
Gliadin, through the Activation of Innate Immunity, Triggers lncRNA NEAT1 Expression in Celiac Disease Duodenal Mucosa

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Long Non-Coding RNA Expression Varies in CD Mucosa
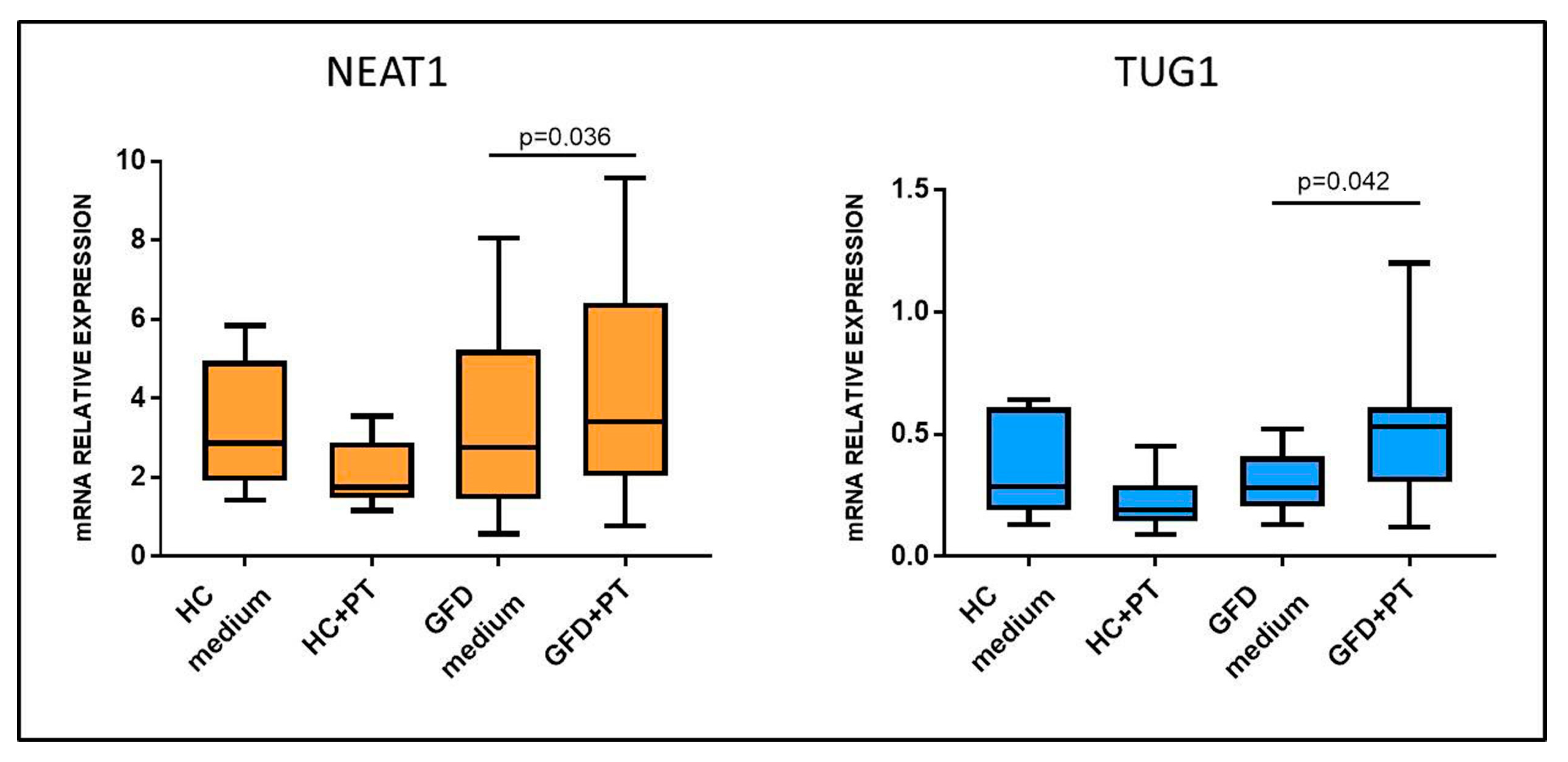
2.2. Gluten Exposure Alters NEAT1 and TUG1 Expression
2.3. NEAT1 and TUG1 Are Preferentially Expressed in the Epithelium
2.4. NEAT1 and TUG1 Expression Is Triggered by Cytokines
2.5. IL-15 Causes the Binding of STAT3 to NEAT1 Promoter in HIEC-6 Cell Model
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Biopsy Processing and Ex Vivo Stimulation
4.3. Laser Capture Microdissection
4.4. Cell Culture
4.5. RNA Extraction and Gene Expression
| HPRT | F: TGAAAAGGACCCCACGAAGT, R: TTGAACTCTCATCTTAGGCTT; |
| NEAT1 | F:CTTCCTCCCTTTAACTTATCCATTCAC, R: CTCTTCCTCCACCATTACCAACAATAC; |
| TUG1 | F:ACGACTGAGCAAGCACTACC, R: CTCAGCAATCAGGAGGCACA; |
| IL-8 | F: GGAAGGAACCATCTCACTGT, R: CCACTCTCAATCACTCTCAG; |
| IL-15 | F: TTCACTTGAGTCCGGAGATGC; R: CCTCCAGTTCCTCACATTCTTTG; |
| IFNγ | F: AAGAGTGTGGAGACCATCAAGG; R: ACTCCTTTTTCGCTTCCCTGT. |
4.6. Protein Extraction and Western Blot
4.7. Promoter Region Analysis
4.8. DNA Retardation Assay
- F:GGTGTTAACCAGGGAGAGGTTCCTGGCAGGAGTTCCTGTCAGATGCCATTTTCCATTCTG;
- R:CAGAATGGAAAATGGCATCTGACAGGAACTCCTGCCAGGAACCTCTCCCTGGTTAACACC;
- F: AGGGAGAGGTTCCTGGCAGG;
- R: CCTGCCAGGAACCTCTCCCT.
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abadie, V.; Jabri, B. IL-15: A Central Regulator of Celiac Disease Immunopathology. Immunol. Rev. 2014, 260, 221–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebwohl, B.; Sanders, D.S.; Green, P.H.R. Coeliac Disease. Lancet 2018, 391, 70–81. [Google Scholar] [CrossRef]
- Buoli Comani, G.; Panceri, R.; Dinelli, M.; Biondi, A.; Mancuso, C.; Meneveri, R.; Barisani, D. MiRNA-Regulated Gene Expression Differs in Celiac Disease Patients According to the Age of Presentation. Genes Nutr. 2015, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaira, V.; Roncoroni, L.; Barisani, D.; Gaudioso, G.; Bosari, S.; Bulfamante, G.; Doneda, L.; Conte, D.; Tomba, C.; Bardella, M.T.; et al. MicroRNA Profiles in Coeliac Patients Distinguish Different Clinical Phenotypes and Are Modulated by Gliadin Peptides in Primary Duodenal Fibroblasts. Clin. Sci. 2013, 126, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, L.; Ding, Y.; Lu, X.; Zhang, G.; Yang, J.; Zheng, H.; Wang, H.; Jiang, Y.; Xu, L. LncRNA Structural Characteristics in Epigenetic Regulation. Int. J. Mol. Sci. 2017, 18, 2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Bajic, V.B.; Zhang, Z. On the Classification of Long Non-Coding RNAs. RNA Biol. 2013, 10, 1–10. [Google Scholar] [CrossRef]
- Chi, Y.; Wang, D.; Wang, J.; Yu, W.; Yang, J. Long Non-Coding RNA in the Pathogenesis of Cancers. Cells 2019, 8, 1015. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Ling, A.; Pareek, S.; Huang, R.S. Oncogene or Tumor Suppressor? Long Noncoding RNAs Role in Patient’s Prognosis Varies Depending on Disease Type. Transl. Res. 2020, in press. [Google Scholar] [CrossRef]
- Wu, G.C.; Pan, H.F.; Leng, R.X.; Wang, D.G.; Li, X.P.; Li, X.M.; Ye, D.Q. Emerging Role of Long Noncoding RNAs in Autoimmune Diseases. Autoimmun. Rev. 2015, 14, 798–805. [Google Scholar] [CrossRef]
- Li, S.; Gao, Y.; Zhang, Z.; Yu, X.; Zheng, J. The Role of Long Non-Coding RNAs in the Pathogenesis of RA, SLE, and SS. Front. Med. 2018, 5, 1–14. [Google Scholar] [CrossRef]
- Chen, S.W.; Wang, P.Y.; Liu, Y.C.; Sun, L.; Zhu, J.; Zuo, S.; Ma, J.; Li, T.Y.; Zhang, J.L.; Chen, G.W.; et al. Effect of Long Noncoding RNA H19 Overexpression on Intestinal Barrier Function and Its Potential Role in the Pathogenesis of Ulcerative Colitis. Inflamm. Bowel Dis. 2016, 22, 2582–2592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, T.X.; Chung, H.K.; Xiao, L.; Piao, J.J.; Lan, S.; Jaladanki, S.K.; Turner, D.J.; Raufman, J.P.; Gorospe, M.; Wang, J.Y. Long Noncoding RNA H19 Impairs the Intestinal Barrier by Suppressing Autophagy and Lowering Paneth and Goblet Cell Function. Cell. Mol. Gastronenterol. Hepatol. 2020, 9, 611–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellanos-Rubio, A.; Fernandez-Jimenez, N.; Kratchmarov, R.; Luo, X.; Bhagat, G.; Green, P.H.; Schneider, R.; Kiledjian, M.; Bilbao, G.S., Jr. A Long Noncoding RNA Associated with Susceptibility to Celiac Disease. Science 2016, 352, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Santin, I.; Jauregi-Miguel, A.; Velayos, T.; Castellanos-Rubio, A.; Garcia-Etxebarria, K.; Romero-Garmendia, I.; Fernandez-Jimenez, N.; Irastorza, I.; Castaño, L.; Bilbao, J. Celiac Disease Associates LncRNA Named HCG14. J. Pediatr. Gastroenterol. Nutr. 2018, 67, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Deng, W.; Zhang, W. Long Non-Coding RNA TUG1 Protects Renal Tubular Epithelial Cells against Injury Induced by Lipopolysaccharide via Regulating MicroRNA-223. Biomed. Pharmacother. 2018, 104, 509–519. [Google Scholar] [CrossRef]
- Shen, N.; Zhang, F.; Zhu, W.; Xia, S.; Tang, Y.; Huang, J.; Yang, W.; Wu, Y.; La, T.; Yao, Y.; et al. Identification of the Long Noncoding RNA NEAT1 as a Novel Inflammatory Regulator Acting through MAPK Pathway in Human Lupus. J. Autoimmun. 2016, 75, 96–104. [Google Scholar] [CrossRef]
- Santoro, M.; Nociti, V.; Lucchini, M.; De Fino, C.; Losavio, F.A.; Mirabella, M. Expression Profile of Long Non-Coding RNAs in Serum of Patients with Multiple Sclerosis. J. Mol. Neurosci. 2016, 59, 18–23. [Google Scholar] [CrossRef]
- Shui, X.; Chen, S.; Lin, J.; Kong, J.; Zhou, C.; Wu, J. Knockdown of LncRNA NEAT1 Inhibits Th17/CD4+ T Cell Differentiation through Reducing the STAT3 Protein Level. J. Cell. Physiol. 2019, 234, 22477–22484. [Google Scholar] [CrossRef]
- Lo, P.K.; Wolfson, B.; Zhou, Q. Cellular, Physiological and Pathological Aspects of the Long Non-Coding RNA NEAT1. Front. Biol. 2016, 11, 413–426. [Google Scholar] [CrossRef] [Green Version]
- Imamura, K.; Imamachi, N.; Akizuki, G.; Kumakura, M.; Kawaguchi, A.; Nagata, K.; Kato, A.; Kawaguchi, Y.; Sato, H.; Yoneda, M.; et al. Long Noncoding RNA NEAT1-Dependent SFPQ Relocation from Promoter Region to Paraspeckle Mediates IL8 Expression upon Immune Stimuli. Mol. Cell 2014, 53, 393–406. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Li, Y.; Zhang, B.; Liu, H.; Wu, M.; Zhang, X. LncRNA TUG1 Regulates Ulcerative Colitis through MiR-142-5p/SOCS1 Axis. Microb. Pathog. 2020, 143, 104139. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liao, S.; Li, H.; Chen, Y.; Yu, J. Long Non-Coding RNA TUG1 Sponges Mir-145a-5p to Regulate Microglial Polarization After Oxygen-Glucose Deprivation. Front. Mol. Neurosci. 2019, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Carlevaro-Fita, J.; Liu, L.; Zhou, Y.; Zhang, S.; Chouvardas, P.; Johnson, R.; Li, J. LnCompare: Gene Set Feature Analysis for Human Long Non-Coding RNAs. Nucleic Acids Res. 2019, 47, W523–W529. [Google Scholar] [CrossRef] [PubMed]
- Johnston, J.A.; Bacon, C.M.; Finbloom, D.S.; Rees, R.C.; Kaplan, D.; Shibuya, K.; Ortaldo, J.R.; Gupta, S.; Chen, Y.Q.; Giri, J.D.; et al. Tyrosine Phosphorylation and Activation of STAT5, STAT3, and Janus Kinases by Interleukins 2 and 15. Proc. Natl. Acad. Sci. USA 1995, 92, 8705–8709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waugh, D.J.J.; Wilson, C. The Interleukin-8 Pathway in Cancer. Clin. Cancer Res. 2008, 14, 6735–6741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro, F.; Cardoso, A.P.; Gonçalves, R.M.; Serre, K.; Oliveira, M.J. Interferon-Gamma at the Crossroads of Tumor Immune Surveillance or Evasion. Front. Immunol. 2018, 9, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Perreault, N.; Beaulieu, J.-F. Use of the Dissociating Enzyme Thermolysin to Generate Viable Human Normal Intestinal Epithelial Cell Cultures. Exp. Cell Res. 1996, 224, 354–364. [Google Scholar] [CrossRef]
- Kumar, V.; Wijmenga, C.; Withoff, S. From Genome-Wide Association Studies to Disease Mechanisms: Celiac Disease as a Model for Autoimmune Diseases. Semin. Immunopathol. 2012, 34, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Plaza-Izurieta, L.; Fernandez-Jimenez, N.; Irastorza, I.; Jauregi-Miguel, A.; Romero-Garmendia, I.; Vitoria, J.C.; Bilbao, J.R. Expression Analysis in Intestinal Mucosa Reveals Complex Relations among Genes under the Association Peaks in Celiac Disease. Eur. J. Hum. Genet. 2015, 23, 1100–1105. [Google Scholar] [CrossRef] [Green Version]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. LncRNA: Insights Into Functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef]
- Xiyuan, L.; Dechao, B.; Liang, S.; Yang, W.; Shuangsang, F.; Hui, L.; Haitao, L.; Chunlong, L.; Wenzheng, F.; Runsheng, C.; et al. Using the NONCODE Database Resource. Curr. Protoc. Bioinform. 2017, 58, 12.16.1–12.16.19. [Google Scholar] [CrossRef] [PubMed]
- Frankish, A.; Diekhans, M.; Ferreira, A.M.; Johnson, R.; Jungreis, I.; Loveland, J.; Mudge, J.M.; Sisu, C.; Wright, J.; Armstrong, J.; et al. GENCODE Reference Annotation for the Human and Mouse Genomes. Nucleic Acids Res. 2019, 47, D766–D773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Liu, S.; Hu, X. Long Non-Coding RNAs: Crucial Regulators of Gastrointestinal Cancer Cell Proliferation. Cell Death Discov. 2018, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, E.K.; Covarrubias, S.; Carpenter, S. The How and Why of LncRNA Function: An Innate Immune Perspective. BBA 2020, 1863, 194419. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ao, L.; Yang, J. Long Non-Coding RNAs in Diseases Related to Inflammation and Immunity. Ann. Transl. Med. 2019, 7, 494. [Google Scholar] [CrossRef]
- Yarani, R.; Mirza, A.H.; Kaur, S.; Pociot, F. The Emerging Role of LncRNAs in Inflammatory Bowel Disease. Exp. Mol. Med. 2018, 50, 161. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Qi, Y.; Qu, J.; Gai, L.; Shi, Y.; Yuan, C. Pathophysiological Functions of the LncRNA TUG1. Curr. Pharm. Des. 2019, 26, 688–700. [Google Scholar] [CrossRef]
- Qiang, Z.; Ma, H.; Cao, X.; Chen, F.; Fang, B.; Chang, Y.; Jia, H.; Li, Z. Downregulation of LncRNA TUG1 Inhibited TLR4 Signaling Pathway-Mediated Inflammatory Damage After Spinal Cord Ischemia Reperfusion in Rats via Suppressing TRIL Expression. J. Neuropathol. Exp. Neurol. 2019, 78, 268–282. [Google Scholar] [CrossRef]
- Fuschi, P.; Carrara, M.; Voellenkle, C.; Garcia-Manteiga, J.M.; Righini, P.; Maimone, B.; Sangalli, E.; Villa, F.; Specchia, C.; Picozza, M.; et al. Central Role of the P53 Pathway in the Noncoding-RNA Response to Oxidative Stress. Aging 2017, 9, 2559–2586. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Wang, S.; Hong, L.; Yu, S.; Li, B.; Zeng, H.; Yang, X.; Zhang, P.; Shao, L. Long Noncoding RNA Taurine-Upregulated Gene 1 Knockdown Protects Cardiomyocytes Against Hypoxia/Reoxygenation-Induced Injury Through Regulating MiR-532-5p/Sox8 Axis. J. Cardiovasc. Pharmacol. 2020, 76, 556–563. [Google Scholar] [CrossRef]
- Yang, B.; Liang, R.-S.; Wu, X.-Y.; Lin, Y.-J. LncRNA TUG1 Inhibits Neuronal Apoptosis in Status Epilepticus Rats via Targeting the MiR-421/MTOR Axis. Cell. Signal. 2020, 76, 109787. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Cai, J.; Wang, Q.; Wang, Y.; Liu, M.; Yang, J.; Zhou, J.; Kang, C.; Li, M.; Jiang, C. Long Noncoding RNA NEAT1, Regulated by the EGFR Pathway, Contributes to Glioblastoma Progression through the WNT/b-Catenin Pathway by Scaffolding EZH2. Clin. Cancer Res. 2018, 24, 684–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.J.; Lin, Q.; Ma, J.X.; Shi, W.W.; Yang, B.; Li, F. Long Non-Coding RNA NEAT1 Acts as Oncogene in NSCLC by Regulating the Wnt Signaling Pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 504–510. [Google Scholar]
- Xiao, C.H.; Yu, H.Z.; Guo, C.Y.; Wu, Z.M.; Cao, H.Y.; Li, W.B.; Yuan, J.F. Long Non-Coding RNA TUG1 Promotes the Proliferation of Colorectal Cancer Cells through Regulating Wnt/β-Catenin Pathway. Oncol. Lett. 2018, 16, 5317–5324. [Google Scholar] [CrossRef] [PubMed]
- Dotsenko, V.; Oittinen, M.; Taavela, J.; Popp, A.; Peräaho, M.; Staff, S.; Sarin, J.; Leon, F.; Isola, J.; Mäki, M.; et al. Genome-Wide Transcriptomic Analysis of Intestinal Mucosa in Celiac Disease Patients on a Gluten-Free Diet and Postgluten Challenge. Cell. Mol. Gatroenterol. Hepatol. 2021, 11, 13–32. [Google Scholar] [CrossRef]
- Magni, S.; Comani, G.B.; Elli, L.; Vanessi, S.; Ballarini, E.; Nicolini, G.; Rusconi, M.; Castoldi, M.; Meneveri, R.; Muckenthaler, M.U.; et al. MIRNAs Affect the Expression of Innate and Adaptive Immunity Proteins in Celiac Disease. Am. J. Gastroenterol. 2014, 109, 1662. [Google Scholar] [CrossRef]
- Frazer, A.C.; Fletcher, R.F.; Ross, C.A.; Shaw, B.; Sammons, H.G.; Schneider, R. Gluten-Induced Enteropathy the Effect of Partially Digested Gluten. Lancet 1959, 274, 252–255. [Google Scholar] [CrossRef]
- Yates, A.D.; Achuthan, P.; Akanni, W.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; et al. Ensembl 2020. Nucleic Acids Res. 2020, 48, D682–D688. [Google Scholar] [CrossRef]
- Cartharius, K.; Frech, K.; Grote, K.; Klocke, B.; Haltmeier, M.; Klingenhoff, A.; Frisch, M.; Bayerlein, M.; Werner, T. MatInspector and beyond: Promoter Analysis Based on Transcription Factor Binding Sites. Bioinformatics 2005, 21, 2933–2942. [Google Scholar] [CrossRef] [Green Version]
- Messeguer, X.; Escudero, R.; Farré, D.; Núñez, O.; Martínez, J.; Albà, M.M. PROMO: Detection of Known Transcription Regulatory Elements Using Species-Tailored Searches. Bioinformatics 2002, 18, 333–334. [Google Scholar] [CrossRef]
- Fornes, O.; Castro-Mondragon, J.A.; Khan, A.; Van Der Lee, R.; Zhang, X.; Richmond, P.A.; Modi, B.P.; Correard, S.; Gheorghe, M.; Baranašić, D.; et al. JASPAR 2020: Update of the Open-Access Database of Transcription Factor Binding Profiles. Nucleic Acids Res. 2020, 48, D87–D92. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Yang, B.; Huang, D.; Wang, D.; Tian, J.; Chen, F.; Wang, X. STAT3-Induced up-Regulation of LncRNA NEAT1 as a CeRNA Facilitates Abdominal Aortic Aneurysm Formation by Elevating TULP3. Biosci. Rep. 2020, 40, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gnodi, E.; Mancuso, C.; Elli, L.; Ballarini, E.; Meneveri, R.; Beaulieu, J.F.; Barisani, D. Gliadin, through the Activation of Innate Immunity, Triggers lncRNA NEAT1 Expression in Celiac Disease Duodenal Mucosa. Int. J. Mol. Sci. 2021, 22, 1289. https://doi.org/10.3390/ijms22031289
Gnodi E, Mancuso C, Elli L, Ballarini E, Meneveri R, Beaulieu JF, Barisani D. Gliadin, through the Activation of Innate Immunity, Triggers lncRNA NEAT1 Expression in Celiac Disease Duodenal Mucosa. International Journal of Molecular Sciences. 2021; 22(3):1289. https://doi.org/10.3390/ijms22031289
Chicago/Turabian StyleGnodi, Elisa, Clara Mancuso, Luca Elli, Elisa Ballarini, Raffaella Meneveri, Jean François Beaulieu, and Donatella Barisani. 2021. "Gliadin, through the Activation of Innate Immunity, Triggers lncRNA NEAT1 Expression in Celiac Disease Duodenal Mucosa" International Journal of Molecular Sciences 22, no. 3: 1289. https://doi.org/10.3390/ijms22031289
APA StyleGnodi, E., Mancuso, C., Elli, L., Ballarini, E., Meneveri, R., Beaulieu, J. F., & Barisani, D. (2021). Gliadin, through the Activation of Innate Immunity, Triggers lncRNA NEAT1 Expression in Celiac Disease Duodenal Mucosa. International Journal of Molecular Sciences, 22(3), 1289. https://doi.org/10.3390/ijms22031289