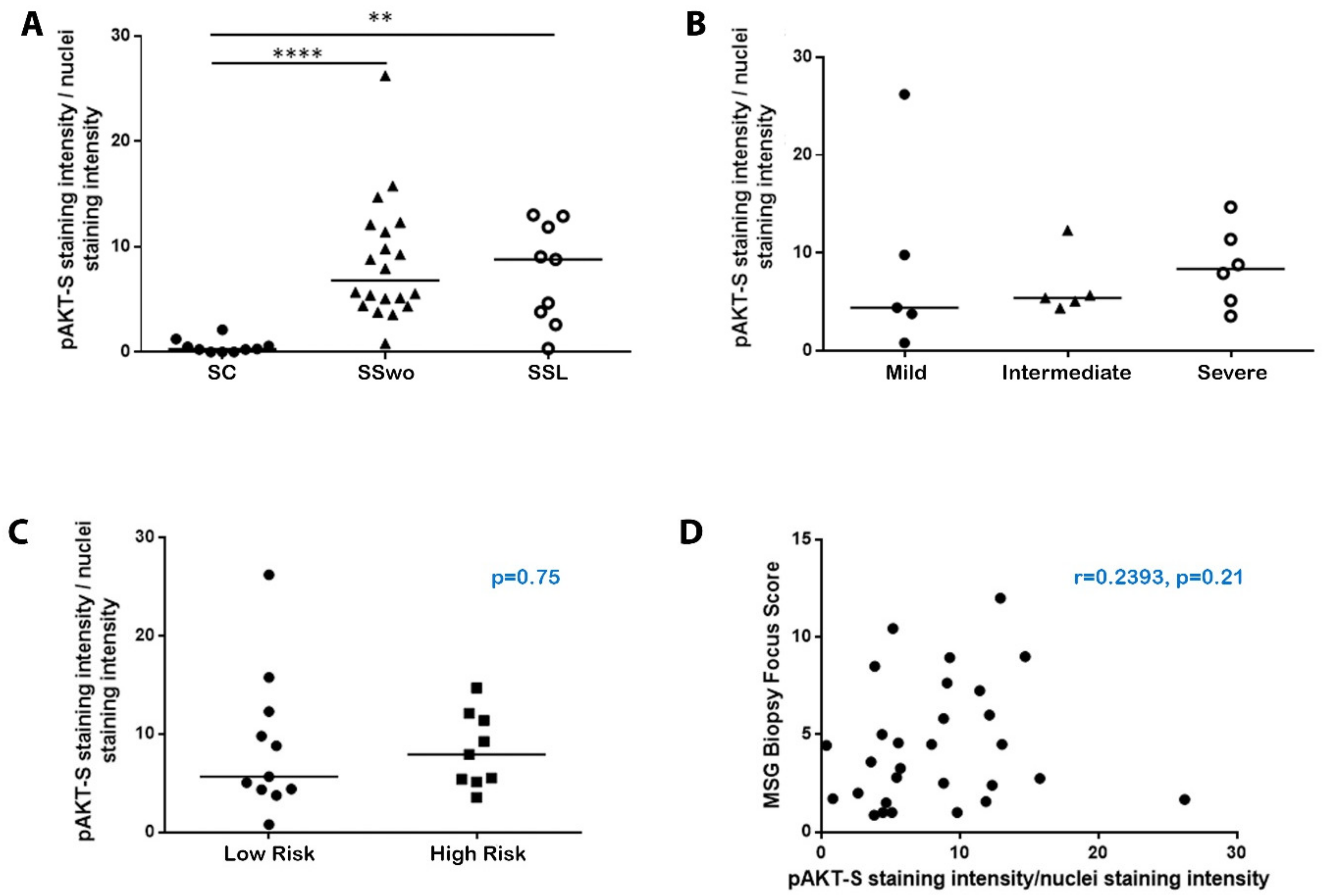
2.1. The Molecules Related to Active Akt Pathway Are Expressed in pSS Patients, but Not in Sicca-Controls
2.2. The Activation of the Akt Pathway in pSS Does Not Associate with Histologic, Clinical, or Other Disease Features, including NHL Development
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Antibodies
4.3. Immunohistochemical Detection of the Molecules Implicated in the Akt Signaling Pathway
4.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tzioufas, A.G.; Kapsogeorgou, E.K.; Moutsopoulos, H.M. Pathogenesis of Sjogren’s syndrome: What we know and what we should learn. J. Autoimmun. 2012, 39, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Papageorgiou, A.; Voulgarelis, M.; Tzioufas, A.G. Clinical picture, outcome and predictive factors of lymphoma in Sjgren syndrome. Autoimmun. Rev. 2015, 14, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Christodoulou, M.I.; Kapsogeorgou, E.K.; Moutsopoulos, H.M. Characteristics of the minor salivary gland infiltrates in Sjogren’s syndrome. J. Autoimmun. 2010, 34, 400–407. [Google Scholar] [CrossRef]
- Bombardieri, M.; Pitzalis, C. Ectopic lymphoid neogenesis and lymphoid chemokines in Sjogren’s syndrome: At the interplay between chronic inflammation, autoimmunity and lymphomagenesis. Curr. Pharm. Biotechnol. 2012, 13, 1989–1996. [Google Scholar] [CrossRef] [PubMed]
- Gerli, R.; Muscat, C.; Giansanti, M.; Danieli, M.G.; Sciuto, M.; Gabrielli, A.; Fiandra, E.; Vitali, C. Quantitative assessment of salivary gland inflammatory infiltration in primary Sjogren’s syndrome: Its relationship to different demographic, clinical and serological features of the disorder. Br. J. Rheumatol. 1997, 36, 969–975. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.; Ikebe-Hiroki, A.; Shinohara, M.; Ohyama, Y.; Mouri, T.; Sasaki, M.; Shirasuna, K.; Nomoto, K. An association between salivary gland disease and serological abnormalities in Sjogren’s syndrome. J. Oral Pathol. Med. 1997, 26, 426–430. [Google Scholar] [CrossRef]
- Risselada, A.P.; Kruize, A.A.; Goldschmeding, R.; Lafeber, F.P.; Bijlsma, J.W.; van Roon, J.A. The prognostic value of routinely performed minor salivary gland assessments in primary Sjogren’s syndrome. Ann. Rheum. Dis. 2014, 73, 1537–1540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theander, E.; Vasaitis, L.; Baecklund, E.; Nordmark, G.; Warfvinge, G.; Liedholm, R.; Brokstad, K.; Jonsson, R.; Jonsson, M.V. Lymphoid organisation in labial salivary gland biopsies is a possible predictor for the development of malignant lymphoma in primary Sjogren’s syndrome. Ann. Rheum. Dis. 2011, 70, 1363–1368. [Google Scholar] [CrossRef] [PubMed]
- Kapsogeorgou, E.K.; Tzioufas, A.G. Glandular epithelium: Innocent bystander or leading actor. In Sjogren’s Syndrome: Novel Insights in Pathogenic, Clinical and Therapeutic Aspects; Alunno, A., Bartoloni, E., Gerli, R., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 189–198. [Google Scholar]
- Bellacosa, A.; Chan, T.O.; Ahmed, N.N.; Datta, K.; Malstrom, S.; Stokoe, D.; McCormick, F.; Feng, J.; Tsichlis, P. Akt activation by growth factors is a multiple-step process: The role of the PH domain. Oncogene 1998, 17, 313–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manning, B.D.; Toker, A. AKT/PKB Signaling: Navigating the Network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef] [Green Version]
- Risso, G.; Blaustein, M.; Pozzi, B.; Mammi, P.; Srebrow, A. Akt/PKB: One kinase, many modifications. Biochem. J. 2015, 468, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Sarbassov, D.D.; Guertin, D.A.; Ali, S.M.; Sabatini, D.M. Phosphorylation and regulation of Akt/PKB by the rictor-mTOR complex. Science 2005, 307, 1098–1101. [Google Scholar] [CrossRef] [Green Version]
- Patel, R.K.; Mohan, C. PI3K/AKT signaling and systemic autoimmunity. Immunol. Res. 2005, 31, 47–55. [Google Scholar] [CrossRef]
- Hawkins, P.T.; Stephens, L.R. PI3K signalling in inflammation. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2015, 1851, 882–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fruman, D.A.; Rommel, C. PI3K and cancer: Lessons, challenges and opportunities. Nat. Rev. Drug Discov. 2014, 13, 140–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blachly, J.S.; Baiocchi, R.A. Targeting PI3-kinase (PI3K), AKT and mTOR axis in lymphoma. Br. J. Haematol. 2014, 167, 19–32. [Google Scholar] [CrossRef]
- Naruse, T.; Yanamoto, S.; Okuyama, K.; Yamashita, K.; Omori, K.; Nakao, Y.; Yamada, S.I.; Umeda, M. Therapeutic implication of mTORC2 in oral squamous cell carcinoma. Oral Oncol. 2017, 65, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Kawakami, A.; Ida, H.; Koji, T.; Eguchi, K. EGF activates PI3K-Akt and NF-kappaB via distinct pathways in salivary epithelial cells in Sjogren’s syndrome. Rheumatol. Int. 2007, 28, 127–136. [Google Scholar] [CrossRef]
- Nakamura, H.; Horai, Y.; Suzuki, T.; Okada, A.; Ichinose, K.; Yamasaki, S.; Koji, T.; Kawakami, A. TLR3-mediated apoptosis and activation of phosphorylated Akt in the salivary gland epithelial cells of primary Sjogren’s syndrome patients. Rheumatol. Int. 2013, 33, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Blokland, S.L.M.; Hillen, M.R.; Wichers, C.G.K.; Zimmermann, M.; Kruize, A.A.; Radstake, T.; Broen, J.C.A.; van Roon, J.A.G. Increased mTORC1 activation in salivary gland B cells and T cells from patients with Sjogren’s syndrome: mTOR inhibition as a novel therapeutic strategy to halt immunopathology? RMD Open 2019, 5, e000701. [Google Scholar] [CrossRef] [Green Version]
- Nayar, S.; Campos, J.; Smith, C.G.; Iannizzotto, V.; Gardner, D.H.; Colafrancesco, S.; Pipi, E.; Kollert, F.; Hunter, K.J.; Brewer, C.; et al. Phosphatidylinositol 3-kinase delta pathway: A novel therapeutic target for Sjogren’s syndrome. Ann. Rheum. Dis. 2019, 78, 249–260. [Google Scholar] [CrossRef] [Green Version]
- Bartok, B.; Boyle, D.L.; Liu, Y.; Ren, P.; Ball, S.T.; Bugbee, W.D.; Rommel, C.; Firestein, G.S. PI3 kinase delta is a key regulator of synoviocyte function in rheumatoid arthritis. Am. J. Pathol. 2012, 180, 1906–1916. [Google Scholar] [CrossRef] [PubMed]
- Karonitsch, T.; Kandasamy, R.K.; Kartnig, F.; Herdy, B.; Dalwigk, K.; Niederreiter, B.; Holinka, J.; Sevelda, F.; Windhager, R.; Bilban, M.; et al. mTOR Senses Environmental Cues to Shape the Fibroblast-like Synoviocyte Response to Inflammation. Cell Rep. 2018, 23, 2157–2167. [Google Scholar] [CrossRef]
- Mitra, A.; Luna, J.I.; Marusina, A.I.; Merleev, A.; Kundu-Raychaudhuri, S.; Fiorentino, D.; Raychaudhuri, S.P.; Maverakis, E. Dual mTOR Inhibition Is Required to Prevent TGF-beta-Mediated Fibrosis: Implications for Scleroderma. J. Investig. Dermatol. 2015, 135, 2873–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, J.B.; Kuechle, M.; Min, J.; Shim, S.C.; Kim, G.; Montenegro, V.; Korn, J.H.; Elkon, K.B. Scleroderma fibroblasts demonstrate enhanced activation of Akt (protein kinase B) in situ. J. Investig. Dermatol. 2005, 124, 298–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, H.; Perl, A. Mechanistic target of rapamycin complex 1 expands Th17 and IL-4+ CD4−CD8− double-negative T cells and contracts regulatory T cells in systemic lupus erythematosus. J. Immunol. 2014, 192, 4134–4144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Jiao, Y.; Zhang, X. Immunometabolic Pathways and Its Therapeutic Implication in Autoimmune Diseases. Clin. Rev. Allergy Immunol. 2021, 60, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.C.; Casciola-Rosen, L.; Berger, A.E.; Kapsogeorgou, E.K.; Cheadle, C.; Tzioufas, A.G.; Baer, A.N.; Rosen, A. Precise probes of type II interferon activity define the origin of interferon signatures in target tissues in rheumatic diseases. Proc. Natl. Acad. Sci. USA 2012, 109, 17609–17614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyriakidis, N.C.; Kapsogeorgou, E.K.; Gourzi, V.C.; Konsta, O.D.; Baltatzis, G.E.; Tzioufas, A.G. Toll-like receptor 3 stimulation promotes Ro52/TRIM21 synthesis and nuclear redistribution in salivary gland epithelial cells, partially via type I interferon pathway. Clin. Exp. Immunol. 2014, 178, 548–560. [Google Scholar] [CrossRef] [Green Version]
- Ezell, S.A.; Polytarchou, C.; Hatziapostolou, M.; Guo, A.; Sanidas, I.; Bihani, T.; Comb, M.J.; Sourvinos, G.; Tsichlis, P.N. The protein kinase Akt1 regulates the interferon response through phosphorylation of the transcriptional repressor EMSY. Proc. Natl. Acad. Sci. USA 2012, 109, E613–E621. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; Sassano, A.; Dolniak, B.; Joshi, S.; Majchrzak-Kita, B.; Baker, D.P.; Hay, N.; Fish, E.N.; Platanias, L.C. Role of the Akt pathway in mRNA translation of interferon-stimulated genes. Proc. Natl. Acad. Sci. USA 2008, 105, 4808–4813. [Google Scholar] [CrossRef] [Green Version]
- Larue, L.; Bellacosa, A. Epithelial-mesenchymal transition in development and cancer: Role of phosphatidylinositol 3’ kinase/AKT pathways. Oncogene 2005, 24, 7443–7454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sisto, M.; Lorusso, L.; Ingravallo, G.; Ribatti, D.; Lisi, S. TGFbeta1-Smad canonical and -Erk noncanonical pathways participate in interleukin-17-induced epithelial-mesenchymal transition in Sjogren’s syndrome. Lab. Investig. 2020, 100, 824–836. [Google Scholar] [CrossRef] [PubMed]
- Sisto, M.; Lorusso, L.; Tamma, R.; Ingravallo, G.; Ribatti, D.; Lisi, S. Interleukin-17 and -22 synergy linking inflammation and EMT-dependent fibrosis in Sjogren’s syndrome. Clin. Exp. Immunol. 2019, 198, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, M.; Hisatsune, J.; Yamasaki, E.; Isomoto, H.; Kurazono, H.; Hatakeyama, M.; Azuma, T.; Yamaoka, Y.; Yahiro, K.; Moss, J.; et al. Helicobacter pylori VacA-induced inhibition of GSK3 through the PI3K/Akt signaling pathway. J. Biol. Chem. 2009, 284, 1612–1619. [Google Scholar] [CrossRef] [Green Version]
- Nagy, T.A.; Frey, M.R.; Yan, F.; Israel, D.A.; Polk, D.B.; Peek, R.M., Jr. Helicobacter pylori regulates cellular migration and apoptosis by activation of phosphatidylinositol 3-kinase signaling. J. Infect. Dis. 2009, 199, 641–651. [Google Scholar] [CrossRef] [Green Version]
- Shu, X.; Yang, Z.; Li, Z.H.; Chen, L.; Zhou, X.D.; Xie, Y.; Lu, N.H. Helicobacter pylori Infection Activates the Akt-Mdm2-p53 Signaling Pathway in Gastric Epithelial Cells. Dig. Dis. Sci. 2015, 60, 876–886. [Google Scholar] [CrossRef] [PubMed]
- Tabassam, F.H.; Graham, D.Y.; Yamaoka, Y. Helicobacter pylori-associated regulation of forkhead transcription factors FoxO1/3a in human gastric cells. Helicobacter 2012, 17, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Wen, S.; So, Y.; Singh, K.; Slingerland, J.M.; Resnick, M.B.; Zhang, S.; Ruiz, V.; Moss, S.F. Promotion of cytoplasmic mislocalization of p27 by Helicobacter pylori in gastric cancer. Oncogene 2012, 31, 1771–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Nagy, T.A.; Vilgelm, A.; Zaika, E.; Ogden, S.R.; Romero-Gallo, J.; Piazuelo, M.B.; Correa, P.; Washington, M.K.; El-Rifai, W.; et al. Regulation of p53 tumor suppressor by Helicobacter pylori in gastric epithelial cells. Gastroenterology 2010, 139, 1333–1343. [Google Scholar] [CrossRef] [Green Version]
- Ben Younes, K.; Doghri, R.; Mrad, K.; Bedhiafi, W.; Benammar-Elgaaied, A.; Sola, B.; Ben Aissa-Fennira, F. PTEN Loss and Cyclin A2 Upregulation Define a PI3K/AKT Pathway Activation in Helicobacter pylori-induced MALT and DLBCL Gastric Lymphoma With Features of MALT. Appl. Immunohistochem. Mol. Morphol. 2021, 29, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, U.S.; Nandula, S.R.; Thimmalapura, P.R.; Scindia, Y.M.; Bagavant, H. Activation of innate immune responses through Toll-like receptor 3 causes a rapid loss of salivary gland function. J. Oral Pathol. Med. 2009, 38, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Nandula, S.R.; Dey, P.; Corbin, K.L.; Nunemaker, C.S.; Bagavant, H.; Deshmukh, U.S. Salivary gland hypofunction induced by activation of innate immunity is dependent on type I interferon signaling. J. Oral Pathol. Med. 2013, 42, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Nandula, S.R.; Scindia, Y.M.; Dey, P.; Bagavant, H.; Deshmukh, U.S. Activation of innate immunity accelerates sialoadenitis in a mouse model for Sjogren’s syndrome-like disease. Oral Dis. 2011, 17, 801–807. [Google Scholar] [CrossRef]
- Sander, S.; Calado, D.P.; Srinivasan, L.; Köchert, K.; Zhang, B.; Rosolowski, M.; Rodig, S.J.; Holzmann, K.; Stilgenbauer, S.; Siebert, R.; et al. Synergy between PI3K signaling and MYC in Burkitt lymphomagenesis. Cancer Cell 2012, 22, 167–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuyama, K.; Suzuki, K.; Naruse, T.; Tsuchihashi, H.; Yanamoto, S.; Kaida, A.; Miura, M.; Umeda, M.; Yamashita, S. Prolonged cetuximab treatment promotes p27(Kip1)-mediated G1 arrest and autophagy in head and neck squamous cell carcinoma. Sci. Rep. 2021, 11, 5259. [Google Scholar] [CrossRef]
- Juarez, M.; Diaz, N.; Johnston, G.I.; Nayar, S.; Payne, A.; Helmer, E.; Cain, D.; Williams, P.; Devauchelle-Pensec, V.; Fisher, B.A.; et al. A phase 2 randomized, double-blind, placebo-controlled, proof-of-concept study of oral seletalisib in primary Sjogren’s syndrome. Rheumatology 2021, 60, 1364–1375. [Google Scholar] [CrossRef] [PubMed]
- Shiboski, C.H.; Shiboski, S.C.; Seror, R.; Criswell, L.A.; Labetoulle, M.; Lietman, T.M.; Rasmussen, A.; Scofield, H.; Vitali, C.; Bowman, S.J.; et al. 2016 American College of Rheumatology/European League Against Rheumatism classification criteria for primary Sjogren’s syndrome: A consensus and data-driven methodology involving three international patient cohorts. Ann. Rheum. Dis. 2017, 76, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Fragkioudaki, S.; Mavragani, C.P.; Moutsopoulos, H.M. Predicting the risk for lymphoma development in Sjogren syndrome: An easy tool for clinical use. Medicine 2016, 95, e3766. [Google Scholar] [CrossRef]
- Crowe, A.R.; Yue, W. Semi-quantitative Determination of Protein Expression using Immunohistochemistry Staining and Analysis: An Integrated Protocol. Bio.-Protoc. 2019, 9, e3465. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Features | pSS Patients | |||
|---|---|---|---|---|
| pSS Low-Risk (n = 10) | pSS High-Risk (n = 10) | SSL (n = 9) | ||
| General | Age (years), median (range) | 60 (38–72) | 53.5 (27–71) | 71.5 (58–77) |
| Men/women | 1/10 | 0/10 | 1/9 | |
| Disease duration (years), median (range) | 11.5 (2.0–21.0) | 4.5 (2.0–16.0) | 13.0 (3.0–37.0) | |
| Histological (MSG biopsy) | Biopsy focus score (number of lymphocytic foci/4 mm2), median (range) | 1.67 (1.0–3.27) | 4.57 (2.75–10.44) | 4.44 (1.5–12.0) |
| Germinal center formation, No (%) | 2(20.0) | 4(40.0) | 3(33.3) | |
| Clinical | Arthralgias, No (%) | 6(60.0) | 5(50.0) | 7(77.8) |
| Arthritis, No, (%) | 2(20.0) | 3(30.0) | 2(22.2) | |
| SG enlargement (SGE), No (%) | 0(0.0) | 6(60.0) | 7(77.8) | |
| Raynaud’s phenomenon, No (%) | 3(30.0) | 2(20.0) | 2(22.2) | |
| Lymphadenopathy, No (%) | 1(10.0) | 2(20.0) | 2(22.2) | |
| Parenchymal organ involvement, No (%) | 2(20.0) | 2(20.0) | 1(11.1) | |
| Lung involvement, No (%) | 1(10.0) | 2(20.0) | 1(11.1) | |
| Renal involvement, No (%) | 0(0.0) | 0(0.0) | 0(0.0) | |
| Liver involvement, No (%) | 1(10.0) | 0(0.0) | 0(0.0) | |
| Indicative of vasculitic involvement, No (%) | 0(0.0) | 2(20.0) | 4(44.4) | |
| Palpable purpura, No (%) | 0(0.0) | 1(10.0) | 2(20.0) | |
| Vasculitis, No (%) | 0(0.0) | 2(20.0) | 0(0.0) | |
| Glomerulonephritis, No (%) | 0(0.0) | 0(0.0) | 0(0.0) | |
| Peripheral neuropathy, No (%) | 0(0.0) | 2(20.0) | 2(22.2) | |
| ESSDAI score, median (range) | 3(0–5) | 10.5(5–20) | 17(15–22) | |
| Laboratory | Anti-Ro/SSA and/or La/SSB positive, No (%) | 9(90.0) | 9(90.0) | 7(77.8) |
| Anti-Ro/SSA positive, No (%) | 9(90.0) | 9(90.0) | 7(77.8) | |
| Anti-La/SSB positive, No (%) | 3(30.0) | 8(80.0) | 4(44.4) | |
| Rheumatoid Factor positive, No (%) | 4(40.0) | 8(80.0) | 8(88.9) | |
| C4-hypocomplementemia, No (%) | 0(0.0) | 7(70.0) | 7(77.8) | |
| Cryoglobulinemia, No (%) | 0(0.0) | 3(30.0) | 6(66.7) | |
| Leukopenia, No, (%) | 1(10.0) | 1(10.0) | 1(11.1) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stergiou, I.E.; Chatzis, L.; Papanikolaou, A.; Giannouli, S.; Tzioufas, A.G.; Voulgarelis, M.; Kapsogeorgou, E.K. Akt Signaling Pathway Is Activated in the Minor Salivary Glands of Patients with Primary Sjögren’s Syndrome. Int. J. Mol. Sci. 2021, 22, 13441. https://doi.org/10.3390/ijms222413441
Stergiou IE, Chatzis L, Papanikolaou A, Giannouli S, Tzioufas AG, Voulgarelis M, Kapsogeorgou EK. Akt Signaling Pathway Is Activated in the Minor Salivary Glands of Patients with Primary Sjögren’s Syndrome. International Journal of Molecular Sciences. 2021; 22(24):13441. https://doi.org/10.3390/ijms222413441
Chicago/Turabian StyleStergiou, Ioanna E., Loukas Chatzis, Asimina Papanikolaou, Stavroula Giannouli, Athanasios G. Tzioufas, Michael Voulgarelis, and Efstathia K. Kapsogeorgou. 2021. "Akt Signaling Pathway Is Activated in the Minor Salivary Glands of Patients with Primary Sjögren’s Syndrome" International Journal of Molecular Sciences 22, no. 24: 13441. https://doi.org/10.3390/ijms222413441
APA StyleStergiou, I. E., Chatzis, L., Papanikolaou, A., Giannouli, S., Tzioufas, A. G., Voulgarelis, M., & Kapsogeorgou, E. K. (2021). Akt Signaling Pathway Is Activated in the Minor Salivary Glands of Patients with Primary Sjögren’s Syndrome. International Journal of Molecular Sciences, 22(24), 13441. https://doi.org/10.3390/ijms222413441