
CLytA-DAAO Chimeric Enzyme Bound to Magnetic Nanoparticles. A New Therapeutical Approach for Cancer Patients?

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
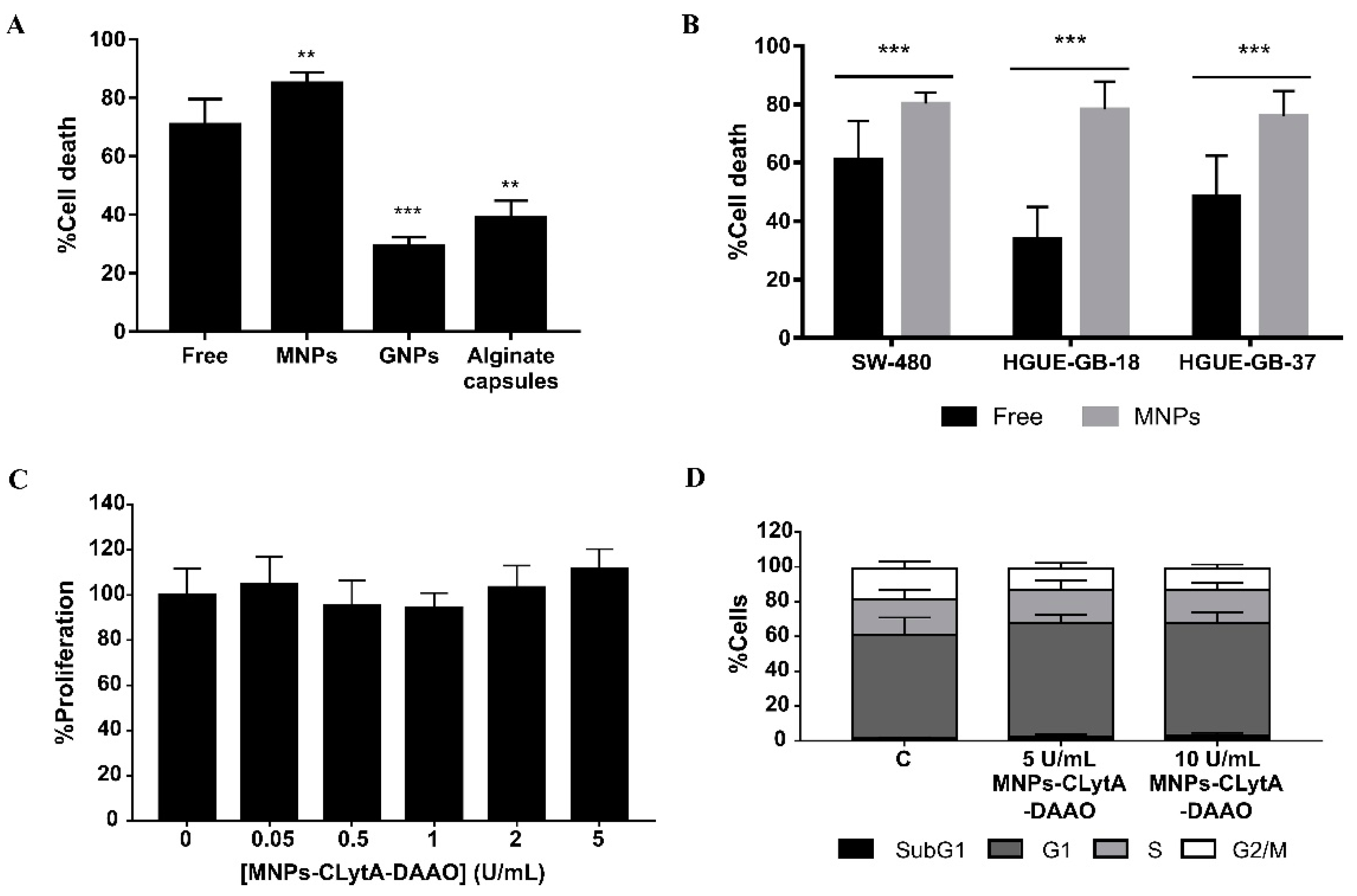
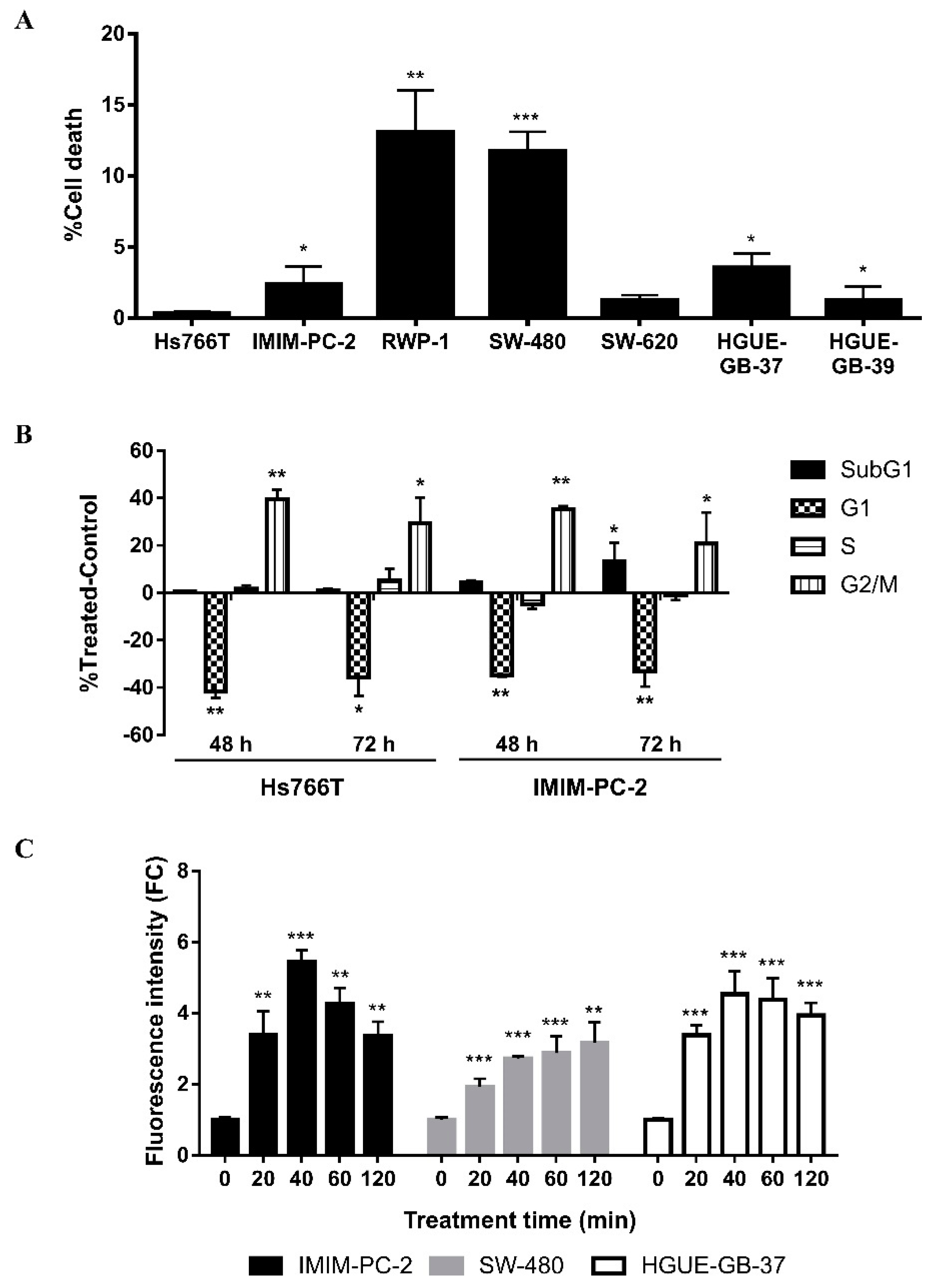
2.1. The Cytotoxic Effect of CLytA-DAAO Is Higher When It Is Bound to Magnetic Nanoparticles than When It Is Immobilized in Gold Nanoparticles or Alginate Capsules
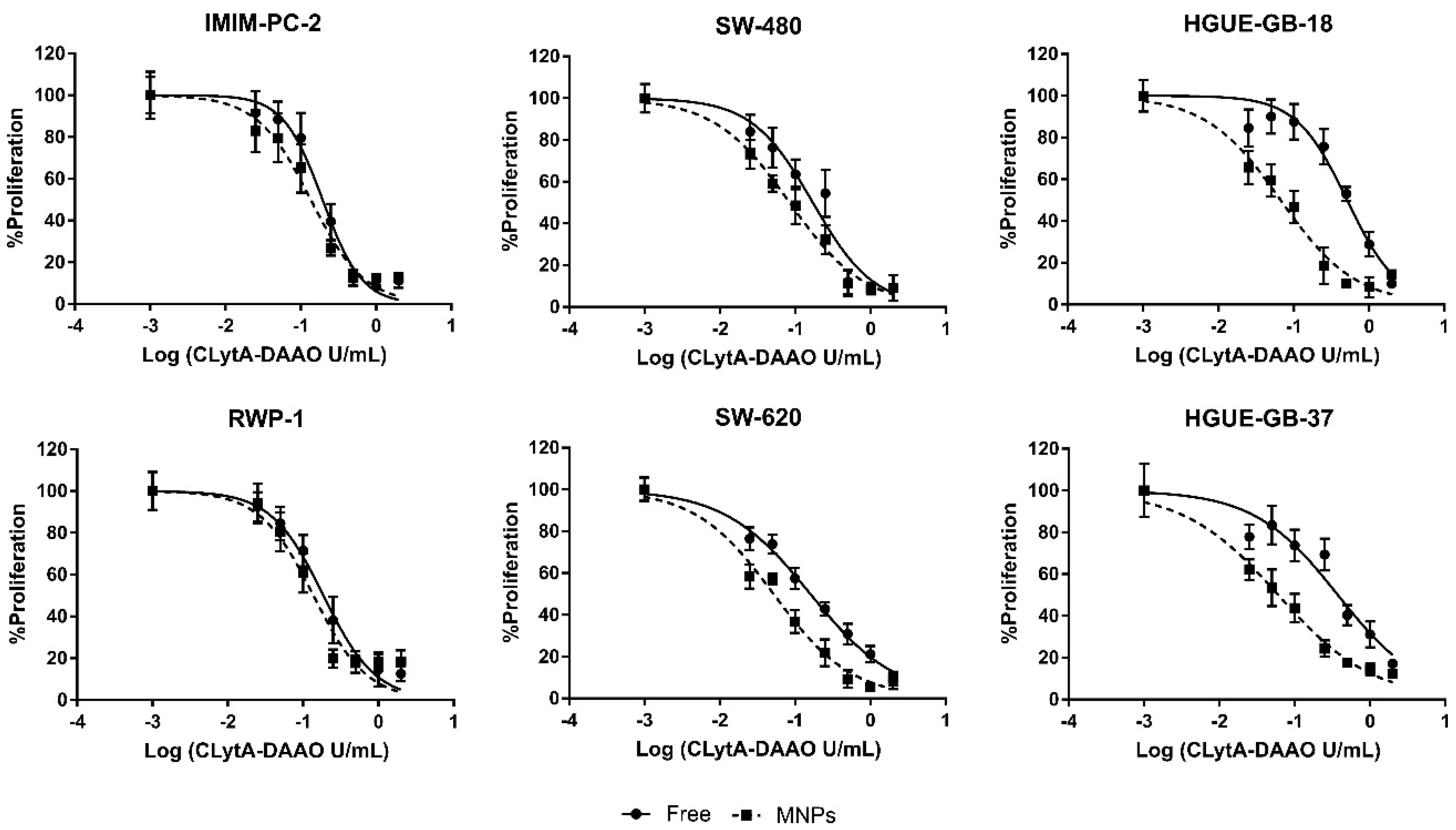
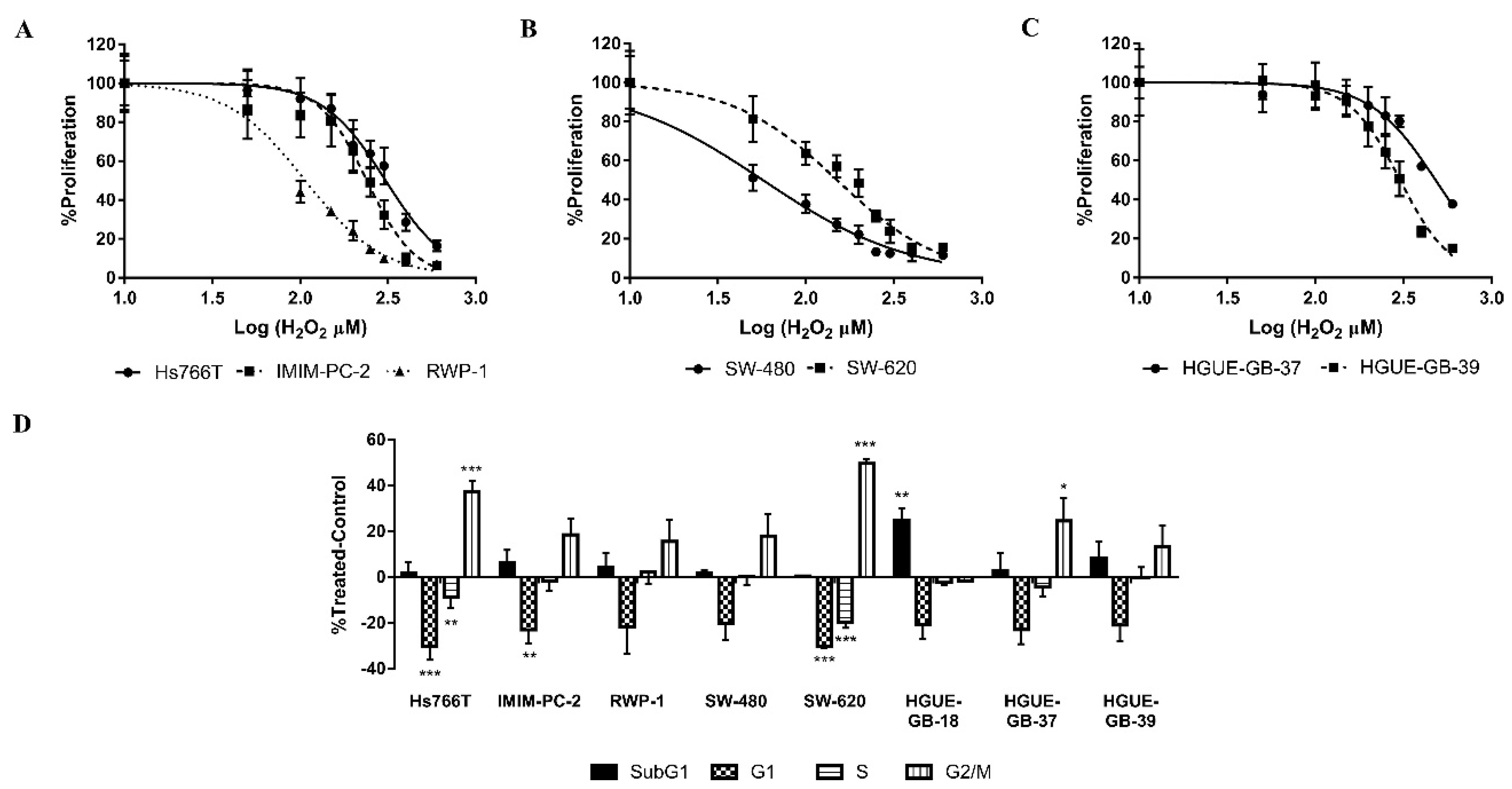
2.2. The IC50 Value for the Enzyme Immobilized in MNPs Is Lower than for the Free Enzyme
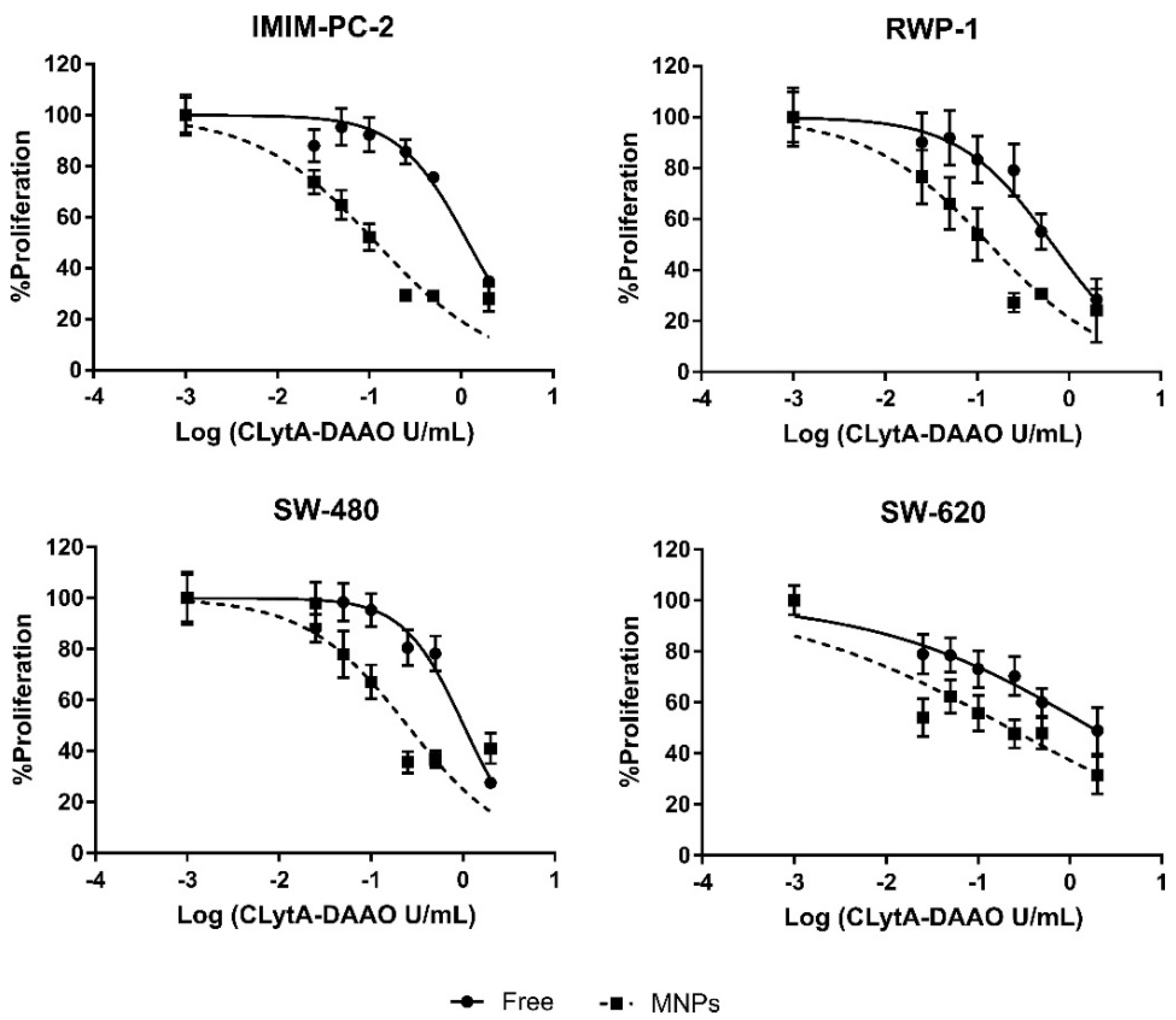
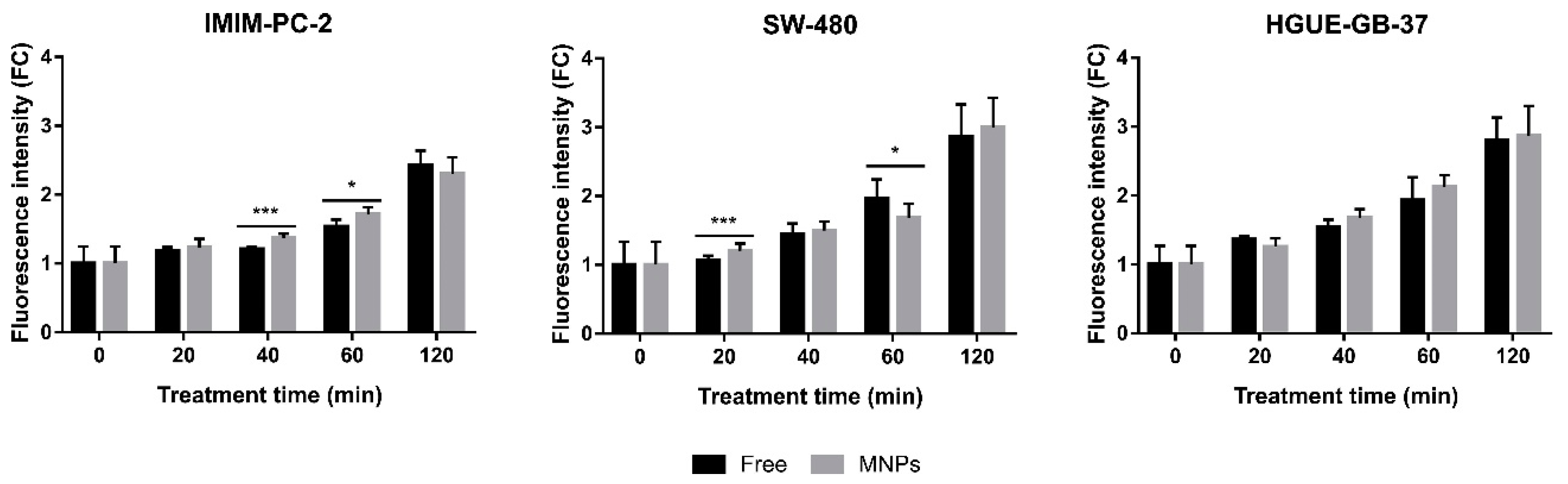
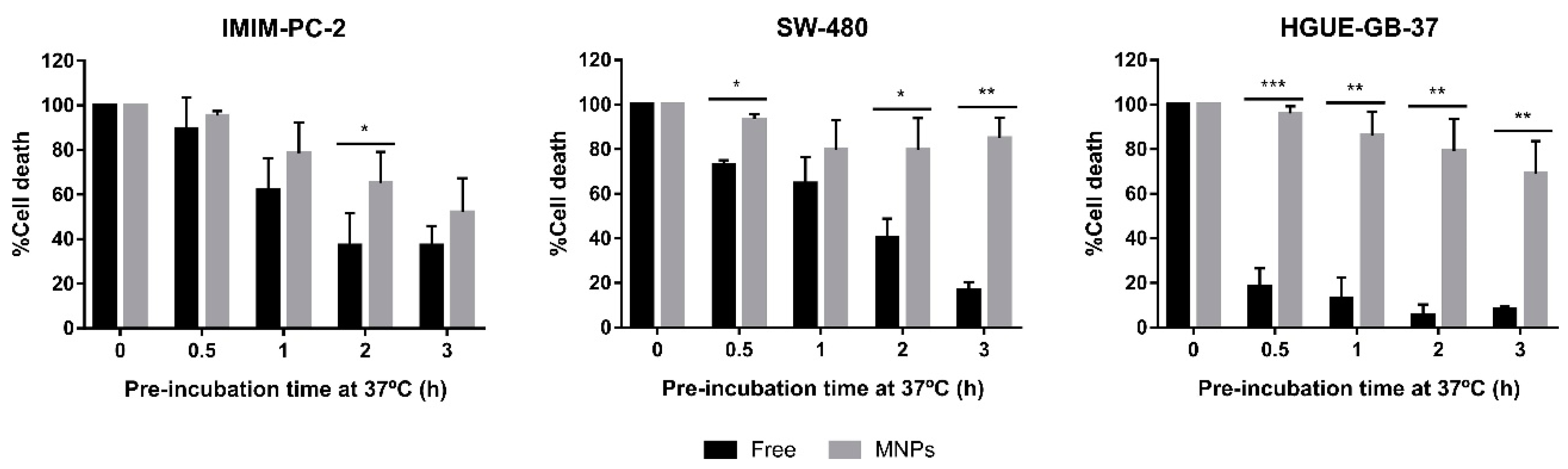
2.3. The Greater Effect Induced by the Treatment with CLytA-DAAO Bound to MNPs Is Due to an Increase in the Enzyme Stability
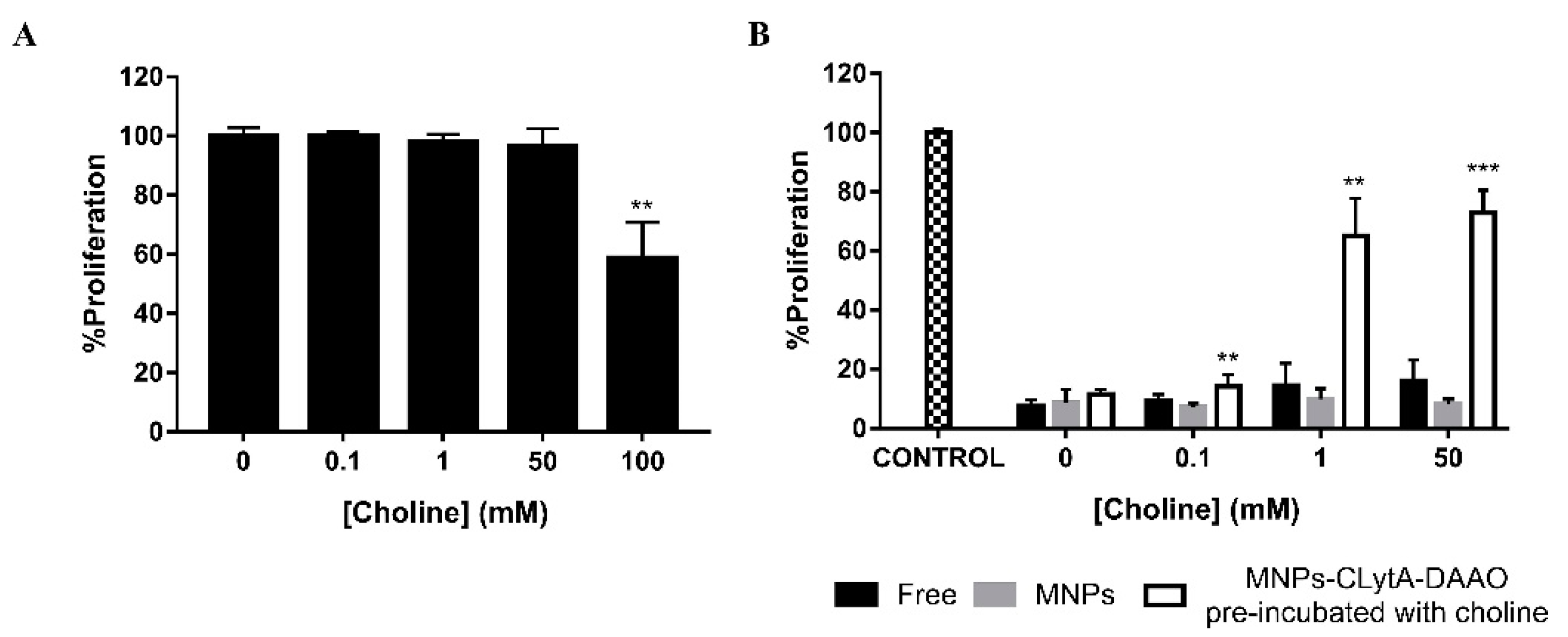
2.4. CLytA-DAAO Can Be Released from MNPs Through the External Addition of Choline
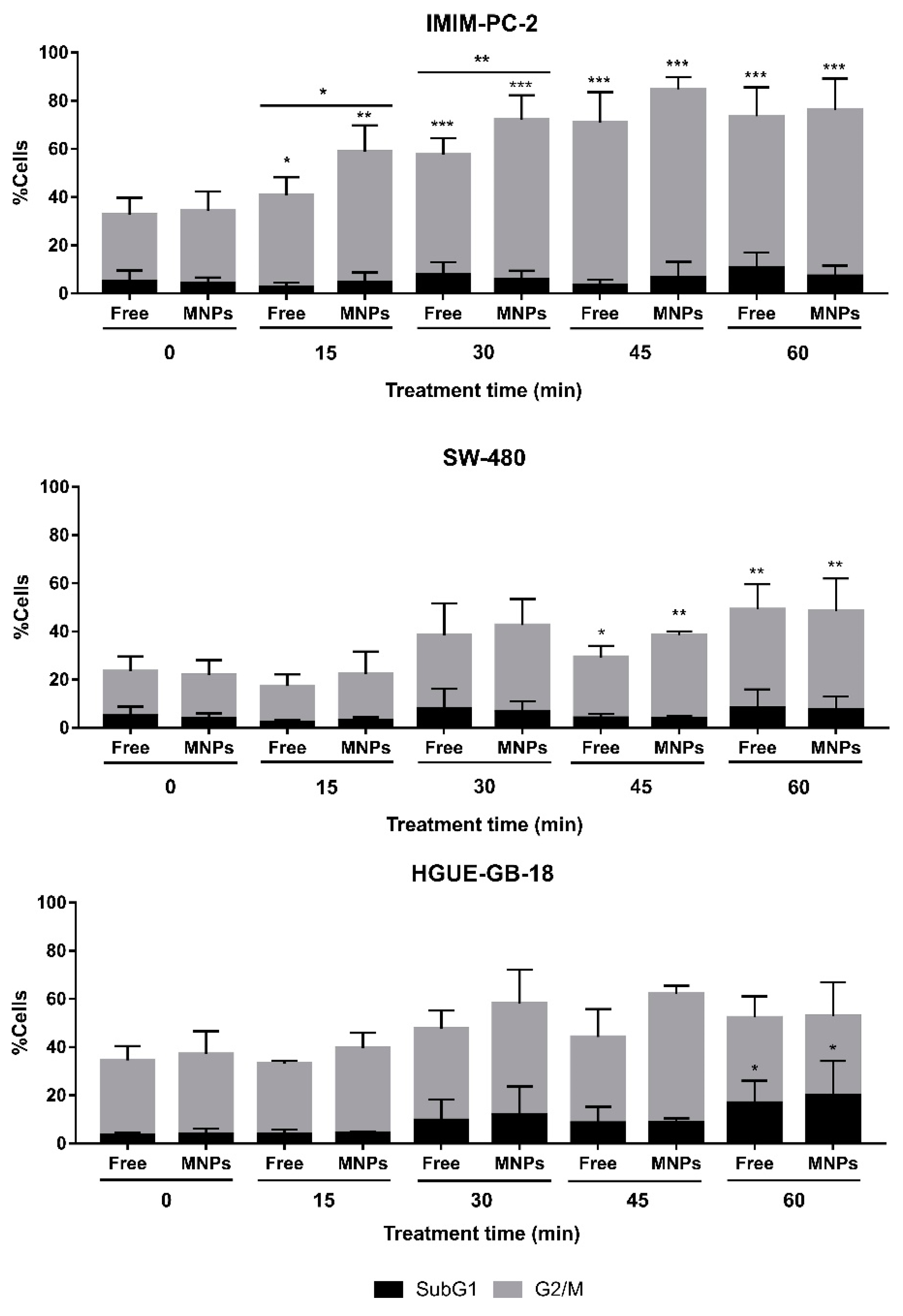
2.5. CLytA-DAAO and D-Ala Induce Cell Death in a Different Way than Direct H2O2 Addition
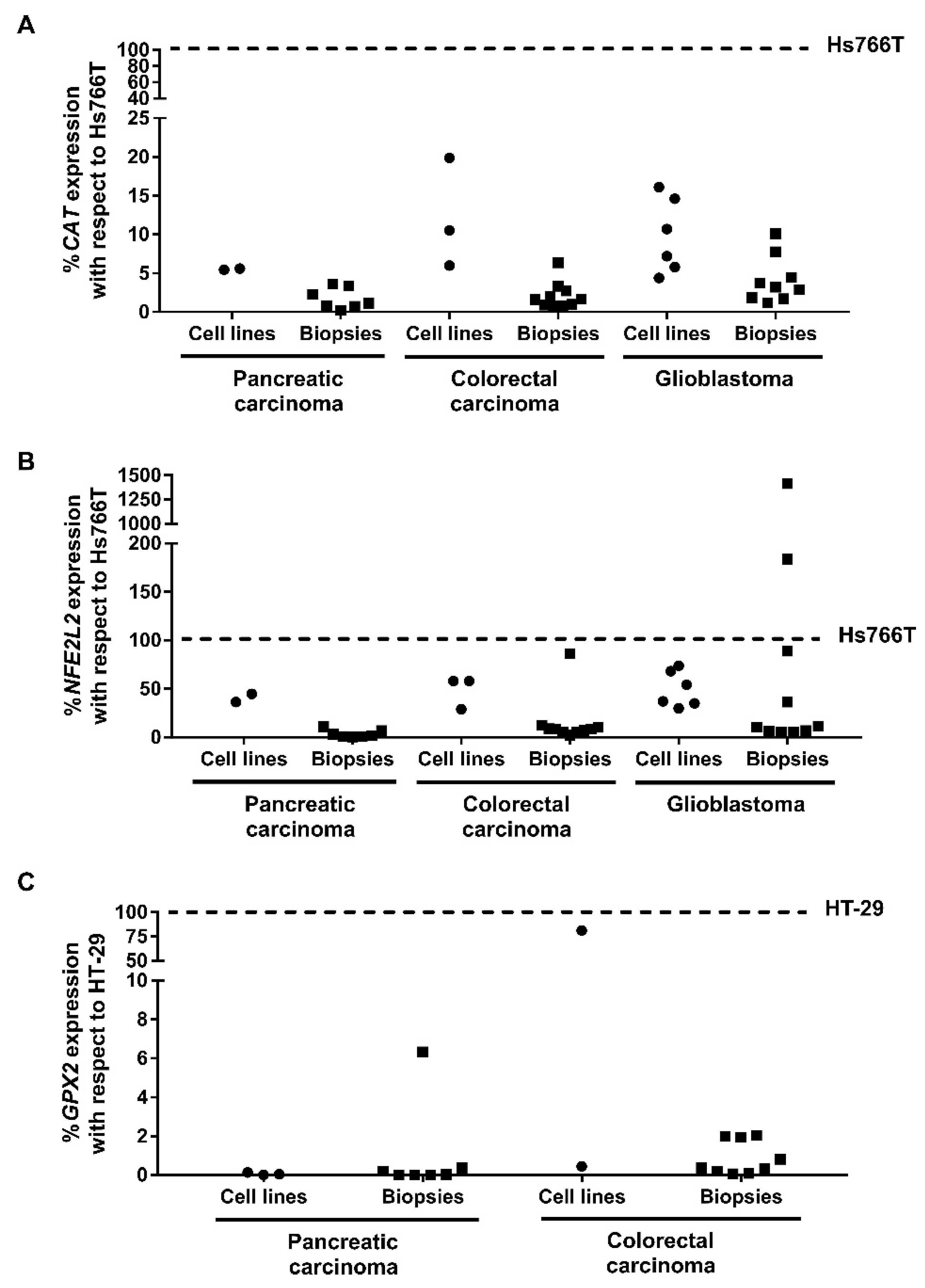
2.6. Expression of Genes Involved in Resistance to CLytA-DAAO-Induced Cell Death in Patient Samples
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Patient Biopsies
4.3. Treatments
4.4. Cell Death
4.5. Proliferation Assays
4.6. Cell Cycle Distribution
4.7. Intracellular Free Radicals’ Measurement
4.8. Gene Expression Analysis
4.9. Statistical Analysis
5. Conclusions
- Our results demonstrate that CLytA-DAAO bound to MNPs is more effective at inducing cytotoxicity than free CLytA-DAAO in pancreatic carcinoma, colorectal carcinoma, and glioblastoma cell models.
- The higher effect is due to the fact that immobilization increases the stability of the enzyme at 37 °C, maintaining its catalytic activity for a longer time.
- The enzyme is bound to the MNPs by a non-covalent immobilization system, which allows the release of the enzyme once it has reached its target by the external addition of choline.
- The cytotoxic effect induced by CLytA-DAAO is due to the prolonged production of ROS over time and is not comparable to adding H2O2 directly.
- Expression analysis of genes, which we have previously found to be related to CLytA-DAAO resistance, performed in biopsies as well as data extracted from UALCAN, suggest that CLytA-DAAO bound to MNPs could be effective as an anti-cancer therapy in a wide range of patients from pancreatic carcinoma, colorectal carcinoma, and glioblastoma. However, NFE2L2 expression has to be taken in consideration, especially in glioblastoma patients.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Illanes, A.; Cauerhff, A.; Wilson, L.; Castro, G.R. Recent trends in biocatalysis engineering. Bioresour. Thechnol. 2012, 115, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Ali Khan, A.; Alzohairy, M.A. Recent Advances and Applications of Immobilized Enzyme Technologies: A Review. Res. J. Biol. Sci. 2010, 5, 565–575. [Google Scholar] [CrossRef]
- Ansari, S.A.; Husain, Q. Potential applications of enzymes immobilized on/in nano materials: A review. Biotechnol. Adv. 2012, 30, 512–523. [Google Scholar] [CrossRef] [PubMed]
- De la Rica, R.; Aili, D.; Stevens, M.M. Enzyme-responsive nanoparticles for drug release and diagnostics. Adv. Drug Deliv. Rev. 2012, 64, 967–978. [Google Scholar] [CrossRef]
- Koutsopoulos, S. Molecular fabrications of smart nanobiomaterials and applications in personalized medicine. Adv. Drug Deliv. Rev. 2012, 64, 1459–1476. [Google Scholar] [CrossRef]
- Martinelli, C.; Pucci, C.; Ciofani, G. Nanostructured carriers as innovative tools for cancer diagnosis and therapy. APL Bioeng. 2019, 3, 011502. [Google Scholar] [CrossRef] [Green Version]
- Lu, A.H.; Salabas, E.L.; Schüth, F. Magnetic nanoparticles: Synthesis, protection, functionalization, and application. Angew. Chem. Int. Ed. Engl. 2007, 46, 1222–1244. [Google Scholar] [CrossRef]
- Sassolas, A.; Blum, L.J.; Leca-Bouvier, B.D. Immobilization strategies to develop enzymatic biosensors. Biotechnol. Adv. 2012, 30, 489–511. [Google Scholar] [CrossRef]
- Akbarzadeh, A.; Samiei, M.; Davaran, S. Magnetic nanoparticles: Preparation, physical properties, and applications in biomedicine. Nanoscale Res. Lett. 2012, 7, 144. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.E.; Shin, J.Y.; Cho, M.H. Magnetic nanoparticles: An update of application for drug delivery and possible toxic effects. Arch. Toxicol. 2012, 86, 685–700. [Google Scholar] [CrossRef]
- Fuentes-Baile, M.; Bello-Gil, D.; Pérez-Valenciano, E.; Sanz, J.M.; García-Morales, P.; Maestro, B.; Ventero, M.P.; Alenda, C.; Barberá, V.M.; Saceda, M. CLytA-DAAO, free and immobilized in magnetic nanoparticles, induces cell death in human cancer cells. Biomolecules 2020, 10, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollegioni, L.; Piubelli, L.; Sacchi, S.; Pilone, M.S.; Molla, G. Physiological functions of D-amino acid oxidases: From yeast to humans. Cell. Mol. Life Sci. 2007, 64, 1373–1394. [Google Scholar] [CrossRef] [PubMed]
- Stegman, L.D.; Zheng, H.; Neal, E.R.; Ben-Yoseph, O.; Pollegioni, L.; Pilone, M.S.; Ross, B.D. Induction of cytotoxic oxidative stress by D-alanine in brain tumor cells expressing Rhodotorula gracilis D-amino acid oxidase: A cancer gene therapy strategy. Hum. Gene Ther. 1998, 9, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Yoseph, O.; Ross, B.D. Oxidation therapy: The use of a reactive oxygen species-generating enzyme system for tumour treatment. Br. J. Cancer 1994, 70, 1131–1135. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, T.; Kokura, S.; Tainaka, K.; Naito, Y.; Kondo, M. A novel cancer therapy based on oxygen radicals. Cancer Res. 1995, 55, 1617–1620. [Google Scholar] [PubMed]
- Fuentes-baile, M.; García-morales, P.; Pérez-valenciano, E.; Ventero, M.P.; Sanz, J.M.; Romero, C.d.J.; Barberá, V.M.; Alenda, C.; Saceda, M. Cell death mechanisms induced by CLytA-DAAO chimeric enzyme in human tumor cell lines. Int. J. Mol. Sci. 2020, 21, 8522. [Google Scholar] [CrossRef]
- López, R.; García, E. Recent trends on the molecular biology of pneumococcal capsules, lytic enzymes, and bacteriophage. FEMS Microbiol. Rev. 2004, 28, 553–580. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Puelles, J.M.; Sanz, J.M.; Garcia, J.L.; Garcia, E. Immobilization and single-step purification of fusion proteins using DEAE-cellulose. Eur. J. Biochem. 1992, 203, 153–159. [Google Scholar] [CrossRef]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- Pollegioni, L.; Molla, G. New biotech applications from evolved D-amino acid oxidases. Trends Biotechnol. 2011, 29, 276–283. [Google Scholar] [CrossRef]
- Golini, P.; Bianchi, D.; Battistel, E.; Cesti, P.; Tassinari, R. Immobilization of d-amino acid oxidase from different yeasts: Characterization and application in the deamination of cephalosporin C. Enzym. Microb. Technol. 1995, 17, 324–329. [Google Scholar] [CrossRef]
- Fernández-Lafuente, R.; Rodríguez, V.; Mateo, C.; Fernández-Lorente, G.; Arminsen, P.; Sabuquillo, P.; Guisán, J.M. Stabilization of enzymes (D-amino acid oxidase) against hydrogen peroxide via immobilization and post-immobilization techniques. J. Mol. Catal. B Enzym. 1999, 7, 173–179. [Google Scholar] [CrossRef]
- Mateo, C.; Abian, O.; Fernandez-Lafuente, R.; Guisan, J.M. Reversible enzyme immobilization via a very strong and nondistorting ionic adsorption on support-polyethylenimine composites. Biotechnol. Bioeng. 2000, 68, 98–105. [Google Scholar] [CrossRef]
- López-Gallego, F.; Betancor, L.; Mateo, C.; Hidalgo, A.; Alonso-Morales, N.; Dellamora-Ortiz, G.; Guisán, J.M.; Fernández-Lafuente, R. Enzyme stabilization by glutaraldehyde crosslinking of adsorbed proteins on aminated supports. J. Biotechnol. 2005, 119, 70–75. [Google Scholar] [CrossRef]
- Zheng, H.; Wang, X.; Chen, J.; Zhu, K.; Zhao, Y.; Yang, Y.; Yang, S.; Jiang, W. Expression, purification, and immobilization of His-tagged D-amino acid oxidase of Trigonopsis variabilis in Pichia pastoris. Appl. Microbiol. Biotechnol. 2006, 70, 683–689. [Google Scholar] [CrossRef]
- Kuan, I.; Liao, R.; Hsieh, H.; Chen, K.; Yu, C. Properties of Rhodotorula gracilis d-Amino Acid Oxidase Immobilized on Magnetic Beads through His-Tag. J. Biosci. Bioeng. 2008, 105, 110–115. [Google Scholar] [CrossRef]
- Hsieh, H.C.; Kuan, I.C.; Lee, S.L.; Tien, G.Y.; Wang, Y.J.; Yu, C.Y. Stabilization of d-amino acid oxidase from Rhodosporidium toruloides by immobilization onto magnetic nanoparticles. Biotechnol. Lett. 2009, 31, 557–563. [Google Scholar] [CrossRef]
- Bava, A.; Gornati, R.; Cappellini, F.; Caldinelli, L.; Pollegioni, L.; Bernardini, G. D-amino acid oxidase-nanoparticle system: A potential novel approach for cancer enzymatic therapy. Nanomedicine 2013, 8, 1797–1806. [Google Scholar] [CrossRef] [Green Version]
- Chien, L.-J.; Lee, C.-K. Biosilicification of dual-fusion enzyme immobilized on magnetic nanoparticle. Biotechnol. Bioeng. 2008, 100, 223–230. [Google Scholar] [CrossRef]
- Wang, S.J.; Yu, C.Y.; Kuan, I.C. Stabilization of native and double d-amino acid oxidases from Rhodosporidium toruloides and Trigonopsis variabilis by immobilization on streptavidin-coated magnetic beads. Biotechnol. Lett. 2008, 30, 1973–1981. [Google Scholar] [CrossRef]
- De Jong, W.H.; Borm, P.J.A. Drug delivery and nanoparticles: Applications and hazards. Int. J. Nanomed. 2008, 3, 133–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lankveld, D.P.K.; Oomen, A.G.; Krystek, P.; Neigh, A.; Troost-de Jong, A.; Noorlander, C.W.; Van Eijkeren, J.C.H.; Geertsma, R.E.; De Jong, W.H. The kinetics of the tissue distribution of silver nanoparticles of different sizes. Biomaterials 2010, 31, 8350–8361. [Google Scholar] [CrossRef] [PubMed]
- Hosta-Rigau, L.; Olmedo, I.; Arbiol, J.; Cruz, L.J.; Kogan, M.J.; Albericio, F. Multifunctionalized gold nanoparticles with peptides targeted to gastrin-releasing peptide receptor of a tumor cell line. Bioconjug. Chem. 2010, 21, 1070–1078. [Google Scholar] [CrossRef] [PubMed]
- Saxena, V.K.; Deb, R.; Shrivastava, S.; Kantaraja, C.; Kumar, A.; Kumar, S. Functionalizing gold nanoparticles with bluetongue virus multiple peptide antigens utilizing gold-thiol interaction: A novel approach to develop pen side test. Res. Vet. Sci. 2012, 93, 1531–1536. [Google Scholar] [CrossRef]
- Shukla, R.; Bansal, V.; Chaudhary, M.; Basu, A.; Bhonde, R.R.; Sastry, M. Biocompatibility of gold nanoparticles and their endocytotic fate inside the cellular compartment: A microscopic overview. Langmuir 2005, 21, 10644–10654. [Google Scholar] [CrossRef]
- Sun, T.; Zhang, Y.S.; Pang, B.; Hyun, D.C.; Yang, M.; Xia, Y. Engineered nanoparticles for drug delivery in cancer therapy. Angew. Chem. Int. Ed. 2014, 53, 12320–12364. [Google Scholar] [CrossRef]
- Connor, E.E.; Mwamuka, J.; Gole, A.; Murphy, C.J.; Wyatt, M.D. Gold nanoparticles are taken up by human cells but do not cause acute cytotoxicity. Small 2005, 1, 325–327. [Google Scholar] [CrossRef]
- Abadeer, N.S.; Murphy, C.J. Recent Progress in Cancer Thermal Therapy Using Gold Nanoparticles. J. Phys. Chem. C 2016, 120, 4691–4716. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, Y.; Ding, T.; Liu, J.; Zhao, H. Multifunctional Gold Nanoparticles: A Novel Nanomaterial for Various Medical Applications and Biological Activities. Front. Bioeng. Biotechnol. 2020, 8, 990. [Google Scholar] [CrossRef]
- Ramalingam, V. Multifunctionality of gold nanoparticles: Plausible and convincing properties. Adv. Colloid Interface Sci. 2019, 271, 101989. [Google Scholar] [CrossRef]
- Brown, S.D.; Nativo, P.; Smith, J.A.; Stirling, D.; Edwards, P.R.; Venugopal, B.; Flint, D.J.; Plumb, J.A.; Graham, D.; Wheate, N.J. Gold nanoparticles for the improved anticancer drug delivery of the active component of oxaliplatin. J. Am. Chem. Soc. 2010, 132, 4678–4684. [Google Scholar] [CrossRef]
- Stuchinskaya, T.; Moreno, M.; Cook, M.J.; Edwards, D.R.; Russell, D.A. Targeted photodynamic therapy of breast cancer cells using antibody-phthalocyanine-gold nanoparticle conjugates. Photochem. Photobiol. Sci. 2011, 10, 822–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordeiro, M.; Carlos, F.F.; Pedrosa, P.; Lopez, A.; Baptista, P.V. Gold nanoparticles for diagnostics: Advances towards points of care. Diagnostics 2016, 6, 43. [Google Scholar] [CrossRef] [PubMed]
- Pereda, M.; Poncelet, D.; Renard, D. Characterization of Core-Shell Alginate Capsules. Food Biophys. 2019, 14, 467–478. [Google Scholar] [CrossRef]
- Dobson, J. Magnetic Nanoparticles for Drug Delivery. Drug Dev. Res. 2006, 67, 55–60. [Google Scholar] [CrossRef]
- Arruebo, M.; Fernández-Pacheco, R.; Ibarra, M.R.; Santamaria, J. Magnetic Nanoparticles for Drug Delivery Applications. Nanotoday 2007, 2, 22–32. [Google Scholar] [CrossRef]
- Yoo, D.; Lee, J.H.; Shin, T.H.; Cheon, J. Theranostic magnetic nanoparticles. Acc. Chem. Res. 2011, 44, 863–874. [Google Scholar] [CrossRef]
- Dennis, C.L.; Ivkov, R. Physics of heat generation using magnetic nanoparticles for hyperthermia. Int. J. Hyperth. 2013, 29, 715–729. [Google Scholar] [CrossRef]
- Sanchez, C.; Belleville, P.; Popall, M.; Nicole, L. Applications of advanced hybrid organic–inorganic nanomaterials: From laboratory to market. Chem. Soc. Rev. 2011, 40, 696–753. [Google Scholar] [CrossRef]
- Thiesen, B.; Jordan, A. Clinical applications of magnetic nanoparticles for hyperthermia. Int. J. Hyperth. 2008, 24, 467–474. [Google Scholar] [CrossRef]
- Pollegioni, L.; Caldinelli, L.; Molla, G.; Sacchi, S.; Pilone, M.S. Catalytic properties of D-amino acid oxidase in cephalosporin C bioconversion: A comparison between proteins from different sources. Biotechnol. Prog. 2004, 20, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Betancor, L.; Hidalgo, A.; Fernandez-Lorente, G.; Mateo, C.; Rodriguez, V.; Fuentes, M.; Lopez-Gallego, F.; Fernandez-Lafuente, R.; Guisan, J.M. Use of Physicochemical Tools to Determine the Choice of Optimal Enzyme: Stabilization of D-Amino Acid Oxidase. Biotechnol. Prog. 2003, 19, 784–788. [Google Scholar] [CrossRef] [PubMed]
- Park, W.H. Hydrogen peroxide inhibits the growth of lung cancer cells via the induction of cell death and G1-phase arrest. Oncol. Rep. 2018, 40, 1787–1794. [Google Scholar] [CrossRef] [PubMed]
- Park, W.H. Anti-apoptotic effect of caspase inhibitors on H2O2-treated HeLa cells through early suppression of its oxidative stress. Oncol. Rep. 2014, 31, 2413–2421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinaldi, M.; Caffo, M.; Minutoli, L.; Marini, H.; Abbritti, R.V.; Squadrito, F.; Trichilo, V.; Valenti, A.; Barresi, V.; Altavilla, D.; et al. ROS and brain gliomas: An overview of potential and innovative therapeutic strategies. Int. J. Mol. Sci. 2016, 17, 984. [Google Scholar] [CrossRef] [Green Version]
- Ventero, M.P.; Fuentes-Baile, M.; Quereda, C.; Perez-Valeciano, E.; Alenda, C.; Garcia-Morales, P.; Esposito, D.; Dorado, P.; Manuel Barbera, V.; Saceda, M. Radiotherapy resistance acquisition in Glioblastoma. Role of SOCS1 and SOCS3. PLoS ONE 2019, 14, e0212581. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin | Cell Line | %Inhibition (0.025 U/mL) | %Inhibition (0.25 U/mL) | IC50 (U/mL) | |||
|---|---|---|---|---|---|---|---|
| Free | MNPs | Free | MNPs | Free | MNPs | ||
| PC | IMIM-PC-2 | 8.6 ± 10.5 | 17.0 ± 10.1 | 60.5 ± 8.5 | 73.4 ± 3.5 | 0.19 ± 0.04 | 0.13 ± 0.02 |
| RWP-1 | 7.5 ± 6.9 | 5.9 ± 9.5 | 61.8 ± 11.1 | 80.2 ± 4.3 | 0.19 ± 0.02 | 0.14 ± 0.03 | |
| Hs766T | 33.8 ± 14.4 | 47.7 ± 3.1 | 62.7 ± 13.5 | 62.5 ± 14.6 | 0.10 ± 0.02 | 0.05 ± 0.02 | |
| CRC | SW-480 | 16.1 ± 8.1 | 26.9 ± 6.9 | 45.6 ± 11.3 | 67.7 ± 7.0 | 0.17 ± 0.03 | 0.09 ± 0.01 |
| SW-620 | 23.6 ± 5.4 | 41.7 ± 5.8 | 57.3 ± 3.3 | 78.1 ± 6.4 | 0.16 ± 0.02 | 0.05 ± 0.01 | |
| HT-29 | 0.0 ± 5.6 | 33.9 ± 8.5 | 22.4 ± 8.1 | 64.9 ± 8.1 | 0.68 ± 0.10 | 0.10 ± 0.02 | |
| GBM | HGUE-GB-18 | 15.4 ± 8.9 | 34.3 ± 7.9 | 24.4 ± 8.4 | 81.4 ± 8.7 | 0.50 ± 0.08 | 0.07 ± 0.01 |
| HGUE-GB-37 | 22.2 ± 5.8 | 37.8 ± 4.9 | 30.7 ± 7.6 | 75.7 ± 3.9 | 0.38 ± 0.06 | 0.07 ± 0.01 | |
| HGUE-GB-39 | 35.8 ± 7.9 | 50.1 ± 1.5 | 41.0 ± 8.4 | 81.2 ± 4.5 | 0.27 ± 0.06 | 0.03 ± 0.01 | |
| HGUE-GB-42 | 7.8 ± 6.4 | 30.8 ± 6.1 | 26.9 ± 9.0 | 72.9 ± 7.1 | 0.54 ± 0.05 | 0.08 ± 0.01 | |
| Cell Line | IC50 (U/mL) | |
|---|---|---|
| Free | MNPs | |
| IMIM-PC-2 | 1.26 ± 0.17 | 0.11 ± 0.03 |
| RWP-1 | 0.70 ± 0.08 | 0.12 ± 0.02 |
| SW-480 | 1.12 ± 0.21 | 0.24 ± 0.08 |
| SW-620 | 0.78 ± 0.30 | 0.17 ± 0.08 |
| Origin | Cell Line | IC50 (μM) |
|---|---|---|
| PC | Hs766T | 358.0 ± 76.5 |
| IMIM-PC-2 | 222.1 ± 55.9 | |
| RWP-1 | 83.71 ± 1.92 | |
| CRC | SW-480 | 54.1 ± 2.77 |
| SW-620 | 145.3 ± 17.0 | |
| GBM | HGUE-GB-37 | 776.6 ± 157.5 |
| HGUE-GB-39 | 362.3 ± 95.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuentes-Baile, M.; Pérez-Valenciano, E.; García-Morales, P.; de Juan Romero, C.; Bello-Gil, D.; Barberá, V.M.; Rodríguez-Lescure, Á.; Sanz, J.M.; Alenda, C.; Saceda, M. CLytA-DAAO Chimeric Enzyme Bound to Magnetic Nanoparticles. A New Therapeutical Approach for Cancer Patients? Int. J. Mol. Sci. 2021, 22, 1477. https://doi.org/10.3390/ijms22031477
Fuentes-Baile M, Pérez-Valenciano E, García-Morales P, de Juan Romero C, Bello-Gil D, Barberá VM, Rodríguez-Lescure Á, Sanz JM, Alenda C, Saceda M. CLytA-DAAO Chimeric Enzyme Bound to Magnetic Nanoparticles. A New Therapeutical Approach for Cancer Patients? International Journal of Molecular Sciences. 2021; 22(3):1477. https://doi.org/10.3390/ijms22031477
Chicago/Turabian StyleFuentes-Baile, María, Elizabeth Pérez-Valenciano, Pilar García-Morales, Camino de Juan Romero, Daniel Bello-Gil, Víctor M. Barberá, Álvaro Rodríguez-Lescure, Jesús M. Sanz, Cristina Alenda, and Miguel Saceda. 2021. "CLytA-DAAO Chimeric Enzyme Bound to Magnetic Nanoparticles. A New Therapeutical Approach for Cancer Patients?" International Journal of Molecular Sciences 22, no. 3: 1477. https://doi.org/10.3390/ijms22031477
APA StyleFuentes-Baile, M., Pérez-Valenciano, E., García-Morales, P., de Juan Romero, C., Bello-Gil, D., Barberá, V. M., Rodríguez-Lescure, Á., Sanz, J. M., Alenda, C., & Saceda, M. (2021). CLytA-DAAO Chimeric Enzyme Bound to Magnetic Nanoparticles. A New Therapeutical Approach for Cancer Patients? International Journal of Molecular Sciences, 22(3), 1477. https://doi.org/10.3390/ijms22031477