
Regulation of Seed Dormancy and Germination Mechanisms in a Changing Environment


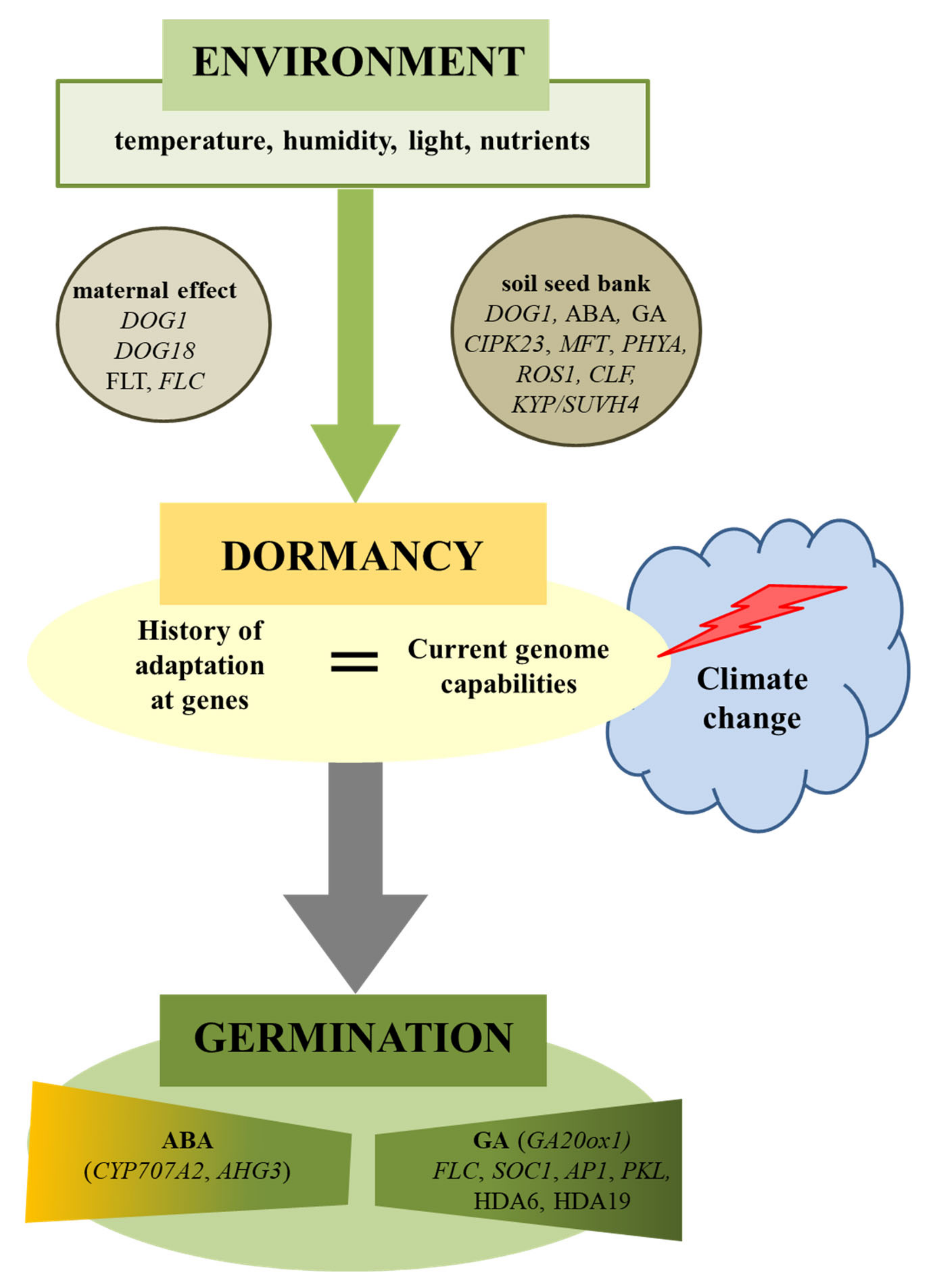{kind=link}
Abstract
:1. Introduction
2. Adaptations of Seed Germination to the Changing Environment
3. Mechanisms of Adaptation of Seed Germination to the Changing Environment
3.1. Physiological Control of Seed Dormancy and Germination
3.2. Molecular Control of Seed Dormancy and Germination
3.3. Epigenetic Control of Seed Dormancy and Germination
4. Conclusions
Funding
Conflicts of Interest
References
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed Dormancy and the Control of Germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef] [PubMed]
- Job, D.; Caboche, M. The Seeds of Life. C. R. Biol. 2008, 331, 711–714. [Google Scholar] [CrossRef] [PubMed]
- Rajjou, L.; Debeaujon, I. Seed Longevity: Survival and Maintenance of High Germination Ability of Dry Seeds. C. R. Biol. 2008, 331, 796–805. [Google Scholar] [CrossRef] [PubMed]
- Donohue, K.; de Casas, R.R.; Burghardt, L.; Kovach, K.; Willis, C.G. Germination, Postgermination Adaptation, and Species Ecological Ranges. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 293–319. [Google Scholar] [CrossRef]
- Chen, S.-C.; Poschlod, P.; Antonelli, A.; Liu, U.; Dickie, J.B. Trade-off between Seed Dispersal in Space and Time. Ecol. Lett. 2020, 23, 1635–1642. [Google Scholar] [CrossRef] [PubMed]
- Graeber, K.; Nakabayashi, K.; Miatton, E.; Leubner-Metzger, G.; Soppe, W.J.J. Molecular Mechanisms of Seed Dormancy. Plant Cell Environ. 2012, 35, 1769–1786. [Google Scholar] [CrossRef]
- Long, R.L.; Gorecki, M.J.; Renton, M.; Scott, J.K.; Colville, L.; Goggin, D.E.; Commander, L.E.; Westcott, D.A.; Cherry, H.; Finch-Savage, W.E. The Ecophysiology of Seed Persistence: A Mechanistic View of the Journey to Germination or Demise. Biol. Rev. 2015, 90, 31–59. [Google Scholar] [CrossRef]
- Walck, J.; Hidayati, S.; Dixon, K.; Thompson, K.; Poschlod, P. Climate Change and Plant Regeneration from Seed. Glob. Chang. Biol. 2011, 17, 2145–2161. [Google Scholar] [CrossRef]
- Montague, J.L.; Barrett, S.C.H.; Eckert, C.G. Re-Establishment of Clinal Variation in Flowering Time among Introduced Populations of Purple Loosestrife (Lythrum Salicaria, Lythraceae). J. Evol. Biol. 2008, 21, 234–245. [Google Scholar] [CrossRef]
- Fenner, M. The Effects of the Parent Environment on Seed Germinability. Seed Sci. Res. 1991, 1, 75–84. [Google Scholar] [CrossRef]
- Donohue, K. Completing the Cycle: Maternal Effects as the Missing Link in Plant Life Histories. Philos. Trans. R Soc. Lond. B Biol. Sci. 2009, 364, 1059–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Ölçer-Footitt, H.; Footitt, S.; Finch-Savage, W.E. Seed Dormancy Is a Dynamic State: Variable Responses to Pre- and Post-Shedding Environmental Signals in Seeds of Contrasting Arabidopsis Ecotypes. Seed Sci. Res. 2015, 25, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Ooi, M.K.J. Seed Bank Persistence and Climate Change. Seed Sci. Res. 2012, 22, S53–S60. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Pascual, E.; Jimenez-Alfaro, B.; Caujape-Castells, J.; Jaen-Molina, R.; Emilio Diaz, T. A Local Dormancy Cline Is Related to the Seed Maturation Environment, Population Genetic Composition and Climate. Ann. Bot. 2013, 112, 937–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biere, A. Parental Effects in Lychnis Flos-Cuculi. I: Seed Size, Germination and Seedling Performance in a Controlled Environment. J. Evol. Biol. 1991, 4, 447–465. [Google Scholar] [CrossRef]
- Lacey, E.; Smith, S.; Case, A. Parental Effects on Seed Mass: Seed Coat but Not Embryo/Endosperm Effects. Am. J. Bot. 1997, 84, 1617. [Google Scholar] [CrossRef] [Green Version]
- Vange, V.; Heuch, I.; Vandvik, V. Do Seed Mass and Family Affect Germination and Juvenile Performance in Knautia arvensis? A Study Using Failure-Time Methods. Acta Oecologica 2004, 25, 169–178. [Google Scholar] [CrossRef]
- Daws, M.I.; Cleland, H.; Chmielarz, P.; Gorian, F.; Leprince, O.; Mullins, C.E.; Thanos, C.A.; Vandvik, V.; Pritchard, H.W. Variable Desiccation Tolerance in Acer pseudoplatanus Seeds in Relation to Developmental Conditions: A Case of Phenotypic Recalcitrance? Funct. Plant Biol. 2006, 33, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Fenner, M.; Thompson, K. The Ecology of Seeds. Available online: /core/books/the-ecology-of-seeds/61AA99663FC86CA7E0E0484AAF37F791 (accessed on 31 January 2018).
- Huang, Z.; Footitt, S.; Finch-Savage, W.E. The Effect of Temperature on Reproduction in the Summer and Winter Annual Arabidopsis thaliana Ecotypes Bur and Cvi. Ann. Bot. 2014, 113, 921–929. [Google Scholar] [CrossRef] [Green Version]
- Springthorpe, V.; Penfield, S. Flowering Time and Seed Dormancy Control Use External Coincidence to Generate Life History Strategy. Elife 2015, 4. [Google Scholar] [CrossRef]
- Rodríguez, M.V.; Barrero, J.M.; Corbineau, F.; Gubler, F.; Benech-Arnold, R.L. Dormancy in Cereals (Not Too Much, Not so Little): About the Mechanisms behind This Trait. Seed Sci. Res. 2015, 25, 99–119. [Google Scholar] [CrossRef] [Green Version]
- Peters, N.C.B. The Dormancy of Wild Oat Seed (Avena fatua L.) from Plants Grown under Various Temperature and Soil Moisture Conditions. Weed Res. 1982, 22, 205–212. [Google Scholar] [CrossRef]
- Alboresi, A.; Gestin, C.; Leydecker, M.-T.; Bedu, M.; Meyer, C.; Truong, H.-N. Nitrate, a Signal Relieving Seed Dormancy in Arabidopsis. Plant Cell Environ. 2005, 28, 500–512. [Google Scholar] [CrossRef]
- Matakiadis, T.; Alboresi, A.; Jikumaru, Y.; Tatematsu, K.; Pichon, O.; Renou, J.-P.; Kamiya, Y.; Nambara, E.; Truong, H.-N. The Arabidopsis Abscisic Acid Catabolic Gene CYP707A2 Plays a Key Role in Nitrate Control of Seed Dormancy. Plant Physiol. 2009, 149, 949–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and, Evolution of Dormancy and Germination; Elsevier: San Diego, CA, USA, 1998; ISBN 978-0-08-054086-3. [Google Scholar]
- Finch-Savage, W.E.; Footitt, S. Seed Dormancy Cycling and the Regulation of Dormancy Mechanisms to Time Germination in Variable Field Environments. J. Exp. Bot. 2017, 68, 843–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Probert, R.J. The role of temperature in the regulation of seed dormancy and germination. In Seeds: The Ecology of Regeneration in Plant Communities; Fenner, M., Ed.; CABI: Wallingford, UK, 2000; pp. 261–292. ISBN 978-0-85199-432-1. [Google Scholar]
- Evans, M.E.K.; Dennehy, J.J. Germ Banking: Bet-Hedging and Variable Release from Egg and Seed Dormancy. Q. Rev. Biol. 2005, 80, 431–451. [Google Scholar] [CrossRef] [Green Version]
- Vleeshouwers, L.M.; Bouwmeester, H.J.; Karssen, C.M. Redefining Seed Dormancy: An Attempt to Integrate Physiology and Ecology. J. Ecol. 1995, 83, 1031–1037. [Google Scholar]
- Benech-Arnold, R.L.; Sánchez, R.A.; Forcella, F.; Kruk, B.C.; Ghersa, C.M. Environmental Control of Dormancy in Weed Seed Banks in Soil. Field Crop. Res. 2000, 67, 105–122. [Google Scholar] [CrossRef]
- Vleeshouwers, L.M.; Bouwmeester, H.J. A Simulation Model for Seasonal Changes in Dormancy and Germination of Weed Seeds. Seed Sci. Res. 2001, 11, 77–92. [Google Scholar] [CrossRef]
- Footitt, S.; Douterelo-Soler, I.; Clay, H.; Finch-Savage, W.E. Dormancy Cycling in Arabidopsis Seeds Is Controlled by Seasonally Distinct Hormone-Signaling Pathways. Proc. Natl. Acad. Sci. USA 2011, 108, 20236–20241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Footitt, S.; Huang, Z.; Clay, H.A.; Mead, A.; Finch-Savage, W.E. Temperature, Light and Nitrate Sensing Coordinate Arabidopsis Seed Dormancy Cycling, Resulting in Winter and Summer Annual Phenotypes. Plant J. 2013, 74, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Long, R.L.; Stevens, J.C.; Griffiths, E.M.; Adamek, M.; Gorecki, M.J.; Powles, S.B.; Merritt, D.J. Seeds of Brassicaceae Weeds Have an Inherent or Inducible Response to the Germination Stimulant Karrikinolide. Ann. Bot. 2011, 108, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Long, R.L.; Stevens, J.C.; Griffiths, E.M.; Adamek, M.; Powles, S.B.; Merritt, D.J. Detecting Karrikinolide Responses in Seeds of the Poaceae. Aust. J. Bot. 2011, 59, 610. [Google Scholar] [CrossRef]
- Fernández Farnocchia, R.B.; Benech-Arnold, R.L.; Batlla, D. Regulation of Seed Dormancy by the Maternal Environment Is Instrumental for Maximizing Plant Fitness in Polygonum Aviculare. J. Exp. Bot. 2019, 70, 4793–4806. [Google Scholar] [CrossRef] [PubMed]
- Batlla, D.; Benech-Arnold, R.L. Predicting Changes in Dormancy Level in Natural Seed Soil Banks. Plant Mol. Biol. 2010, 73, 3–13. [Google Scholar] [CrossRef]
- Frachon, L.; Bartoli, C.; Carrère, S.; Bouchez, O.; Chaubet, A.; Gautier, M.; Roby, D.; Roux, F. A Genomic Map of Climate Adaptation in Arabidopsis thaliana at a Micro-Geographic Scale. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Sundqvist, L.; Godhe, A.; Jonsson, P.R.; Sefbom, J. The Anchoring Effect—Long-Term Dormancy and Genetic Population Structure. ISME J. 2018, 12, 2929–2941. [Google Scholar] [CrossRef]
- Huang, X.; Schmitt, J.; Dorn, L.; Griffith, C.; Effgen, S.; Takao, S.; Koornneef, M.; Donohue, K. The Earliest Stages of Adaptation in an Experimental Plant Population: Strong Selection on QTLS for Seed Dormancy. Mol. Ecol. 2010, 19, 1335–1351. [Google Scholar] [CrossRef]
- Staszak, A.M.; Pawłowski, T. Forest Tree Research in Post Genomic Era. Introduction to Systems Biology of Broadleaves. Dendrobiology 2012, 68, 113–123. [Google Scholar]
- Mondoni, A.; Pedrini, S.; Bernareggi, G.; Rossi, G.; Abeli, T.; Probert, R.J.; Ghitti, M.; Bonomi, C.; Orsenigo, S. Climate Warming Could Increase Recruitment Success in Glacier Foreland Plants. Ann. Bot. 2015, 116, 907–916. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.T.; Harper, J.L. Seed Polymorphism and Germination. Weed Res. 1965, 5, 141–150. [Google Scholar] [CrossRef]
- Jurado, E.; Flores, J. Is Seed Dormancy under Environmental Control or Bound to Plant Traits? J. Veg. Sci. 2005, 16, 559–564. [Google Scholar] [CrossRef]
- Rubio de Casas, R.; Willis, C.G.; Pearse, W.D.; Baskin, C.C.; Baskin, J.M.; Cavender-Bares, J. Global Biogeography of Seed Dormancy Is Determined by Seasonality and Seed Size: A Case Study in the Legumes. New Phytol. 2017, 214, 1527–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawłowski, T.A.; Klupczyńska, E.A.; Staszak, A.M.; Suszka, J. Proteomic Analysis of Black poplar (Populus nigra L.) Seed Storability. Ann. For. Sci. 2019, 76, 104. [Google Scholar] [CrossRef] [Green Version]
- Dyderski, M.K.; Paz, S.; Frelich, L.E.; Jagodzinski, A.M. How Much Does Climate Change Threaten European Forest Tree Species Distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Fernández-Pascual, E.; Seal, C.E.; Pritchard, H.W. Simulating the Germination Response to Diurnally Alternating Temperatures under Climate Change Scenarios: Comparative Studies on Carex diandra Seeds. Ann. Bot. 2015, 115, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Pawłowski, T.A.; Bujarska-Borkowska, B.; Suszka, J.; Tylkowski, T.; Chmielarz, P.; Klupczyńska, E.A.; Staszak, A.M. Temperature Regulation of Primary and Secondary Seed Dormancy in Rosa canina L.: Findings from Proteomic Analysis. Int. J. Mol. Sci. 2020, 21, 7008. [Google Scholar] [CrossRef]
- Gareca, E.E.; Vandelook, F.; Fernández, M.; Hermy, M.; Honnay, O. Seed Germination, Hydrothermal Time Models and the Effects of Global Warming on a Threatened High Andean Tree Species. Seed Sci. Res. 2012, 22, 287–298. [Google Scholar] [CrossRef]
- Ooi, M.K.J.; Auld, T.D.; Denham, A.J. Climate Change and Bet-Hedging: Interactions between Increased Soil Temperatures and Seed Bank Persistence. Glob. Chang. Biol. 2009, 15, 2375–2386. [Google Scholar] [CrossRef]
- Penfield, S.; MacGregor, D.R. Effects of Environmental Variation during Seed Production on Seed Dormancy and Germination. J. Exp. Bot. 2017, 68, 819–825. [Google Scholar] [CrossRef] [Green Version]
- Lippmann, R.; Babben, S.; Menger, A.; Delker, C.; Quint, M. Development of Wild and Cultivated Plants under Global Warming Conditions. Curr. Biol. 2019, 29, R1326–R1338. [Google Scholar] [CrossRef]
- Chamorro, D.; Luna, B.; Moreno, J.M. Germination Responses to Current and Future Temperatures of Four Seeder Shrubs across a Latitudinal Gradient in Western Iberia. Am. J. Bot. 2017, 104, 83–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, L.; Milberg, P. Variation in Seed Dormancy among Mother Plants, Populations and Years of Seed Collection. Seed Sci. Res. 1998, 8, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Schütz, W.; Rave, G. Variation in Seed Dormancy of the Wetland Sedge, Carex Elongata, between Populations and Individuals in Two Consecutive Years. Seed Sci. Res. 2003, 13, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Koutecká, E.; Lepš, J. Effect of Light and Moisture Conditions and Seed Age on Germination of Three Closely Related Myosotis Species. Folia Geobot. 2009, 44, 109. [Google Scholar] [CrossRef]
- Herranz, J.M.; Copete, M.Á.; Ferrandis, P.; Copete, E. Intermediate Complex Morphophysiological Dormancy in the Endemic Iberian Aconitum napellus Subsp. castellanum (Ranunculaceae). Seed Sci. Res. 2010, 20, 109–121. [Google Scholar] [CrossRef]
- Ren, Z.; Abbott, R.J. Seed Dormancy in Mediterranean Senecio vulgaris L. New Phytol. 1991, 117, 673–678. [Google Scholar] [CrossRef]
- Skordilis, A.; Thanos, C.A. Seed Stratification and Germination Strategy in the Mediterranean Pines Pinus brutia and P. halepensis. Seed Sci. Res. 1995, 5, 151–160. [Google Scholar] [CrossRef]
- Wagmann, K.; Hautekèete, N.-C.; Piquot, Y.; Meunier, C.; Schmitt, S.E.; Van Dijk, H. Seed Dormancy Distribution: Explanatory Ecological Factors. Ann. Bot. 2012, 110, 1205–1219. [Google Scholar] [CrossRef] [Green Version]
- Beardsell, D.; Mullet, J. Seed Generation of Eucalyptus Pauciflora Sieb. Ex Spreng. From Low and High Altitude Populations in Victoria. Aust. J. Bot. 1984, 32, 475–480. [Google Scholar] [CrossRef]
- Holm, S.-O. Reproductive Patterns of Betula Pendula and B. Pubescens Coll. along a Regional Altitudinal Gradient in Northern Sweden. Ecography 1994, 17, 60–72. [Google Scholar] [CrossRef]
- Cavieres, L.A.; Arroyo, M.T.K. Seed Germination Response to Cold Stratification Period and Thermal Regime in Phacelia Secunda (Hydrophyllaceae)—Altitudinal Variation in the Mediterranean Andes of Central Chile. Plant Ecol. 2000, 149, 1–8. [Google Scholar] [CrossRef]
- Meyer, S.E.; Monsen, S.B. Habitat-Correlated Variation in Mountain Big Sagebrush (Artemisia tridentata ssp. Vaseyana) Seed Germination Patterns. Ecology 1991, 72, 739–742. [Google Scholar]
- El-Keblawy, A.A.; Shaltout, K.H.; Doust, J.L.; Doust, L.L. Maternal Effects on Progeny in Thymelaea hirsuta. New Phytol. 1996, 132, 77–85. [Google Scholar] [CrossRef]
- Arana, M.V.; Gonzalez-Polo, M.; Martinez-Meier, A.; Gallo, L.A.; Benech-Arnold, R.L.; Sánchez, R.A.; Batlla, D. Seed Dormancy Responses to Temperature Relate to Nothofagus Species Distribution and Determine Temporal Patterns of Germination across Altitudes in Patagonia. New Phytol. 2016, 209, 507–520. [Google Scholar] [CrossRef] [PubMed]
- Chamorro, D.; Luna, B.; Moreno, J.M. Local Climate Controls Among-Population Variation in Germination Patterns in Two Erica Species across Western Iberia. Seed Sci. Res. 2018, 28, 112–122. [Google Scholar] [CrossRef]
- Dwyer, J.M.; Erickson, T.E. Warmer Seed Environments Increase Germination Fractions in Australian Winter Annual Plant Species. Ecosphere 2016, 7, e01497. [Google Scholar] [CrossRef]
- Seal, C.E.; Daws, M.I.; Flores, J.; Ortega-Baes, P.; Galíndez, G.; León-Lobos, P.; Sandoval, A.; Stuva, A.C.; Bullón, N.R.; Dávila-Aranda, P.; et al. Thermal Buffering Capacity of the Germination Phenotype across the Environmental Envelope of the Cactaceae. Glob. Chang. Biol. 2017, 23, 5309–5317. [Google Scholar] [CrossRef]
- Dantas, B.F.; Moura, M.S.B.; Pelacani, C.R.; Angelotti, F.; Taura, T.A.; Oliveira, G.M.; Bispo, J.S.; Matias, J.R.; Silva, F.F.S.; Pritchard, H.W.; et al. Rainfall, Not Soil Temperature, Will Limit the Seed Germination of Dry Forest Species with Climate Change. Oecologia 2020, 192, 529–541. [Google Scholar] [CrossRef]
- Stevens, N.; Seal, C.E.; Archibald, S.; Bond, W. Increasing Temperatures Can Improve Seedling Establishment in Arid-Adapted Savanna Trees. Oecologia 2014, 175, 1029–1040. [Google Scholar] [CrossRef]
- Carta, A.; Probert, R.; Puglia, G.; Peruzzi, L.; Bedini, G. Local Climate Explains Degree of Seed Dormancy in Hypericum elodes L. (Hypericaceae). Plant Biol. 2016, 18 (Suppl. S1), 76–82. [Google Scholar] [CrossRef] [PubMed]
- Orsenigo, S.; Abeli, T.; Rossi, G.; Bonasoni, P.; Pasquaretta, C.; Gandini, M.; Mondoni, A. Effects of Autumn and Spring Heat Waves on Seed Germination of High Mountain Plants. PLoS ONE 2015, 10, e0133626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondoni, A.; Rossi, G.; Orsenigo, S.; Probert, R.J. Climate Warming Could Shift the Timing of Seed Germination in Alpine Plants. Ann. Bot. 2012, 110, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Orrù, M.; Mattana, E.; Pritchard, H.W.; Bacchetta, G. Thermal Thresholds as Predictors of Seed Dormancy Release and Germination Timing: Altitude-Related Risks from Climate Warming for the Wild Grapevine Vitis vinifera Subsp. Sylvestris. Ann. Bot. 2012, 110, 1651–1660. [Google Scholar] [CrossRef] [Green Version]
- Gosling, P.G.; McCartan, S.A.; Peace, A.J. Seed Dormancy and Germination Characteristics of Common Alder (Alnus glutinosa L.) Indicate Some Potential to Adapt to Climate Change in Britain. Forestry 2009, 82, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Vidigal, D.S.; Marques, A.C.S.S.; Willems, L.A.J.; Buijs, G.; Méndez-Vigo, B.; Hilhorst, H.W.M.; Bentsink, L.; Picó, F.X.; Alonso-Blanco, C. Altitudinal and Climatic Associations of Seed Dormancy and Flowering Traits Evidence Adaptation of Annual Life Cycle Timing in Arabidopsis thaliana. Plant Cell Environ. 2016, 39, 1737–1748. [Google Scholar] [CrossRef] [Green Version]
- Carta, A.; Hanson, S.; Müller, J.V. Plant Regeneration from Seeds Responds to Phylogenetic Relatedness and Local Adaptation in Mediterranean Romulea (Iridaceae) Species. Ecol. Evol. 2016, 6, 4166–4178. [Google Scholar] [CrossRef]
- Escudero, A.; Pérez-García, F.; Luzuriaga, A.L. Effects of Light, Temperature and Population Variability on the Germination of Seven Spanish Pines. Seed Sci. Res. 2002, 12, 261–271. [Google Scholar] [CrossRef]
- Cochrane, A.; Yates, C.J.; Hoyle, G.L.; Nicotra, A.B. Will Among-Population Variation in Seed Traits Improve the Chance of Species Persistence under Climate Change? Glob. Ecol. Biogeogr. 2015, 24, 12–24. [Google Scholar] [CrossRef]
- Chiang, G.C.K.; Bartsch, M.; Barua, D.; Nakabayashi, K.; Debieu, M.; Kronholm, I.; Koornneef, M.; Soppe, W.J.J.; Donohue, K.; De MEAUX, J. DOG1 Expression Is Predicted by the Seed-Maturation Environment and Contributes to Geographical Variation in Germination in Arabidopsis thaliana. Mol. Ecol. 2011, 20, 3336–3349. [Google Scholar] [CrossRef]
- Baskin, J.; Baskin, C. A Classification System for Seed Dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Willis, C.G.; Baskin, C.C.; Baskin, J.M.; Auld, J.R.; Venable, D.L.; Cavender-Bares, J.; Donohue, K.; Rubio de Casas, R.; The NESCent Germination Working Group. The Evolution of Seed Dormancy: Environmental Cues, Evolutionary Hubs, and Diversification of the Seed Plants. New Phytol. 2014, 203, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Staszak, A.M.; Guzicka, M.; Pawłowski, T.A. Signalling Regulators of Abscisic and Gibberellic Acid Pathways Are Involved in Dormancy Breaking of Norway Maple (Acer platanoides L.) Seeds. Acta Physiol. Plant 2017, 39, 251. [Google Scholar] [CrossRef] [Green Version]
- Staszak, A.M.; Rewers, M.; Sliwinska, E.; Klupczyńska, E.A.; Pawłowski, T.A. DNA Synthesis Pattern, Proteome, and ABA and GA Signalling in Developing Seeds of Norway Maple (Acer platanoides). Funct. Plant Biol. 2019, 46, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Donohue, K. Seeds and Seasons: Interpreting Germination Timing in the Field. Seed Sci. Res. 2005, 15, 175–187. [Google Scholar] [CrossRef] [Green Version]
- North, H.; Baud, S.; Debeaujon, I.; Dubos, C.; Dubreucq, B.; Grappin, P.; Jullien, M.; Lepiniec, L.; Marion-Poll, A.; Miquel, M.; et al. Arabidopsis Seed Secrets Unravelled after a Decade of Genetic and Omics-Driven Research. Plant J. 2010, 61, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, B.J.W.; Bentsink, L. Regulation of Seed Dormancy by Abscisic Acid and Delay of Germination 1. Seed Sci. Res. 2015, 25, 82–98. [Google Scholar] [CrossRef]
- Johnston, I.G.; Bassel, G.W. Identification of a Bet-Hedging Network Motif Generating Noise in Hormone Concentrations and Germination Propensity in Arabidopsis. J. R. Soc. Interface 2018, 15, 20180042. [Google Scholar] [CrossRef] [Green Version]
- Nambara, E.; Okamoto, M.; Tatematsu, K.; Yano, R.; Seo, M.; Kamiya, Y. Abscisic Acid and the Control of Seed Dormancy and Germination. Seed Sci. Res. 2010, 20, 55–67. [Google Scholar] [CrossRef]
- Bassel, G.W.; Lan, H.; Glaab, E.; Gibbs, D.J.; Gerjets, T.; Krasnogor, N.; Bonner, A.J.; Holdsworth, M.J.; Provart, N.J. Genome-Wide Network Model Capturing Seed Germination Reveals Coordinated Regulation of Plant Cellular Phase Transitions. PNAS 2011, 108, 9709–9714. [Google Scholar] [CrossRef] [Green Version]
- Morris, K.; Linkies, A.; Müller, K.; Oracz, K.; Wang, X.; Lynn, J.R.; Leubner-Metzger, G.; Finch-Savage, W.E. Regulation of Seed Germination in the Close Arabidopsis Relative Lepidium sativum: A Global Tissue-Specific Transcript Analysis. Plant Physiol. 2011, 155, 1851–1870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekkers, B.J.W.; Pearce, S.; van Bolderen-Veldkamp, R.P.; Marshall, A.; Widera, P.; Gilbert, J.; Drost, H.-G.; Bassel, G.W.; Müller, K.; King, J.R.; et al. Transcriptional Dynamics of Two Seed Compartments with Opposing Roles in Arabidopsis Seed Germination. Plant Physiol. 2013, 163, 205–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finch-Savage, W.E.; Bassel, G.W. Seed Vigour and Crop Establishment: Extending Performance beyond Adaptation. J. Exp. Bot. 2016, 67, 567–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassel, G.W. To Grow or Not to Grow? Trends Plant Sci. 2016, 21, 498–505. [Google Scholar] [CrossRef]
- Bentsink, L.; Jowett, J.; Hanhart, C.J.; Koornneef, M. Cloning of DOG1, a Quantitative Trait Locus Controlling Seed Dormancy in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 17042–17047. [Google Scholar] [CrossRef] [Green Version]
- Graeber, K.; Linkies, A.; Müller, K.; Wunchova, A.; Rott, A.; Leubner-Metzger, G. Cross-Species Approaches to Seed Dormancy and Germination: Conservation and Biodiversity of ABA-Regulated Mechanisms and the Brassicaceae DOG1 Genes. Plant Mol. Biol. 2010, 73, 67–87. [Google Scholar] [CrossRef]
- Nonogaki, H. Seed Dormancy and Germination—Emerging Mechanisms and New Hypotheses. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Blanco, C.; Bentsink, L.; Hanhart, C.J.; Vries, H.B.; Koornneef, M. Analysis of Natural Allelic Variation at Seed Dormancy Loci of Arabidopsis Thaliana. Genetics 2003, 164, 711–729. [Google Scholar]
- Bentsink, L.; Hanson, J.; Hanhart, C.J.; Vries, H.B.; Coltrane, C.; Keizer, P.; El-Lithy, M.; Alonso-Blanco, C.; de Andrés, M.T.; Reymond, M.; et al. Natural Variation for Seed Dormancy in Arabidopsis Is Regulated by Additive Genetic and Molecular Pathways. PNAS 2010, 107, 4264–4269. [Google Scholar] [CrossRef] [Green Version]
- Chiang, G.C.K.; Barua, D.; Dittmar, E.; Kramer, E.M.; de Casas, R.R.; Donohue, K. Pleiotropy in the Wild: The Dormancy Gene Dog1 Exerts Cascading Control on Life Cycles. Evolution 2013, 67, 883–893. [Google Scholar] [CrossRef]
- Kronholm, I.; Picó, F.X.; Alonso-Blanco, C.; Goudet, J.; de Meaux, J. Genetic Basis of Adaptation in Arabidopsis Thaliana: Local Adaptation at the Seed Dormancy QTL DOG1. Evolution 2012, 66, 2287–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Footitt, S.; Clay, H.A.; Dent, K.; Finch-Savage, W.E. Environment Sensing in Spring-Dispersed Seeds of a Winter Annual Arabidopsis Influences the Regulation of Dormancy to Align Germination Potential with Seasonal Changes. New Phytol. 2014, 202, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Nakabayashi, K.; Bartsch, M.; Xiang, Y.; Miatton, E.; Pellengahr, S.; Yano, R.; Seo, M.; Soppe, W.J.J. The Time Required for Dormancy Release in Arabidopsis Is Determined by DELAY OF GERMINATION1 Protein Levels in Freshly Harvested Seeds[OA]. Plant Cell 2012, 24, 2826–2838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Yang, S.; Liu, X.; Wu, K. Arabidopsis Histone Demethylases LDL1 and LDL2 Control Primary Seed Dormancy by Regulating DELAY OF GERMINATION 1 and ABA Signaling-Related Genes. Front. Plant Sci. 2015, 6, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graeber, K.; Linkies, A.; Steinbrecher, T.; Mummenhoff, K.; Tarkowská, D.; Turečková, V.; Ignatz, M.; Sperber, K.; Voegele, A.; de Jong, H.; et al. DELAY OF GERMINATION 1 Mediates a Conserved Coat-Dormancy Mechanism for the Temperature- and Gibberellin-Dependent Control of Seed Germination. PNAS 2014, 111, E3571–E3580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barua, D.; Butler, C.; Tisdale, T.E.; Donohue, K. Natural Variation in Germination Responses of Arabidopsis to Seasonal Cues and Their Associated Physiological Mechanisms. Ann. Bot. 2012, 109, 209–226. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; MacGregor, D.R.; Dave, A.; Florance, H.; Moore, K.; Paszkiewicz, K.; Smirnoff, N.; Graham, I.A.; Penfield, S. Maternal Temperature History Activates Flowering Locus T in Fruits to Control Progeny Dormancy According to Time of Year. PNAS 2014, 111, 18787–18792. [Google Scholar] [CrossRef] [Green Version]
- Chiang, G.C.K.; Barua, D.; Kramer, E.M.; Amasino, R.M.; Donohue, K. Major Flowering Time Gene, FLOWERING LOCUS C, Regulates Seed Germination in Arabidopsis Thaliana. PNAS 2009, 106, 11661–11666. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Berdeja, A.; Stitzer, M.C.; Taylor, M.A.; Okada, M.; Ezcurra, E.; Runcie, D.E.; Schmitt, J. Functional Variants of DOG1 Control Seed Chilling Responses and Variation in Seasonal Life-History Strategies in Arabidopsis Thaliana. PNAS 2020, 117, 2526–2534. [Google Scholar] [CrossRef] [Green Version]
- Postma, F.M.; Ågren, J. Early Life Stages Contribute Strongly to Local Adaptation in Arabidopsis Thaliana. Proc. Natl. Acad. Sci. USA 2016, 113, 7590–7595. [Google Scholar] [CrossRef] [Green Version]
- Donohue, K.; Heschel, M.S.; Butler, C.M.; Barua, D.; Sharrock, R.A.; Whitelam, G.C.; Chiang, G.C.K. Diversification of Phytochrome Contributions to Germination as a Function of Seed-Maturation Environment. New Phytol. 2008, 177, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Kendall, S.L.; Hellwege, A.; Marriot, P.; Whalley, C.; Graham, I.A.; Penfield, S. Induction of Dormancy in Arabidopsis Summer Annuals Requires Parallel Regulation of DOG1 and Hormone Metabolism by Low Temperature and CBF Transcription Factors. Plant Cell 2011, 23, 2568–2580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Cai, M.; Long, Q.; Liu, L.; Lin, Q.; Jiang, L.; Chen, S.; Wan, J. OsLOX2, a Rice Type I Lipoxygenase, Confers Opposite Effects on Seed Germination and Longevity. Transgenic Res. 2014, 23, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Finch-Savage, W.E.; Footitt, S. Regulation of Seed Dormancy Cycling in Seasonal Field Environments. In Advances in Plant Dormancy; Springer: Cham, Switzerland, 2015; pp. 35–47. ISBN 978-3-319-14450-4. [Google Scholar]
- Murphey, M.; Kovach, K.; Elnacash, T.; He, H.; Bentsink, L.; Donohue, K. DOG1-Imposed Dormancy Mediates Germination Responses to Temperature Cues. Environ. Exp. Bot. 2015, 33–43. [Google Scholar] [CrossRef]
- Huo, H.; Wei, S.; Bradford, K.J. Delay of germination1 (DOG1) Regulates Both Seed Dormancy and Flowering Time through MicroRNA Pathways. Proc. Natl. Acad. Sci. USA 2016, 113, E2199–E2206. [Google Scholar] [CrossRef] [Green Version]
- Martel, C.; Blair, L.K.; Donohue, K. PHYD Prevents Secondary Dormancy Establishment of Seeds Exposed to High Temperature and Is Associated with Lower PIL5 Accumulation. J. Exp. Bot. 2018, 69, 3157–3169. [Google Scholar] [CrossRef]
- He, H.; Willems, L.A.J.; Batushansky, A.; Fait, A.; Hanson, J.; Nijveen, H.; Hilhorst, H.W.M.; Bentsink, L. Effects of Parental Temperature and Nitrate on Seed Performance Are Reflected by Partly Overlapping Genetic and Metabolic Pathways. Plant Cell Physiol. 2016, 57, 473–487. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Y.; Song, B.; Née, G.; Kramer, K.; Finkemeier, I.; Soppe, W.J.J. Sequence Polymorphisms at the REDUCED DORMANCY5 Pseudophosphatase Underlie Natural Variation in Arabidopsis Dormancy. Plant Physiol. 2016, 171, 2659–2670. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Y.; Nakabayashi, K.; Ding, J.; He, F.; Bentsink, L.; Soppe, W.J.J. REDUCED DORMANCY5 Encodes a Protein Phosphatase 2C That Is Required for Seed Dormancy in Arabidopsis. Plant Cell 2014, 26, 4362–4375. [Google Scholar] [CrossRef] [Green Version]
- Buijs, G.; Vogelzang, A.; Nijveen, H.; Bentsink, L. Dormancy Cycling: Translation-Related Transcripts Are the Main Difference between Dormant and Non-Dormant Seeds in the Field. Plant J. 2020, 102, 327–339. [Google Scholar] [CrossRef]
- Footitt, S.; Ölçer-Footitt, H.; Hambidge, A.J.; Finch-Savage, W.E. A Laboratory Simulation of Arabidopsis Seed Dormancy Cycling Provides New Insight into Its Regulation by Clock Genes and the Dormancy-Related Genes DOG1, MFT, CIPK23 and PHYA. Plant Cell Environ. 2017, 40, 1474–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, S.; Abe, F.; Kawahigashi, H.; Nakazono, K.; Tagiri, A.; Matsumoto, T.; Utsugi, S.; Ogawa, T.; Handa, H.; Ishida, H.; et al. A Wheat Homolog of MOTHER OF FT AND TFL1 Acts in the Regulation of Germination. Plant Cell 2011, 23, 3215–3229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chono, M.; Matsunaka, H.; Seki, M.; Fujita, M.; Kiribuchi-Otobe, C.; Oda, S.; Kojima, H.; Nakamura, S. Molecular and Genealogical Analysis of Grain Dormancy in Japanese Wheat Varieties, with Specific Focus on MOTHER OF FT AND TFL1 on Chromosome 3A. Breed. Sci. 2015, 65, 103–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penfield, S.; Josse, E.; Kannangara, R.; Gilday, A.; Halliday, K.; Graham, I. Cold and Light Control Seed Germination through the BHLH Transcription Factor SPATULA. Curr. Biol. 2005, 15, 1998–2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaistij, F.E.; Gan, Y.; Penfield, S.; Gilday, A.D.; Dave, A.; He, Z.; Josse, E.-M.; Choi, G.; Halliday, K.J.; Graham, I.A. Differential Control of Seed Primary Dormancy in Arabidopsis Ecotypes by the Transcription Factor SPATULA. Proc. Natl. Acad. Sci. USA 2013, 110, 10866–10871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dave, A.; Vaistij, F.E.; Gilday, A.D.; Penfield, S.D.; Graham, I.A. Regulation of Arabidopsis Thaliana Seed Dormancy and Germination by 12-Oxo-Phytodienoic Acid. J. Exp. Bot. 2016, 67, 2277–2284. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Dong, A.; Shen, W.-H. Histone Variants and Chromatin Assembly in Plant Abiotic Stress Responses. Biochim. Biophys. Acta 2013, 1819, 343–348. [Google Scholar] [CrossRef]
- Probst, A.V.; Mittelsten Scheid, O. Stress-Induced Structural Changes in Plant Chromatin. Curr. Opin. Plant Biol. 2015, 27, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-M.; Sasaki, T.; Ueda, M.; Sako, K.; Seki, M. Chromatin Changes in Response to Drought, Salinity, Heat, and Cold Stresses in Plants. Front. Plant Sci. 2015, 6, 114. [Google Scholar] [CrossRef] [Green Version]
- Whittle, C.A.; Otto, S.P.; Johnston, M.O.; Krochko, J.E. Adaptive Epigenetic Memory of Ancestral Temperature Regime in Arabidopsis Thaliana. Botany 2009, 87, 650–657. [Google Scholar] [CrossRef] [Green Version]
- Angel, A.; Song, J.; Dean, C.; Howard, M. A Polycomb-Based Switch Underlying Quantitative Epigenetic Memory. Nature 2011, 476, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Barski, A.; Cuddapah, S.; Cui, K.; Roh, T.-Y.; Schones, D.E.; Wang, Z.; Wei, G.; Chepelev, I.; Zhao, K. High-Resolution Profiling of Histone Methylations in the Human Genome. Cell 2007, 129, 823–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soria, G.; Polo, S.E.; Almouzni, G. Prime, Repair, Restore: The Active Role of Chromatin in the DNA Damage Response. Mol. Cell 2012, 46, 722–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, A.; Yin, H.; Meng, Q.; Yu, X.; Huang, S.; Wang, J.; Ahmad, R.; Liu, B.; Xu, Z.-Y. Trithorax-Group Proteins ARABIDOPSIS TRITHORAX4 (ATX4) and ATX5 Function in Abscisic Acid and Dehydration Stress Responses. New Phytol. 2018, 217, 1582–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolny, E.; Braszewska-Zalewska, A.; Kroczek, D.; Hasterok, R. Histone H3 and H4 Acetylation Patterns Are More Dynamic than Those of DNA Methylation in Brachypodium Distachyon Embryos during Seed Maturation and Germination. Protoplasma 2017, 254, 2045–2052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Footitt, S.; Müller, K.; Kermode, A.R.; Finch-Savage, W.E. Seed Dormancy Cycling in Arabidopsis: Chromatin Remodelling and Regulation of DOG1 in Response to Seasonal Environmental Signals. Plant J. 2015, 81, 413–425. [Google Scholar] [CrossRef]
- Li, H.-C.; Chuang, K.; Henderson, J.T.; Rider, S.D.; Bai, Y.; Zhang, H.; Fountain, M.; Gerber, J.; Ogas, J. PICKLE Acts during Germination to Repress Expression of Embryonic Traits. Plant J. 2005, 44, 1010–1022. [Google Scholar] [CrossRef]
- Zhang, H.; Rider, S.D.; Henderson, J.T.; Fountain, M.; Chuang, K.; Kandachar, V.; Simons, A.; Edenberg, H.J.; Romero-Severson, J.; Muir, W.M.; et al. The CHD3 Remodeler PICKLE Promotes Trimethylation of Histone H3 Lysine 27. J. Biol. Chem. 2008, 283, 22637–22648. [Google Scholar] [CrossRef] [Green Version]
- Tai, H.H.; Tai, G.C.C.; Beardmore, T. Dynamic Histone Acetylation of Late Embryonic Genes during Seed Germination. Plant Mol. Biol. 2005, 59, 909–925. [Google Scholar] [CrossRef]
- Tanaka, M.; Kikuchi, A.; Kamada, H. The Arabidopsis Histone Deacetylases HDA6 and HDA19 Contribute to the Repression of Embryonic Properties after Germination. Plant Physiol. 2008, 146, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Sura, W.; Kabza, M.; Karlowski, W.M.; Bieluszewski, T.; Kuś-Slowinska, M.; Pawełoszek, Ł.; Sadowski, J.; Ziolkowski, P.A. Dual Role of the Histone Variant H2A.Z in Transcriptional Regulation of Stress-Response Genes. Plant Cell 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, N.; Tsuchiya, W.; Moresco, J.J.; Hayashi, Y.; Satoh, K.; Kaiwa, N.; Irisa, T.; Kinoshita, T.; Schroeder, J.I.; Yates, J.R.; et al. Control of Seed Dormancy and Germination by DOG1-AHG1 PP2C Phosphatase Complex via Binding to Heme. Nat. Commun. 2018, 9, 2132. [Google Scholar] [CrossRef]
- Yatusevich, R.; Fedak, H.; Ciesielski, A.; Krzyczmonik, K.; Kulik, A.; Dobrowolska, G.; Swiezewski, S. Antisense Transcription Represses Arabidopsis Seed Dormancy QTL DOG1 to Regulate Drought Tolerance. EMBO Rep. 2017, 18, 2186–2196. [Google Scholar] [CrossRef] [PubMed]
- Arc, E.; Galland, M.; Godin, B.; Cueff, G.; Rajjou, L. Nitric Oxide Implication in the Control of Seed Dormancy and Germination. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkelstein, R.; Reeves, W.; Ariizumi, T.; Steber, C. Molecular Aspects of Seed Dormancy. Annu. Rev. Plant. Biol. 2008, 59, 387–415. [Google Scholar] [CrossRef] [Green Version]
- Linkies, A.; Graeber, K.; Knight, C.; Leubner-Metzger, G. The Evolution of Seeds. New Phytol. 2010, 186, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Simons, A.M. Fluctuating Natural Selection Accounts for the Evolution of Diversification Bet Hedging. Proc. R. Soc. Lond. B Biol. Sci. 2009, 276, 1987–1992. [Google Scholar] [CrossRef] [Green Version]
- Intergovernmental Panel on Climate Change. Climate Change 2013—The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014; ISBN 978-1-107-41532-4. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klupczyńska, E.A.; Pawłowski, T.A. Regulation of Seed Dormancy and Germination Mechanisms in a Changing Environment. Int. J. Mol. Sci. 2021, 22, 1357. https://doi.org/10.3390/ijms22031357
Klupczyńska EA, Pawłowski TA. Regulation of Seed Dormancy and Germination Mechanisms in a Changing Environment. International Journal of Molecular Sciences. 2021; 22(3):1357. https://doi.org/10.3390/ijms22031357
Chicago/Turabian StyleKlupczyńska, Ewelina A., and Tomasz A. Pawłowski. 2021. "Regulation of Seed Dormancy and Germination Mechanisms in a Changing Environment" International Journal of Molecular Sciences 22, no. 3: 1357. https://doi.org/10.3390/ijms22031357
APA StyleKlupczyńska, E. A., & Pawłowski, T. A. (2021). Regulation of Seed Dormancy and Germination Mechanisms in a Changing Environment. International Journal of Molecular Sciences, 22(3), 1357. https://doi.org/10.3390/ijms22031357