
A Role for Human DNA Polymerase λ in Alternative Lengthening of Telomeres

 , , , , ,
, , , , ,  ,
, 
Abstract
:1. Introduction
2. Results
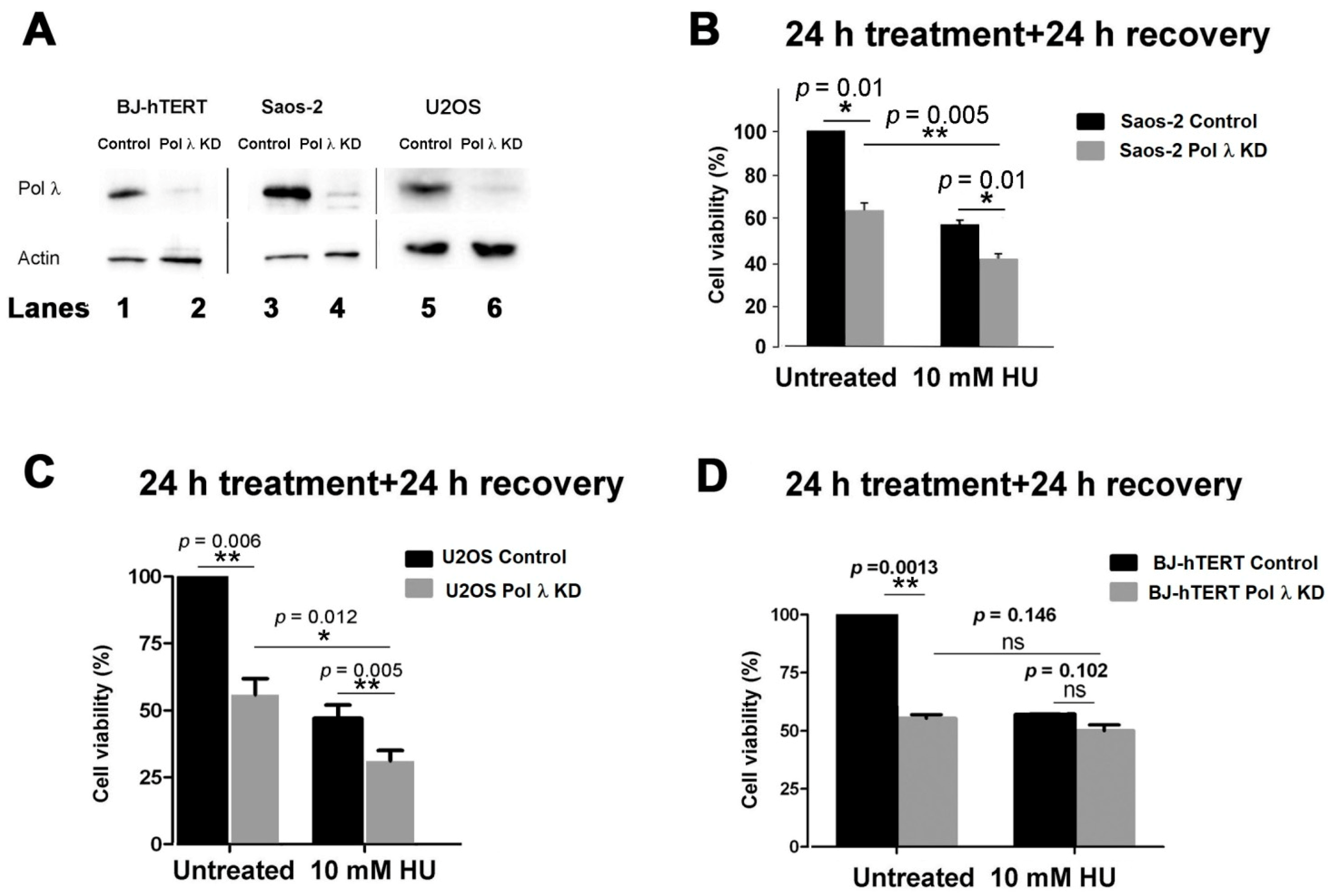
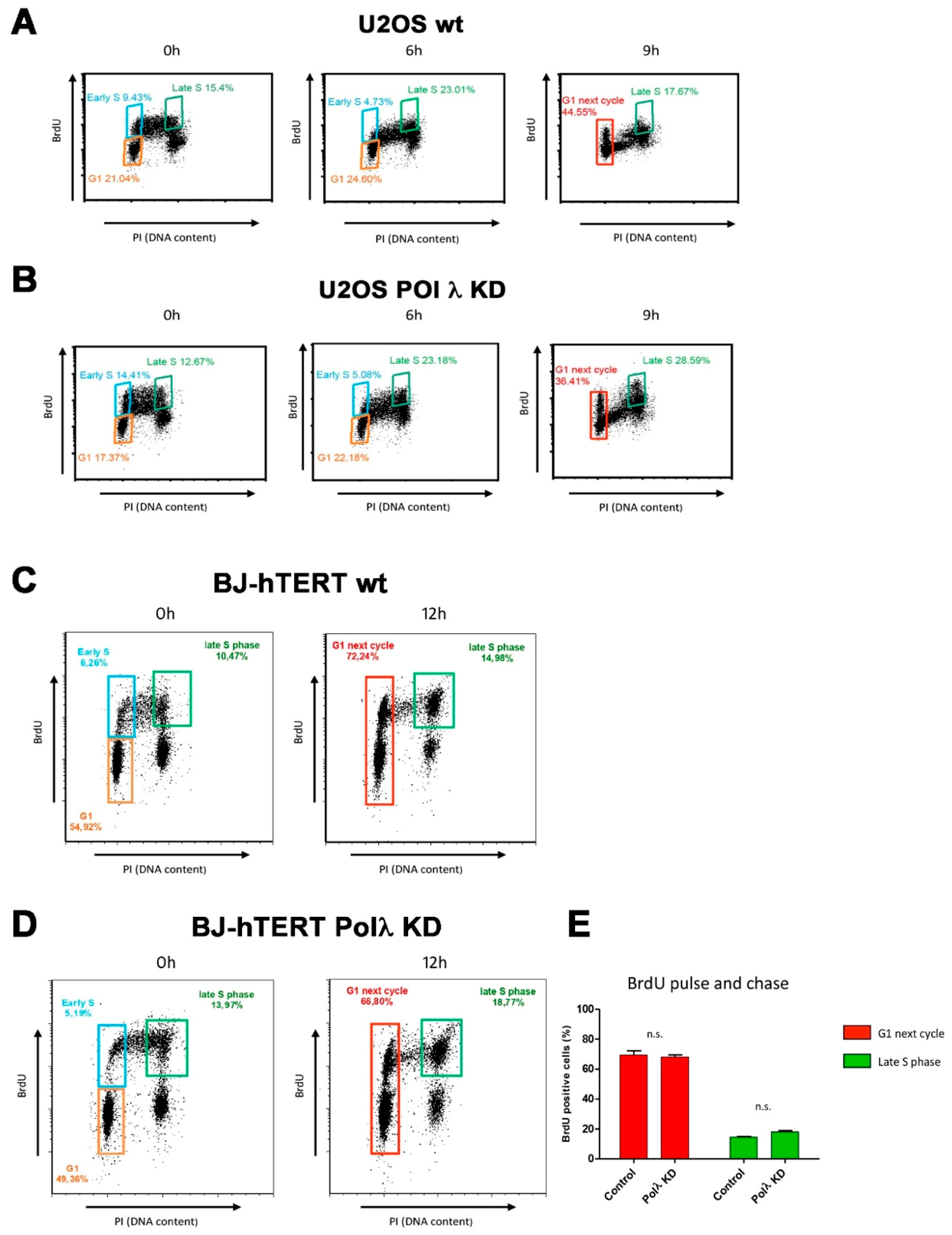
2.1. Silencing of Human DNA Polymerase λ Specifically Affects the Viability of ALT Cell Lines in the Presence of Replication Stress
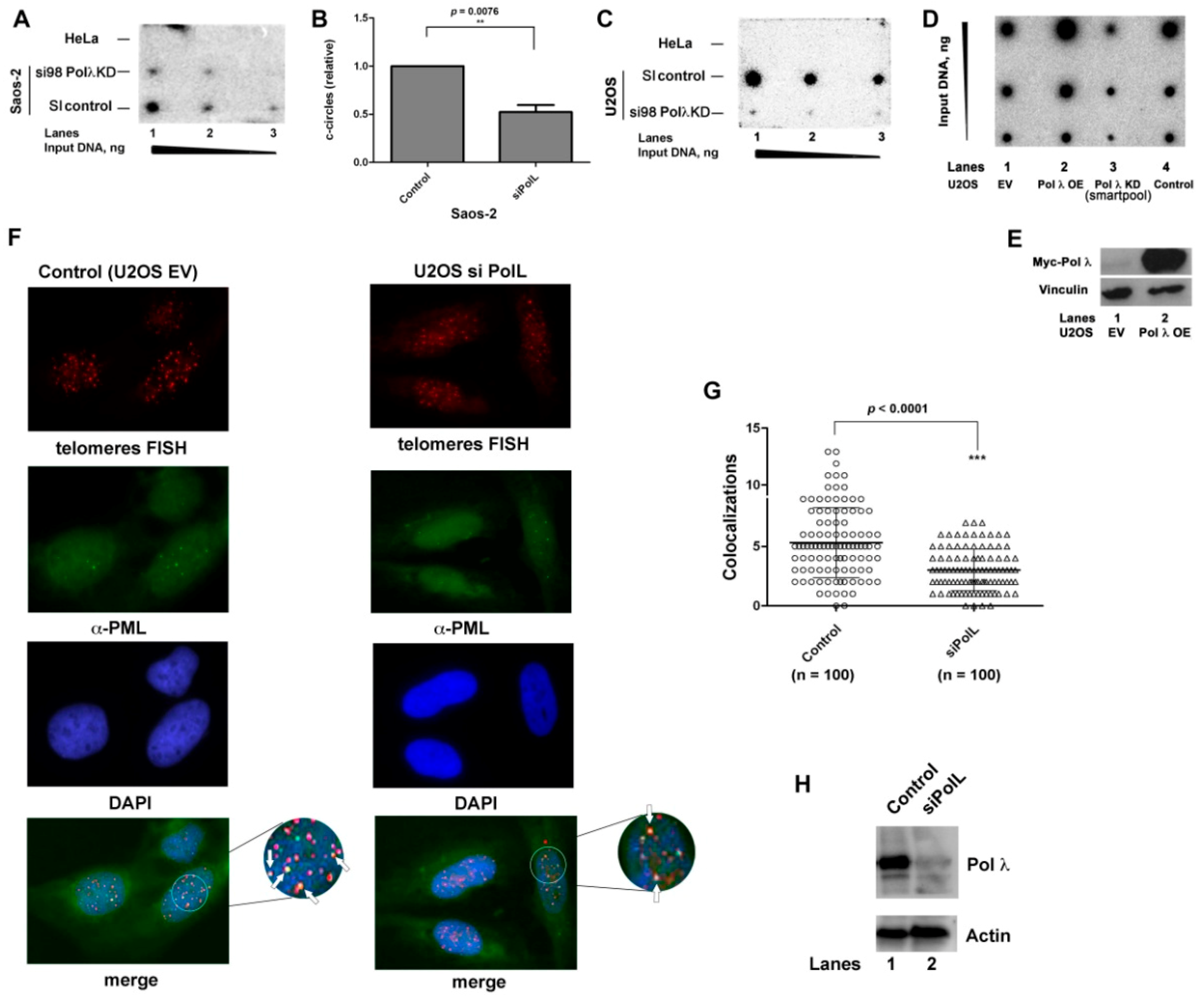
2.2. The Levels of DNA Polymerase λ Correlate with the Cellular ALT Activity
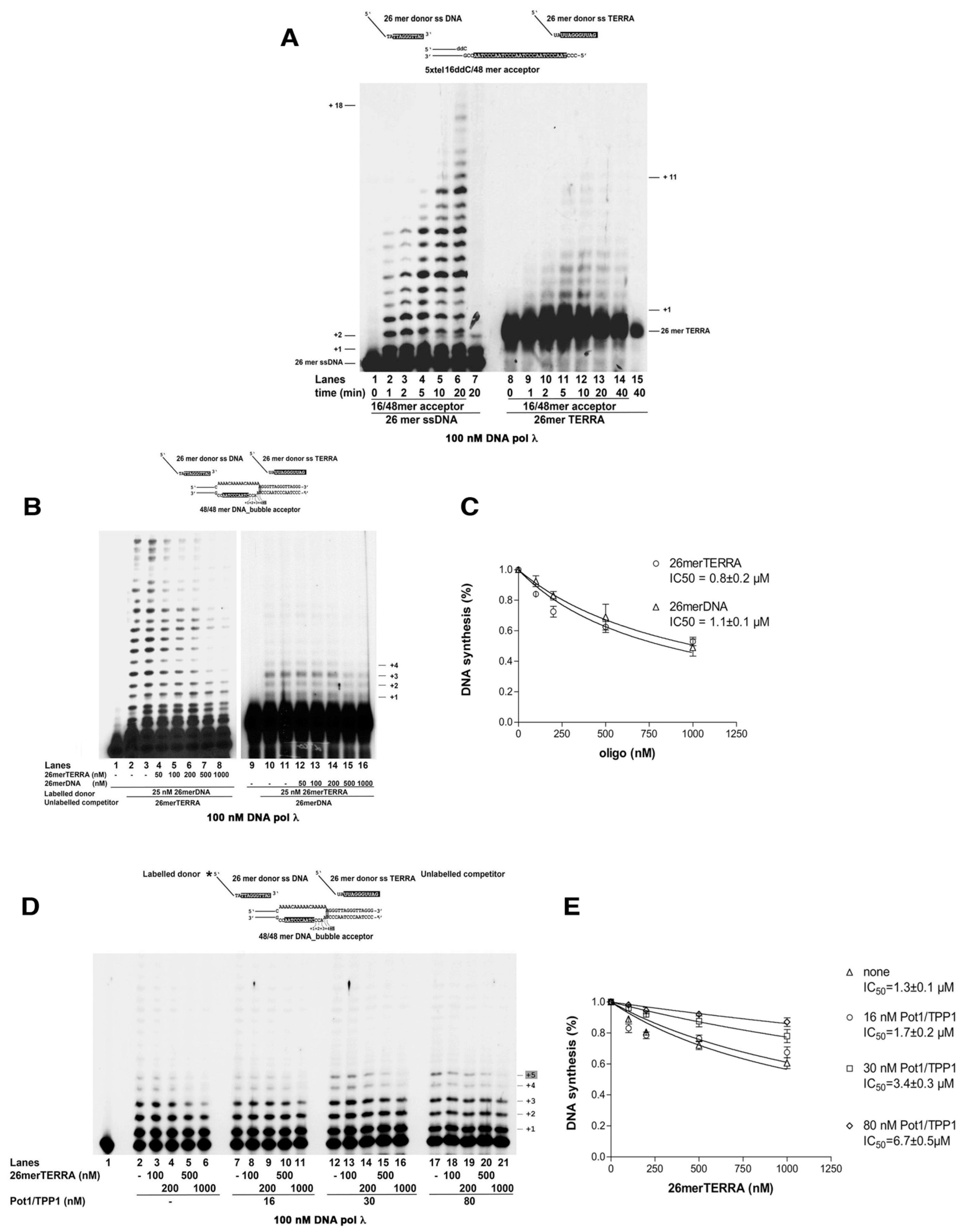
2.3. DNA Polymerase λ Can Catalyze Annealing and Elongation of a G-Rich Telomeric Strand to a Complementary C-Rich Telomeric Template Sequence
2.4. Replication Protein A Restricts DNA Polymerase λ Microhomology-Mediated Strand Transfer Activity
2.5. The POT1/TPP1 Complex Stimulates the Microhomology-Mediated Strand Transfer Activity of DNA Polymerase λ
2.6. TERRA RNA Can Function as a Substrate for the Microhomology-Mediated Strand Transfer Activity by DNA Polymerase λ
2.7. The POT1/TPP1 Complex Favors the Microhomology-Mediated Strand Transfer Activity by DNA Polymerase λ Starting from Telomeric DNA over TERRA RNA
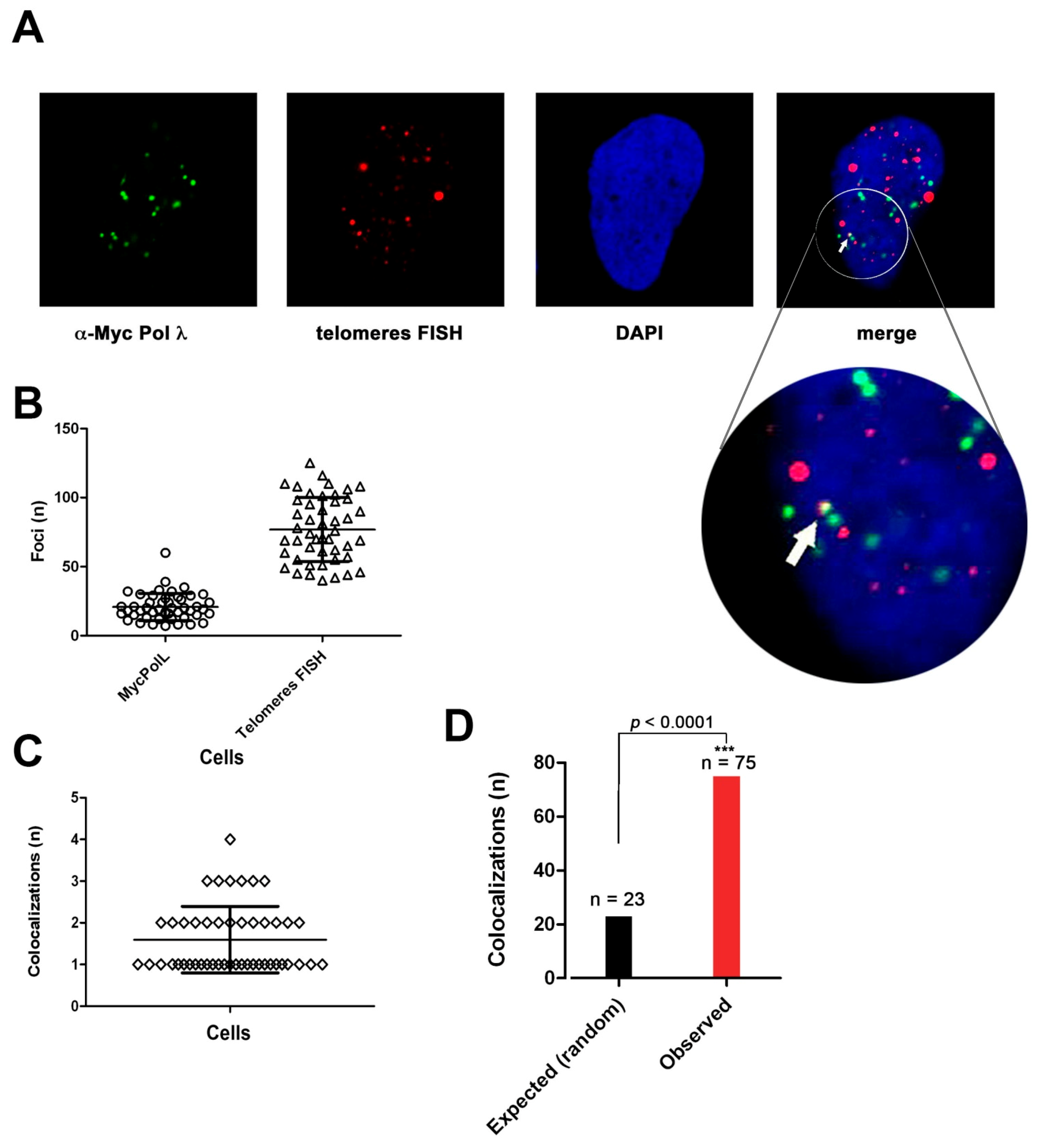
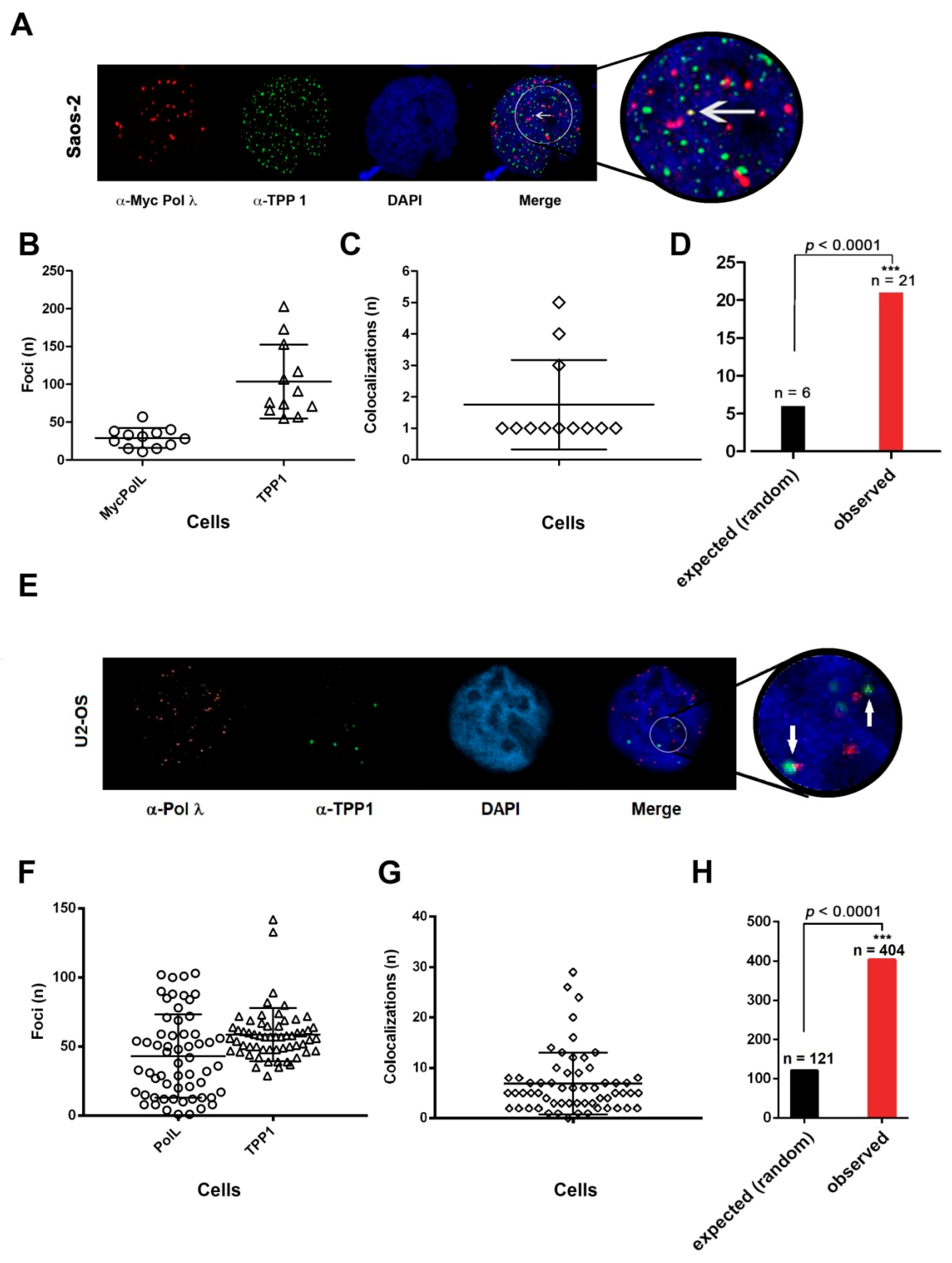
2.8. DNA Polymerase λ Associates with Telomeres and TPP1 in ALT Cells
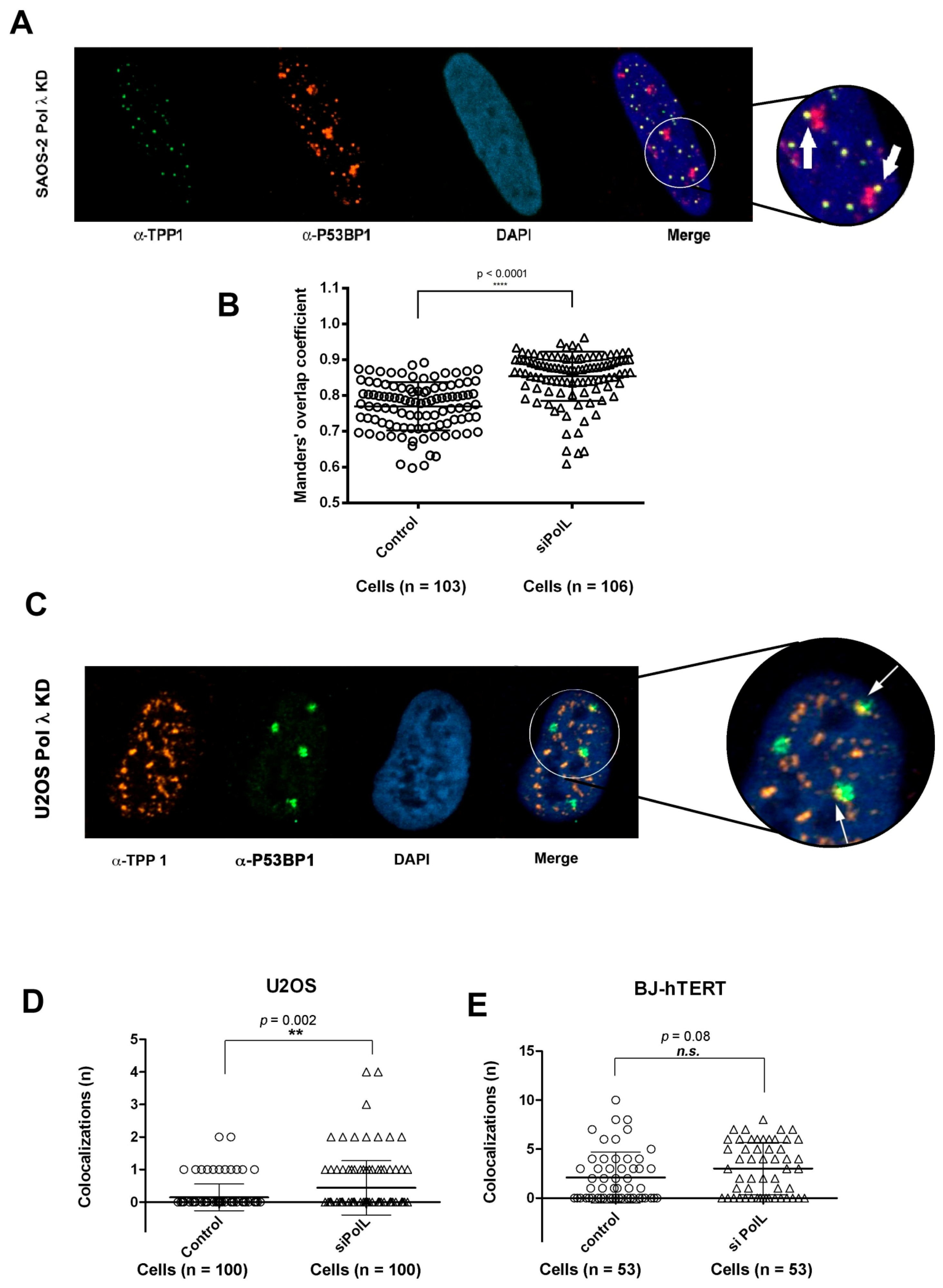
2.9. DNA Polymerase λ Silencing Induces Telomere Stress Specifically in ALT Cells
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals
- Acceptor linear (16ddC/48mer)
- 5′-CAGTCGATCGATCGAc (c = ddC)
- 3′-GTCAGCTAGCTAGCTGCCAATCCCAATCCCAACTATAGAGCGCTGTCA-5′
- Acceptor bubble (48mer_comp_bub/48mer)
- 5′-CAGTCGATCGATCGACAAAACAAAAACAAAAAGATATCTCGCGACAGT-3′
- 3′-GTCAGCTAGCTAGCTGCCAATCCCAATCCCAACTATAGAGCGCTGTCA-5′
- 5xtel16ddC/48mer (16ddC/5xtel48mer)
- 5′-CAGTCGATCGATCGAc (c = ddC)
- 3′-GTCAGCTAGCTAGCTGCCAATCCCAATCCCAATCCCAATCCCAATCCC-5′
- 5xtel acceptor bubble (5xtel48mer_comp_bub/5xtel48mer)
- 5′-CAGTCGATCGATCGACAAAACAAAAACAAAAAAGGGTTAGGGTTAGGG-3′
- 3′-GTCAGCTAGCTAGCTGCCAATCCCAATCCCAATCCCAATCCCAATCCC-5′
- 25 mer donor
- 5′-GTCAGCTAGCTAGCTATTAGGGTTA-3′
- 26 mer donor
- 5′-GTCAGCTAGCTAGCTATTAGGGTTAG-3′
- 26 mer TERRA
- 5′-GUCAGCUAGCUAGCUAUUAGGGUUAG-3′
5.2. Proteins Production and Purification
5.3. Cell Lines and Treatments
5.4. Protein Extracts
5.5. Cytotoxicity Assay
5.6. RNA Interference
5.7. Pol λ over Expression Stable Cell Lines
5.8. Immunofluorescence
5.8.1. Pol λ and TPP1 Colocalization
5.8.2. TPP1 and P53BP1 Colocalization
5.8.3. Immunofluorescence-FISH and APBs Detection
5.9. Terra RNA FISH
5.10. Cell Cycle Analysis
5.11. CC-Assay
5.12. Enzymatic Assays
5.13. Steady State Kinetic Analysis
5.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALT | Alternative Lengthening of Telomeres |
| Pol | DNA Polymerase |
| POT1/TTP1 | Protection of Telomeres protein 1 |
| HR | Homologous Recombination |
| RP-A | Replication Protein A |
| ECRTs | Extra-Chromosomal Telomeric Repeats |
| DSBs | Double Strand Breaks |
| aNHEJ | Alternative Microhomology-Mediated Non-Homologous End Joining Pathway |
| PARP-1 | Poly (ADP-Ribose) Polymerase |
| BIR | Break Induced Replication |
| TLS | Translesion Synthesis |
| MMST | Microhomology Mediated Strand Transfer |
| HU | Hydroxyurea |
| PML | ALT-associated promyelocytic leukemia |
| TERRA | Telomeric Repeats Containing RNA |
| TIFs | telomere dysfunction-induced foci |
References
- Apte, M.S.; Cooper, J.P. Life and cancer without telomerase: ALT and other strategies for making sure ends (don’t) meet. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 57–73. [Google Scholar] [CrossRef]
- Sobinoff, A.P.; Pickett, H.A. Alternative Lengthening of Telomeres: DNA Repair Pathways Converge. Trends Genet. 2017, 33, 921–932. [Google Scholar] [CrossRef]
- Flynn, R.L.; Centore, R.C.; O’Sullivan, R.J.; Rai, R.; Tse, A.; Zhou, S.Y.; Chang, S.; Karlseder, J.; Zou, L. TERRA and hnRNPA1 orchestrate an RPA-to-POT1 switch on telomeric single-stranded DNA. Nature 2011, 471, 532–536. [Google Scholar] [CrossRef] [Green Version]
- Cesare, A.J.; Griffith, J.D. Telomeric DNA in ALT cells is characterized by free telomeric circles and heterogeneous t-loops. Mol. Cell. Biol. 2004, 24, 9948–9957. [Google Scholar] [CrossRef] [Green Version]
- Nabetani, A.; Ishikawa, F. Unusual Telomeric DNAs in Human Telomerase-Negative Immortalized Cells. Mol. Cell. Biol. 2009, 29, 703–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomaska, L.; McEachern, M.J.; Nosek, J. Alternatives to telomerase: Keeping linear chromosomes via telomeric circles. Febs Lett. 2004, 567, 142–146. [Google Scholar] [CrossRef]
- Henson, J.D.; Cao, Y.; Huschtscha, L.I.; Chang, A.C.; Au, A.Y.M.; Pickett, H.A.; Reddel, R.R. DNA C-circles are specific and quantifiable markers of alternative-lengthening-of-telomeres activity. Nat. Biotechnol. 2009, 27, 1181–1185. [Google Scholar] [CrossRef] [PubMed]
- Nosek, J.; Rycovska, A.; Makhov, A.M.; Griffith, J.D.; Tomaska, L. Amplification of telomeric arrays via rolling-circle mechanism. J. Biol. Chem. 2005, 280, 10840–10845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natarajan, S.; McEachern, M.J. Recombinational telomere elongation promoted by DNA circles. Mol. Cell. Biol. 2002, 22, 4512–4521. [Google Scholar] [CrossRef] [Green Version]
- Sfeir, A.; Kosiyatrakul, S.T.; Hockemeyer, D.; MacRae, S.L.; Karlseder, J.; Schildkraut, C.L.; de Lange, T. Mammalian Telomeres Resemble Fragile Sites and Require TRF1 for Efficient Replication. Cell 2009, 138, 90–103. [Google Scholar] [CrossRef] [Green Version]
- Fumagalli, M.; Rosiello, F.; Clerici, M.; Barozzi, S.; Cittaro, D.; Kaplunov, J.M.; Bucci, G.; Dobreva, M.; Matti, V.; Beausejour, C.M.; et al. Telomeric DNA damage is irreparable and causes persistent DNA-damage-response activation. Nat. Cell Biol. 2012, 14, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Gan, W.J.; Guan, Z.S.; Liu, J.; Gui, T.; Shen, K.; Manley, J.L.; Li, X.L. R-loop-mediated genomic instability is caused by impairment of replication fork progression. Genes Dev. 2011, 25, 2041–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rippe, K.; Luke, B. TERRA and the state of the telomere. Nat. Struct. Mol. Biol. 2015, 22, 853–858. [Google Scholar] [CrossRef]
- Hu, Y.; Shi, G.; Zhang, L.C.; Li, F.; Jiang, Y.L.; Jiang, S.; Ma, W.B.; Zhao, Y.; Zhou, S.Y.; Huang, J.J. Switch telomerase to ALT mechanism by inducing telomeric DNA damages and dysfunction of ATRX and DAXX. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cho, N.W.; Dilley, R.L.; Lampson, M.A.; Greenberg, R.A. Interchromosomal Homology Searches Drive Directional ALT Telomere Movement and Synapsis. Cell 2014, 159, 108–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sfeir, A.; de Lange, T. Removal of Shelterin Reveals the Telomere End-Protection Problem. Science 2012, 336, 593–597. [Google Scholar] [CrossRef] [Green Version]
- Doksani, Y.; de Lange, T. Telomere-Internal Double-Strand Breaks Are Repaired by Homologous Recombination and PARP1/Lig3-Dependent End-Joining. Cell Rep. 2016, 17, 1646–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roumelioti, F.M.; Sotiriou, S.K.; Katsini, V.; Chiourea, M.; Halazonetis, T.D.; Gagos, S. Alternative lengthening of human telomeres is a conservative DNA replication process with features of break-induced replication. Embo Rep. 2016, 17, 1731–1737. [Google Scholar] [CrossRef] [PubMed]
- Dilley, R.L.; Verma, P.; Cho, N.W.; Winters, H.D.; Wondisford, A.R.; Greenberg, R.A. Break-induced telomere synthesis underlies alternative telomere maintenance. Nature 2016, 539, 54–58. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.M.; Yadav, T.; Ouyang, J.; Lan, L.; Zou, L. Alternative Lengthening of Telomeres through Two Distinct Break-Induced Replication Pathways. Cell Rep. 2019, 26, 955–968. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.P.; Zhang, Z.P.; Gong, S.Z.; Li, X.C.; Liu, H.Y.; Zhao, Y. Strand break-induced replication fork collapse leads to C-circles, C-overhangs and telomeric recombination. PLoS Genet. 2019, 15, e1007925. [Google Scholar] [CrossRef] [Green Version]
- Kent, T.; Chandramouly, G.; McDevitt, S.M.; Ozdemir, A.Y.; Pomerantz, R.T. Mechanism of microhomology-mediated end-joining promoted by human DNA polymerase theta. Nat. Struct. Mol. Biol. 2015, 22, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Mateos-Gomez, P.A.; Gong, F.D.; Nair, N.; Miller, K.M.; Lazzerini-Denchi, E.; Sfeir, A. Mammalian polymerase theta promotes alternative NHEJ and suppresses recombination. Nature 2015, 518, 254–U285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Exposito, L.; Bournique, E.; Bergoglio, V.; Bose, A.; Barroso-Gonzalez, J.; Zhang, S.F.; Roncaioli, J.L.; Lee, M.; Wallace, C.T.; Watkins, S.C.; et al. Proteomic Profiling Reveals a Specific Role for Translesion DNA Polymerase eta in the Alternative Lengthening of Telomeres. Cell Rep. 2016, 17, 1858–1871. [Google Scholar] [CrossRef] [Green Version]
- van Loon, B.; Hubscher, U.; Maga, G. Living on the Edge: DNA Polymerase Lambda between Genome Stability and Mutagenesis. Chem. Res. Toxicol. 2017, 30, 1936–1941. [Google Scholar] [CrossRef]
- Crespan, E.; Czabany, T.; Maga, G.; Hubscher, U. Microhomology-mediated DNA strand annealing and elongation by human DNA polymerases lambda and beta on normal and repetitive DNA sequences. Nucleic Acids Res. 2012, 40, 5577–5590. [Google Scholar] [CrossRef] [Green Version]
- Crespan, E.; Hubscher, U.; Maga, G. Expansion of CAG triplet repeats by human DNA polymerases lambda and beta in vitro, is regulated by flap endonuclease 1 and DNA ligase 1. Dna Repair 2015, 29, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Zucca, E.; Bertoletti, F.; Wimmer, U.; Ferrari, E.; Mazzini, G.; Khoronenkova, S.; Grosse, N.; van Loon, B.; Dianov, G.; Hubscher, U.; et al. Silencing of human DNA polymerase lambda causes replication stress and is synthetically lethal with an impaired S phase checkpoint. Nucleic Acids Res. 2013, 41, 229–241. [Google Scholar] [CrossRef]
- Chung, I.; Osterwald, S.; Deeg, K.I.; Rippe, K. PML body meets telomere. The beginning of an ALTernate ending? Nucl. Austin 2012, 3, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; de Lange, T. A Shld1-Controlled POT1a Provides Support for Repression of ATR Signaling at Telomeres through RPA Exclusion. Mol. Cell 2010, 40, 377–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flynn, R.L.; Cox, K.E.; Jeitany, M.; Wakimoto, H.; Bryll, A.R.; Ganem, N.J.; Bersani, F.; Pineda, J.R.; Suva, M.L.; Benes, C.H.; et al. Alternative lengthening of telomeres renders cancer cells hypersensitive to ATR inhibitors. Science 2015, 347, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Pinzaru, A.M.; Hom, R.A.; Beal, A.; Phillips, A.F.; Ni, E.; Cardozo, T.; Nair, N.; Choi, J.; Wuttke, D.S.; Sfeir, A.; et al. Telomere Replication Stress Induced by POT1 Inactivation Accelerates Tumorigenesis. Cell Rep. 2016, 15, 2170–2184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiang, W.G.; Wu, Q.Q.; Zhou, F.X.; Xie, C.H.; Wu, C.P.; Zhou, Y.F. Suppression of telomere-binding protein TPP1 resulted in telomere dysfunction and enhanced radiation sensitivity in telomerase-negative osteosarcoma cell line. Biochem. Biophys. Res. Commun. 2014, 445, 363–368. [Google Scholar] [CrossRef]
- Arora, R.; Azzalin, C.M. Telomere elongation chooses TERRA ALTernatives. Rna Biol. 2015, 12, 938–941. [Google Scholar] [CrossRef] [Green Version]
- Ramadan, K.; Maga, G.; Shevelev, I.V.; Villani, G.; Blanco, L.; Hubscher, U. Human DNA polymerase lambda possesses terminal deoxyribonucleotidyl transferase activity and can elongate RNA primers: Implications for novel functions. J. Mol. Biol. 2003, 328, 63–72. [Google Scholar] [CrossRef]
- Heaphy, C.M.; Subhawong, A.P.; Hong, S.M.; Goggins, M.G.; Montgomery, E.A.; Gabrielson, E.; Netto, G.J.; Epstein, J.I.; Lotan, T.L.; Westra, W.H.; et al. Prevalence of the Alternative Lengthening of Telomeres Telomere Maintenance Mechanism in Human Cancer Subtypes. Am. J. Pathol. 2011, 179, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Hakin-Smith, V.; Jellinek, D.A.; Levy, D.; Carroll, T.; Teo, M.; Timperley, W.R.; McKay, M.J.; Reddel, R.R.; Royds, J.A. Alternative lengthening of telomeres and survival in patients with glioblastoma multiforme. Lancet 2003, 361, 836–838. [Google Scholar] [CrossRef]
- Henson, J.D.; Reddel, R.R. Assaying and investigating Alternative Lengthening of Telomeres activity in human cells and cancers. Febs Lett. 2010, 584, 3800–3811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doksani, Y. The Response to DNA Damage at Telomeric Repeats and Its Consequences for Telomere Function. Genes 2019, 10, 318. [Google Scholar] [CrossRef] [Green Version]
- Grudic, A.; Jul-Larsen, A.; Haring, S.J.; Wold, M.S.; Lonning, P.E.; Bjerkvig, R.; Boe, S.O. Replication protein A prevents accumulation of single-stranded telomeric DNA in cells that use alternative lengthening of telomeres. Nucleic Acids Res. 2007, 35, 7267–7278. [Google Scholar] [CrossRef]
- Albertella, M.R.; Lau, A.; O’Connor, M.J. The overexpression of specialized DNA polymerases in cancer. Dna Repair 2005, 4, 583–593. [Google Scholar] [CrossRef] [PubMed]
- M’kacher, R.; Cuceu, C.; Al Jawhari, M.; Morat, L.; Frenzel, M.; Shim, G.; Lenain, A.; Hempel, W.M.; Junker, S.; Girinsky, T.; et al. The Transition between Telomerase and ALT Mechanisms in Hodgkin Lymphoma and Its Predictive Value in Clinical Outcomes. Cancers 2018, 10, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawlor, R.T.; Veronese, N.; Pea, A.; Nottegar, A.; Smith, L.; Pilati, C.; Demurtas, J.; Fassan, M.; Cheng, L.; Luchini, C. Alternative lengthening of telomeres (ALT) influences survival in soft tissue sarcomas: A systematic review with meta-analysis. BMC Cancer 2019, 19, 232. [Google Scholar] [CrossRef] [Green Version]
- Minasi, S.; Baldi, C.; Pietsch, T.; Donofrio, V.; Pollo, B.; Antonelli, M.; Massimino, M.; Giangaspero, F.; Buttarelli, F.R. Telomere elongation via alternative lengthening of telomeres (ALT) and telomerase activation in primary metastatic medulloblastoma of childhood. J. Neuro-Oncol. 2019, 142, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Maga, G.; Shevelev, I.; Villani, G.; Spadari, S.; Hubscher, U. Human replication protein A can suppress the intrinsic in vitro mutator phenotype of human DNA polymerase lambda. Nucleic Acids Res. 2006, 34, 1405–1415. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.Y.; Redon, S.; Lingner, J. The human CST complex is a terminator of telomerase activity. Nature 2012, 488, 540–544. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Bolte, S.; Cordelières, F.P. A guided tour into subcellular colocalization analysis in light microscopy. J. Microsc. 2006, 224, 213–232. [Google Scholar] [CrossRef]
- Cordelières, F.; Bolte, S. JACoP v 2.0: Improving the User Experience with Co-Localization Studies. 2008. Available online: https://imagejdocu.tudor.lu/_media/plugin/analysis/jacop_2.0/just_another_colocalization_plugin/jacop_ijconf2008.pdf (accessed on 1 July 2020).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acceptor Template | 16/48 | 16/48 5xtel | 48/48 Bubble | ||
|---|---|---|---|---|---|
| −RP-A | +RP-A | +RP-A | −RP-A | +RP-A | |
| Km (nM) | 12 ± 1 | 21 ± 2 | 11 ± 1 | 43 ± 4 | 124 ± 10 |
| Vmax (nM × min−1) | 9 ± 1 | 7 ± 1 | 7.5 ± 0.7 | 9 ± 1 | 10 ± 1 |
| Vmax/Km (min−1) | 0.75 | 0.33 | 0.68 | 0.21 | 0.08 |
| -fold reduction Vmax/Km | 2.27 | 1.1 | 2.62 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mentegari, E.; Bertoletti, F.; Kissova, M.; Zucca, E.; Galli, S.; Tagliavini, G.; Garbelli, A.; Maffia, A.; Bione, S.; Ferrari, E.; et al. A Role for Human DNA Polymerase λ in Alternative Lengthening of Telomeres. Int. J. Mol. Sci. 2021, 22, 2365. https://doi.org/10.3390/ijms22052365
Mentegari E, Bertoletti F, Kissova M, Zucca E, Galli S, Tagliavini G, Garbelli A, Maffia A, Bione S, Ferrari E, et al. A Role for Human DNA Polymerase λ in Alternative Lengthening of Telomeres. International Journal of Molecular Sciences. 2021; 22(5):2365. https://doi.org/10.3390/ijms22052365
Chicago/Turabian StyleMentegari, Elisa, Federica Bertoletti, Miroslava Kissova, Elisa Zucca, Silvia Galli, Giulia Tagliavini, Anna Garbelli, Antonio Maffia, Silvia Bione, Elena Ferrari, and et al. 2021. "A Role for Human DNA Polymerase λ in Alternative Lengthening of Telomeres" International Journal of Molecular Sciences 22, no. 5: 2365. https://doi.org/10.3390/ijms22052365
APA StyleMentegari, E., Bertoletti, F., Kissova, M., Zucca, E., Galli, S., Tagliavini, G., Garbelli, A., Maffia, A., Bione, S., Ferrari, E., d’Adda di Fagagna, F., Francia, S., Sabbioneda, S., Chen, L. -Y., Lingner, J., Bergoglio, V., Hoffmann, J. -S., Hübscher, U., Crespan, E., & Maga, G. (2021). A Role for Human DNA Polymerase λ in Alternative Lengthening of Telomeres. International Journal of Molecular Sciences, 22(5), 2365. https://doi.org/10.3390/ijms22052365