
Function of Connexin-43 in Macrophages


{kind=link}
Abstract
:1. Introduction
2. Dawdling and Devouring: Rolling around for Initiative
2.1. Migration
2.2. Phagocytosis
2.3. Antigen-Presentation
3. Intercellular Communication Is a Two-Way Street
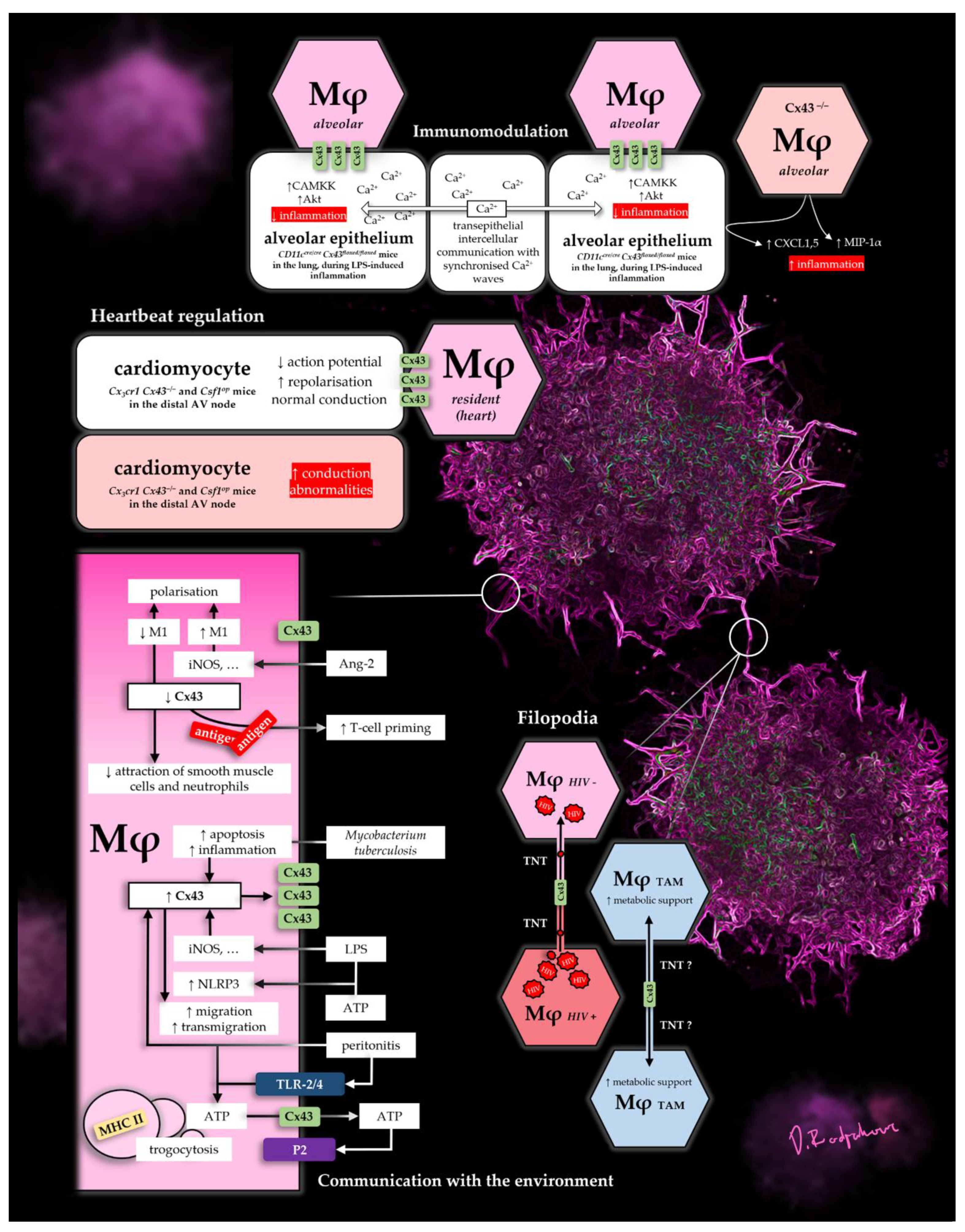
3.1. Physiological Communication in Heart, Lung and Intestine
3.1.1. Electrochemical Communication of the Heart
3.1.2. Immunomodulatory Communication in the Lungs
3.1.3. Intercellular Communication in the Intestine
3.2. Pathological Communication in IBD, Tumors and HIV
3.2.1. IBD
3.2.2. Filopodia in Tumor Networks and HIV
4. A Microenvironment with Macro-Consequences
4.1. The Environment Changes the Mφ: LPS, Acute Peritonitis and Sepsis
4.2. The Mφ Changes the Environment: ATP Release
5. Mφs Dress for the Job They Want
5.1. Adapting to Their Environment: Polarisation
- M1 Mφs accumulate at the inflammation site by strong adhesion, which promotes cell retention and the progression of inflammation [62];
5.2. Adapting to Their Environment: Differentiation
5.2.1. Foam Cells
5.2.2. Foreign Body Giant Cells
6. Conclusions
Funding
Conflicts of Interest
Abbreviations
| Ang-2 | Angiotensin-2 |
| ATP | Adenosine triphosphate |
| Cx43 | Connexin-43 |
| DC | Dendritic cell |
| FBGC | Foreign body giant cell |
| IBD | Inflammatory bowel disease |
| Mφ | Macrophage |
| TAM | Tumor-associated-macrophage |
| TNT | Tunneling-nanotube |
References
- Mečnikov, É. Leçons sur la Pathologie Comparée de L’inflammation; Bibliothèque des Annales de L’Institut Pasteur: Paris, France, 1892. [Google Scholar]
- Evans, H.M.; Bowman, F.B.; Winternitz, M.C. An experimental study of the histogenesis of the miliary tubercle in vitally stained rabbits. J. Exp. Med. 1914, 19, 283–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cecil, R.L. A study of experimental non-hemolytic streptococcus lesions in vitally stained rabbits. J. Exp. Med. 1916, 24, 739–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynn, T.A.; Vannella, K.M. Macrophages in Tissue Repair, Regeneration, and Fibrosis. Immunity 2016, 44, 450–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, L.C.; Jenkins, S.J.; Allen, J.E.; Taylor, P.R. Tissue-resident macrophages. Nat. Immunol. 2013, 14, 986–995. [Google Scholar]
- Bang, F.B.; Warwick, A. Macrophages and mouse hepatitis. Virology 1959, 9, 715–717. [Google Scholar] [CrossRef]
- Evans, R.; Alexander, P. Mechanism of immunologically specific killing of tumour cells by macrophages. Nature 1972, 236, 168–170. [Google Scholar] [CrossRef]
- Polacek, D.; Bech, F.; McKinsey, J.F.; Davies, P.F. Connexin43 gene expression in the rabbit arterial wall: Effects of hypercholesterolemia, balloon injury and their combination. J. Vasc. Res. 1997, 34, 19–30. [Google Scholar] [CrossRef]
- Dosch, M.; Zindel, J.; Jebbawi, F.; Melin, N.; Sanchez-Taltavull, D.; Stroka, D.; Candinas, D.; Beldi, G. Connexin-43-dependent ATP release mediates macrophage activation during sepsis. elife 2019, 8, e42670. [Google Scholar] [CrossRef]
- Hopper, K.E.; Wood, P.R.; Nelson, D.S. Macrophage heterogeneity. Vox Sang. 1979, 36, 257–274. [Google Scholar] [CrossRef]
- Laird, D.W.; Lampe, P.D. Therapeutic strategies targeting connexins. Nat. Rev. Drug Discov. 2018, 17, 905–921. [Google Scholar] [CrossRef]
- Dbouk, H.A.; Mroue, R.M.; El-Sabban, M.E.; Talhouk, R.S. Connexins: A myriad of functions extending beyond assembly of gap junction channels. Cell Commun. Signal. 2009, 12, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro-Rodrigues, T.M.; Martins-Marques, T.; Morel, S.; Kwak, B.R.; Girão, H. Role of connexin 43 in different forms of intercellular communication—Gap junctions, extracellular vesicles and tunnelling nanotubes. J. Cell. Sci. 2017, 130, 3619–3630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stout, C.; Goodenough, D.A.; Paul, D.L. Connexins: Functions without junctions. Curr. Opin. Cell Biol. 2004, 16, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Pfenniger, A.; Wohlwend, A.; Kwak, B.R. Mutations in connexin genes and disease. Eur. J. Clin. Investig. 2011, 41, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Reaume, A.G.; de Sousa, P.A.; Kulkarni, S.; Langille, B.L.; Zhu, D.; Davies, T.C.; Juneja, S.C.; Kidder, G.M.; Rossant, J. Cardiac malformation in neonatal mice lacking connexin43. Science 1995, 267, 1831–1834. [Google Scholar] [CrossRef]
- Tittarelli, A.; Navarrete, M.; Gleisner, M.A.; Gebicke-Haerter, P. Connexin-Mediated Signaling at the Immunological Synapse. Int. J. Mol. Sci. 2020, 21, 3736. [Google Scholar] [CrossRef]
- Westphalen, K.; Gusarova, G.A.; Islam, M.N.; Subramanian, M.; Cohen, T.S.; Prince, A.S.; Bhattacharya, J. Sessile alveolar macrophages communicate with alveolar epithelium to modulate immunity. Nature 2014, 506, 503–506. [Google Scholar] [CrossRef] [Green Version]
- Beckmann, A.; Grissmer, A.; Meier, C.; Tschernig, T. Intercellular communication between alveolar epithelial cells and macrophages. Ann. Anat. 2020, 227, 151417. [Google Scholar] [CrossRef]
- Hulsmans, M.; Clauss, S.; Xiao, L.; Aguirre, A.D.; King, K.R.; Hanley, A.; Nahrendorf, M. Macrophages Facilitate Electrical Conduction in the Heart. Cell 2017, 169, 510–522.e20. [Google Scholar] [CrossRef] [Green Version]
- Eugenín, E.A.; Brañes, M.C.; Berman, J.W.; Sáez, J.C. TNF-alpha plus IFN-gamma induce connexin43 expression and formation of gap junctions between human monocytes/macrophages that enhance physiological responses. J. Immunol. 2003, 170, 1320–1328. [Google Scholar] [CrossRef] [Green Version]
- Qin, J.; Zhang, G.; Zhang, X.; Tan, B.; Lv, Z.; Liu, M.; Ren, H.; Qian, M.; Du, B. TLR-Activated Gap Junction Channels Protect Mice against Bacterial Infection through Extracellular UDP Release. J. Immunol. 2016, 196, 1790–1798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutstein, D.E.; Morley, G.E.; Tamaddon, H.; Vaidya, D.; Schneider, M.D.; Chen, J.; Chien, K.R.; Stuhlmann, H.; Fishman, G.I. Conduction slowing and sudden arrhythmic death in mice with cardiac-restricted inactivation of connexin43. Circ. Res. 2001, 88, 333–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chadjichristos, C.E.; Matter, C.M.; Roth, I.; Sutter, E.; Pelli, G.; Lüscher, T.F.; Chanson, M.; Kwak, B.R. Reduced connexin43 expression limits neointima formation after balloon distension injury in hypercholesterolemic mice. Circulation 2006, 113, 2835–2843. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Chen, J.H.; Lee, Y.; Hassan, M.M.; Kim, S.J.; Choi, E.Y.; Hong, S.T.; Park, B.H.; Park, J.H. mTOR- and SGK-Mediated Connexin 43 Expression Participates in Lipopolysaccharide-Stimulated Macrophage Migration through the iNOS/Src/FAK Axis. J. Immunol. 2018, 201, 2986–2997. [Google Scholar] [CrossRef] [PubMed]
- Glass, A.M.; Wolf, B.J.; Schneider, K.M.; Princiotta, M.F.; Taffet, S.M. Connexin43 is dispensable for phagocytosis. J. Immunol. 2014, 190, 4830–4835. [Google Scholar] [CrossRef] [Green Version]
- Oviedo-Orta, E.; Errington, R.J.; Evans, W.H. Gap junction intercellular communication during lymphocyte transendothelial migration. Cell Biol. Int. 2002, 26, 253–263. [Google Scholar] [CrossRef]
- Morel, S.; Chanson, M.; Nguyen, T.D.; Glass, A.M.; Richani Sarieddine, M.Z.; Meens, M.J.; Burnier, L.; Kwak, B.R.; Taffet, S.M. Titration of the gap junction protein Connexin43 reduces atherogenesis. Thromb. Haemost. 2014, 112, 390–401. [Google Scholar] [CrossRef]
- Huang, Y.; Mao, Z.; Zhang, Z.; Obata, F.; Yang, X.; Zhang, X.; Huang, Y.; Mitsui, T.; Fan, J.; Takeda, M.; et al. Connexin43 Contributes to Inflammasome Activation and Lipopolysaccharide-Initiated Acute Renal Injury via Modulation of Intracellular Oxidative Status. Antioxid. Redox Signal. 2019, 31, 1194–1212. [Google Scholar] [CrossRef]
- Anand, R.J.; Dai, S.; Gribar, S.C.; Richardson, W.; Kohler, J.W.; Hoffman, R.A.; Hackam, D.J. A role for connexin43 in macrophage phagocytosis and host survival after bacterial peritoneal infection. J. Immunol. 2008, 181, 8534–8543. [Google Scholar] [CrossRef] [Green Version]
- Morris, L.; Crocker, P.R.; Gordon, S. Murine fetal liver macrophages bind developing erythroblasts by a divalent cation-dependent hemagglutinin. J. Cell Biol. 1988, 106, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Austyn, J.M.; Gordon, S. F4/80, a monoclonal antibody directed specifically against the mouse macrophage. Eur. J. Immunol. 1981, 11, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Mazzini, E.; Massimiliano, L.; Penna, G.; Rescigno, M. Oral tolerance can be established via gap junction transfer of fed antigens from CX3CR1+ macrophages to CD103+ dendritic cells. Immunity 2014, 40, 248–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joly, E.; Hudrisier, D. What is trogocytosis and what is its purpose? Nat. Immunol. 2003, 4, 815. [Google Scholar] [CrossRef]
- Smyth, L.A.; Hervouet, C.; Hayday, T.; Becker, P.D.; Ellis, R.; Lechler, R.I.; Lombardi, G.; Klavinskis, L.S. Acquisition of MHC:peptide complexes by dendritic cells contributes to the generation of antiviral CD8+ T cell immunity in vivo. J. Immunol. 2012, 189, 2274–2282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakim, L.M.; Bevan, M.J. Cross-dressed dendritic cells drive memory CD8+ T-cell activation after viral infection. Nature 2011, 471, 629–632. [Google Scholar] [CrossRef] [Green Version]
- Okafo, G.; Prevedel, L.; Eugenin, E. Tunneling nanotubes (TNT) mediate long-range gap junctional communication: Implications for HIV cell to cell spread. Sci. Rep. 2017, 7, 16660. [Google Scholar] [CrossRef]
- De Couto, G. Macrophages in cardiac repair: Environmental cues and therapeutic strategies. Exp. Mol. Med. 2019, 51, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rosenthal, N. A Guardian of the Heartbeat. N. Engl. J. Med. 2017, 377, 84–86. [Google Scholar] [CrossRef] [Green Version]
- Manning, B.D.; Cantley, L.C. AKT/PKB signaling: Navigating downstream. Cell 2007, 129, 1261–1274. [Google Scholar] [CrossRef] [Green Version]
- Al-Ghadban, S.; Kaissi, S.; Homaidan, F.R.; Naim, H.Y.; El-Sabban, M.E. Cross-talk between intestinal epithelial cells and immune cells in inflammatory bowel disease. Sci. Rep. 2016, 6, 29783. [Google Scholar] [CrossRef] [Green Version]
- Caillou, B.; Talbot, M.; Weyemi, U.; Pioche-Durieu, C.; Al Ghuzlan, A.; Bidart, J.M.; Chouaib, S.; Schlumberger, M.; Dupuy, C. Tumor-associated macrophages (TAMs) form an interconnected cellular supportive network in anaplastic thyroid carcinoma. PLoS ONE 2011, 6, e22567. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Fajardo, A.; Ylihärsilä, M.; Evans, W.H.; Newby, A.C.; Oviedo-Orta, E. CD4+ T lymphocyte subsets express connexin 43 and establish gap junction channel communication with macrophages in vitro. J. Leukoc. Biol. 2007, 82, 608–612. [Google Scholar] [CrossRef] [PubMed]
- Eugenín, E.A.; González, H.E.; Sánchez, H.A.; Brañes, M.C.; Sáez, J.C. Inflammatory conditions induce gap junctional communication between rat Kupffer cells both in vivo and in vitro. Cell. Immunol. 2007, 247, 103–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Wang, X.M.; Yang, P.; Han, L.; Wang, Y.Z.; Zheng, Z.H.; Wu, F.; Zhang, W.J.; Zhang, L. Effect of gap junctions on RAW264.7 macrophages infected with H37Rv. Medicine (Baltimore) 2018, 97, e12125. [Google Scholar] [CrossRef]
- Zumerle, S.; Calì, B.; Munari, F.; Angioni, R.; Di Virgilio, F.; Molon, B.; Viola, A. Intercellular Calcium Signaling Induced by ATP Potentiates Macrophage Phagocytosis. Cell Rep. 2019, 27, 1–10.e4. [Google Scholar] [CrossRef] [Green Version]
- Junger, W.G. Immune cell regulation by autocrine purinergic signalling. Nat. Rev. Immunol. 2011, 11, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Dosch, M.; Gerber, J.; Jebbawi, F.; Beldi, G. Mechanisms of ATP Release by Inflammatory Cells. Int. J. Mol. Sci. 2018, 19, 1222. [Google Scholar] [CrossRef] [Green Version]
- Corriden, R.; Insel, P.A. Basal release of ATP: An autocrine-paracrine mechanism for cell regulation. Sci. Signal. 2010, 3, re1. [Google Scholar] [CrossRef] [Green Version]
- Fortes, F.S.; Pecora, I.L.; Persechini, P.M.; Hurtado, S.; Costa, V.; Coutinho-Silva, R.; Goldenberg, R.C. Modulation of intercellular communication in macrophages: Possible interactions between GAP junctions and P2 receptors. J. Cell Sci. 2004, 117, 4717–4726. [Google Scholar] [CrossRef] [Green Version]
- Csóka, B.; Németh, Z.H.; Törő, G.; Idzko, M.; Zech, A.; Koscsó, B.; Spolarics, Z.; Antonioli, L.; Cseri, K.; Erdélyi, K.; et al. Extracellular ATP protects against sepsis through macrophage P2X7 purinergic receptors by enhancing intracellular bacterial killing. FASEB J. 2015, 29, 3626–3637. [Google Scholar] [CrossRef] [Green Version]
- Greve, A.S.; Skals, M.; Fagerberg, S.K.; Tonnus, W.; Ellermann-Eriksen, S.; Evans, R.J.; Linkermann, A.; Praetorius, H.A. P2X1, P2X4, and P2X7 Receptor Knock Out Mice Expose Differential Outcome of Sepsis Induced by α-Haemolysin Producing Escherichia coli. Front. Cell. Infect. Microbiol. 2017, 7, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecut, C.; Faccinetto, C.; Delierneux, C.; van Oerle, R.; Spronk, H.M.; Evans, R.J.; El Benna, J.; Bours, V.; Oury, C. ATP-gated P2X1 ion channels protect against endotoxemia by dampening neutrophil activation. J. Thromb. Haemost. 2012, 10, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Maître, B.; Magnenat, S.; Heim, V.; Ravanat, C.; Evans, R.J.; de la Salle, H.; Gachet, C.; Hechler, B. The P2X1 receptor is required for neutrophil extravasation during lipopolysaccharide-induced lethal endotoxemia in mice. J. Immunol. 2015, 194, 739–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, D.A.; Song, Z.; Stevens, W.; Sladek, C.D. Sustained stimulation of vasopressin and oxytocin release by ATP and phenylephrine requires recruitment of desensitization-resistant P2X purinergic receptors. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R940–R949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Rodríguez, R.; Yarova, P.; Winter, P.; Dora, K.A. Desensitization of endothelial P2Y1 receptors by PKC-dependent mechanisms in pressurized rat small mesenteric arteries. Br. J. Pharmacol. 2009, 158, 1609–1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.; Kang, N.; Lovatt, D.; Torres, A.; Zhao, Z.; Lin, J.; Nedergaard, M. Connexin 43 hemichannels are permeable to ATP. J. Neurosci. 2008, 28, 4702–4711. [Google Scholar] [CrossRef]
- Li, W.; Bao, G.; Chen, W.; Qiang, X.; Zhu, S.; Wang, S.; He, M.; Ma, G.; Ochani, M.; Al-Abed, Y.; et al. Connexin 43 Hemichannel as a Novel Mediator of Sterile and Infectious Inflammatory Diseases. Sci. Rep. 2018, 8, 166. [Google Scholar] [CrossRef]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Invest. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Cui, K.; Ardell, C.L.; Podolnikova, N.P.; Yakubenko, V.P. Distinct Migratory Properties of M1, M2, and Resident Macrophages Are Regulated by αDβ2 and αMβ2 Integrin-Mediated Adhesion. Front. Immunol. 2018, 9, 2650. [Google Scholar] [CrossRef] [PubMed]
- Novak, M.L.; Koh, T.J. Macrophage phenotypes during tissue repair. J. Leukoc. Biol. 2013, 93, 875–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Chen, K.; Xiao, J.; Xin, J.; Zhang, L.; Li, X.; Li, L.; Si, J.; Wang, L.; Ma, K. Angiotensin II induces RAW264.7 macrophage polarization to the M1-type through the connexin 43/NF-κB pathway. Mol. Med. Rep. 2020, 21, 2103–2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dandona, P.; Dhindsa, S.; Ghanim, H.; Chaudhuri, A. Angiotensin II and inflammation: The effect of angiotensin-converting enzyme inhibition and angiotensin II receptor blockade. Ann. NY Acad. Sci. 2007, 21, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS−) vs. Alternatively Activated Macrophages. Front. Immunol. 2020, 25, 234. [Google Scholar] [CrossRef]
- Yu, X.H.; Fu, Y.C.; Zhang, D.W.; Yin, K.; Tang, C.K. Foam cells in atherosclerosis. Clin. Chim. Acta 2013, 424, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Herde, K.; Hartmann, S.; Brehm, R.; Kilian, O.; Heiss, C.; Hild, A.; Alt, V.; Bergmann, M.; Schnettler, R.; Wenisch, S. Connexin 43 expression of foreign body giant cells after implantation of nanoparticulate hydroxyapatite. Biomaterials 2007, 28, 4912–4921. [Google Scholar] [CrossRef]
- Wenisch, S.; Cavalcanti-Adam, E.A.; Tryankowski, E.; Raabe, O.; Kilian, O.; Heiss, C.; Alt, V.; Arnhold, S.; Schnettler, R. Light- and transmission-electron-microscopic investigations on distribution of CD44, connexin 43 and actin cytoskeleton during the foreign body reaction to a nanoparticular hydroxyapatite in mini-pigs. Acta Biomater. 2012, 8, 2807–2814. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodjakovic, D.; Salm, L.; Beldi, G. Function of Connexin-43 in Macrophages. Int. J. Mol. Sci. 2021, 22, 1412. https://doi.org/10.3390/ijms22031412
Rodjakovic D, Salm L, Beldi G. Function of Connexin-43 in Macrophages. International Journal of Molecular Sciences. 2021; 22(3):1412. https://doi.org/10.3390/ijms22031412
Chicago/Turabian StyleRodjakovic, Daniel, Lilian Salm, and Guido Beldi. 2021. "Function of Connexin-43 in Macrophages" International Journal of Molecular Sciences 22, no. 3: 1412. https://doi.org/10.3390/ijms22031412
APA StyleRodjakovic, D., Salm, L., & Beldi, G. (2021). Function of Connexin-43 in Macrophages. International Journal of Molecular Sciences, 22(3), 1412. https://doi.org/10.3390/ijms22031412