
Physiological and Gene Expression Responses of Six Annual Ryegrass Cultivars to Cobalt, Lead, and Nickel Stresses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
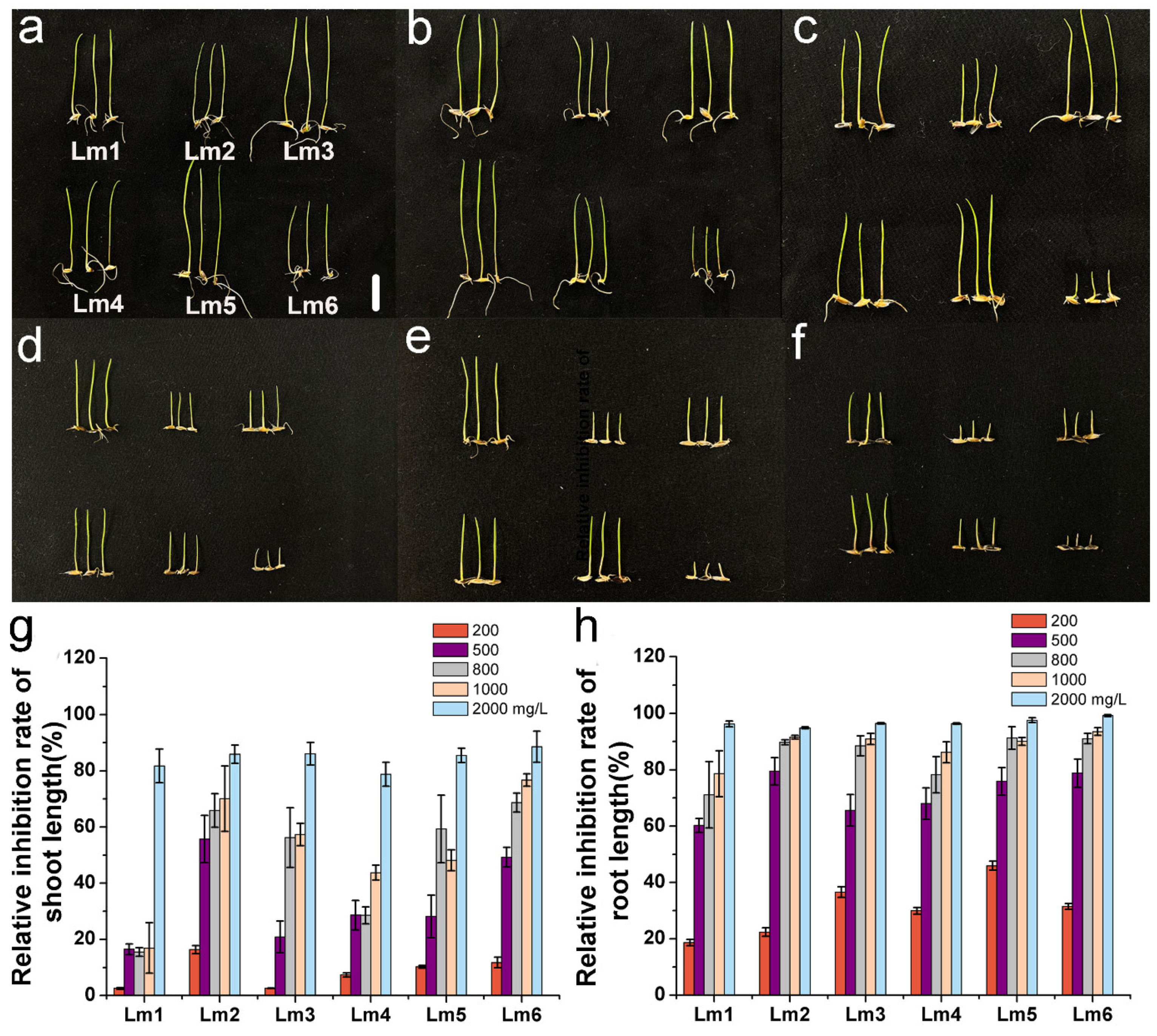
2.1. Effects of Co2+, Pb2+, and Ni2+ on Growth of Six Ryegrass Cultivars
2.2. Effects of Co2+, Pb2+, Ni2+ on Physiological Indexes of Six Ryegrass Cultivars
2.3. Effects of Heavy Metals on the Total Antioxidant Capacity of Six Ryegrass Cultivars
2.4. Toxicity Symptoms in Roots of Six Ryegrass Cultivars Treated with Heavy Metals
2.5. Comprehensive Evaluation of Co2+, Pb2+, and Ni2+ Tolerance of Six Ryegrass Cultivars
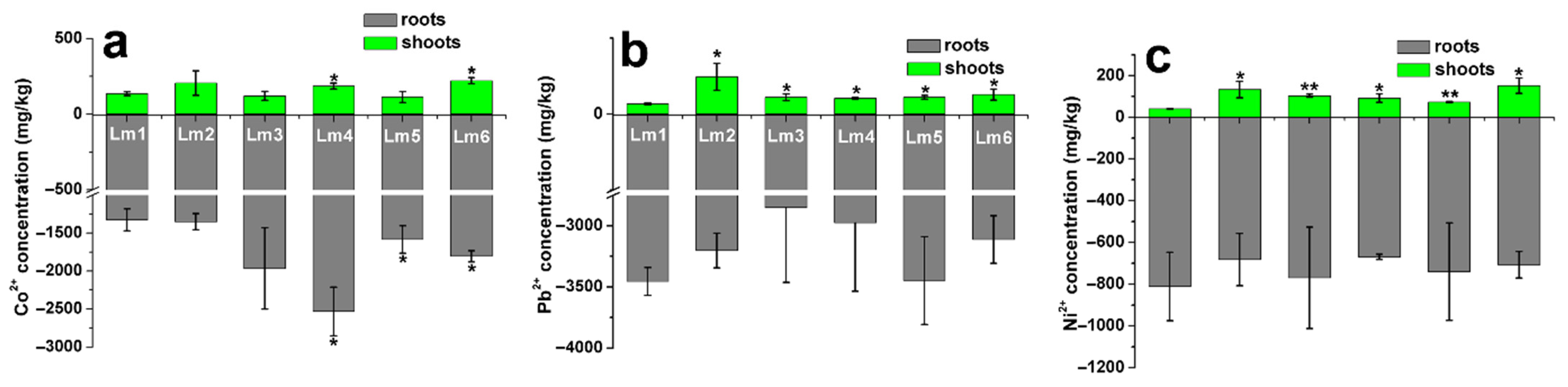
2.6. Heavy Metal Accumulation in Shoots and Roots of Six Ryegrass Cultivars
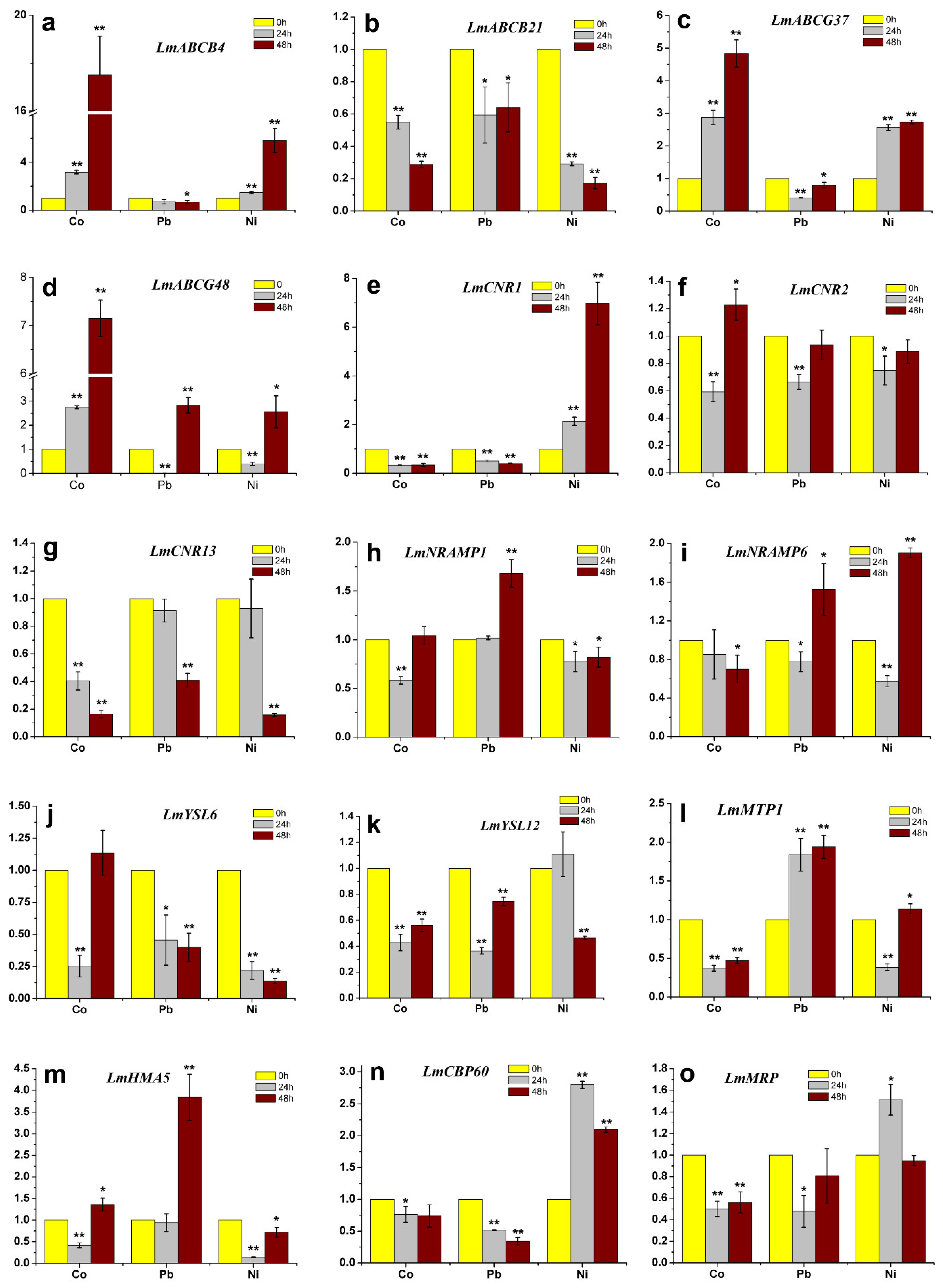
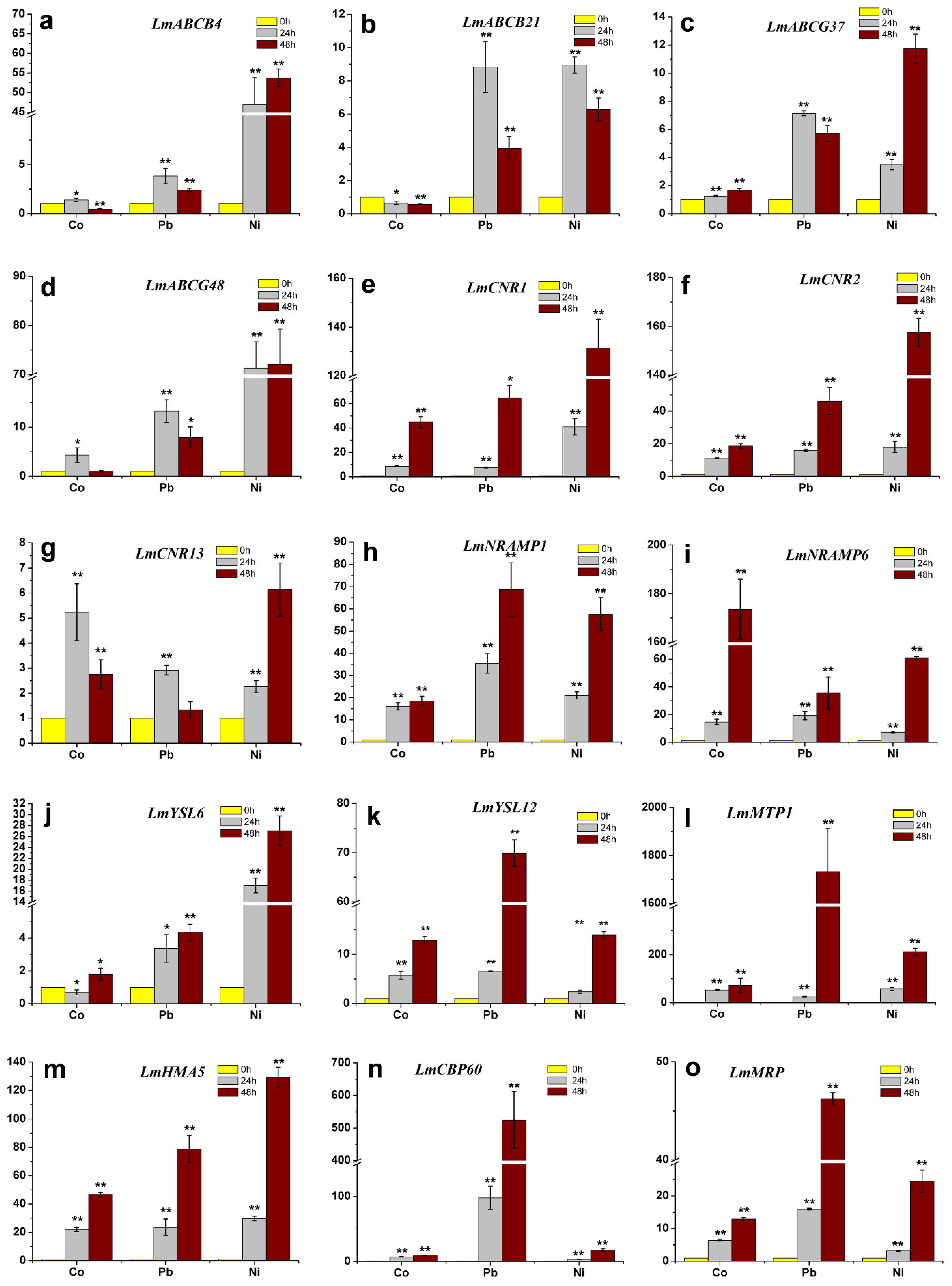
2.7. Changes in Transcript Levels of Genes Encoding Heavy Metal Transporters in Response to Heavy Metal Treatments
3. Discussion
3.1. Effects of Heavy Metals on Seed Germination of Six Ryegrass Cultivars
3.2. Effects of Co2+, Pb2+, and Ni2+ Stresses on Ryegrass Growth
3.3. Effects of Co2+, Pb2+, and Ni2+ Stresses on Total Antioxidant Capacity of Ryegrass
3.4. Accumulation and Translocation of Heavy Metals in Ryegrass Plants
4. Materials and Methods
4.1. Plant Materials and Tolerance Analysis
4.2. Determination of Total Antioxidant Capacity
4.3. Evans Blue Staining Analyses
4.4. Determination of Heavy Metal Concentrations in Seedlings’ Shoots and Roots
4.5. Gene Expression Analyses in Lm1
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, Q.Q.; Li, Z.Y.; Lu, X.N.; Duan, Q.N.; Huang, L.; Bi, J. A review of soil heavy metal pollution from industrial and agricultural regions in China: Pollution and risk assessment. Sci. Total Environ. 2018, 642, 690–700. [Google Scholar] [CrossRef]
- Chonokhuu, S.; Batbold, C.; Chuluunpurev, B.; Battsengel, E.; Dorjsuren, B.; Byambaa, B. Contamination and health risk assessment of heavy metals in the soil of major cities in Mongolia. Int. J. Environ. Res. Public Health 2019, 16, 2552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ai, S.W.; Liu, B.L.; Yang, Y.; Ding, J.; Yang, W.Z.; Bai, X.J.; Naeem, S.; Zhang, Y.M. Temporal variations and spatial distributions of heavy metals in a waste water-irrigated soil-eggplant system and associated influencing factors. Ecotoxicol. Environ. Saf. 2018, 158, 204–214. [Google Scholar] [CrossRef]
- Liu, G.N.; Tao, L.; Liu, X.H.; Hou, J.; Wang, A.J.; Li, R.P. Heavy metal speciation and pollution of agricultural soils along Jishui River in non-ferrous metal mine area in Jiangxi Province. China J. Geochem. Explor. 2013, 132, 156–163. [Google Scholar] [CrossRef]
- Xue, H.Q.; Zhao, L.; Liu, X.D. Characteristics of heavy metal pollution in road runoff in the Nanjing urban area, East China. Water Sci. Technol. 2020, 81, 1961–1971. [Google Scholar] [CrossRef]
- Kosiorek, M.; Wyszkowski, M. Effect of cobalt on environment and living organisms—A review. Appl. Ecol. Environ. Res. 2019, 17, 11419–11449. [Google Scholar] [CrossRef]
- Gupta, V.; Jatav, P.K.; Verma, R.; Kothari, S.L.; Kachhwaha, S. Nickel accumulation and its effect on growth, physiological and biochemical parameters in millets and oats. Environ. Sci. Pollut. Res. 2017, 24, 23915–23925. [Google Scholar] [CrossRef]
- Shahzad, B.; Tanveer, M.; Rehman, A.; Cheema, S.A.; Fahad, S.; Rehman, S.; Sharma, A. Nickel: Whether toxic or essential for plants and environment—A review. Plant Physiol. Biochem. 2018, 132, 641–651. [Google Scholar] [CrossRef]
- Fiala, R.; Repka, V.; Ciamporova, M.; Martinka, M.; Pavlovkin, J. The effect of cadmium-nickel interactions on superoxide production, cell viability and membrane potential (EM) in roots of two maize cultivars. Acta Biol. Hung. 2015, 66, 192–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilmaz, D.D.; Parlak, K.U. Nickel-induced changes in lipid peroxidation, antioxidative enzymes, and metal accumulation in Lemna gibba. Int. J. Phytoremediat. 2011, 13, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Gruenhage, L.; Jaeger, H.J. Effect of heavy metals on growth and heavy metal content of Allium porrum L. and Pisum sativum L. J. Appl. Bot. Angew. Bot. 1985, 59, 11–27. [Google Scholar]
- Kumar, A.; Prasad, M. Plant-lead interactions: Transport, toxicity, tolerance, and detoxification mechanisms. Ecotoxicol. Environ. Saf. 2018, 166, 401–418. [Google Scholar] [CrossRef]
- Illi, J.C.; Vancetta, T.; Alves, D.D.; Osorio, D.M.M.; Bianchin, L.; de Quevedo, D.M.; Juchem, F. Integrated assessment of air pollution by metals and source apportionment using ryegrass (Lolium multiflorum Lam.) in southern Brazil. Environ. Sci. Pollut. Res. 2017, 24, 2790–2803. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.L.; He, X.Y.; Chen, W.; Zhao, M.Z. Ecotoxicological responses of three ornamental herb species to cadmium. Environ. Toxicol. Chem. 2013, 32, 1746–1751. [Google Scholar] [CrossRef]
- Wang, J.G.; Zhao, J.C.; Feng, S.; Zhang, J.Z.; Gong, S.F.; Qiao, K.; Zhou, A.M. Comparison of cadmium uptake and transcriptional responses in roots reveal key transcripts from high and low-cadmium tolerance ryegrass cultivars. Ecotoxicol. Environ. Saf. 2020, 203, 110961. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.S.; Xu, Y.C.; Gui, H.P.; Zhang, H.H.; Dong, Q.; Sikder, R.K.; Wang, X.R.; Yang, G.Z.; Song, M.Z. Evaluation of cotton (Gossypium hirsutum L.) leaf abscission sensitivity triggered by thidiazuron through membership function value. Plants 2021, 10, 49. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.J.; Song, S.H.; Wang, W.B.; Wang, C.L.; Li, H.B.; Wang, F.; Li, S.Y.; Sun, X.G. Screening diverse soybean genotypes for drought tolerance by membership function value based on multiple traits and drought tolerant coefficient of yield. BMC Plant Biol. 2020, 20, 321. [Google Scholar] [CrossRef]
- Zhang, H.; Kou, J.T.; Shi, S.L. Physiological and biochemical responses of Medicago sativa seed to cobalt stress. Acta Pratacul. Sin. 2015, 24, 146–153. [Google Scholar]
- Mathad, P.; Pratima, H. Copper toxicity causes oxidative stress in Brassica juncea L. seedings. Indian J. Plant Physiol. 2009, 14, 397–401. [Google Scholar]
- Kumar, M.; Jayaraman, P. Toxic Effect of Lead Nitrate [Pb(NO3)2] on the Black Gram Seedlings (Vigna mungo (L.) Hepper). Int. J. Adv. Res. Biol. Sci. 2014, 1, 209–213. [Google Scholar]
- Sidhu, G.P.S.; Bali, A.S.; Bhardwaj, R.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Bioaccumulation and physiological responses to lead (Pb) in Chenopodium murale L. Ecotoxicol. Environ. Saf. 2018, 151, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Imtiaz, M.; Dai, Z.H.; Mehmood, S.; Adeel, M.; Liu, J.C.; Tu, S.X. Nickel stressed responses of rice in Ni subcellular distribution, antioxidant production, and osmolyte accumulation. Environ. Sci. Pollut. Res. 2017, 24, 20587–20598. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, X.Y.; Liu, Y.Y.; Wei, J.Y.; Shen, W.B.; Shen, Z.G.; Cui, J. Hemin-mediated alleviation of zinc, lead and chromium toxicity is associated with elevated photosynthesis, antioxidative capacity; suppressed metal uptake and oxidative stress in rice seedlings. Plant Growth Regul. 2017, 81, 253–264. [Google Scholar] [CrossRef]
- Duman, F.; Ozturk, F. Nickel accumulation and its effect on biomass, protein content and antioxidative enzymes in roots and leaves of watercress. J. Environ. Sci. 2010, 22, 526–532. [Google Scholar] [CrossRef]
- Georgiadou, E.C.; Ewa, K.; Katarzyna, P.; Katarzyna, P.; Kamila, K.; Beata, S.; Joanna, L.; Vasileios, F. Influence of heavy metals (Ni, Cu, and Zn) on nitro-oxidative stress responses, proteome regulation and allergen production in basil (Ocimum basilicum L.) plants. Front. Plant Sci. 2018, 9, 862. [Google Scholar] [CrossRef] [Green Version]
- Abolghassem, E.; Ding, Y.; Mokhberdoran, F.; Xie, Y.F. Heavy metal stress and some mechanisms of plant defense response. Sci. World J. 2015, 2015, 756120. [Google Scholar]
- Fu, S.; Lu, Y.S.; Zhang, X.; Yang, G.Z.; Chao, D.; Wang, Z.G.; Shi, M.X.; Chen, J.G.; Chao, D.Y.; Li, R.B.; et al. The ABC transporter ABCG36 is required for cadmium tolerance in rice. J. Exp. Bot. 2019, 70, 5909–5918. [Google Scholar] [CrossRef]
- Ruzicka, K.; Strader, L.C.; Bailly, A.; Yang, H.B.; Blakeslee, J.; Langowski, L.; Nejedla, E.; Fujita, H.; Itoh, H.; Syono, K.; et al. Arabidopsis PIS1 encodes the ABCG37 transporter of auxinic compounds including the auxin precursor indole-3-butyric acid. Proc. Natl. Acad. Sci. USA 2010, 107, 10749–10753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamimoto, Y.; Terasaka, K.; Hamamoto, M.; Takanashi, K.; Fukuda, S.; Shitan, N.; Sugiyama, A.; Suzuki, H.; Shibata, D.; Wang, B.; et al. Arabidopsis ABCB21 is a facultative auxin importer/exporter regulated by cytoplasmic auxin concentration. Plant Cell Physiol. 2012, 53, 2090–2100. [Google Scholar] [CrossRef] [Green Version]
- Deng, F.L.; Yamaji, N.; Xia, J.X.; Ma, J.F. A member of the heavy metal P-type ATPase OsHMA5 is involved in xylem loading of copper in rice. Plant Physiol. 2013, 163, 1353–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.F.; Aarts, M.G.M. The molecular mechanism of zinc and cadmium stress response in plants. Cell. Mol. Life Sci. 2012, 69, 3187–3206. [Google Scholar] [CrossRef] [PubMed]
- Li, D.D.; Xu, X.M.; Hu, X.Q.; Liu, Q.G.; Wang, Z.C.; Zhang, H.Z.; Wang, H.; Wei, M.; Wang, H.Z.; Liu, H.M.; et al. Genome-wide analysis and heavy metal-induced expression profiling of the HMA gene family in Populus trichocarpa. Front. Plant Sci. 2015, 6, 1149. [Google Scholar] [CrossRef]
- Chang, P.; Yin, H.; Imanaka, T.; Igarashi, Y.; Li, N.N.; Luo, F. The metal transporter CrNRAMP1 is involved in zinc and cobalt transports in Chlamydomonas reinhardtii. Biochem. Biophys. Res. Commun. 2020, 523, 880–886. [Google Scholar] [CrossRef]
- Shi, W.; Zhou, J.; Li, J.; Ma, C.F.; Zhang, Y.H.; Deng, S.R.; Yu, W. Lead exposure-induced defense responses result in low lead translocation from the roots to aerial tissues of two contrasting poplar species. Environ. Pollut. 2020, 271, 116346. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, T.; Usui, K.; Horie, K.; Nosaka, S.; Mizuno, N.; Obata, H. Cloning of three ZIP/Nramp transporter genes from a Ni hyperaccumulator plant Thlaspi japonicum and their Ni2+-transport abilities. Plant Physiol. Biochem. 2005, 43, 793–801. [Google Scholar] [CrossRef]
- Tian, E.J.; He, G.D.; Qin, L.J.; Li, D.D.; Meng, L.L.; Huang, Y.; He, T.B. Genome-wide analysis of the NRAMP gene family in potato (Solanum tuberosum): Identification, expression analysis and response to five heavy metals stress. Ecotoxicol. Environ. Saf. 2021, 208, 111661. [Google Scholar] [CrossRef]
- Montanini, B.; Blaudez, D.; Jeandroz, S.; Sanders, D.; Chalot, M. Phylogenetic and functional analysis of the Cation Diffusion Facilitator (CDF) family: Improved signature and prediction of substrate specificity. BMC Genom. 2007, 8, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.H.; Qiao, K.; Liang, S.; Tian, S.Q.; Tian, Y.B.; Wang, H.; Chai, T.Y. Triticum urartu MTP1: Its ability to maintain Zn2+ and Co2+ homeostasis and metal selectivity determinants. Plant Cell Rep. 2018, 37, 1653–1666. [Google Scholar] [CrossRef]
- Jaquinod, M.; Villiers, F.; Kieffer-Jaquinod, S.; Hugouvieu, V.; Bruley, C.; Garin, J.; Bourguignon, J. A proteomics dissection of Arabidopsis thaliana vacuoles isolated from cell culture. Mol. Cell. Proteom. 2007, 6, 394–412. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.W.; Li, Y.; Zhang, Y.X.; Chai, T.Y. Molecular cloning and characterization of a Brassica juncea yellow stripe-like gene, BjYSL7, whose overexpression increases heavy metal tolerance of tobacco. Plant Cell Rep. 2013, 32, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Qiao, K.; Tian, Y.B.; Hu, Z.L.; Chai, T.Y. Wheat cell number regulator CNR10 enhances the tolerance, translocation, and accumulation of heavy metals in plants. Environ. Sci. Technol. 2019, 53, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Qiao, K.; Wang, F.H.; Liang, S.; Wang, H.; Hu, Z.L.; Chai, T.Y. New biofortification tool: Wheat TaCNR5 enhances zinc and manganese tolerance and increases zinc and manganese accumulation in rice grains. J. Agric. Food Chem. 2019, 67, 9877–9884. [Google Scholar] [CrossRef]
- Wan, D.L.; Li, R.L.; Zou, B.; Zhang, X.; Cong, J.Y.; Wang, R.G.; Xia, Y.J. Calmodulin-binding protein CBP60g is a positive regulator of both disease resistance and drought tolerance in Arabidopsis. Plant Cell Rep. 2012, 31, 1269–1281. [Google Scholar] [CrossRef] [PubMed]
- Sunkar, R.; Kaplan, B.; Bouche, N.; Arazi, T.; Dolev, D.; Talke, I.N.; Maathuis, F.J.M.; Sanders, D.; Bouchez, D.; Fromm, H. Expression of a truncated tobacco NtCBP4 channel in transgenic plants and disruption of the homologous Arabidopsis CNGC1 gene confer Pb2+ tolerance. Plant J. 2000, 24, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Kolukisaoglu, H.U.; Bovet, L.; Klein, M.; Eggmann, T.; Geisler, M.; Wanke, D.; Martinoia, E.; Schulz, B. Family business: The multidrug-resistance related protein (MRP) ABC transporter genes in Arabidopsis thaliana. Planta 2002, 216, 107–119. [Google Scholar] [CrossRef]
- Zientaraa, K.; Wawrzynska, A.; Łukomskaa, J.; López-Moya, J.R.; Liszewska, F.; Assuncão, A.G.L.; Aarts, M.G.M.; Sirko, A. Activity of the AtMRP3 promoter in transgenic Arabidopsis thaliana and Nicotiana tabacum plants is increased by cadmium, nickel, arsenic, cobalt and lead but not by zinc and iron. J. Biotechnol. 2009, 139, 258–263. [Google Scholar] [CrossRef]
- Cui, E.P.; Cui, B.J.; Fan, X.Y.; Li, S.J.; Gao, F. Ryegrass (Lolium multiflflorum L.) and Indian mustard (Brassica juncea L.) intercropping can improve the phytoremediation of antibiotics and antibiotic resistance genes but not heavy metals. Sci. Total Environ. 2021, 784, 147093. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiao, S.; Tao, Y.; Shan, Q.; Wang, J.; Chai, T.; Gong, S.; Qiao, K. Physiological and Gene Expression Responses of Six Annual Ryegrass Cultivars to Cobalt, Lead, and Nickel Stresses. Int. J. Mol. Sci. 2021, 22, 13583. https://doi.org/10.3390/ijms222413583
Qiao S, Tao Y, Shan Q, Wang J, Chai T, Gong S, Qiao K. Physiological and Gene Expression Responses of Six Annual Ryegrass Cultivars to Cobalt, Lead, and Nickel Stresses. International Journal of Molecular Sciences. 2021; 22(24):13583. https://doi.org/10.3390/ijms222413583
Chicago/Turabian StyleQiao, Siyu, Ye Tao, Qinghua Shan, Jingang Wang, Tuanyao Chai, Shufang Gong, and Kun Qiao. 2021. "Physiological and Gene Expression Responses of Six Annual Ryegrass Cultivars to Cobalt, Lead, and Nickel Stresses" International Journal of Molecular Sciences 22, no. 24: 13583. https://doi.org/10.3390/ijms222413583
APA StyleQiao, S., Tao, Y., Shan, Q., Wang, J., Chai, T., Gong, S., & Qiao, K. (2021). Physiological and Gene Expression Responses of Six Annual Ryegrass Cultivars to Cobalt, Lead, and Nickel Stresses. International Journal of Molecular Sciences, 22(24), 13583. https://doi.org/10.3390/ijms222413583