
Molecular Characterization of Paralichthys olivaceus MAF1 and Its Potential Role as an Anti-Viral Hemorrhagic Septicaemia Virus Factor in Hirame Natural Embryo Cells

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Isolation and Characterization of Maf1 Gene from P. olivaceus
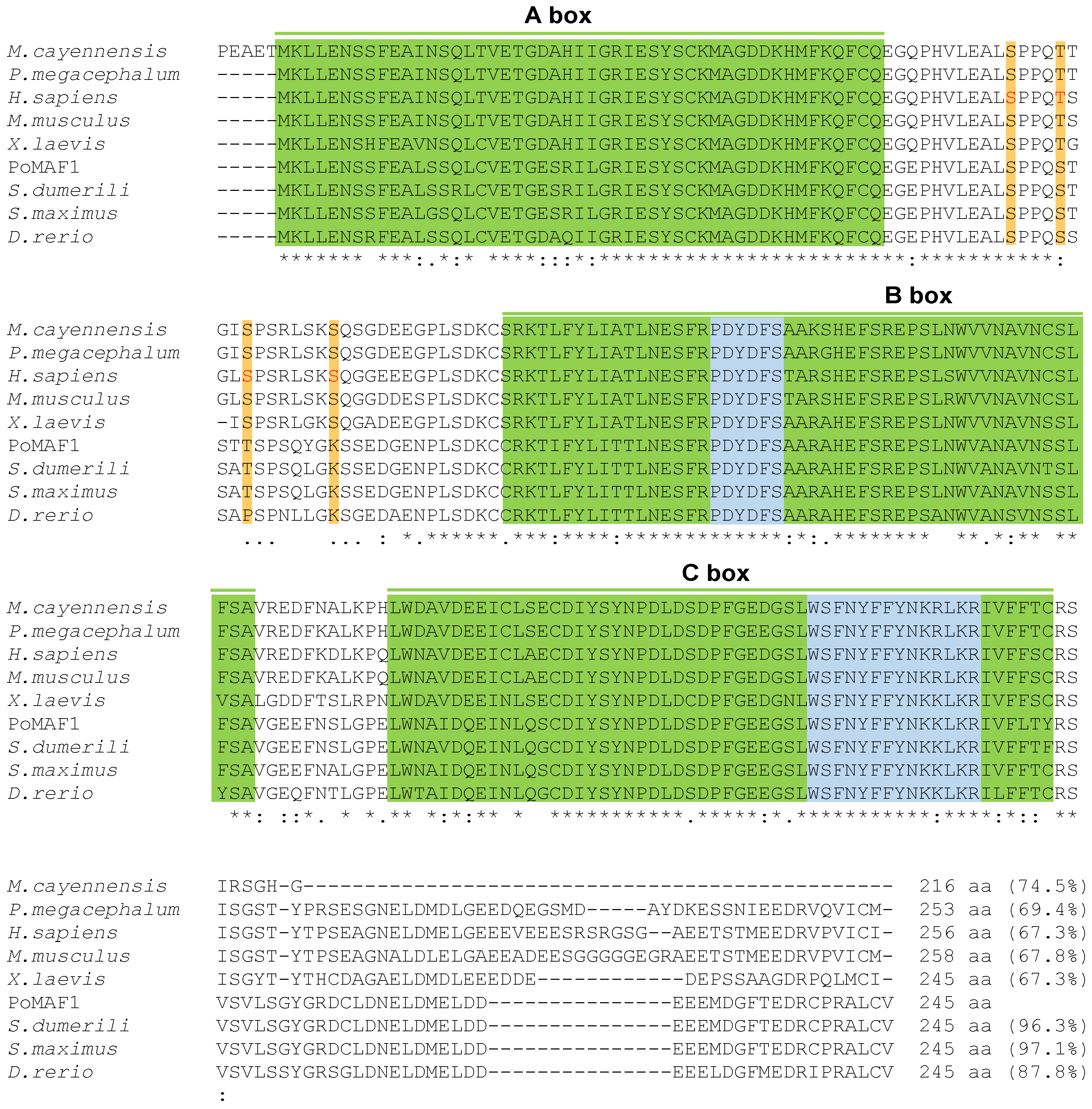
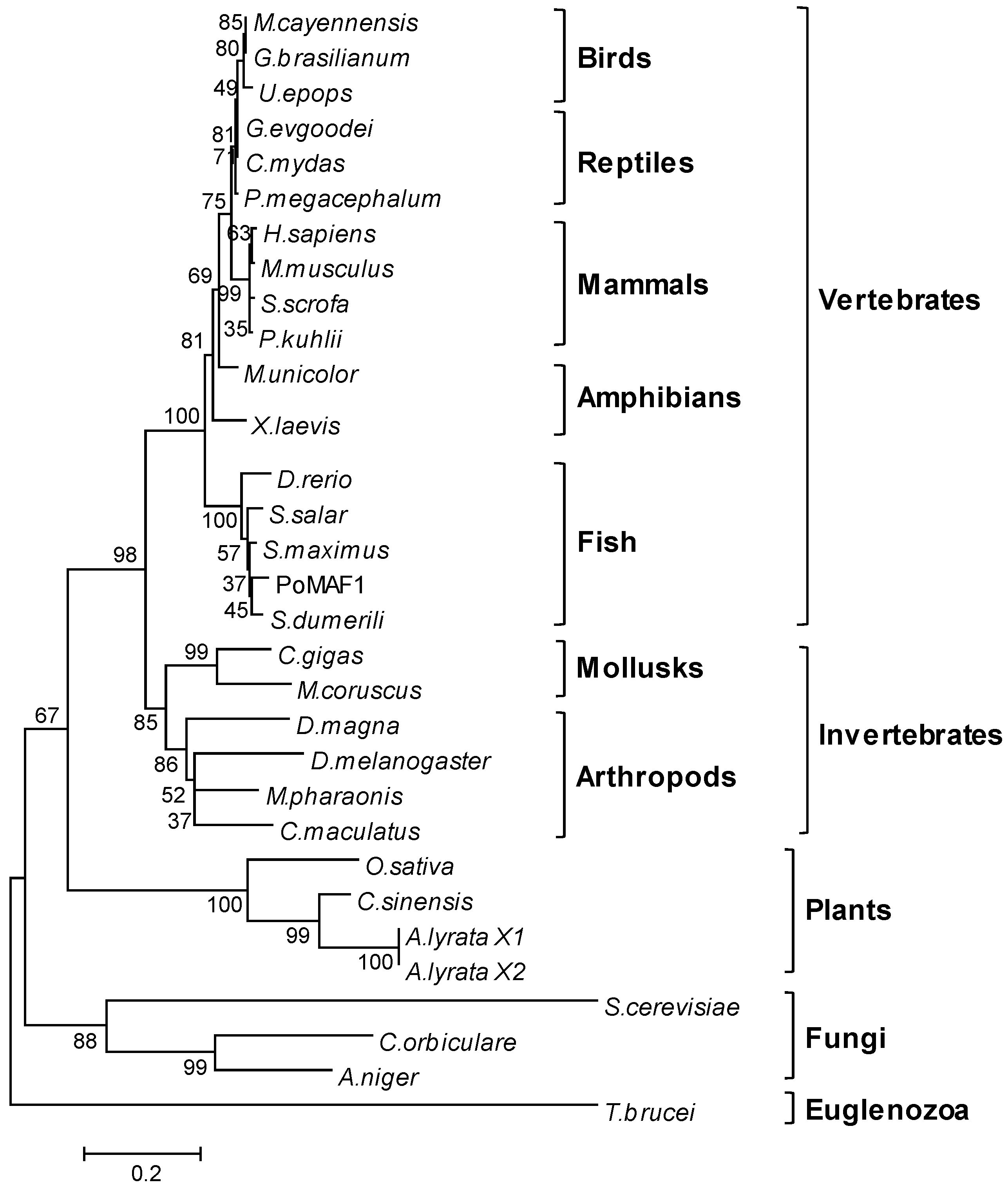
2.2. Amino Acids Sequence and Phylogenetic Analysis of PoMAF1
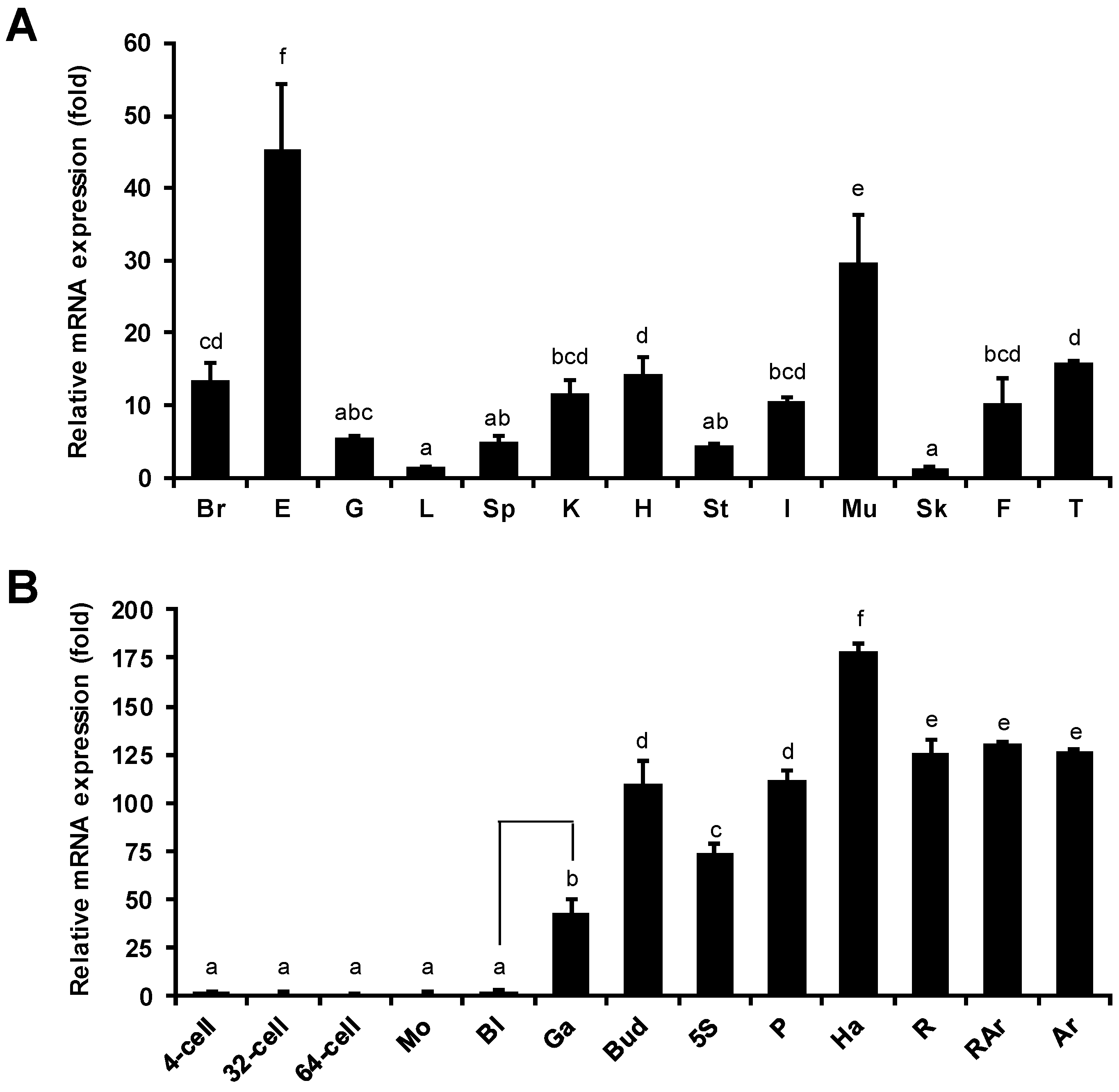
2.3. Tissue Distribution and Developmental Stage-Specific Expression of PoMaf1
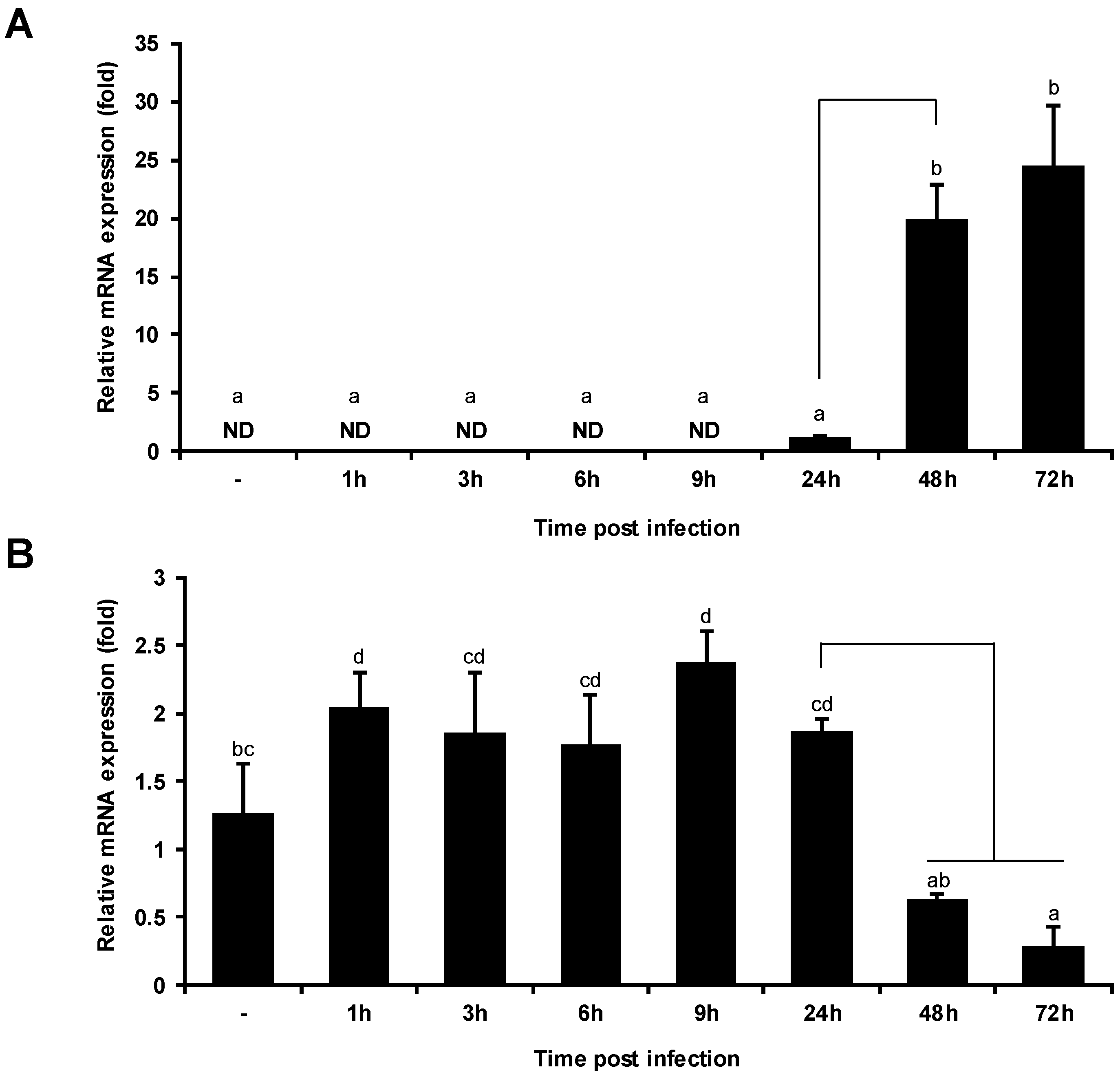
2.4. VHSV Infection Decreases PoMaf1 mRNA Level
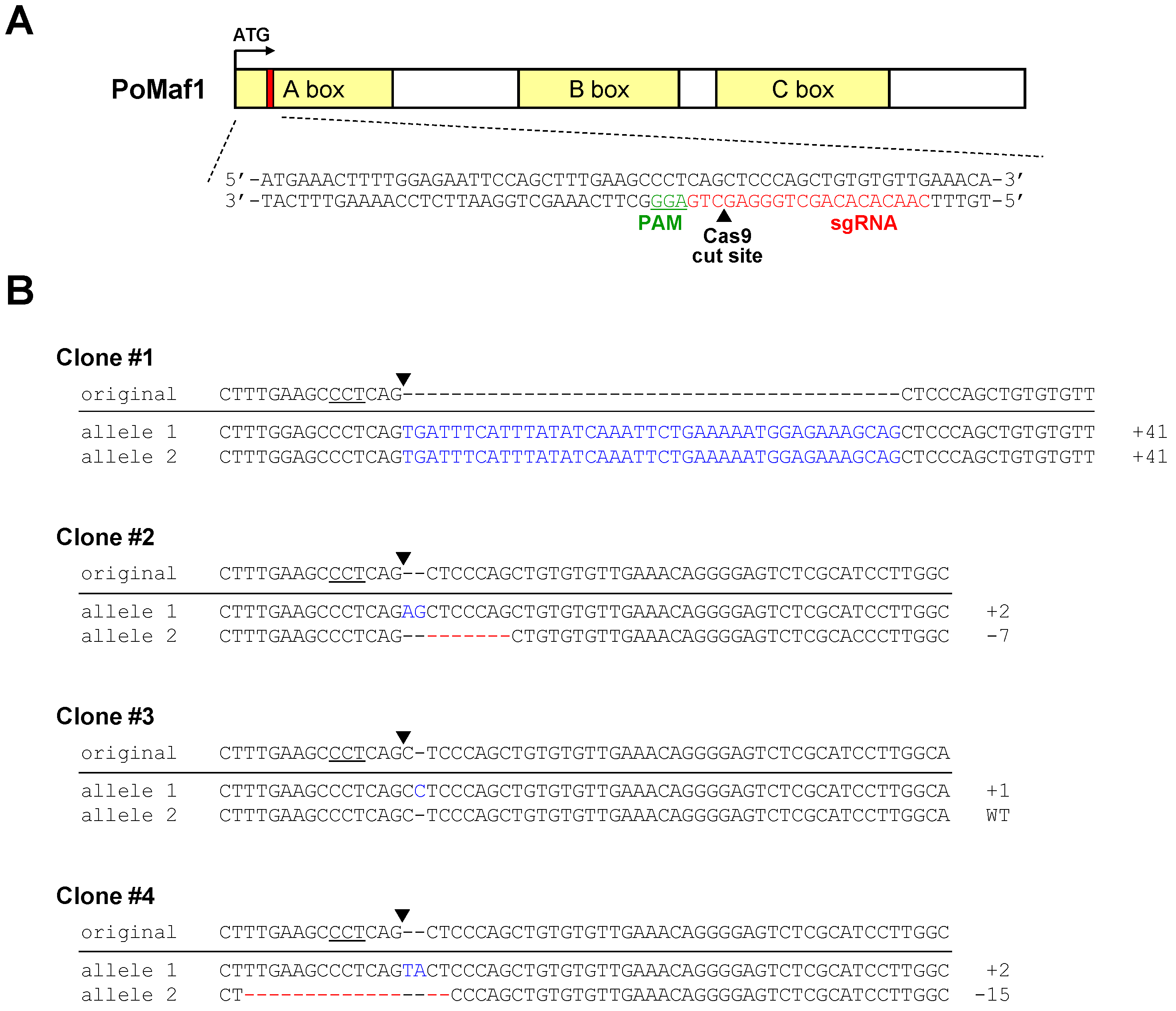
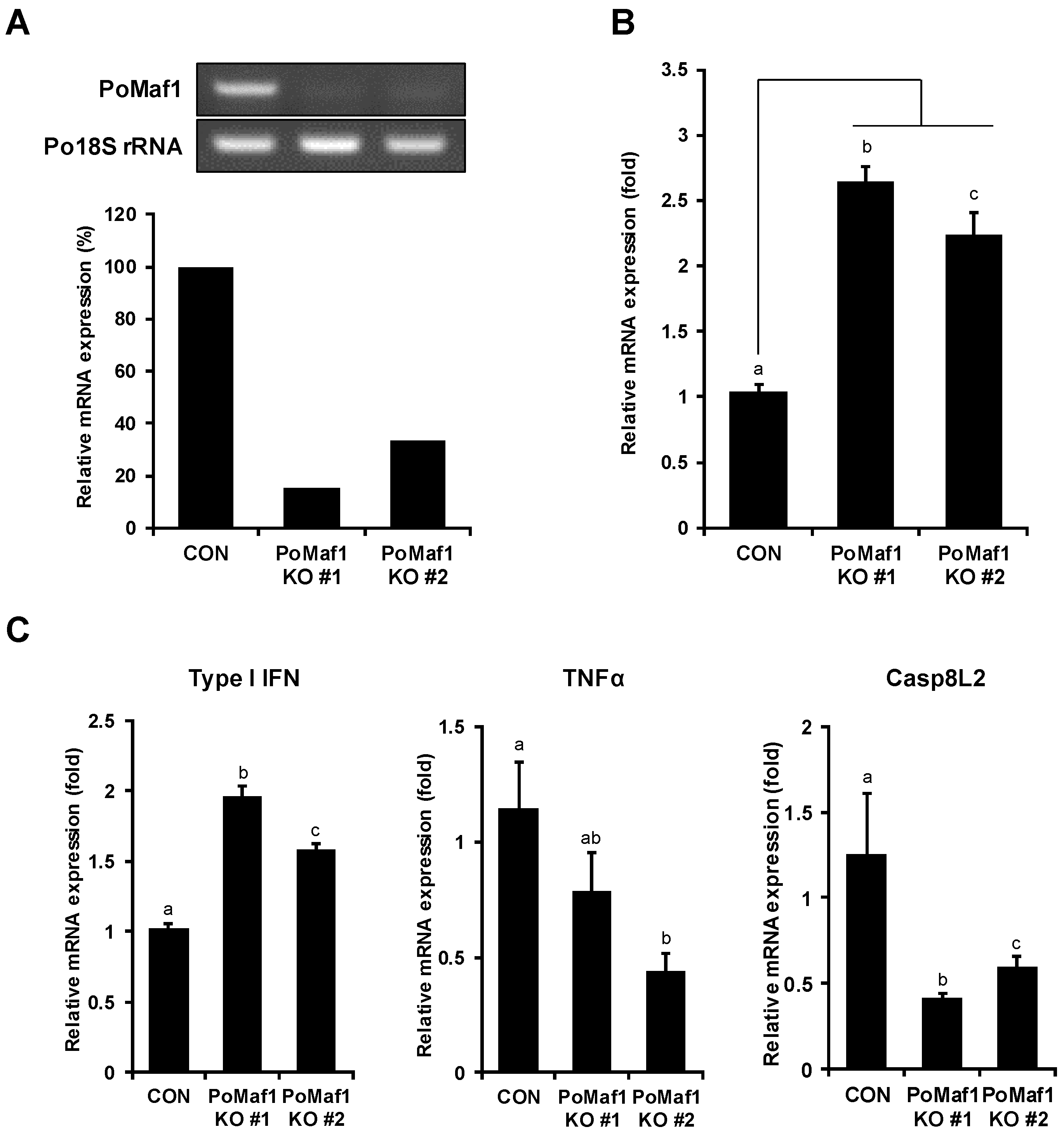
2.5. Disruption of PoMaf1 in HINAE Cells Using CRISPR/Cas9 System
2.6. PoMaf1 Disruption Enhances VHSV G mRNA Level in HINAE Cells
3. Materials and Methods
3.1. Fish and Sample Preparation
3.2. Cloning and Sequencing of PoMaf1 from P. olivaceus
3.3. Amino Acid Sequence Analysis
3.4. Multiple Alignment and Phylogenetic Analysis
3.5. RNA Isolation, Reverse Transcriptase PCR and Quantitative Real-Time PCR
3.6. sgRNA Design and Vector Construction for the CRISPR/Cas9 System
3.7. Cell Culture and Transfection
3.8. Production of Single-Cell Clones of KO Cells
3.9. VHSV Preparation
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vannini, A.; Ringel, R.; Kusser, A.G.; Berninghausen, O.; Kassavetis, G.A.; Cramer, P. Molecular Basis of RNA Polymerase III Transcription Repression by Maf1. Cell 2010, 143, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Pluta, K.; Lefebvre, O.; Martin, N.C.; Smagowicz, W.J.; Stanford, D.R.; Ellis, S.R.; Hopper, A.K.; Sentenac, A.; Boguta, M. Maf1p, a Negative Effector of RNA Polymerase III in Saccharomyces cerevisiae. Mol. Cell. Biol. 2001, 21, 5031–5040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhya, R.; Lee, J.; Willis, I.M. Maf1 Is an Essential Mediator of Diverse Signals that Repress RNA Polymerase III Transcription. Mol. Cell 2002, 10, 1489–1494. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.S.; Zhang, C.; Fromm, J.; Willis, I.M.; Johnson, D.L. Mammalian Maf1 Is a Negative Regulator of Transcription by All Three Nuclear RNA Polymerases. Mol. Cell 2007, 26, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, X.; Wang, H.Y.; Zheng, X.F.S. Beyond regulation of pol III: Role of MAF1 in growth, metabolism, aging and cancer. Biochim. Biophys. Acta Gene Regul. Mech. 2018, 1861, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Kantidakis, T.; Ramsbottom, B.A.; Birch, J.L.; Dowding, S.N.; White, R.J. mTOR associates with TFIIIC, is found at tRNA and 5S rRNA genes, and targets their repressor Maf1. Proc. Natl. Acad. Sci. USA 2010, 107, 11823–11828. [Google Scholar] [CrossRef] [Green Version]
- Palian, B.M.; Rohira, A.D.; Johnson, S.A.S.; He, L.; Zheng, N.; Dubeau, L.; Stiles, B.L.; Johnson, D.L. Maf1 Is a Novel Target of PTEN and PI3K Signaling That Negatively Regulates Oncogenesis and Lipid Metabolism. PLoS Genet. 2014, 10, e1004789. [Google Scholar] [CrossRef] [Green Version]
- Bonhoure, N.; Byrnes, A.; Moir, R.D.; Hodroj, W.; Preitner, F.; Praz, V.; Marcelin, G.; Chua, S.C., Jr.; Martinez-Lopez, N.; Singh, R.; et al. Loss of the RNA polymerase III repressor MAF1 confers obesity resistance. Genes Dev. 2015, 29, 934–947. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Tsang, C.K.; Wang, S.; Li, X.; Yang, Y.; Fu, L.; Huang, W.; Li, M.; Wang, H.; Zheng, X.S. MAF1 suppresses AKT-mTOR signaling and liver cancer through activation of PTEN transcription. Hepatology 2016, 63, 1928–1942. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Chen, C.; Xue, R.; Wang, Y.; Dong, B.; Li, J.; Chen, C.; Jiang, J.; Fan, W.; Liang, Z.; et al. Maf1 ameliorates cardiac hypertrophy by inhibiting RNA polymerase III through ERK1/2. Theranostics 2019, 9, 7268–7281. [Google Scholar] [CrossRef]
- Martin, S.; Saha, B.; Riley, J.L. The Battle over mTOR: An Emerging Theatre in Host–Pathogen Immunity. PLoS Pathog. 2012, 8, e1002894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skall, H.F.; Olesen, N.J.; Mellergaard, S. Viral haemorrhagic septicaemia virus in marine fish and its implications for fish farming—A review. J. Fish Dis. 2005, 28, 509–529. [Google Scholar] [CrossRef] [PubMed]
- Office International des Epizooties (OIE). Available online: https://www.oie.int/en/standard-setting/aquatic-manual/access-online/ (accessed on 10 December 2020).
- Takano, R.; Nishizawa1, T.; Arimoto, M.; Muroga, K. Isolation of viral haemorrhagic septicaemia virus (VHSV) from wild Jap-anese flounder, Paralichthys olivaceus. Bull. Eur. Assoc. Fish Pathol. 2000, 20, 186. [Google Scholar]
- Kim, M.S.; Kim, K.H. Genetically engineered viral hemorrhagic septicemia virus (VHSV) vaccines. Fish Shellfish Immunol. 2019, 95, 11–15. [Google Scholar] [CrossRef]
- Kurita, J.; Iida, Y.; Nakajima, K.; Inouye, K. Virucidal Effects of Various Disinfectants on Viral Hemorrhagic Septicemia Virus(VHSV) Isolated from Japanese Flounder. Fish Pathol. 2002, 37, 175–181. [Google Scholar] [CrossRef]
- Yan, M.; Li, J. The evolving CRISPR technology. Protein Cell 2019, 10, 783–786. [Google Scholar] [CrossRef] [Green Version]
- Gratacap, R.L.; Wargelius, A.; Edvardsen, R.B.; Houston, R.D. Potential of Genome Editing to Improve Aquaculture Breeding and Production. Trends Genet. 2019, 35, 672–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedell, V.M.; Wang, Y.; Campbell, J.M.; Poshusta, T.L.; Starker, C.G.; Krug, R.G., 2nd; Tan, W.; Penheiter, S.G.; Ma, A.C.; Leung, A.Y.; et al. In vivo genome editing using a high-efficiency TALEN system. Nat. Cell Biol. 2012, 491, 114–118. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, M.R.; Oakes, B.L.; Sternberg, S.H.; East-Seletsky, A.; Kaplan, M.; Doudna, J.A. Programmable RNA recognition and cleavage by CRISPR/Cas9. Nat. Cell Biol. 2014, 516, 263–266. [Google Scholar] [CrossRef]
- Cleveland, B.M. A perspective of the future value and challenges of genetic engineering in aquaculture. J. World Aquac. Soc. 2018, 50, 890–893. [Google Scholar] [CrossRef]
- Kim, J.; Cho, J.Y.; Kim, J.-W.; Kim, H.-C.; Noh, J.K.; Kim, Y.-O.; Hwang, H.-K.; Kim, W.-J.; Yeo, S.-Y.; An, C.M.; et al. CRISPR/Cas9-mediated myostatin disruption enhances muscle mass in the olive flounder Paralichthys olivaceus. Aquaculture 2019, 512, 734336. [Google Scholar] [CrossRef]
- Dehler, C.E.; Boudinot, P.; Martin, S.A.M.; Collet, B. Development of an Efficient Genome Editing Method by CRISPR/Cas9 in a Fish Cell Line. Mar. Biotechnol. 2016, 18, 449–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moir, R.D.; Lee, J.; Haeusler, R.A.; Desai, N.; Engelke, D.R.; Willis, I. Protein kinase A regulates RNA polymerase III transcription through the nuclear localization of Maf1. Proc. Natl. Acad. Sci. USA 2006, 103, 15044–15049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Zheng, X.F.S. Maf1 regulation: A model of signal transduction inside the nucleus. Nucleus 2010, 1, 162–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanna, A.; Pradhan, A.; Curran, S.P. Emerging Roles for Maf1 beyond the Regulation of RNA Polymerase III Activity. J. Mol. Biol. 2015, 427, 2577–2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shor, B.; Wu, J.; Shakey, Q.; Toral-Barza, L.; Shi, C.; Follettie, M.; Yu, K. Requirement of the mTOR Kinase for the Regulation of Maf1 Phosphorylation and Control of RNA Polymerase III-dependent Transcription in Cancer Cells. J. Biol. Chem. 2010, 285, 15380–15392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reina, J.H.; Azzouz, T.N.; Hernandez, N. Maf1, a New Player in the Regulation of Human RNA Polymerase III Transcription. PLoS ONE 2006, 1, e134. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, A.; Hammerquist, A.M.; Khanna, A.; Curran, S.P. The C-Box Region of MAF1 Regulates Transcriptional Activity and Protein Stability. J. Mol. Biol. 2017, 429, 192–207. [Google Scholar] [CrossRef] [Green Version]
- Uhlen, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Cardoso, T.F.; Quintanilla, R.; Tibau, J.; Gil, M.; Marmol-Sanchez, E.; González-Rodríguez, O.; González-Prendes, R.; Amills, M. Nutrient supply affects the mRNA expression profile of the porcine skeletal muscle. BMC Genom. 2017, 18, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Willis, I.; Moir, R.D. Integration of nutritional and stress signaling pathways by Maf1. Trends Biochem. Sci. 2007, 32, 51–53. [Google Scholar] [CrossRef]
- Chen, K.; Zhu, L.; Guo, L.; Pan, Y.-B.; Feng, D.-F. Maf1 regulates dendritic morphogenesis and influences learning and memory. Cell Death Dis. 2020, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Hinaux, H.; Devos, L.; Blin, M.; Elipot, Y.; Bibliowicz, J.; Alié, A.; Rétaux, S. Sensory evolution in blind cavefish is driven by early embryonic events during gastrulation and neurulation. Development 2016, 143, 4521–4532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Gao, T.-T.; Fu, X.-Y.; Xu, Z.-H.; Ren, H.; Zhao, P.; Qi, Z.-T.; Qin, Z.-L. PTEN Lipid Phosphatase Activity Enhances Dengue Virus Production through Akt/FoxO1/Maf1 Signaling. Virol. Sin. 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ji, W.-T. PI3K-Akt Signaling and Viral Infection. Recent Patents Biotechnol. 2008, 2, 218–226. [Google Scholar] [CrossRef]
- Fu, X.; Ming, Y.; Li, C.; Niu, Y.; Lin, Q.; Liu, L.; Liang, H.; Huang, Z.; Li, N.-Q. Siniperca chuatsi rhabdovirus (SCRV) induces autophagy via PI3K/Akt-mTOR pathway in CPB cells. Fish Shellfish Immunol. 2020, 102, 381–388. [Google Scholar] [CrossRef]
- Moir, R.D.; Willis, I.M. Regulating Maf1 Expression and Its Expanding Biological Functions. PLoS Genet. 2015, 11, e1004896. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.P.; Fodor, E. Interplay between Influenza Virus and the Host RNA Polymerase II Transcriptional Machinery. Trends Microbiol. 2019, 27, 398–407. [Google Scholar] [CrossRef] [Green Version]
- Sikora, D.; Zhang, D.; Bojic, T.; Beeharry, Y.; Tanara, A.; Pelchat, M. Identification of a Binding Site for ASF/SF2 on an RNA Fragment Derived from the Hepatitis delta Virus Genome. PLoS ONE 2013, 8, e54832. [Google Scholar] [CrossRef] [Green Version]
- Graczyk, D.; White, R.J.; Ryan, K.M. Involvement of RNA Polymerase III in Immune Responses. Mol. Cell. Biol. 2015, 35, 1848–1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, Y.-H.; Macmillan, J.B.; Chen, Z. RNA Polymerase III Detects Cytosolic DNA and Induces Type I Interferons through the RIG-I Pathway. Cell 2009, 138, 576–591. [Google Scholar] [CrossRef] [Green Version]
- Hokonohara, K.; Nishida, N.; Miyoshi, N.; Takahashi, H.; Haraguchi, N.; Hata, T.; Matsuda, C.; Mizushima, T.; Doki, Y.; Mori, M. Involvement of MAF1 homolog, negative regulator of RNA polymerase III in colorectal cancer progression. Int. J. Oncol. 2019, 54, 1001–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtani, M.; Hikima, J.-I.; Kondo, H.; Hirono, I.; Jung, T.-S.; Aoki, T. Evolutional Conservation of Molecular Structure and Antiviral Function of a Viral RNA Receptor, LGP2, in Japanese Flounder, Paralichthys olivaceus. J. Immunol. 2010, 185, 7507–7517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, J.Y.; Lee, U.H.; Heo, M.J.; Jeong, J.M.; Kwon, M.G.; Jee, B.Y.; Park, C.-I.; Park, J.W. RNA-seq transcriptome analysis in flounder cells to compare innate immune responses to low- and high-virulence viral hemorrhagic septicemia virus. Arch. Virol. 2021, 166, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Osborn, M.J.; Lees, C.; McElroy, A.N.; Nielsen, S.C.; Eide, C.R.; Mathews, W.; Feser, C.J.; Tschann, M.; McElmurry, R.; Webber, B.R.; et al. CRISPR/Cas9-Based Cellular Engineering for Targeted Gene Overexpression. Int. J. Mol. Sci. 2018, 19, 946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Xie, Y.; Zhu, Q.; Liu, Y. Targeted Genome Editing in Genes and cis-Regulatory Regions Improves Qualitative and Quantitative Traits in Crops. Mol. Plant 2017, 10, 1368–1370. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Lai, L.; Li, Z. Large-scale genomic deletions mediated by CRISPR/Cas9 system. Oncotarget 2017, 8, 5647. [Google Scholar] [CrossRef]
- Kumar, S.; Tamura, K.; Nei, M. MEGA3: Integrated software for Molecular Evolutionary Genetics Analysis and sequence alignment. Briefings Bioinform. 2004, 5, 150–163. [Google Scholar] [CrossRef] [Green Version]
- Ran, F.A.; Hsu, P.D.; Wright, J.D.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.K.; Kim, J.; Moon, J.Y.; Nam, B.-H.; Kim, Y.-O.; Kim, W.-J.; Park, J.Y.; An, C.M.; Cheong, J.; Kong, H.J. Microarray analysis of gene expression in olive flounder liver infected with viral haemorrhagic septicaemia virus (VHSV). Fish Shellfish Immunol. 2016, 49, 66–78. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Accession No. | Species | Accession No. |
|---|---|---|---|
| Seriola dumerili | XP_022622762.1 | Oryza sativa | XP_015636424.1 |
| Scophthalmus maximus | XP_035474856.1 | Drosophila melanogaster | NP_001015167.2 |
| Homo sapiens | AAH14082.1 | Aspergillus niger | PYH58464.1 |
| Danio rerio | NP_001025410.1 | Citrus sinensis | NP_001275774.1 |
| Mus musculus | AAH16260.1 | Upupa epops | NWU98758.1 |
| Mesembrinibis cayennensis | NXL10327.1 | Glaucidium brasilianum | NXL38974.1 |
| Platysternon megacephalum | TFJ99060.1 | Microcaecilia unicolor | XP_030076765.1 |
| Salmo salar | ACI33843.1 | Xenopus laevis | NP_001080708.1 |
| Saccharomyces cerevisiae | QHB07477.1 | Gopherus evgoodei | XP_030409929.1 |
| Trypanosoma brucei | RHW73349.1 | Chelonia mydas | XP_027690307.1 |
| Colletotrichum orbiculare | TDZ26909.1 | Monomorium pharaonis | XP_012522189.1 |
| Arabidopsis lyrata (MAF1 isoform X1) | XP_020878685.1 | Callosobruchus maculatus | VEN63387.1 |
| Arabidopsis lyrata (MAF1 isoform X2) | XP_020878686.1 | Daphnia magna | XP_032783504.1 |
| Sus scrofa | XP_003125492.2 | Crassostrea gigas | XP_011417596.2 |
| Pipistrellus kuhlii | KAF6276919.1 | Mytilus coruscus | CAC5403110.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Cho, J.Y.; Kim, J.-W.; Kim, D.-G.; Nam, B.-H.; Kim, B.-S.; Kim, W.-J.; Kim, Y.-O.; Cheong, J.; Kong, H.J. Molecular Characterization of Paralichthys olivaceus MAF1 and Its Potential Role as an Anti-Viral Hemorrhagic Septicaemia Virus Factor in Hirame Natural Embryo Cells. Int. J. Mol. Sci. 2021, 22, 1353. https://doi.org/10.3390/ijms22031353
Kim J, Cho JY, Kim J-W, Kim D-G, Nam B-H, Kim B-S, Kim W-J, Kim Y-O, Cheong J, Kong HJ. Molecular Characterization of Paralichthys olivaceus MAF1 and Its Potential Role as an Anti-Viral Hemorrhagic Septicaemia Virus Factor in Hirame Natural Embryo Cells. International Journal of Molecular Sciences. 2021; 22(3):1353. https://doi.org/10.3390/ijms22031353
Chicago/Turabian StyleKim, Julan, Ja Young Cho, Ju-Won Kim, Dong-Gyun Kim, Bo-Hye Nam, Bong-Seok Kim, Woo-Jin Kim, Young-Ok Kim, JaeHun Cheong, and Hee Jeong Kong. 2021. "Molecular Characterization of Paralichthys olivaceus MAF1 and Its Potential Role as an Anti-Viral Hemorrhagic Septicaemia Virus Factor in Hirame Natural Embryo Cells" International Journal of Molecular Sciences 22, no. 3: 1353. https://doi.org/10.3390/ijms22031353
APA StyleKim, J., Cho, J. Y., Kim, J. -W., Kim, D. -G., Nam, B. -H., Kim, B. -S., Kim, W. -J., Kim, Y. -O., Cheong, J., & Kong, H. J. (2021). Molecular Characterization of Paralichthys olivaceus MAF1 and Its Potential Role as an Anti-Viral Hemorrhagic Septicaemia Virus Factor in Hirame Natural Embryo Cells. International Journal of Molecular Sciences, 22(3), 1353. https://doi.org/10.3390/ijms22031353