
Human Herpesvirus-6 and -7 in the Brain Microenvironment of Persons with Neurological Pathology and Healthy People



Abstract
:1. Introduction
2. Results
2.1. Nested and Real Time Polymerase Chain Reactions
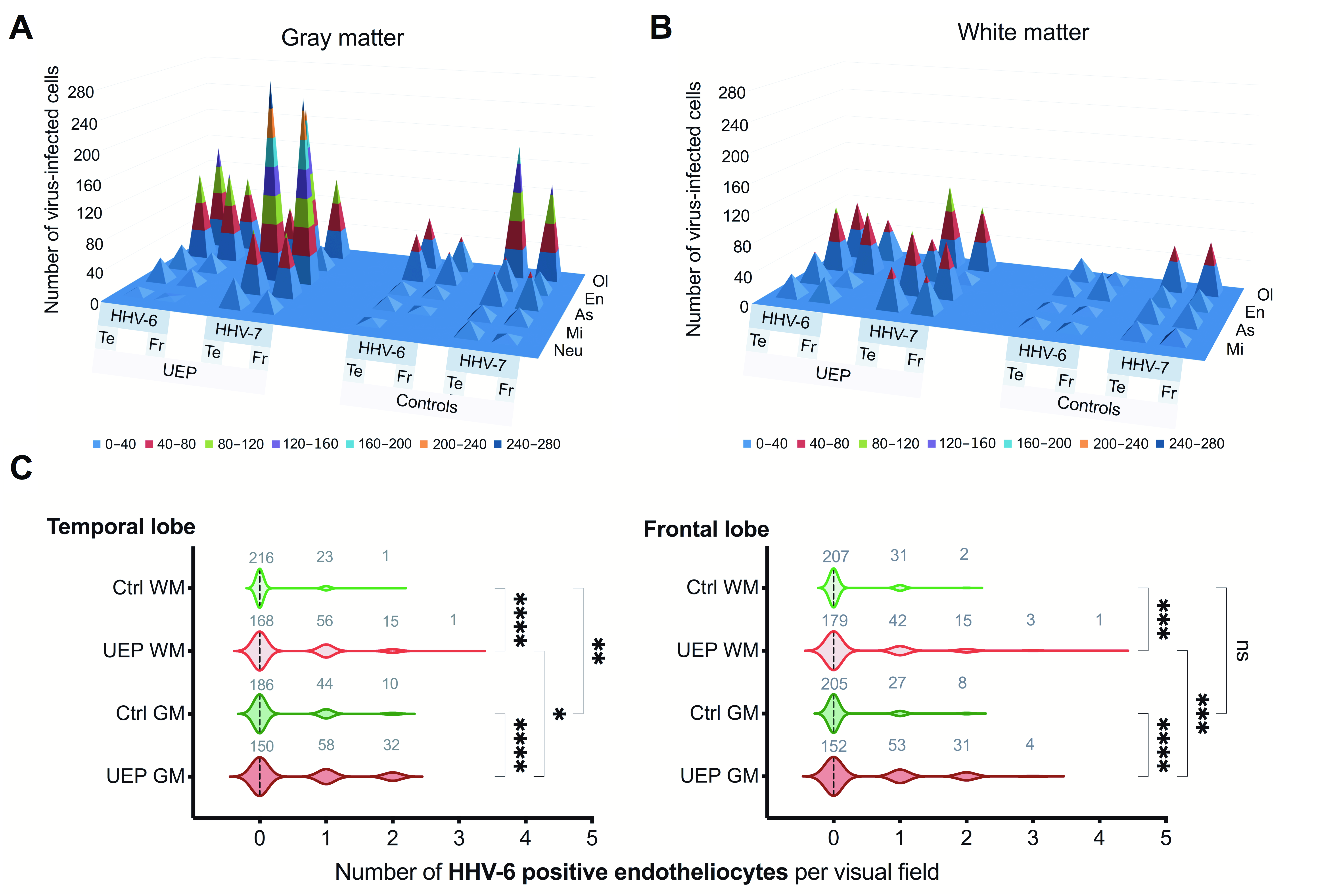
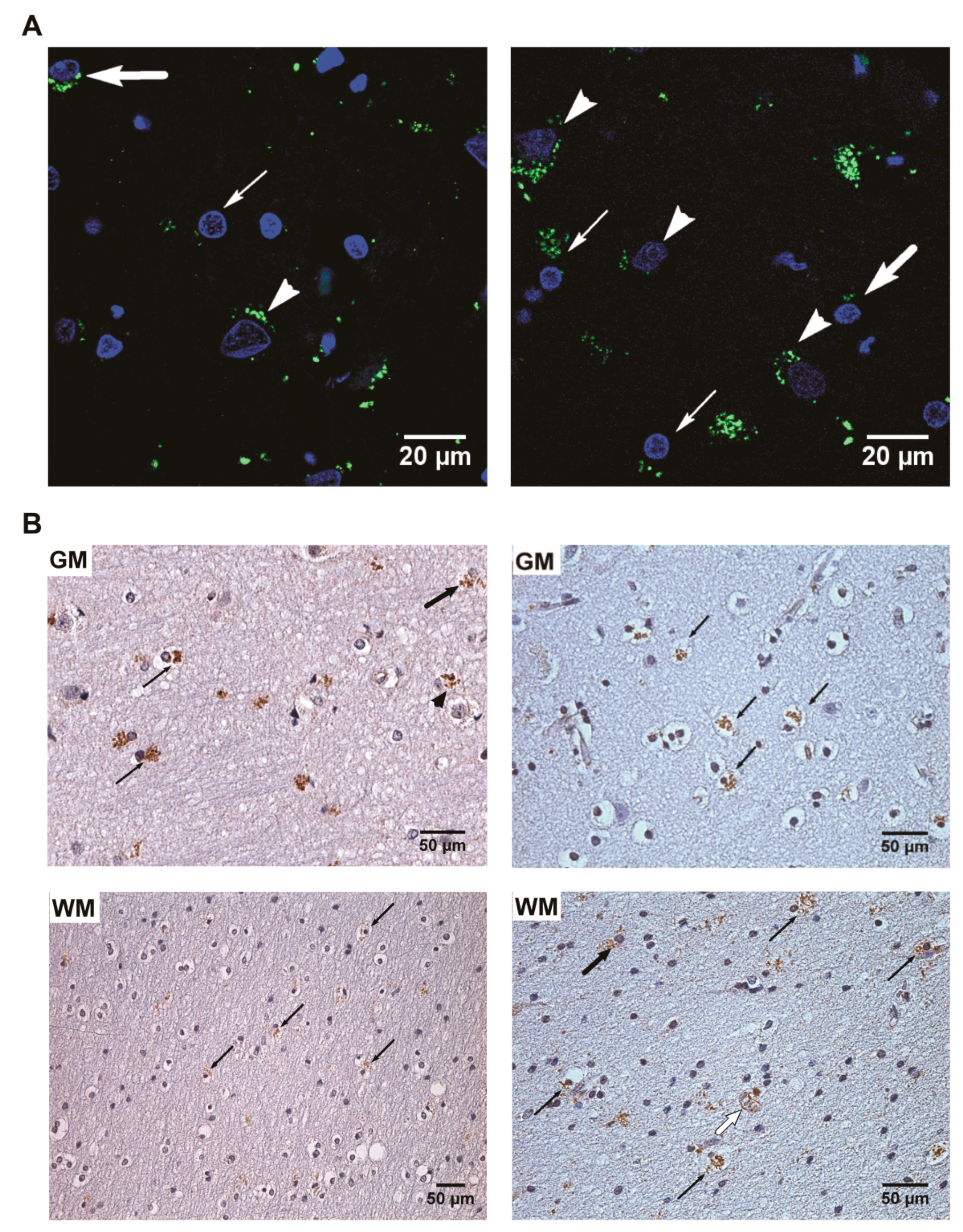
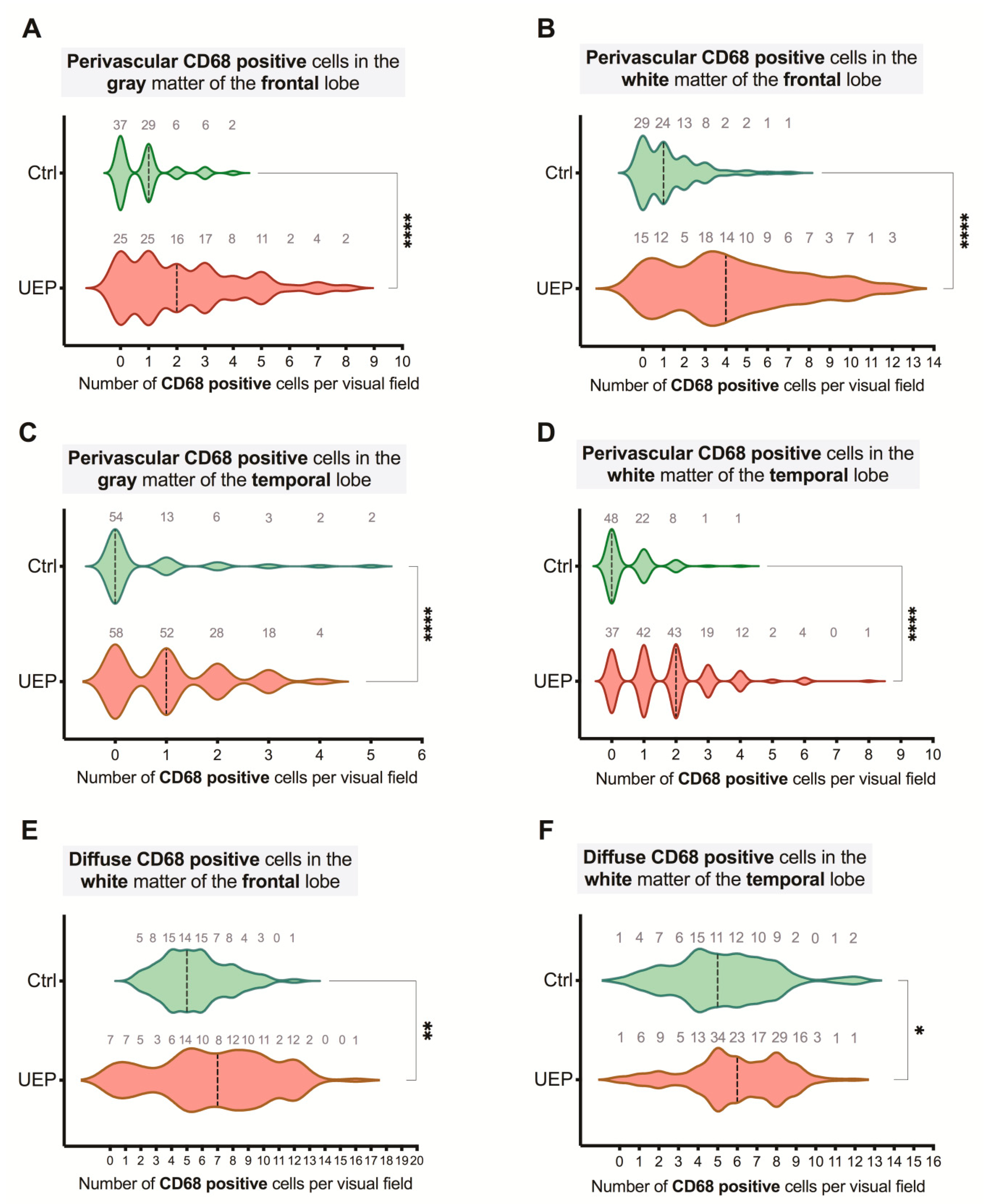
2.2. Immunohistochemistry
3. Discussion
4. Materials and Methods
4.1. Tissue Material and Sampling
4.2. Nested and Real Time Polymerase Chain Reactions
4.3. Immunohistochemistry
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Ablashi, D.; Agut, H.; Alvarez-Lafuente, R.; Clark, D.A.; Dewhurst, S.; DiLuca, D.; Flamand, L.; Frenkel, N.; Gallo, R.; Gompels, U.A.; et al. Classification of HHV-6A and HHV-6B as distinct viruses. Arch. Virol. 2014, 159, 863–870. [Google Scholar] [CrossRef]
- Reynaud, J.M.; Jégou, J.-F.; Welsch, J.C.; Horvat, B. Human herpesvirus 6A infection in CD46 transgenic mice: Viral persistence in the brain and increased production of proinflammatory chemokines via Toll-like receptor 9. J. Virol. 2014, 88, 5421–5436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoeckle, M.Y. The Spectrum of Human Herpesvirus 6 Infection: From Roseola Infantum to Adult Disease. Annu. Rev. Med. 2000, 51, 423–430. [Google Scholar] [CrossRef]
- Stone, R.C.; Micali, G.A.; Schwartz, R.A. Roseola infantum and its causal human herpesviruses. Int. J. Dermatol. 2014, 53, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Wang, X.; Wang, Y.; Wang, P.; Fan, D.; Chen, S.; Guan, Y.; Li, T.; An, J.; Luan, G. Detection of EBV and HHV6 in the Brain Tissue of Patients with Rasmussen’s Encephalitis. Virol. Sin. 2018, 33, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Eliassen, E.; Lum, E.; Pritchett, J.; Ongradi, J.; Krueger, G.; Crawford, J.R.; Phan, T.L.; Ablashi, D.; Hudnall, S.D. Human herpesvirus 6 and malignancy: A review. Front. Oncol. 2018, 8, 512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, N.; Kharlamova, N.; Fogdell-Hahn, A. The role of herpesvirus 6A and 6B in multiple sclerosis and epilepsy. Scand. J. Immunol. 2020, 92, 1–7. [Google Scholar] [CrossRef]
- Santpere, G.; Telford, M.; Andrés-Benito, P.; Navarro, A.; Ferrer, I. The presence of human herpesvirus 6 in the brain in health and disease. Biomolecules 2020, 10, 1520. [Google Scholar] [CrossRef]
- Eimer, W.A.; Vijaya Kumar, D.K.; Navalpur Shanmugam, N.K.; Rodriguez, A.S.; Mitchell, T.; Washicosky, K.J.; György, B.; Breakefield, X.O.; Tanzi, R.E.; Moir, R.D. Alzheimer’s Disease-Associated β-Amyloid Is Rapidly Seeded by Herpesviridae to Protect against Brain Infection. Neuron 2018, 99, 56–63.e3. [Google Scholar] [CrossRef] [Green Version]
- Readhead, B.; Haure-Mirande, J.V.; Funk, C.C.; Richards, M.A.; Shannon, P.; Haroutunian, V.; Sano, M.; Liang, W.S.; Beckmann, N.D.; Price, N.D.; et al. Multiscale Analysis of Independent Alzheimer’s Cohorts Finds Disruption of Molecular, Genetic, and Clinical Networks by Human Herpesvirus. Neuron 2018, 99, 64–82.e7. [Google Scholar] [CrossRef] [Green Version]
- Opsahl, M.L.; Kennedy, P.G.E. Early and late HHV-6 gene transcripts in multiple sclerosis lesions and normal appearing white matter. Brain 2005, 128, 516–527. [Google Scholar] [CrossRef] [Green Version]
- Hogestyn, J.M.; Mock, D.J.; Mayer-Proschel, M. Contributions of neurotropic human herpesviruses herpes simplex virus 1 and human herpesvirus 6 to neurodegenerative disease pathology. Neural Regen. Res. 2018, 13, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Strausbaugh, L.J.; Caserta, M.T.; Mock, D.J.; Dewhurst, S. Human Herpesvirus 6. Clin. Infect. Dis. 2001, 33, 829–833. [Google Scholar] [CrossRef] [Green Version]
- Tyler, K.L. Human Herpesvirus 6 and Multiple Sclerosis: The Continuing Conundrum. J. Infect. Dis. 2003, 187, 1360–1364. [Google Scholar] [CrossRef] [Green Version]
- Prusty, B.K.; Gulve, N.; Govind, S.; Krueger, G.R.F.; Feichtinger, J.; Larcombe, L.; Aspinall, R.; Ablashi, D.V.; Toro, C.T. Active HHV-6 infection of cerebellar Purkinje cells in mood disorders. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, N.; Oka, N.; Takahashi, M.; Shimada, K.; Ishii, A.; Tatebayashi, Y.; Shigeta, M.; Yanagisawa, H.; Kondo, K. Human Herpesvirus 6B Greatly Increases Risk of Depression by Activating Hypothalamic-Pituitary-Adrenal Axis during Latent Phase of Infection. iScience 2020, 23, 101187. [Google Scholar] [CrossRef] [PubMed]
- De Bolle, L.; Naesens, L.; De Clercq, E. Update on human herpesvirus 6 biology, clinical features, and therapy. Clin. Microbiol. Rev. 2005, 18, 217–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, D.A.; Griffiths, P.D. Human herpesvirus 6: Relevance of infection in the immunocompromised host. Br. J. Haematol. 2003, 120, 384–395. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.-Z.; Pellett, P.E. HHV-6A, 6B, and 7: Immunobiology and host response. In Human Herpesviruses; Arvin, A., Campadelli-Fiume, G., Mocarski, E., Moore, P.S., Roizman, B., Whitley, R., Yamanishi, K., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 850–874. [Google Scholar]
- Taródi, B. Human herpesvirus 6 and human herpesvirus 7. In Latency Strategies of Herpesviruses; Minarovits, J., Gonczol, E., Valyi-Nagy, T., Eds.; Springer: New York, NY, USA, 2007; pp. 86–101. ISBN 9780387341279. [Google Scholar]
- Traylen, C.M.; Patel, H.R.; Fondaw, W.; Mahatme, S.; Williams, J.F.; Walker, L.R.; Dyson, O.F.; Arce, S.; Akula, S.M. Virus reactivation: A panoramic view in human infections. Future Virol. 2011, 6, 451–463. [Google Scholar] [CrossRef] [Green Version]
- Reynaud, J.M.; Horvat, B. Human Herpesvirus 6 and Neuroinflammation. ISRN Virol. 2013, 2013, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Sehrawat, S.; Kumar, D.; Rouse, B.T. Herpesviruses: Harmonious pathogens but relevant cofactors in other diseases? Front. Cell. Infect. Microbiol. 2018, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Leibovitch, E.C.; Caruso, B.; Ha, S.K.; Schindler, M.K.; Lee, N.J.; Luciano, N.J.; Billioux, B.J.; Guy, J.R.; Yen, C.; Sati, P.; et al. Herpesvirus trigger accelerates neuroinflammation in a nonhuman primate model of multiple sclerosis. Proc. Natl. Acad. Sci. USA 2018, 115, 11292–11297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Zhong, D.; Li, G. The role of microglia in viral encephalitis: A review. J. Neuroinflamm. 2019, 16, 1–12. [Google Scholar] [CrossRef]
- Bortolotti, D.; Gentili, V.; Rotola, A.; Caselli, E.; Rizzo, R. HHV-6A infection induces amyloid-beta expression and activation of microglial cells. Alzheimer’s Res. Ther. 2019, 11, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hüfner, K.; Arbusow, V.; Himmelein, S.; Derfuss, T.; Sinicina, I.; Strupp, M.; Brandt, T.; Theil, D. The prevalence of human herpesvirus 6 in human sensory ganglia and its co-occurrence with alpha-herpesviruses. J. Neurovirol. 2007, 13, 462–467. [Google Scholar] [CrossRef]
- Ptaszynska-Sarosiek, I.; Dunaj, J.; Zajkowska, A.; Niemcunowicz-Janica, A.; Król, M.; Pancewicz, S.; Zajkowska, J. Post-mortem detection of six human herpesviruses (HSV-1, HSV-2, VZV, EBV, CMV, HHV-6) in trigeminal and facial nerve ganglia by PCR. PeerJ 2019, 2019, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Koyuncu, O.O.; Hogue, I.B.; Enquist, L.W. Virus infections in the nervous system. Cell Host Microbe 2013, 13, 379–393. [Google Scholar] [CrossRef] [Green Version]
- Bello-Morales, R.; Andreu, S.; López-Guerrero, J.A. The role of herpes simplex virus type 1 infection in demyelination of the central nervous system. Int. J. Mol. Sci. 2020, 21, 5026. [Google Scholar] [CrossRef]
- Martin, J.R.; Mitchell, W.J.; Henken, D.B. Neurotropic Herpesviruses, Neural Mechanisms and Arteritis. Brain Pathol. 1990, 1, 6–10. [Google Scholar] [CrossRef]
- Albright, A.V.; Lavi, E.; Black, J.B.; Goldberg, S.; O’Connor, M.J.; González-Scarano, F. The effect of human herpesvirus-6 (HHV-6) on cultured human neural cells: Oligodendrocytes and microglia. J. Neurovirol. 1998, 4, 486–494. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Asano, Y.; Ihira, M.; Suzuki, K.; Ohashi, M.; Suga, S.; Kudo, K.; Horibe, K.; Kojima, S.; Kato, K.; et al. Human Herpesvirus 6 Viremia in Bone Marrow Transplant Recipients: Clinical Features and Risk Factors. J. Infect. Dis. 2002, 185, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Ahlqvist, J.; Fotheringham, J.; Akhyani, N.; Yao, K.; Fogdell-Hahn, A.; Jacobson, S. Differential tropism of human herpesvirus 6 (HHV-6) variants and induction of latency by HHV-6A in oligodendrocytes. J. Neurovirol. 2005, 11, 384–394. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.; Crawford, J.R.; Komaroff, A.L.; Ablashi, D.V.; Jacobson, S. Review part 2: Human herpesvirus-6 in central nervous system diseases. J. Med. Virol. 2010, 82, 1669–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campadelli-Fiume, G.; Mirandola, P.; Menotti, L. Human herpesvirus 6: An emerging pathogen. Emerg. Infect. Dis. 1999, 5, 353–366. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Muñoz, M.E.; Fuentes-Pananá, E.M. Beta and gamma human herpesviruses: Agonistic and antagonistic interactions with the host immune system. Front. Microbiol. 2018, 8, 2521. [Google Scholar] [CrossRef]
- Cassiani-Ingoni, R.; Greenstone, H.L.; Donati, D.; Fogdell-Hahn, A.; Martinelli, E.; Refai, D.; Martin, R.; Berger, E.A.; Jacobson, S. CD46 on glial cells can function as a receptor for viral glycoprotein-mediated cell-cell fusion. Glia 2005, 52, 252–258. [Google Scholar] [CrossRef]
- Greenstone, H.L.; Santoro, F.; Lusso, P.; Berger, E.A. Human herpesvirus 6 and measles virus employ distinct CD46 domains for receptor function. J. Biol. Chem. 2002, 277, 39112–39118. [Google Scholar] [CrossRef] [Green Version]
- Pantry, S.N.; Medveczky, P.G. Latency, integration, and reactivation of human herpesvirus-6. Viruses 2017, 9, 194. [Google Scholar] [CrossRef] [Green Version]
- Lusso, P.; Secchiero, P.; Crowley, R.W.; Garzino-Demo, A.; Berneman, Z.N.; Gallo, R.C. CD4 is a critical component of the receptor for human herpesvirus 7: Interference with human immunodeficiency virus. Proc. Natl. Acad. Sci. USA 1994, 91, 3872–3876. [Google Scholar] [CrossRef] [Green Version]
- Krummenacher, C.; Carfí, A.; Eisenberg, R.J.; Cohen, G.H. Entry of herpesviruses into cells: The enigma variations. Adv. Exp. Med. Biol. 2013, 790, 178–195. [Google Scholar] [CrossRef]
- Eisenberg, R.J.; Atanasiu, D.; Cairns, T.M.; Gallagher, J.R.; Krummenacher, C.; Cohen, G.H. Herpes virus fusion and entry: A story with many characters. Viruses 2012, 4, 800–832. [Google Scholar] [CrossRef] [PubMed]
- Sobhy, H. A comparative review of viral entry and attachment during large and giant dsDNA virus infections. Arch. Virol. 2017, 162, 3567–3585. [Google Scholar] [CrossRef] [PubMed]
- Donati, D.; Martinelli, E.; Cassiani-Ingoni, R.; Ahlqvist, J.; Hou, J.; Major, E.O.; Jacobson, S. Variant-Specific Tropism of Human Herpesvirus 6 in Human Astrocytes. J. Virol. 2005, 79, 9439–9448. [Google Scholar] [CrossRef] [Green Version]
- Reynaud, J.M.; Horvat, B. Animal models for human herpesvirus 6 infection. Front. Microbiol. 2013, 4, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allnutt, M.A.; Johnson, K.; Bennett, D.A.; Connor, S.M.; Troncoso, J.C.; Pletnikova, O.; Albert, M.S.; Resnick, S.M.; Scholz, S.W.; De Jager, P.L.; et al. Human Herpesvirus 6 Detection in Alzheimer’s Disease Cases and Controls across Multiple Cohorts. Neuron 2020, 105, 1027–1035.e2. [Google Scholar] [CrossRef] [PubMed]
- Knox, K.K.; Harrington, D.P.; Carrigan, D.R. Fulminant human herpesvirus six encephalitis in a human immunodeficiency virus-infected infant. J. Med. Virol. 1995, 45, 288–292. [Google Scholar] [CrossRef]
- Skuja, S.; Zieda, A.; Ravina, K.; Chapenko, S.; Roga, S.; Teteris, O.; Groma, V.; Murovska, M. Structural and ultrastructural alterations in human olfactory pathways and possible associations with herpesvirus 6 infection. PLoS ONE 2017, 12, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agut, H.; Bonnafous, P.; Gautheret-Dejean, A. Laboratory and clinical aspects of human herpesvirus 6 infections. Clin. Microbiol. Rev. 2015, 28, 313–335. [Google Scholar] [CrossRef] [Green Version]
- Becerra, A.; Gibson, L.; Stern, L.J.; Calvo-Calle, J.M. Immune response to HHV-6 and implications for immunotherapy. Curr. Opin. Virol. 2014, 9, 154–161. [Google Scholar] [CrossRef] [Green Version]
- Nikitina, E.; Larionova, I.; Choinzonov, E.; Kzhyshkowska, J. Monocytes and macrophages as viral targets and reservoirs. Int. J. Mol. Sci. 2018, 19, 2821. [Google Scholar] [CrossRef] [Green Version]
- Cone, R.W.; Huang, M.L.W.; Ashley, R.; Corey, L. Human herpesvirus 6 DNA in peripheral blood cells and saliva from immunocompetent individuals. J. Clin. Microbiol. 1993, 31, 1262–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lusso, P. HHV-6 and the immune system: Mechanisms of immunomodulation and viral escape. J. Clin. Virol. 2006, 37 (Suppl. 1), 4–10. [Google Scholar] [CrossRef]
- Wang, F.; Chi, J.; Peng, G.; Zhou, F.; Wang, J.; Li, L.; Feng, D.; Xie, F.; Gu, B.; Qin, J.; et al. Development of Virus-Specific CD4+ and CD8+ Regulatory T Cells Induced by Human Herpesvirus 6 Infection. J. Virol. 2014, 88, 1011–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, D.J.; Hill, J.A.; Koelle, D.M. Advances in the characterization of the T-cell response to human herpesvirus-6. Front. Immunol. 2018, 9, 4–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkins, B.T.; Davis, T.P. The blood-brain barrier/neurovascular unit in health and disease. Pharmacol. Rev. 2005, 57, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.K.; Jin, H.K.; Jeong, A.P.; Lee, S.W.; Woo, J.K.; Young, S.Y.; Kim, K.W. Blood-neural barrier: Intercellular communication at glio-vascular interface. J. Biochem. Mol. Biol. 2006, 39, 339–345. [Google Scholar] [CrossRef] [Green Version]
- Domingues, H.S.; Portugal, C.C.; Socodato, R.; Relvas, J.B. Oligodendrocyte, astrocyte, and microglia crosstalk in myelin development, damage, and repair. Front. Cell Dev. Biol. 2016, 4, 1–16. [Google Scholar] [CrossRef]
- Chapenko, S.; Roga, S.; Skuja, S.; Rasa, S.; Cistjakovs, M.; Svirskis, S.; Zaserska, Z.; Groma, V.; Murovska, M. Detection frequency of human herpesviruses-6A, -6B, and -7 genomic sequences in central nervous system DNA samples from post-mortem individuals with unspecified encephalopathy. J. Neurovirol. 2016, 22, 488–497. [Google Scholar] [CrossRef]
- Secchiero, P.; Carrigan, D.R.; Asano, Y.; Benedetti, L.; Crowley, R.W.; Komaroff, A.L.; Gallo, R.C.; Lusso, P. Detection of human herpesvirus 6 in plasma of children with primary infection and immunosuppressed patients by polymerase chain reaction. J. Infect. Dis. 1995, 171, 273–280. [Google Scholar] [CrossRef]
- Berneman, Z.N.; Ablashi, D.V.; Li, G.; Eger-Fletcher, M.; Reitz, M.S.; Hung, C.L.; Brus, I.; Komaroff, A.L.; Gallo, R.C. Human herpesvirus 7 is a T-lymphotropic virus and is related to, but significantly different from, human herpesvirus 6 and human cytomegalovirus. Proc. Natl. Acad. Sci. USA 1992, 89, 10552–10556. [Google Scholar] [CrossRef] [Green Version]
- Skuja, S.; Vilmane, A.; Svirskis, S.; Groma, V.; Murovska, M. Evidence of human parvovirus B19 infection in the post-mortem brain tissue of the elderly. Viruses 2018, 10, 582. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Lobe | Samples Containing Viral Genomic Sequences (n) | |||||
|---|---|---|---|---|---|---|---|
| n | HHV-6 (a) | HHV-7 (b) | HHV-6 + HHV-7 (c) | Total ((a+b)-c) | p-Value (Chi2) (vs. Control) | ||
| UEP | Frontal | 24 | 9 | 4 | 2 | 11 | 0.3759 |
| Temporal | 24 | 11 | 6 | 1 | 16 | 0.0209 | |
| Controls | Frontal | 24 | 4 | 4 | 0 | 8 | |
| Temporal | 24 | 7 | 4 | 3 | 8 | ||
| Groups | Lobes | Viral Genomic Sequence | Gray Matter | White Matter | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Neu (n) | Mi (n) | As (n) | En (n) | Ol (n) | Total (n) | Mi (n) | As (n) | En (n) | Ol (n) | Total (n) | |||
| UEP | Te | HHV-6 | 10 | 32 | 33 | 122 | 148 | 345 | 30 | 42 | 89 | 79 | 240 |
| Fr | HHV-6 | 2 | 21 | 26 | 127 | 105 | 281 | 31 | 21 | 85 | 58 | 195 | |
| Te | HHV-7 | 39 | 86 | 278 | 85 | 218 | 706 | 55 | 86 | 57 | 116 | 314 | |
| Fr | HHV-7 | 27 | 87 | 257 | 33 | 115 | 519 | 49 | 76 | 15 | 89 | 229 | |
| Controls | Te | HHV-6 | 7 | 15 | 15 | 64 | 70 | 171 | 11 | 9 | 25 | 30 | 75 |
| Fr | HHV-6 | 0 | 7 | 15 | 43 | 47 | 112 | 10 | 9 | 35 | 15 | 69 | |
| Te | HHV-7 | 10 | 18 | 42 | 44 | 184 | 298 | 12 | 30 | 26 | 61 | 129 | |
| Fr | HHV-7 | 8 | 20 | 48 | 29 | 134 | 239 | 18 | 29 | 26 | 71 | 144 | |
| Groups | Lobes | Viral Genomic Sequence | Gray Matter | White Matter | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Neu (n) | Mi (n) | As (n) | En (n) | OL (n) | Total (n) | Mi (n) | As (n) | En (n) | Ol (n) | Total (n) | |||
| UEP | Te | HHV-6 | 10 | 24 | 28 | 70 | 124 | 256 | 20 | 39 | 61 | 63 | 183 |
| Fr | HHV-6 | 2 | 12 | 21 | 55 | 56 | 146 | 17 | 15 | 31 | 27 | 90 | |
| Te | HHV-7 | 19 | 61 | 96 | 41 | 91 | 308 | 35 | 39 | 26 | 47 | 147 | |
| Fr | HHV-7 | 19 | 36 | 66 | 18 | 41 | 180 | 24 | 24 | 10 | 18 | 76 | |
| Controls | Te | HHV-6 | 5 | 12 | 9 | 33 | 65 | 124 | 8 | 9 | 10 | 30 | 57 |
| Fr | HHV-6 | 0 | 1 | 7 | 23 | 36 | 67 | 2 | 7 | 12 | 8 | 29 | |
| Te | HHV-7 | 3 | 6 | 20 | 7 | 56 | 92 | 1 | 2 | 4 | 11 | 18 | |
| Fr | HHV-7 | 1 | 9 | 26 | 9 | 41 | 86 | 12 | 10 | 9 | 19 | 50 | |
| Group | Lobe | Gray Matter | White Matter | ||||
|---|---|---|---|---|---|---|---|
| Diffuse (n) | Perivascular (n) | Total (n) | Diffuse (n) | Perivascular (n) | Total (n) | ||
| UEP | Fr | 314 | 466 | 780 | 1458 | 864 | 2322 |
| Te | 431 | 245 | 676 | 1289 | 376 | 1665 | |
| Controls | Fr | 273 | 193 | 466 | 901 | 323 | 1224 |
| Te | 284 | 185 | 469 | 842 | 233 | 1075 | |
| Group | Lobe | Gray Matter | White Matter | ||||
|---|---|---|---|---|---|---|---|
| Diffuse (n) | Perivascular (n) | Total (n) | Diffuse (n) | Perivascular (n) | Total (n) | ||
| UEP | Fr_HHV+ | 188 | 251 | 439 | 740 | 478 | 1218 |
| Fr_HHV- | 126 | 215 | 341 | 718 | 386 | 1104 | |
| Te_HHV+ | 325 | 178 | 503 | 947 | 275 | 1222 | |
| Te_HHV- | 106 | 67 | 173 | 342 | 101 | 443 | |
| Controls | Fr_HHV+ | 142 | 67 | 209 | 445 | 105 | 550 |
| Fr_HHV- | 131 | 126 | 257 | 456 | 218 | 674 | |
| Te_HHV+ | 170 | 58 | 228 | 418 | 42 | 460 | |
| Te_HHV- | 114 | 127 | 241 | 424 | 191 | 615 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skuja, S.; Svirskis, S.; Murovska, M. Human Herpesvirus-6 and -7 in the Brain Microenvironment of Persons with Neurological Pathology and Healthy People. Int. J. Mol. Sci. 2021, 22, 2364. https://doi.org/10.3390/ijms22052364
Skuja S, Svirskis S, Murovska M. Human Herpesvirus-6 and -7 in the Brain Microenvironment of Persons with Neurological Pathology and Healthy People. International Journal of Molecular Sciences. 2021; 22(5):2364. https://doi.org/10.3390/ijms22052364
Chicago/Turabian StyleSkuja, Sandra, Simons Svirskis, and Modra Murovska. 2021. "Human Herpesvirus-6 and -7 in the Brain Microenvironment of Persons with Neurological Pathology and Healthy People" International Journal of Molecular Sciences 22, no. 5: 2364. https://doi.org/10.3390/ijms22052364
APA StyleSkuja, S., Svirskis, S., & Murovska, M. (2021). Human Herpesvirus-6 and -7 in the Brain Microenvironment of Persons with Neurological Pathology and Healthy People. International Journal of Molecular Sciences, 22(5), 2364. https://doi.org/10.3390/ijms22052364