Green Synthesis of Selenium and Tellurium Nanoparticles: Current Trends, Biological Properties and Biomedical Applications

 ,
, 

 ,
,  ,
,  and
and 
Abstract
:
1. Introduction
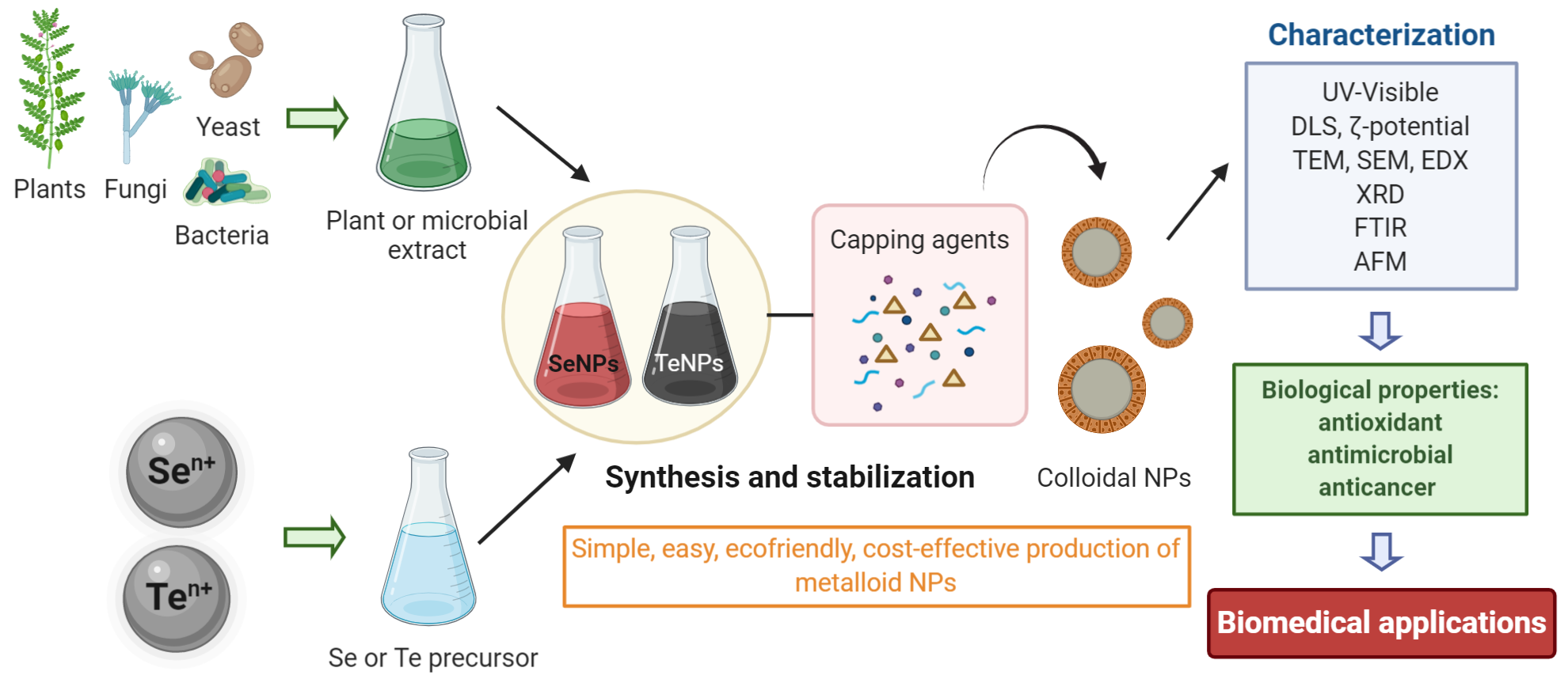
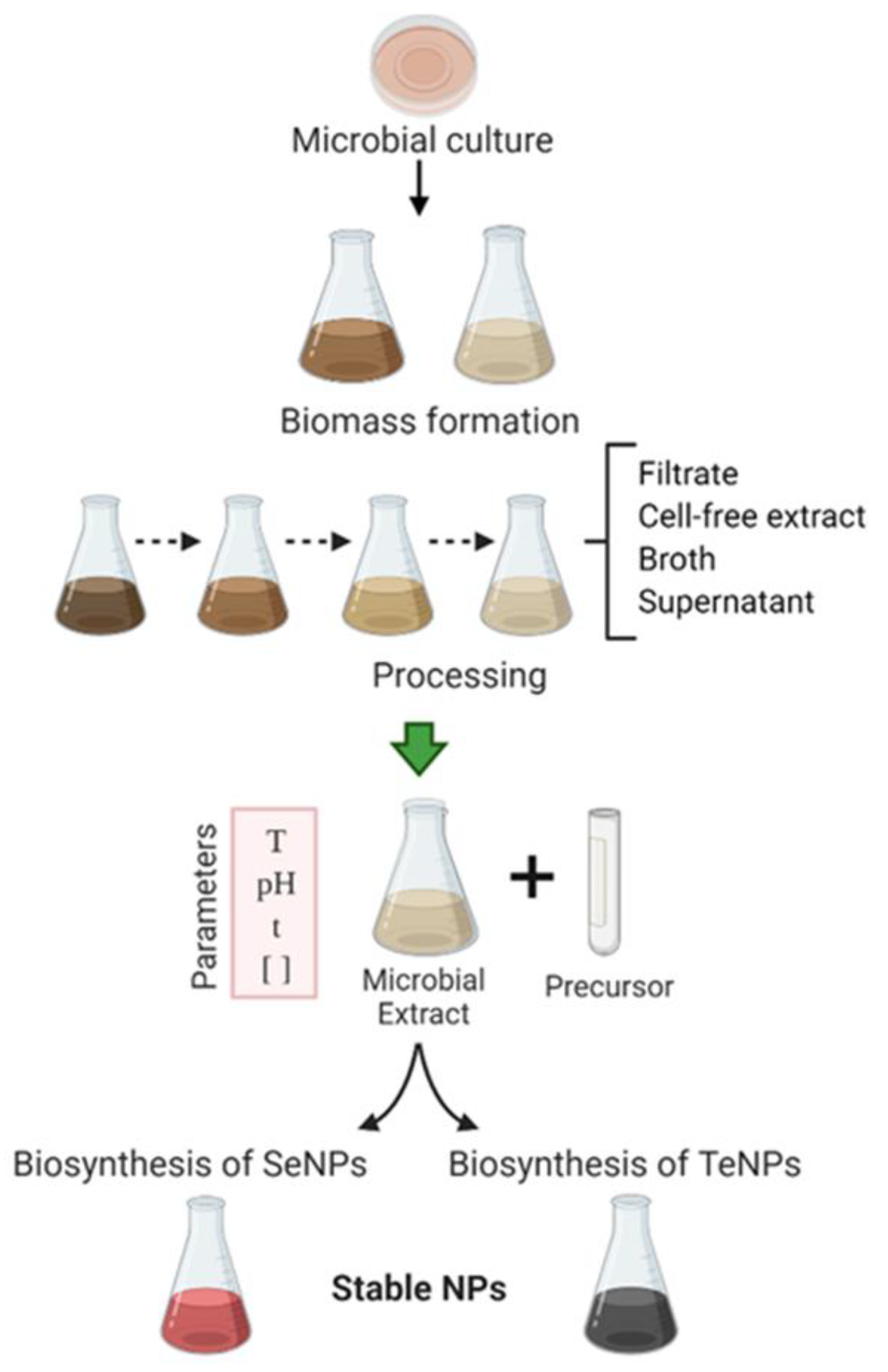
2. Green Synthesis of Inorganic Nanoparticles Using Microorganisms
3. Parameters Affecting the Green Synthesis of Metalloid Nanoparticles
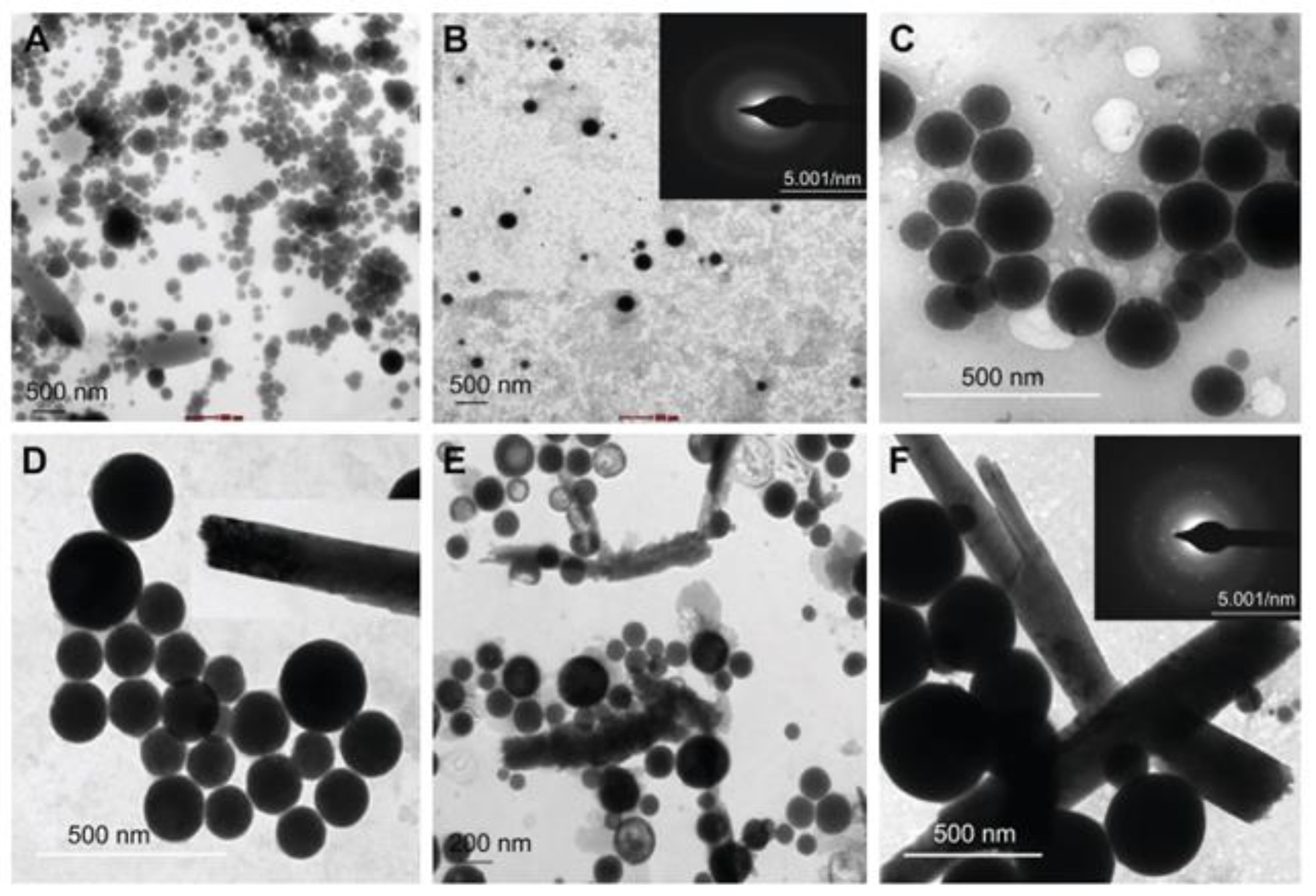
4. Techniques of Characterization
5. Microbial Biosynthesis of Selenium Nanoparticles
5.1. Using Bacteria
5.2. Using Fungi
5.3. Using Yeast
6. Microbial Synthesis of Tellurium Nanoparticles
7. Plant-Mediated Synthesis of Metalloid Nanoparticles
7.1. Plant-Based Synthesis of Selenium Nanoparticles
7.2. Plant-Based Synthesis of Tellurium Nanoparticles
8. Biosynthesis of Bimetallic Se-Te Alloy Nanoparticles
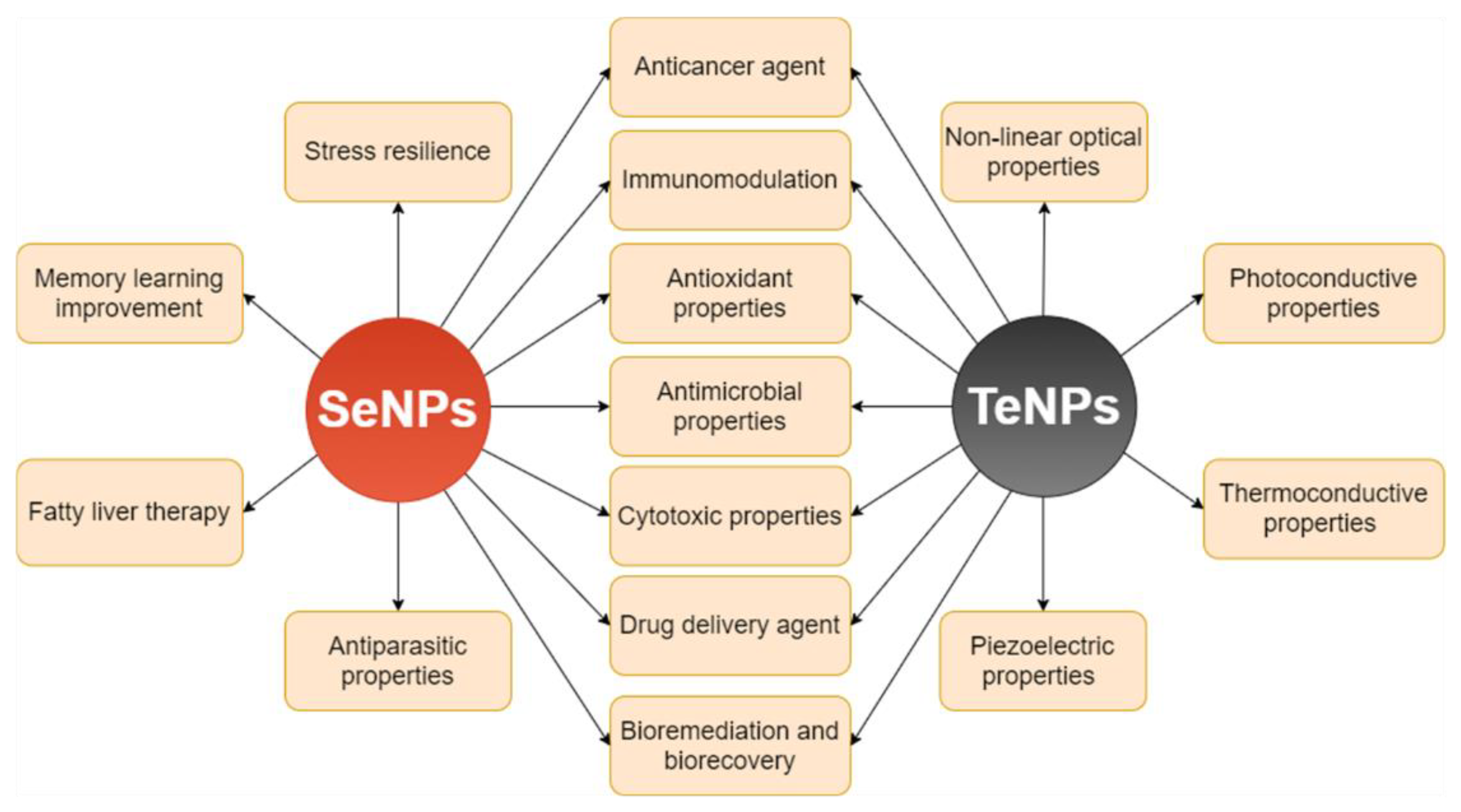
9. Bioapplications of SeNPs and TeNPs
10. Human Cell-Cytotoxicity and Immune Response Induced by SeNPs and TeNPs
11. Conclusions and Perspectives
Funding
Conflicts of Interest
References
- Hosnedlova, B.; Kepinska, M.; Skalickova, S.; Fernandez, C.; Ruttkay-Nedecky, B.; Peng, Q.; Baron, M.; Melcova, M.; Opatrilova, R.; Zidkova, J.; et al. Nano-selenium and its nanomedicine applications: A critical review. Int. J. Nanomed. 2018, 13, 2107–2128. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xu, H.; Chen, Z.-S.; Chen, G. Biosynthesis of Nanoparticles by Microorganisms and Their Applications. J. Nanomater. 2011, 2011, 270974. [Google Scholar] [CrossRef] [Green Version]
- Rashidi, L.; Khosravi-Darani, K. The applications of nanotechnology in food industry. Crit. Rev. Food Sci. Nutr. 2011, 51, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Stark, W.J.; Stoessel, P.R.; Wohlleben, W.; Hafner, A. Industrial Applications of Nanoparticles. Chem. Soc. Rev. 2015, 44, 5793–5805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.; Nalwa, H.S. Medical applications of nanoparticles in biological imaging, cell labeling, antimicrobial agents, and anticancer nanodrugs. J. Biomed. Nanotechnol. 2011, 7, 489–503. [Google Scholar] [CrossRef] [PubMed]
- Sakr, T.M.; Korany, M.; Katti, K.V. Selenium nanomaterials in biomedicine—An overview of new opportunities in nanomedicine of selenium. J. Drug Deliv. Sci. Technol. 2018, 46, 223–233. [Google Scholar] [CrossRef]
- Brito-Silva, A.M.; Gómez, L.A.; de Araújo, C.B.; Galembeck, A. Laser Ablated Silver Nanoparticles with Nearly the Same Size in Different Carrier Media. J. Nanomater. 2010, 2010, 142897. [Google Scholar] [CrossRef]
- Nee, C.H.; Yap, S.L.; Tou, T.Y.; Chang, H.C.; Yap, S.S. Direct synthesis of nanodiamonds by femtosecond laser irradiation of ethanol. Sci. Rep. 2016, 6, 33966. [Google Scholar] [CrossRef]
- Anbarasu, M.; Anandan, M.; Chinnasamy, E.; Gopinath, V.; Balamurugan, K. Synthesis and characterization of polyethylene glycol (PEG) coated Fe3O4 nanoparticles by chemical co-precipitation method for biomedical applications. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2015, 135, 536–539. [Google Scholar] [CrossRef]
- Janjua, M.R.S.A. Synthesis of Co3O4 Nano Aggregates by Co-precipitation Method and its Catalytic and Fuel Additive Applications. Open Chem. 2019, 17, 865–873. [Google Scholar] [CrossRef]
- Li, M.; Gu, L.; Li, T.; Hao, S.; Tan, F.; Chen, D.; Zhu, D.; Xu, Y.; Sun, C.; Yang, Z. TiO2-Seeded Hydrothermal Growth of Spherical BaTiO3 Nanocrystals for Capacitor Energy-Storage Application. Crystals 2020, 10, 202. [Google Scholar] [CrossRef] [Green Version]
- Clark, I.; Smith, J.; Gomes, R.L.; Lester, E. Towards the Continuous Hydrothermal Synthesis of ZnO@Mg2Al-CO3 Core-Shell Composite Nanomaterials. Nanomaterials 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Yang, H.; Rong, K.; Lu, Z.; Yu, X.; Chen, R. Shape-controlled solvothermal synthesis of bismuth subcarbonate nanomaterials. J. Solid State Chem. 2010, 183, 1878–1883. [Google Scholar] [CrossRef]
- Yang, S.; Zhou, X.; Zhang, J.; Liu, Z. Morphology-controlled solvothermal synthesis of LiFePO4 as a cathode material for lithium-ion batteries. J. Mater. Chem. 2010, 20, 8086–8091. [Google Scholar] [CrossRef]
- Riccò, R.; Nizzero, S.; Penna, E.; Meneghello, A.; Cretaio, E.; Enrichi, F. Ultra-small dye-doped silica nanoparticles via modified sol-gel technique. J. Nanopart. Res. 2018, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adnan, R.; Razana, N.A.; Abdul Rahman, I.; Farrukh, M.A. Synthesis and Characterization of High Surface Area Tin Oxide Nanoparticles via the Sol-Gel Method as a Catalyst for the Hydrogenation of Styrene. J. Chin. Chem. Soc. 2010, 57, 222–229. [Google Scholar] [CrossRef]
- Brayner, R.; Dahoumane, S.A.; Nguyen, J.N.; Yepremian, C.; Djediat, C.; Coute, A.; Fievet, F. Ecotoxicological studies of CdS nanoparticles on photosynthetic microorganisms. J. Nanosci. Nanotechnol. 2011, 11, 1852–1858. [Google Scholar] [CrossRef]
- Brayner, R.; Dahoumane, S.A.; Yepremian, C.; Djediat, C.; Meyer, M.; Coute, A.; Fievet, F. ZnO nanoparticles: Synthesis, characterization, and ecotoxicological studies. Langmuir 2010, 26, 6522–6528. [Google Scholar] [CrossRef]
- Khaydarov, R.A.; Khaydarov, R.R.; Gapurova, O.; Estrin, Y.; Scheper, T. Electrochemical method for the synthesis of silver nanoparticles. J. Nanopart. Res. 2008, 11, 1193–1200. [Google Scholar] [CrossRef]
- Starowicz, M.; Starowicz, P.; Zukrowski, J.; Przewoznik, J.; Lemanski, A.; Kapusta, C.; Banas, J. Electrochemical synthesis of magnetic iron oxide nanoparticles with controlled size. J. Nanopart. Res. 2011, 13, 7167–7176. [Google Scholar] [CrossRef] [Green Version]
- Armijo Garcia, D.; Mendoza, L.; Vizuete, K.; Debut, A.; Arias, M.T.; Gavilanes, A.; Terencio, T.; Avila, E.; Jeffryes, C.; Dahoumane, S.A. Sugar-Mediated Green Synthesis of Silver Selenide Semiconductor Nanocrystals under Ultrasound Irradiation. Molecules 2020, 25. [Google Scholar] [CrossRef]
- Yan, Q.; Qiu, M.; Chen, X.; Fan, Y. Ultrasound Assisted Synthesis of Size-Controlled Aqueous Colloids for the Fabrication of Nanoporous Zirconia Membrane. Front. Chem. 2019, 7, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.V.; Bafana, A.P.; Pawar, P.; Faltane, M.; Rahman, A.; Dahoumane, S.A.; Kucknoor, A.; Jeffryes, C.S. Optimized production of antibacterial copper oxide nanoparticles in a microwave-assisted synthesis reaction using response surface methodology. Colloid Surf. A Physicochem. Eng. Aspect 2019, 573, 170–178. [Google Scholar] [CrossRef]
- Bafana, A.; Kumar, S.V.; Temizel-Sekeryan, S.; Dahoumane, S.A.; Haselbach, L.; Jeffryes, C.S. Evaluating microwave-synthesized silver nanoparticles from silver nitrate with life cycle assessment techniques. Sci. Total Environ. 2018, 636, 936–943. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Gomez, B.; Arregui, L.; Serrano, S.; Santos, A.; Perez-Corona, T.; Madrid, Y. Selenium and tellurium-based nanoparticles as interfering factors in quorum sensing-regulated processes: Violacein production and bacterial biofilm formation. Metallomics 2019, 11, 1104–1114. [Google Scholar] [CrossRef]
- Dahoumane, S.A.; Jeffryes, C.; Mechouet, M.; Agathos, S.N. Biosynthesis of Inorganic Nanoparticles: A Fresh Look at the Control of Shape, Size and Composition. Bioengineering 2017, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Rahman, A.; Lin, J.; Jaramillo, F.E.; Bazylinski, D.A.; Jeffryes, C.; Dahoumane, S.A. In Vivo Biosynthesis of Inorganic Nanomaterials Using Eukaryotes-A Review. Molecules 2020, 25. [Google Scholar] [CrossRef]
- Kumar, S.V.; Bafana, A.P.; Pawar, P.; Rahman, A.; Dahoumane, S.A.; Jeffryes, C.S. High conversion synthesis of <10 nm starch-stabilized silver nanoparticles using microwave technology. Sci. Rep. 2018, 8, 5106. [Google Scholar] [CrossRef] [Green Version]
- Rahman, A.; Kumar, S.; Bafana, A.; Lin, J.; Dahoumane, S.A.; Jeffryes, C. A Mechanistic View of the Light-Induced Synthesis of Silver Nanoparticles Using Extracellular Polymeric Substances of Chlamydomonas reinhardtii. Molecules 2019, 24. [Google Scholar] [CrossRef] [Green Version]
- Ramani, M.; Ponnusamy, S.; Muthamizhchelvan, C.; Marsili, E. Amino acid-mediated synthesis of zinc oxide nanostructures and evaluation of their facet-dependent antimicrobial activity. Colloid Surf. B Biointerfaces 2014, 117, 233–239. [Google Scholar] [CrossRef]
- Manivasagan, P.; Oh, J. Marine polysaccharide-based nanomaterials as a novel source of nanobiotechnological applications. Int. J. Biol. Macromol. 2016, 82, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Sharma, A.R.; Bhavesh, R.; Park, J.; Ganbold, B.; Nam, J.S.; Lee, S.S. Biomolecule-mediated synthesis of selenium nanoparticles using dried Vitis vinifera (raisin) extract. Molecules 2014, 19, 2761–2770. [Google Scholar] [CrossRef] [PubMed]
- Kuppusamy, P.; Yusoff, M.M.; Maniam, G.P.; Govindan, N. Biosynthesis of metallic nanoparticles using plant derivatives and their new avenues in pharmacological applications—An updated report. Saudi Pharm. J. 2016, 24, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Kim, Y.J.; Zhang, D.; Yang, D.C. Biological Synthesis of Nanoparticles from Plants and Microorganisms. Trend Biotechnol. 2016, 34, 588–599. [Google Scholar] [CrossRef]
- Jeffryes, C.; Agathos, S.N.; Rorrer, G. Biogenic nanomaterials from photosynthetic microorganisms. Curr. Opin. Biotechnol. 2015, 33, 23–31. [Google Scholar] [CrossRef]
- Dahoumane, S.A.; Mechouet, M.; Wijesekera, K.; Filipe, C.D.M.; Sicard, C.; Bazylinski, D.A.; Jeffryes, C. Algae-mediated biosynthesis of inorganic nanomaterials as a promising route in nanobiotechnology—A review. Green Chem. 2017, 19, 552–587. [Google Scholar] [CrossRef]
- Dahoumane, S.A.; Wujcik, E.K.; Jeffryes, C. Noble metal, oxide and chalcogenide-based nanomaterials from scalable phototrophic culture systems. Enzyme Microb. Technol. 2016, 95, 13–27. [Google Scholar] [CrossRef]
- Dahoumane, S.A.; Djediat, C.; Yéprémian, C.; Couté, A.; Fiévet, F.; Brayner, R. Design of magnetic akaganeite-cyanobacteria hybrid biofilms. Thin Solid Film 2010, 518, 5432–5436. [Google Scholar] [CrossRef]
- Dahoumane, S.A.; Yéprémian, C.; Djédiat, C.; Couté, A.; Fiévet, F.; Coradin, T.; Brayner, R. A global approach of the mechanism involved in the biosynthesis of gold colloids using micro-algae. J. Nanopart. Res. 2014, 16. [Google Scholar] [CrossRef]
- Dahoumane, S.A.; Yéprémian, C.; Djédiat, C.; Couté, A.; Fiévet, F.; Coradin, T.; Brayner, R. Improvement of kinetics, yield, and colloidal stability of biogenic gold nanoparticles using living cells of Euglena gracilis microalga. J. Nanopart. Res. 2016, 18. [Google Scholar] [CrossRef] [Green Version]
- Das, S.K.; Marsili, E. A green chemical approach for the synthesis of gold nanoparticles: Characterization and mechanistic aspect. Rev. Environ. Sci. Biotechnol. 2010, 9, 199–204. [Google Scholar] [CrossRef] [Green Version]
- Gahlawat, G.; Roy Choudhury, A. A review on the biosynthesis of metal and metal salt nanoparticles by microbes. RSC Adv. 2019, 9, 12944–12967. [Google Scholar] [CrossRef] [Green Version]
- Pantidos, N.; Horsfall, L.E. Biological Synthesis of Metallic Nanoparticles by Bacteria, Fungi and Plants. J. Nanomed. Nanotechnol. 2014, 5. [Google Scholar] [CrossRef]
- Kitching, M.; Ramani, M.; Marsili, E. Fungal biosynthesis of gold nanoparticles: Mechanism and scale up. Microb. Biotechnol. 2015, 8, 904–917. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Kumar, S.; Bafana, A.; Dahoumane, S.A.; Jeffryes, C. Biosynthetic Conversion of Ag(+) to highly Stable Ag(0) Nanoparticles by Wild Type and Cell Wall Deficient Strains of Chlamydomonas reinhardtii. Molecules 2019, 24, 98. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Xia, X.; Wu, S.; Zheng, S.; Wang, G. The essentialness of glutathione reductase GorA for biosynthesis of Se(0)-nanoparticles and GSH for CdSe quantum dot formation in Pseudomonas stutzeri TS44. J. Hazard. Mater. 2019, 366, 301–310. [Google Scholar] [CrossRef]
- Chaudhary, S.; Umar, A.; Mehta, S.K. Selenium nanomaterials: An overview of recent developments in synthesis, properties and potential applications. Prog. Mater. Sci. 2016, 83, 270–329. [Google Scholar] [CrossRef]
- Rasouli, M. Biosynthesis of selenium nanoparticles using yeast Nematospora coryli and examination of their anti-candida and anti-oxidant activities. IET Nanobiotechnol. 2019, 13, 214–218. [Google Scholar] [CrossRef]
- Piacenza, E.; Presentato, A.; Zonaro, E.; Lampis, S.; Vallini, G.; Turner, R.J. Selenium and tellurium nanomaterials. Phys. Sci. Rev. 2018, 3. [Google Scholar] [CrossRef]
- Elahian, F.; Reiisi, S.; Shahidi, A.; Mirzaei, S.A. High-throughput bioaccumulation, biotransformation, and production of silver and selenium nanoparticles using genetically engineered Pichia pastoris. Nanomed. Nanobiotechnol. 2017, 13, 853–861. [Google Scholar] [CrossRef]
- Liang, X.; Perez, M.A.M.; Nwoko, K.C.; Egbers, P.; Feldmann, J.; Csetenyi, L.; Gadd, G.M. Fungal formation of selenium and tellurium nanoparticles. Appl. Microbiol. Biotechnol. 2019, 103, 7241–7259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramanujam, J.; Singh, U.P. Copper indium gallium selenide based solar cells—A review. Energy Environ. Sci. 2017, 10, 1306–1319. [Google Scholar] [CrossRef]
- Tan, L.C.; Nancharaiah, Y.V.; Lu, S.; van Hullebusch, E.D.; Gerlach, R.; Lens, P.N.L. Biological treatment of selenium-laden wastewater containing nitrate and sulfate in an upflow anaerobic sludge bed reactor at pH 5.0. Chemosphere 2018, 211, 684–693. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, K.; Foltz, C.M. Selenium as an integral part of factor 3 against dietary necrotic liver degeneration. J. Am. Chem. Soc. 1957, 79, 3292–3293. [Google Scholar] [CrossRef]
- Shoeibi, S.; Mashreghi, M. Biosynthesis of selenium nanoparticles using Enterococcus faecalis and evaluation of their antibacterial activities. J. Trace Elem. Med. Biol. 2017, 39, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Dhanjal, S.; Singh Cameotra, S. Aerobic biogenesis of selenium nanospheres by Bacillus cereus isolated from coalmine soil. Microbial. Cell Fact. 2010, 9, 52. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, E.T.; Safwat, G.M. Assessment of the ameliorative role of selenium nanoparticles on the oxidative stress of acetaminophen in some tissues of male albino rats. Beni-Suef Univ. J. Basic Appl. Sci. 2013, 2, 80–85. [Google Scholar] [CrossRef] [Green Version]
- Wadhwani, S.A.; Gorain, M.; Banerjee, P.; Shedbalkar, U.U.; Singh, R.; Kundu, G.C.; Chopade, B.A. Green synthesis of selenium nanoparticles using Acinetobacter sp. SW30: Optimization, characterization and its anticancer activity in breast cancer cells. Int. J. Nanomed. 2017, 12, 6841–6855. [Google Scholar] [CrossRef] [Green Version]
- Vaigankar, D.C.; Dubey, S.K.; Mujawar, S.Y.; D’Costa, A.; Shyama, S.K. Tellurite biotransformation and detoxification by Shewanella baltica with simultaneous synthesis of tellurium nanorods exhibiting photo-catalytic and anti-biofilm activity. Ecotoxicol. Environ. Saf. 2018, 165, 516–526. [Google Scholar] [CrossRef]
- Huang, W.; Wu, H.; Li, X.; Chen, T. Facile One-Pot Synthesis of Tellurium Nanorods as Antioxidant and Anticancer Agents. Chem. Asian J. 2016, 11, 2301–2311. [Google Scholar] [CrossRef]
- Medina Cruz, D.; Mi, G.; Webster, T.J. Synthesis and characterization of biogenic selenium nanoparticles with antimicrobial properties made by Staphylococcus aureus, methicillin-resistant Staphylococcus aureus (MRSA), Escherichia coli, and Pseudomonas aeruginosa. J. Biomed. Mater. Res. A 2018, 106, 1400–1412. [Google Scholar] [CrossRef] [PubMed]
- Medina Cruz, D.; Tien-Street, W.; Zhang, B.; Huang, X.; Vernet Crua, A.; Nieto-Arguello, A.; Cholula-Diaz, J.L.; Martinez, L.; Huttel, Y.; Ujue Gonzalez, M.; et al. Citric Juice-mediated Synthesis of Tellurium Nanoparticles with Antimicrobial and Anticancer Properties. Green Chem. 2019, 21, 1982–1988. [Google Scholar] [CrossRef] [PubMed]
- Vahidi, H.; Kobarfard, F.; Alizadeh, A.; Saravanan, M.; Barabadi, H. Green nanotechnology-based tellurium nanoparticles: Exploration of their antioxidant, antibacterial, antifungal and cytotoxic potentials against cancerous and normal cells compared to potassium tellurite. Inorg. Chem. Commun. 2021, 124, 108385. [Google Scholar] [CrossRef]
- Mirjani, R.; Faramarzi, M.A.; Sharifzadeh, M.; Setayesh, N.; Khoshayand, M.R.; Shahverdi, A.R. Biosynthesis of tellurium nanoparticles by Lactobacillus plantarum and the effect of nanoparticle-enriched probiotics on the lipid profiles of mice. IET Nanobiotechnol. 2015, 9, 300–305. [Google Scholar] [CrossRef]
- Espinosa-Ortiz, E.J.; Rene, E.R.; Guyot, F.; van Hullebusch, E.D.; Lens, P.N.L. Biomineralization of tellurium and selenium-tellurium nanoparticles by the white-rot fungus Phanerochaete chrysosporium. Int. Biodeter. Biodegr. 2017, 124, 258–266. [Google Scholar] [CrossRef]
- Espinosa-Ortiz, E.J.; Rene, E.R.; van Hullebusch, E.D.; Lens, P.N.L. Removal of selenite from wastewater in a Phanerochaete chrysosporium pellet based fungal bioreactor. Int. Biodeter. Biodegr. 2015, 102, 361–369. [Google Scholar] [CrossRef]
- Espinosa-Ortiz, E.J.; Pechaud, Y.; Lauchnor, E.; Rene, E.R.; Gerlach, R.; Peyton, B.M.; van Hullebusch, E.D.; Lens, P.N. Effect of selenite on the morphology and respiratory activity of Phanerochaete chrysosporium biofilms. Bioresour. Technol. 2016, 210, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Espinosa-Ortiz, E.J.; Gonzalez-Gil, G.; Saikaly, P.E.; van Hullebusch, E.D.; Lens, P.N. Effects of selenium oxyanions on the white-rot fungus Phanerochaete chrysosporium. Appl. Microbiol. Biotechnol. 2015, 99, 2405–2418. [Google Scholar] [CrossRef] [Green Version]
- Gharieb, M.M.; Gadd, G.M. The kinetics of 75[Se]-selenite uptake by Saccharomyces cerevisiae and the vacuolization response to high concentrations. Mycol. Res. 2004, 108, 1415–1422. [Google Scholar] [CrossRef]
- Verma, A.; Gautam, S.P.; Bansal, K.K.; Prabhakar, N.; Rosenholm, J.M. Green Nanotechnology: Advancement in Phytoformulation Research. Medicines 2019, 6. [Google Scholar] [CrossRef] [Green Version]
- Zonaro, E.; Lampis, S.; Turner, R.J.; Qazi, S.J.; Vallini, G. Biogenic selenium and tellurium nanoparticles synthesized by environmental microbial isolates efficaciously inhibit bacterial planktonic cultures and biofilms. Front. Microbiol. 2015, 6, 584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearce, C.I.; Pattrick, R.A.; Law, N.; Charnock, J.M.; Coker, V.S.; Fellowes, J.W.; Oremland, R.S.; Lloyd, J.R. Investigating different mechanisms for biogenic selenite transformations: Geobacter sulfurreducens, Shewanella oneidensis and Veillonella atypica. Environ. Technol. 2009, 30, 1313–1326. [Google Scholar] [CrossRef]
- Pasula, R.R.; Lim, S. Engineering nanoparticle synthesis using microbial factories. Eng. Biol. 2017, 1, 12–17. [Google Scholar] [CrossRef]
- Ruiz Fresneda, M.A.; Delgado Martín, J.; Gómez Bolívar, J.; Fernández Cantos, M.V.; Bosch-Estévez, G.; Martínez Moreno, M.F.; Merroun, M.L. Green synthesis and biotransformation of amorphous Se nanospheres to trigonal 1D Se nanostructures: Impact on Se mobility within the concept of radioactive waste disposal. Environ. Sci. Nano 2018, 5, 2103–2116. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; Wu, S.; Li, N.; Wang, D.; Zheng, S.; Wang, G. Novel bacterial selenite reductase CsrF responsible for Se(IV) and Cr(VI) reduction that produces nanoparticles in Alishewanella sp. WH16-1. J. Hazard. Mater. 2018, 342, 499–509. [Google Scholar] [CrossRef]
- Rajasree, R.S.R.; Gayathri, S. Extracellular biosynthesis of Selenium nanoparticles using some species of Lactobacillus. Indian J. Geo-Marine Sci. 2015, 43, 766–775. [Google Scholar]
- Dahoumane, S.A.; Djediat, C.; Yéprémian, C.; Couté, A.; Fiévet, F.; Coradin, T.; Brayner, R. Species selection for the design of gold nanobioreactor by photosynthetic organisms. J. Nanopart. Res. 2012, 14. [Google Scholar] [CrossRef]
- Dahoumane, S.A.; Mechouet, M.; Alvarez, F.J.; Agathos, S.N.; Jeffryes, C. Microalgae: An outstanding tool in nanotechnology. Bionatura 2016, 1. [Google Scholar] [CrossRef] [Green Version]
- Dahoumane, S.A.; Wijesekera, K.; Filipe, C.D.; Brennan, J.D. Stoichiometrically controlled production of bimetallic Gold-Silver alloy colloids using micro-alga cultures. J. Colloid Interface Sci. 2014, 416, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Alqadi, M.K.; Abo Noqtah, O.A.; Alzoubi, F.Y.; Alzouby, J.; Aljarrah, K. pH effect on the aggregation of silver nanoparticles synthesized by chemical reduction. Mater. Sci. Pol. 2014, 32, 107–111. [Google Scholar] [CrossRef]
- Castro, L.; Blázquez, M.L.; González, F.; Muñoz, J.A.; Ballester, A. Extracellular biosynthesis of gold nanoparticles using sugar beet pulp. Chem. Eng. J. 2010, 164, 92–97. [Google Scholar] [CrossRef]
- Wu, S.; Sun, K.; Wang, X.; Wang, D.; Wan, X.; Zhang, J. Protonation of epigallocatechin-3-gallate (EGCG) results in massive aggregation and reduced oral bioavailability of EGCG-dispersed selenium nanoparticles. J. Agric. Food. Chem. 2013, 61, 7268–7275. [Google Scholar] [CrossRef] [PubMed]
- Akcay, F.A.; Avci, A. Effects of process conditions and yeast extract on the synthesis of selenium nanoparticles by a novel indigenous isolate Bacillus sp. EKT1 and characterization of nanoparticles. Arch. Microbiol. 2020, 202, 2233–2243. [Google Scholar] [CrossRef]
- Kuroda, M.; Notaguchi, E.; Sato, A.; Yoshioka, M.; Hasegawa, A.; Kagami, T.; Narita, T.; Yamashita, M.; Sei, K.; Soda, S.; et al. Characterization of Pseudomonas stutzeri NT-I capable of removing soluble selenium from the aqueous phase under aerobic conditions. J. Biosci. Bioeng. 2011, 112, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.P.; Xu, S.; Liu, L.P.; Li, Y.D. A positive-microemulsion method for preparing nearly uniform Ag2Se nanoparticles at low temperature. Chem. Eur. J. 2006, 12, 3672–3677. [Google Scholar] [CrossRef]
- Mollania, N.; Tayebee, R.; Narenji-Sani, F. An environmentally benign method for the biosynthesis of stable selenium nanoparticles. Res. Chem. Intermed. 2015, 42, 4253–4271. [Google Scholar] [CrossRef]
- Ashour, A.H.; El-Batal, A.I.; Maksoud, M.I.A.A.; El-Sayyad, G.S.; Labib, S.; Abdeltwab, E.; El-Okr, M.M. Antimicrobial activity of metal-substituted cobalt ferrite nanoparticles synthesized by sol–gel technique. Particuology 2018, 40, 141–151. [Google Scholar] [CrossRef]
- Wong, C.W.; Chan, Y.S.; Jeevanandam, J.; Pal, K.; Bechelany, M.; Abd Elkodous, M.; El-Sayyad, G.S. Response Surface Methodology Optimization of Mono-dispersed MgO Nanoparticles Fabricated by Ultrasonic-Assisted Sol–Gel Method for Outstanding Antimicrobial and Antibiofilm Activities. J. Clust. Sci. 2020, 31, 367–389. [Google Scholar] [CrossRef]
- Baer, D.R. Surface Characterization of Nanoparticles: Critical needs and significant challenges. J. Surf. Anal. 2011, 17, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Sadtler, B.; Burgos, S.P.; Batara, N.A.; Beardslee, J.A.; Atwater, H.A.; Lewis, N.S. Phototropic growth control of nanoscale pattern formation in photoelectrodeposited Se-Te films. Proc. Natl. Acad. Sci. USA 2013, 110, 19707–19712. [Google Scholar] [CrossRef] [Green Version]
- Vijayanandan, A.S.; Balakrishnan, R.M. Impact of precursor concentration on biological synthesis of cobalt oxide nanoparticles. Data Brief 2018, 19, 1941–1947. [Google Scholar] [CrossRef] [PubMed]
- Sharifi Dehsari, H.; Halda Ribeiro, A.; Ersöz, B.; Tremel, W.; Jakob, G.; Asadi, K. Effect of precursor concentration on size evolution of iron oxide nanoparticles. CrystEngComm 2017, 19, 6694–6702. [Google Scholar] [CrossRef] [Green Version]
- Moloto, N.; Revaprasadu, N.; Musetha, P.L.; Moloto, M.J. The effect of precursor concentration, temperature and capping group on the morphology of CdS nanoparticles. J. Nanosci. Nanotechnol. 2009, 9, 4760–4766. [Google Scholar] [CrossRef] [PubMed]
- Dahoumane, S.A.; Djediat, C.; Yepremian, C.; Coute, A.; Fievet, F.; Coradin, T.; Brayner, R. Recycling and adaptation of Klebsormidium flaccidum microalgae for the sustained production of gold nanoparticles. Biotechnol. Bioeng. 2012, 109, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Kumar, S.; Bafana, A.; Dahoumane, S.A.; Jeffryes, C. Individual and Combined Effects of Extracellular Polymeric Substances and Whole Cell Components of Chlamydomonas reinhardtii on Silver Nanoparticle Synthesis and Stability. Molecules 2019, 24. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, S.; Mehta, S.K. Selenium nanomaterials: Applications in electronics, catalysis and sensors. J. Nanosci. Nanotechnol. 2014, 14, 1658–1674. [Google Scholar] [CrossRef]
- Wang, X.; Liu, G.; Zhou, J.; Wang, J.; Jin, R.; Lv, H. Quinone-mediated reduction of selenite and tellurite by Escherichia coli. Bioresour. Technol. 2011, 102, 3268–3271. [Google Scholar] [CrossRef]
- Presentato, A.; Piacenza, E.; Darbandi, A.; Anikovskiy, M.; Cappelletti, M.; Zannoni, D.; Turner, R.J. Assembly, growth and conductive properties of tellurium nanorods produced by Rhodococcus aetherivorans BCP1. Sci. Rep. 2018, 8, 3923. [Google Scholar] [CrossRef] [Green Version]
- Presentato, A.; Piacenza, E.; Anikovskiy, M.; Cappelletti, M.; Zannoni, D.; Turner, R.J. Biosynthesis of selenium-nanoparticles and -nanorods as a product of selenite bioconversion by the aerobic bacterium Rhodococcus aetherivorans BCP1. New Biotechnol. 2018, 41, 1–8. [Google Scholar] [CrossRef]
- Prange, A.; Birzele, B.; Hormes, J.; Modrow, H. Investigation of different human pathogenic and food contaminating bacteria and moulds grown on selenite/selenate and tellurite/tellurate by X-ray absorption spectroscopy. Food Control. 2005, 16, 723–728. [Google Scholar] [CrossRef]
- Wang, T.; Yang, L.; Zhang, B.; Liu, J. Extracellular biosynthesis and transformation of selenium nanoparticles and application in H2O2 biosensor. Colloid Surface B Biointerface. 2010, 80, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Menon, S.; Shanmugam, V. Cytotoxicity Analysis of Biosynthesized Selenium Nanoparticles Towards A549 Lung Cancer Cell Line. J. Inorg. Organometal. Polym. Mater. 2019, 30, 1852–1864. [Google Scholar] [CrossRef]
- Rajkumar, K.; Mvs, S.; Koganti, S.; Burgula, S. Selenium Nanoparticles Synthesized Using Pseudomonas stutzeri (MH191156) Show Antiproliferative and Anti-angiogenic Activity Against Cervical Cancer Cells. Int. J. Nanomed. 2020, 15, 4523–4540. [Google Scholar] [CrossRef] [PubMed]
- Sayes, C.M.; Warheit, D.B. Characterization of nanomaterials for toxicity assessment. Adv. Rev. 2009, 1, 960–970. [Google Scholar] [CrossRef] [PubMed]
- Warheit, D.B. Nanoparticles. Mater. Today 2004, 7, 32–35. [Google Scholar] [CrossRef]
- Warheit, D.B.; Sayes, C.M.; Reed, K.L.; Swain, K.A. Health effects related to nanoparticle exposures: Environmental, health and safety considerations for assessing hazards and risks. Pharmacol. Ther. 2008, 120, 35–42. [Google Scholar] [CrossRef]
- Powers, K.W.; Palazuelos, M.; Moudgil, B.M.; Roberts, S.M. Characterization of the size, shape, and state of dispersion of nanoparticles for toxicological studies. Nanotoxicology 2009, 1, 42–51. [Google Scholar] [CrossRef]
- Kapur, M.; Soni, K.; Kohli, K. Green Synthesis of Selenium Nanoparticles from Broccoli, Characterization, Application and Toxicity. Adv. Tech. Biol. Med. 2017, 5. [Google Scholar] [CrossRef]
- Estevam, E.C.; Griffin, S.; Nasim, M.J.; Denezhkin, P.; Schneider, R.; Lilischkis, R.; Dominguez-Alvarez, E.; Witek, K.; Latacz, G.; Keck, C.; et al. Natural selenium particles from Staphylococcus carnosus: Hazards or particles with particular promise? J. Hazard. Mater. 2017, 324, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, M.; Schmidt, S.; Winter, J. Formation of Se (0) Nanoparticles by Duganella sp. and Agrobacterium sp. isolated from Se-laden soil of North-East Punjab, India. Microb. Cell Fact. 2012, 11, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Combs, G.F.J.; Garbisu, C.; Yee, B.C.; Yee, A.; Carlson, D.E.; Smith, N.R.; Magyarosy, A.C.; Leighton, T.; Buchanan, B.B. Bioavailability of Selenium Accumulated by Selenite-reducing Bacteria. Biol. Trace Elem. Res. 1996, 52, 209–225. [Google Scholar] [CrossRef]
- Kenward, P.A.; Fowle, D.A.; Yee, N. Microbial Selenate Sorption and Reduction in Nutrient Limited Systems. Environ. Sci. Technol. 2006, 40, 3782–3786. [Google Scholar] [CrossRef] [PubMed]
- Lampis, S.; Zonaro, E.; Bertolini, C.; Cecconi, D.; Monti, F.; Micaroni, M.; Turner, R.J.; Butler, C.S.; Vallini, G. Selenite biotransformation and detoxification by Stenotrophomonas maltophilia SeITE02: Novel clues on the route to bacterial biogenesis of selenium nanoparticles. J. Hazard. Mater. 2017, 324, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Cremonini, E.; Zonaro, E.; Donini, M.; Lampis, S.; Boaretti, M.; Dusi, S.; Melotti, P.; Lleo, M.M.; Vallini, G. Biogenic selenium nanoparticles: Characterization, antimicrobial activity and effects on human dendritic cells and fibroblasts. Microb. Biotechnol. 2016, 9, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, M.; Fernandez, V.; Arenas-Salinas, M.; Ahumada, D.; Munoz-Villagran, C.; Cornejo, F.; Vargas, E.; Latorre, M.; Morales, E.; Vasquez, C.; et al. Synthesis and Antibacterial Activity of Metal(loid) Nanostructures by Environmental Multi-Metal(loid) Resistant Bacteria and Metal(loid)-Reducing Flavoproteins. Front. Microbiol. 2018, 9, 959. [Google Scholar] [CrossRef]
- Tam, K.; Ho, C.T.; Lee, J.H.; Lai, M.; Chang, C.H.; Rheem, Y.; Chen, W.; Hur, H.G.; Myung, N.V. Growth mechanism of amorphous selenium nanoparticles synthesized by Shewanella sp. HN-41. Biosci. Biotechnol. Biochem. 2010, 74, 696–700. [Google Scholar] [CrossRef] [Green Version]
- Shakibaie, M.; Khorramizadeh, M.R.; Faramarzi, M.A.; Sabzevari, O.; Shahverdi, A.R. Biosynthesis and recovery of selenium nanoparticles and the effects on matrix metalloproteinase-2 expression. Biotechnol. Appl. Biochem. 2010, 56, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Shakibaie, M.; Forootanfar, H.; Golkari, Y.; Mohammadi-Khorsand, T.; Shakibaie, M.R. Anti-biofilm activity of biogenic selenium nanoparticles and selenium dioxide against clinical isolates of Staphylococcus aureus, Pseudomonas aeruginosa, and Proteus mirabilis. J. Trace Elem. Med. Biol. 2015, 29, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Beheshti, N.; Soflaei, S.; Shakibaie, M.; Yazdi, M.H.; Ghaffarifar, F.; Dalimi, A.; Shahverdi, A.R. Efficacy of biogenic selenium nanoparticles against Leishmania major: In vitro and in vivo studies. J. Trace Elem. Med. Biol. 2013, 27, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Forootanfar, H.; Adeli-Sardou, M.; Nikkhoo, M.; Mehrabani, M.; Amir-Heidari, B.; Shahverdi, A.R.; Shakibaie, M. Antioxidant and cytotoxic effect of biologically synthesized selenium nanoparticles in comparison to selenium dioxide. J. Trace Elem. Med. Biol. 2014, 28, 75–79. [Google Scholar] [CrossRef]
- Fernandez-Llamosas, H.; Castro, L.; Blazquez, M.L.; Diaz, E.; Carmona, M. Biosynthesis of selenium nanoparticles by Azoarcus sp. CIB. Microb. Cell Fact. 2016, 15, 109. [Google Scholar] [CrossRef] [PubMed]
- Khoei, N.S.; Lampis, S.; Zonaro, E.; Yrjala, K.; Bernardi, P.; Vallini, G. Insights into selenite reduction and biogenesis of elemental selenium nanoparticles by two environmental isolates of Burkholderia fungorum. New Biotechnol. 2017, 34, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Wang, Y.; Wang, Y.; Xu, D.; Huang, Y.; Wang, D.; Wang, G.; Rensing, C.; Zheng, S. Novel mechanisms of selenate and selenite reduction in the obligate aerobic bacterium Comamonas testosteroni S44. J. Hazard. Mater. 2018, 359, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Kamnev, A.A.; Mamchenkova, P.V.; Dyatlova, Y.A.; Tugarova, A.V. FTIR spectroscopic studies of selenite reduction by cells of the rhizobacterium Azospirillum brasilense Sp7 and the formation of selenium nanoparticles. J. Mol. Struct. 2017, 1140, 106–112. [Google Scholar] [CrossRef]
- Vogel, M.; Fischer, S.; Maffert, A.; Hubner, R.; Scheinost, A.C.; Franzen, C.; Steudtner, R. Biotransformation and detoxification of selenite by microbial biogenesis of selenium-sulfur nanoparticles. J. Hazard. Mater. 2018, 344, 749–757. [Google Scholar] [CrossRef]
- Kora, A.J.; Rastogi, L. Biomimetic synthesis of selenium nanoparticles by Pseudomonas aeruginosa ATCC 27853: An approach for conversion of selenite. J. Environ. Manag. 2016, 181, 231–236. [Google Scholar] [CrossRef]
- Pouri, S.; Motamedi, H.; Honary, S.; Kazeminezhad, I. Biological Synthesis of Selenium Nanoparticles and Evaluation of their Bioavailability. Braz. Arch. Biol. Techn. 2018, 60. [Google Scholar] [CrossRef]
- Srivastava, N.; Mukhopadhyay, M. Biosynthesis and structural characterization of selenium nanoparticles mediated by Zooglea ramigera. Powder Technol. 2013, 244, 26–29. [Google Scholar] [CrossRef]
- Subedi, G.; Taylor, J.; Hatam, I.; Baldwin, S.A. Simultaneous selenate reduction and denitrification by a consortium of enriched mine site bacteria. Chemosphere 2017, 183, 536–545. [Google Scholar] [CrossRef]
- Wadhwani, S.A.; Shedbalkar, U.U.; Singh, R.; Chopade, B.A. Biosynthesis of gold and selenium nanoparticles by purified protein from Acinetobacter sp. SW 30. Enzyme Microb. Technol. 2018, 111, 81–86. [Google Scholar] [CrossRef]
- Ramya, S.; Shanmugasundaram, T.; Balagurunathan, R. Biomedical potential of actinobacterially synthesized selenium nanoparticles with special reference to anti-biofilm, anti-oxidant, wound healing, cytotoxic and anti-viral activities. J. Trace Elem. Med. Biol. 2015, 32, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Ameri, A.; Shakibaie, M.; Ameri, A.; Faramarzi, M.A.; Amir-Heidari, B.; Forootanfar, H. Photocatalytic decolorization of bromothymol blue using biogenic selenium nanoparticles synthesized by terrestrial actinomycete Streptomyces griseobrunneus strain FSHH12. Desalin. Water Treat. 2015, 57, 21552–21563. [Google Scholar] [CrossRef]
- Fernandez-Llamosas, H.; Castro, L.; Blazquez, M.L.; Diaz, E.; Carmona, M. Speeding up bioproduction of selenium nanoparticles by using Vibrio natriegens as microbial factory. Sci. Rep. 2017, 7, 16046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Guo, Y.; Qiao, L.; Ma, L.; Cheng, Y.; Roman, A. Biogenic Synthesis of Novel Functionalized Selenium Nanoparticles by Lactobacillus casei ATCC 393 and Its Protective Effects on Intestinal Barrier Dysfunction Caused by Enterotoxigenic Escherichia coli K88. Front. Microbiol. 2018, 9, 1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaaban, M.; El-Mahdy, A.M. Biosynthesis of Ag, Se, and ZnO nanoparticles with antimicrobial activities against resistant pathogens using waste isolate Streptomyces enissocaesilis. IET Nanobiotechnol. 2018, 12, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Ramya, S.; Shanmugasundaram, T.; Balagurunathan, R. Actinobacterial enzyme mediated synthesis of selenium nanoparticles for antibacterial, mosquito larvicidal and anthelminthic applications. Particul. Sci. Technol. 2019, 38, 63–72. [Google Scholar] [CrossRef]
- San Keskin, N.O.; Akbal Vural, O.; Abaci, S. Biosynthesis of Noble Selenium Nanoparticles from Lysinibacillus sp. NOSK for Antimicrobial, Antibiofilm Activity, and Biocompatibility. Geomicrobiol. J. 2020, 37, 919–928. [Google Scholar] [CrossRef]
- Srivastava, P.; Kowshik, M. Anti-neoplastic selenium nanoparticles from Idiomarina sp. PR58-8. Enzyme Microb. Technol. 2016, 95, 192–200. [Google Scholar] [CrossRef]
- Srivastava, N.; Mukhopadhyay, M. Green synthesis and structural characterization of selenium nanoparticles and assessment of their antimicrobial property. Bioprocess. Biosyst. Eng. 2015, 38, 1723–1730. [Google Scholar] [CrossRef]
- Song, D.; Li, X.; Cheng, Y.; Xiao, X.; Lu, Z.; Wang, Y.; Wang, F. Aerobic biogenesis of selenium nanoparticles by Enterobacter cloacae Z0206 as a consequence of fumarate reductase mediated selenite reduction. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Li, D.B.; Cheng, Y.Y.; Wu, C.; Li, W.W.; Li, N.; Yang, Z.C.; Tong, Z.H.; Yu, H.Q. Selenite reduction by Shewanella oneidensis MR-1 is mediated by fumarate reductase in periplasm. Sci. Rep. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Hnain, A.; Brooks, J.; Lefebvre, D.D. The synthesis of elemental selenium particles by Synechococcus leopoliensis. Appl. Microbiol. Biotechnol. 2013, 97, 10511–10519. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Su, J.; Wang, L.; Yao, R.; Wang, D.; Deng, Y.; Wang, R.; Wang, G.; Rensing, C. Selenite reduction by the obligate aerobic bacterium Comamonas testosteroni S44 isolated from a metal-contaminated soil. BMC Microbiol. 2014, 14, 204. [Google Scholar] [CrossRef] [PubMed]
- Tugarova, A.V.; Mamchenkova, P.V.; Khanadeev, V.A.; Kamnev, A.A. Selenite reduction by the rhizobacterium Azospirillum brasilense, synthesis of extracellular selenium nanoparticles and their characterisation. N. Biotechnol. 2020, 58, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Kora, A.J. Bacillus cereus, selenite-reducing bacterium from contaminated lake of an industrial area: A renewable nanofactory for the synthesis of selenium nanoparticles. Bioresour. Bioprocess. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Eszenyi, P.; Sztrik, A.; Babka, B.; Prokisch, J. Elemental, Nano-Sized (100-500 nm) Selenium Production by Probiotic Lactic Acid Bacteria. Int. J. Biosci. Biochem. Bioinform. 2011, 1, 148–152. [Google Scholar] [CrossRef] [Green Version]
- Shakibaie, M.; Shahverdi, A.R.; Faramarzi, M.A.; Hassanzadeh, G.R.; Rahimi, H.R.; Sabzevari, O. Acute and subacute toxicity of novel biogenic selenium nanoparticles in mice. Pharm. Biol. 2013, 51, 58–63. [Google Scholar] [CrossRef]
- Xu, C.; Qiao, L.; Guo, Y.; Ma, L.; Cheng, Y. Preparation, characteristics and antioxidant activity of polysaccharides and proteins-capped selenium nanoparticles synthesized by Lactobacillus casei ATCC 393. Carbohydr. Polym. 2018, 195, 576–585. [Google Scholar] [CrossRef]
- Wadgaonkar, S.L.; Mal, J.; Nancharaiah, Y.V.; Maheshwari, N.O.; Esposito, G.; Lens, P.N.L. Formation of Se(0), Te(0), and Se(0)-Te(0) nanostructures during simultaneous bioreduction of selenite and tellurite in a UASB reactor. Appl. Microbiol. Biotechnol. 2018, 102, 2899–2911. [Google Scholar] [CrossRef]
- Staicu, L.C.; van Hullebusch, E.D.; Oturan, M.A.; Ackerson, C.J.; Lens, P.N. Removal of colloidal biogenic selenium from wastewater. Chemosphere 2015, 125, 130–138. [Google Scholar] [CrossRef]
- Dessi, P.; Jain, R.; Singh, S.; Seder-Colomina, M.; van Hullebusch, E.D.; Rene, E.R.; Ahammad, S.Z.; Carucci, A.; Lens, P.N.L. Effect of temperature on selenium removal from wastewater by UASB reactors. Water Res. 2016, 94, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Golshan, N.H.; Deng, X.; Hickey, D.J.; Zeimer, K.; Li, H.; Webster, T.J. Selenium nanoparticles incorporated into titania nanotubes inhibit bacterial growth and macrophage proliferation. Nanoscale 2016, 8, 15783–15794. [Google Scholar] [CrossRef] [PubMed]
- Eswayah, A.S.; Smith, T.J.; Gardiner, P.H. Microbial Transformations of Selenium Species of Relevance to Bioremediation. Appl. Environ. Microbiol. 2016, 82, 4848–4859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoeibi, S.; Mozdziak, P.; Golkar-Narenji, A. Biogenesis of Selenium Nanoparticles Using Green Chemistry. Top. Curr. Chem. 2017, 375, 88. [Google Scholar] [CrossRef] [PubMed]
- Nancharaiah, Y.V.; Lens, P.N. Ecology and biotechnology of selenium-respiring bacteria. Microbiol. Mol. Biol. Rev. 2015, 79, 61–80. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.; Yao, R.; Wang, R.; Wang, D.; Wang, G.; Zheng, S. Reduction of selenite to Se(0) nanoparticles by filamentous bacterium Streptomyces sp. ES2-5 isolated from a selenium mining soil. Microb. Cell Fact. 2016, 15, 157. [Google Scholar] [CrossRef] [Green Version]
- Tomei, F.A.; Barton, L.L.; Lemanski, C.L.; Zocco, T.G.; Fink, N.H.; Sillerud, L.O. Transformation of selenate and selenite to elemental selenium by Desulfovibrio desulfuricans. J. Indus. Microbiol. 1995, 14, 329–336. [Google Scholar] [CrossRef]
- Losi, M.E.; Frankenberger, W.T., Jr. Reduction of Selenium Oxyanions by Enterobacter cloacae SLD1a-1: Isolation and Growth of the Bacterium and Its Expulsion of Selenium Particles. Appl. Environ. Microbiol. 1997, 63, 3079–3084. [Google Scholar] [CrossRef] [Green Version]
- Kessi, J.; Ramuz, M.; Wehrli, E.; Spycher, M.; Bachofen, R. Reduction of Selenite and Detoxification of Elemental Selenium by the Phototrophic Bacterium Rhodospirillum rubrum. Appl. Environ. Microbiol. 1999, 65, 4734–4740. [Google Scholar] [CrossRef] [Green Version]
- Tan, L.C.; Nancharaiah, Y.V.; van Hullebusch, E.D.; Lens, P.N.L. Selenium: Environmental significance, pollution, and biological treatment technologies. Biotechnol. Adv. 2016, 34, 886–907. [Google Scholar] [CrossRef]
- Kessi, J.; Hanselmann, K.W. Similarities between the abiotic reduction of selenite with glutathione and the dissimilatory reaction mediated by Rhodospirillum rubrum and Escherichia coli. J. Biol. Chem. 2004, 279, 50662–50669. [Google Scholar] [CrossRef] [PubMed]
- Hunter, W.J. Pseudomonas seleniipraecipitans proteins potentially involved in selenite reduction. Curr. Microbiol. 2014, 69, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Zhou, R.; Webster, T.J. Green Synthesized BSA-Coated Selenium Nanoparticles Inhibit Bacterial Growth While Promoting Mammalian Cell Growth. Int. J. Nanomed. 2020, 15, 115–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.; Wu, X.; Chen, P.; Zhang, L.; Yang, C.S.; Zhang, J. Selenium nanoparticles are more efficient than sodium selenite in producing reactive oxygen species and hyper-accumulation of selenium nanoparticles in cancer cells generates potent therapeutic effects. Free Radic. Biol. Med. 2018, 126, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Krafft, T.; Bowen, A.; Theis, F.; Macy, J.M. Cloning and Sequencing of the Genes Encoding the Periplasmic-Cytochrome B-Containing Selenate Reductase of Thauera selenafis. DNA Seq. 2000, 10, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Hunter, W.J. A Rhizobium selenitireducens protein showing selenite reductase activity. Curr. Microbiol. 2014, 68, 311–316. [Google Scholar] [CrossRef]
- Kuroda, M.; Yamashita, M.; Miwa, E.; Imao, K.; Fujimoto, N.; Ono, H.; Nagano, K.; Sei, K.; Ike, M. Molecular cloning and characterization of the srdBCA operon, encoding the respiratory selenate reductase complex, from the selenate-reducing bacterium Bacillus selenatarsenatis SF-1. J. Bacteriol. 2011, 193, 2141–2148. [Google Scholar] [CrossRef] [Green Version]
- Afkar, E.; Lisak, J.; Saltikov, C.; Basu, P.; Oremland, R.S.; Stolz, J.F. The respiratory arsenate reductase from Bacillus selenitireducens strain MLS10. FEMS Microbiol. Lett. 2003, 226, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Harrison, G.; Curie, C.; Laishley, E.J. Purification and characterization of an inducible dissimilatory type sulfite reductase from Clostridium pasteurianum. Arch. Microbiol. 1984, 138, 72–78. [Google Scholar] [CrossRef]
- Yanke, L.J.; Bryant, R.D.; Laishley, E.J. Hydrogenase I of Clostridium pasteurianum functions as a novel selenite reductase. Anaerobe 1995, 1, 61–67. [Google Scholar] [CrossRef]
- DeMoll-Decker, H.; Macy, J.M. The periplasmic nitrite reductase of Thauera selenatis may catalyze the reduction of selenite to elemental selenium. Arch. Microbiol. 1993, 160, 241–247. [Google Scholar]
- Ridley, H.; Watts, C.A.; Richardson, D.J.; Butler, C.S. Resolution of distinct membrane-bound enzymes from Enterobacter cloacae SLD1a-1 that are responsible for selective reduction of nitrate and selenate oxyanions. Appl. Environ. Microbiol. 2006, 72, 5173–5180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessi, J. Enzymic systems proposed to be involved in the dissimilatory reduction of selenite in the purple non-sulfur bacteria Rhodospirillum rubrum and Rhodobacter capsulatus. Microbiology 2006, 152, 731–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonkusre, P.; Nanduri, R.; Gupta, P.; Singh Cameotra, S. Improved Extraction of Intracellular Biogenic Selenium Nanoparticles and their Specificity for Cancer Chemoprevention. Nanomed. Nanotechnol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Lenz, M.; Kolvenbach, B.; Gygax, B.; Moes, S.; Corvini, P.F. Shedding light on selenium biomineralization: Proteins associated with bionanominerals. Appl. Environ. Microbiol. 2011, 77, 4676–4680. [Google Scholar] [CrossRef] [Green Version]
- Lynch, I.; Dawson, K.A. Protein-nanoparticle interactions. Nano Today 2008, 3, 40–47. [Google Scholar] [CrossRef]
- Xue, L.; Greisler, H.P. Biomaterials in the development and future of vascular grafts. J. Vasc. Surg. 2003, 37, 472–480. [Google Scholar] [CrossRef] [Green Version]
- Dhillon, G.S.; Brar, S.K.; Kaur, S.; Verma, M. Green approach for nanoparticle biosynthesis by fungi: Current trends and applications. Crit. Rev. Biotechnol. 2012, 32, 49–73. [Google Scholar] [CrossRef]
- Boroumand Moghaddam, A.; Namvar, F.; Moniri, M.; Md Tahir, P.; Azizi, S.; Mohamad, R. Nanoparticles Biosynthesized by Fungi and Yeast: A Review of Their Preparation, Properties, and Medical Applications. Molecules 2015, 20, 16540–16565. [Google Scholar] [CrossRef]
- Sarkar, J.; Dey, P.; Saha, S.; Acharya, K. Mycosynthesis of selenium nanoparticles. Micro Nano Lett. 2011, 6, 599. [Google Scholar] [CrossRef]
- Joshi, S.M.; De Britto, S.; Jogaiah, S.; Ito, S.I. Mycogenic Selenium Nanoparticles as Potential New Generation Broad Spectrum Antifungal Molecules. Biomolecules 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diko, C.S.; Zhang, H.; Lian, S.; Fan, S.; Li, Z.; Qu, Y. Optimal synthesis conditions and characterization of selenium nanoparticles in Trichoderma sp. WL-Go culture broth. Mater. Chem. Phys. 2020, 246, 122583. [Google Scholar] [CrossRef]
- Liang, T.; Qiu, X.; Ye, X.; Liu, Y.; Li, Z.; Tian, B.; Yan, D. Biosynthesis of selenium nanoparticles and their effect on changes in urinary nanocrystallites in calcium oxalate stone formation. 3 Biotech. 2020, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Mosallam, F.M.; El-Sayyad, G.S.; Fathy, R.M.; El-Batal, A.I. Biomolecules-mediated synthesis of selenium nanoparticles using Aspergillus oryzae fermented Lupin extract and gamma radiation for hindering the growth of some multidrug-resistant bacteria and pathogenic fungi. Microb. Pathog. 2018, 122, 108–116. [Google Scholar] [CrossRef] [PubMed]
- El-Batal, A.I.; Mosallam, F.M.; Ghorab, M.M.; Hanora, A.; Gobara, M.; Baraka, A.; Elsayed, M.A.; Pal, K.; Fathy, R.M.; Abd Elkodous, M.; et al. Factorial design-optimized and gamma irradiation-assisted fabrication of selenium nanoparticles by chitosan and Pleurotus ostreatus fermented fenugreek for a vigorous in vitro effect against carcinoma cells. Int. J. Biol. Macromol. 2020, 156, 1584–1599. [Google Scholar] [CrossRef] [PubMed]
- El-Sayyad, G.S.; El-Bastawisy, H.S.; Gobara, M.; El-Batal, A.I. Gentamicin-Assisted Mycogenic Selenium Nanoparticles Synthesized Under Gamma Irradiation for Robust Reluctance of Resistant Urinary Tract Infection-Causing Pathogens. Biol. Trace Elem. Res. 2020, 195, 323–342. [Google Scholar] [CrossRef]
- Gao, X.; Li, X.; Mu, J.; Ho, C.T.; Su, J.; Zhang, Y.; Lin, X.; Chen, Z.; Li, B.; Xie, Y. Preparation, physicochemical characterization, and anti-proliferation of selenium nanoparticles stabilized by Polyporus umbellatus polysaccharide. Int. J. Biol. Macromol. 2020, 152, 605–615. [Google Scholar] [CrossRef]
- Jin, Y.; Cai, L.; Yang, Q.; Luo, Z.; Liang, L.; Liang, Y.; Wu, B.; Ding, L.; Zhang, D.; Xu, X.; et al. Anti-leukemia activities of selenium nanoparticles embedded in nanotube consisted of triple-helix β-D-glucan. Carbohydr. Polym. 2020, 240, 116329. [Google Scholar] [CrossRef]
- Liao, W.; Yu, Z.; Lin, Z.; Lei, Z.; Ning, Z.; Regenstein, J.M.; Yang, J.; Ren, J. Biofunctionalization of Selenium Nanoparticle with Dictyophora Indusiata Polysaccharide and Its Antiproliferative Activity through Death-Receptor and Mitochondria-Mediated Apoptotic Pathways. Sci. Rep. 2015, 5, 18629. [Google Scholar] [CrossRef]
- Liu, Y.; Zeng, S.; Liu, Y.; Wu, W.; Shen, Y.; Zhang, L.; Li, C.; Chen, H.; Liu, A.; Shen, L.; et al. Synthesis and antidiabetic activity of selenium nanoparticles in the presence of polysaccharides from Catathelasma ventricosum. Int. J. Biol. Macromol. 2018, 114, 632–639. [Google Scholar] [CrossRef]
- Rosenfeld, C.E.; Kenyon, J.A.; James, B.R.; Santelli, C.M. Selenium (IV,VI) reduction and tolerance by fungi in an oxic environment. Geobiology 2017, 15, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Vetchinkina, E.; Loshchinina, E.; Kupryashina, M.; Burov, A.; Pylaev, T.; Nikitina, V. Green synthesis of nanoparticles with extracellular and intracellular extracts of basidiomycetes. PeerJ 2018, 6, e5237. [Google Scholar] [CrossRef] [PubMed]
- Vetchinkina, E.; Loshchinina, E.; Kursky, V.; Nikitina, V. Reduction of organic and inorganic selenium compounds by the edible medicinal basidiomycete Lentinula edodes and the accumulation of elemental selenium nanoparticles in its mycelium. J. Microbiol. 2013, 51, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Vetchinkina, E.; Loshchinina, E.; Kupryashina, M.; Burov, A.; Nikitina, V. Shape and Size Diversity of Gold, Silver, Selenium, and Silica Nanoparticles Prepared by Green Synthesis Using Fungi and Bacteria. Indus. Eng. Chem. Res. 2019, 58, 17207–17218. [Google Scholar] [CrossRef]
- Xiao, Y.; Huang, Q.; Zheng, Z.; Guan, H.; Liu, S. Construction of a Cordyceps sinensis exopolysaccharide-conjugated selenium nanoparticles and enhancement of their antioxidant activities. Int. J. Biol. Macromol. 2017, 99, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhou, H.; Bai, J.; Li, Y.; Yang, J.; Ma, Q.; Qu, Y. Biosynthesis of selenium nanoparticles mediated by fungus Mariannaea sp. HJ and their characterization. Colloid Surf. A Physicochem. Eng. Aspect 2019, 571, 9–16. [Google Scholar] [CrossRef]
- Srivastava, N.; Mukhopadhyay, M. Biosynthesis and Structural Characterization of Selenium Nanoparticles Using Gliocladium roseum. J. Cluster Sci. 2015, 26, 1473–1482. [Google Scholar] [CrossRef]
- Gharieb, M.M.; Wilkinson, S.C.; Gadd, G.M. Reduction of selenium oxyanions by unicellular, polymorphic and filamentous fungi: Cellular location of reduced selenium and implications for tolerance. J. Indud. Microbiol. 1995, 14, 300–311. [Google Scholar] [CrossRef]
- Das, S.K.; Dickinson, C.; Lafir, F.; Brougham, D.F.; Marsili, E. Synthesis, characterization and catalytic activity of gold nanoparticles biosynthesized with Rhizopus oryzae protein extract. Green Chem. 2012, 14, 1322–1334. [Google Scholar] [CrossRef]
- Sarkar, J.; Acharya, K. Alternaria alternata culture filtrate mediated bioreduction of chloroplatinate to platinum nanoparticles. Inorg. Nano Metal. Chem. 2017, 47, 365–369. [Google Scholar] [CrossRef]
- Sathishkumar, Y.; Devarayan, K.; Ki, C.; Rajagopal, K.; Soo Lee, Y. Shape-controlled extracellular synthesis of silver nanocubes by Mucor circinelloides. Mater. Lett. 2015, 159, 481–483. [Google Scholar] [CrossRef]
- Herrero, E.; Wellinger, R.E. Yeast as a model system to study metabolic impact of selenium compounds. Microb. Cell 2015, 2, 139–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrauzer, G.N. Selenium yeast: Composition, quality, analysis, and safety. Pure Appl. Chem. 2006, 78, 105–109. [Google Scholar] [CrossRef] [Green Version]
- Rayman, M.P. The use of high-selenium yeast to raise selenium status: How does it measure up? Br. J. Nutr. 2004, 92, 557–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Lamana, J.; Abad-Álvaro, I.; Bierla, K.; Laborda, F.; Szpunar, J.; Lobinski, R. Detection and characterization of biogenic selenium nanoparticles in selenium-rich yeast by single particle ICPMS. J. Anal. Atom. Spectrom. 2018, 33, 452–460. [Google Scholar] [CrossRef]
- Bartosiak, M.; Giersz, J.; Jankowski, K. Analytical monitoring of selenium nanoparticles green synthesis using photochemical vapor generation coupled with MIP-OES and UV–Vis spectrophotometry. Microchem. J. 2019, 145, 1169–1175. [Google Scholar] [CrossRef]
- Lian, S.; Diko, C.S.; Yan, Y.; Li, Z.; Zhang, H.; Ma, Q.; Qu, Y. Characterization of biogenic selenium nanoparticles derived from cell-free extracts of a novel yeast Magnusiomyces ingens. 3 Biotech. 2019, 9. [Google Scholar] [CrossRef]
- Faramarzi, S.; Anzabi, Y.; Jafarizadeh-Malmiri, H. Nanobiotechnology approach in intracellular selenium nanoparticle synthesis using Saccharomyces cerevisiae-fabrication and characterization. Arch. Microbiol. 2020, 202, 1203–1209. [Google Scholar] [CrossRef]
- Asghari-Paskiabi, F.; Imani, M.; Rafii-Tabar, H.; Razzaghi-Abyaneh, M. Physicochemical properties, antifungal activity and cytotoxicity of selenium sulfide nanoparticles green synthesized by Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2019, 516, 1078–1084. [Google Scholar] [CrossRef]
- Khakpour, H.; Younesi, H.; Mohammadhosseini, M. Two-stage biosorption of selenium from aqueous solution using dried biomass of the baker’s yeast Saccharomyces cerevisiae. J. Environ. Chem. Eng. 2014, 2, 532–542. [Google Scholar] [CrossRef]
- Zhang, L.; Li, D.; Gao, P. Expulsion of selenium/protein nanoparticles through vesicle-like structures by Saccharomyces cerevisiae under microaerophilic environment. World J. Microbiol. Biotechnol. 2012, 28, 3381–3386. [Google Scholar] [CrossRef] [PubMed]
- Turner, R.J.; Borghese, R.; Zannoni, D. Microbial processing of tellurium as a tool in biotechnology. Biotechnol. Adv. 2012, 30, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Borghese, R.; Brucale, M.; Fortunato, G.; Lanzi, M.; Mezzi, A.; Valle, F.; Cavallini, M.; Zannoni, D. Extracellular production of tellurium nanoparticles by the photosynthetic bacterium Rhodobacter capsulatus. J. Hazard. Mater. 2016, 309, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Maltman, C.; Donald, L.J.; Yurkov, V. Tellurite and Tellurate Reduction by the Aerobic Anoxygenic Phototroph Erythromonas ursincola, Strain KR99 Is Carried out by a Novel Membrane Associated Enzyme. Microorganisms 2017, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borghese, R.; Baccolini, C.; Francia, F.; Sabatino, P.; Turner, R.J.; Zannoni, D. Reduction of chalcogen oxyanions and generation of nanoprecipitates by the photosynthetic bacterium Rhodobacter capsulatus. J. Hazard. Mater. 2014, 269, 24–30. [Google Scholar] [CrossRef]
- Presentato, A.; Piacenza, E.; Anikovskiy, M.; Cappelletti, M.; Zannoni, D.; Turner, R.J. Rhodococcus aetherivorans BCP1 as cell factory for the production of intracellular tellurium nanorods under aerobic conditions. Microb. Cell Fact. 2016, 15, 204. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Zhang, X.; Kislyakov, I.M.; Dong, N.; Zhang, S.; Wang, G.; Fan, J.; Zou, X.; Du, J.; Leng, Y.; et al. Bacterially synthesized tellurium nanostructures for broadband ultrafast nonlinear optical applications. Nat. Commun. 2019, 10, 3985. [Google Scholar] [CrossRef] [Green Version]
- Pugin, B.; Cornejo, F.A.; Muñoz-Díaz, P.; Muñoz-Villagrán, C.M.; Vargas-Pérez, J.I.; Arenas, F.A.; Vásquez, C.C. Glutathione Reductase-Mediated Synthesis of Tellurium-Containing Nanostructures Exhibiting Antibacterial Properties. Appl. Environ. Microbiol. 2014, 80, 7061–7070. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, V.K.; Choi, W.; Ha, Y.; Gu, Y.; Lee, C.; Park, J.; Jang, G.; Shin, C.; Cho, S. Microbial tellurite reduction and production of elemental tellurium nanoparticles by novel bacteria isolated from wastewater. J. Indus. Eng. Chem. 2019, 78, 246–256. [Google Scholar] [CrossRef]
- Castro, L.; Li, J.; González, F.; Muñoz, J.A.; Blázquez, M.L. Green synthesis of tellurium nanoparticles by tellurate and tellurite reduction using Aeromonas hydrophila under different aeration conditions. Hydrometallurgy 2020, 196, 105415. [Google Scholar] [CrossRef]
- Zare, B.; Faramarzi, M.A.; Sepehrizadeh, Z.; Shakibaie, M.; Rezaie, S.; Shahverdi, A.R. Biosynthesis and recovery of rod-shaped tellurium nanoparticles and their bactericidal activities. Mater. Res. Bull. 2012, 47, 3719–3725. [Google Scholar] [CrossRef]
- Ramos-Ruiz, A.; Field, J.A.; Wilkening, J.V.; Sierra-Alvarez, R. Recovery of Elemental Tellurium Nanoparticles by the Reduction of Tellurium Oxyanions in a Methanogenic Microbial Consortium. Environ. Sci. Technol. 2016, 50, 1492–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottosson, L.G.; Logg, K.; Ibstedt, S.; Sunnerhagen, P.; Kall, M.; Blomberg, A.; Warringer, J. Sulfate assimilation mediates tellurite reduction and toxicity in Saccharomyces cerevisiae. Eukaryot. Cell 2010, 9, 1635–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Wong, K.H.; Yang, Y.; Li, X.; Jiang, J.; Zheng, W.; Wu, H.; Chen, T. Purification and in vitro antioxidant activities of tellurium-containing phycobiliproteins from tellurium-enriched Spirulina platensis. Drug Des. Devel. Ther. 2014, 8, 1789–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenas-Salinas, M.; Vargas-Perez, J.I.; Morales, W.; Pinto, C.; Munoz-Diaz, P.; Cornejo, F.A.; Pugin, B.; Sandoval, J.M.; Diaz-Vasquez, W.A.; Munoz-Villagran, C.; et al. Flavoprotein-Mediated Tellurite Reduction: Structural Basis and Applications to the Synthesis of Tellurium-Containing Nanostructures. Front. Microbiol. 2016, 7, 1160. [Google Scholar] [CrossRef]
- Klonowska, A.; Heulin, T.; Vermeglio, A. Selenite and tellurite reduction by Shewanella oneidensis. Appl. Environ. Microbiol. 2005, 71, 5607–5609. [Google Scholar] [CrossRef] [Green Version]
- Alam, H.; Khatoon, N.; Raza, M.; Ghosh, P.C.; Sardar, M. Synthesis and Characterization of Nano Selenium Using Plant Biomolecules and Their Potential Applications. BioNanoScience 2019, 9, 96–104. [Google Scholar] [CrossRef]
- Gunti, L.; Dass, R.S.; Kalagatur, N.K. Phytofabrication of Selenium Nanoparticles From Emblica officinalis Fruit Extract and Exploring Its Biopotential Applications: Antioxidant, Antimicrobial, and Biocompatibility. Front. Microbiol. 2019, 10, 931. [Google Scholar] [CrossRef] [Green Version]
- Sowndarya, P.; Ramkumar, G.; Shivakumar, M.S. Green synthesis of selenium nanoparticles conjugated Clausena dentata plant leaf extract and their insecticidal potential against mosquito vectors. Artif. Cells Nanomed. Biotechnol. 2017, 45, 1490–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marslin, G.; Siram, K.; Maqbool, Q.; Selvakesavan, R.K.; Kruszka, D.; Kachlicki, P.; Franklin, G. Secondary Metabolites in the Green Synthesis of Metallic Nanoparticles. Materials 2018, 11. [Google Scholar] [CrossRef] [Green Version]
- Mittal, A.K.; Kumar, S.; Banerjee, U.C. Quercetin and gallic acid mediated synthesis of bimetallic (silver and selenium) nanoparticles and their antitumor and antimicrobial potential. J. Colloid Interface Sci. 2014, 431, 194–199. [Google Scholar] [CrossRef]
- Cobbett, C.S. Heavy Metal Detoxification in Plants: Phytochelatin Biosynthesis and Function. IUBMB Life 2001, 51, 183–188. [Google Scholar] [CrossRef]
- Branco-Neves, S.; Soares, C.; de Sousa, A.; Martins, V.; Azenha, M.; Gerós, H.; Fidalgo, F. An efficient antioxidant system and heavy metal exclusion from leaves make Solanum cheesmaniae more tolerant to Cu than its cultivated counterpart. Food Energ. Secur. 2017, 6, 123–133. [Google Scholar] [CrossRef]
- Schiavon, M.; Pilon-Smits, E.A. Selenium Biofortification and Phytoremediation Phytotechnologies: A Review. J. Environ. Qual. 2017, 46, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Vogrincic, M.; Cuderman, P.; Kreft, I.; Stibilj, V. Selenium and Its Species Distribution in Above-ground Plant Parts of Selenium Enriched Buckwheat (Fagopyrum esculentum Moench). Anal. Sci. 2009, 25, 1357–1363. [Google Scholar] [CrossRef] [Green Version]
- Hu, T.; Li, H.; Li, J.; Zhao, G.; Wu, W.; Liu, L.; Wang, Q.; Guo, Y. Absorption and Bio-Transformation of Selenium Nanoparticles by Wheat Seedlings (Triticum aestivum L.). Front. Plant. Sci. 2018, 9, 597. [Google Scholar] [CrossRef] [Green Version]
- Anu, K.; Devanesan, S.; Prasanth, R.; AlSalhi, M.S.; Ajithkumar, S.; Singaravelu, G. Biogenesis of selenium nanoparticles and their anti-leukemia activity. J. King Saud Univ. Sci. 2020, 32, 2520–2526. [Google Scholar] [CrossRef]
- Fan, D.; Li, L.; Li, Z.; Zhang, Y.; Ma, X.; Wu, L.; Zhang, H.; Guo, F. Biosynthesis of selenium nanoparticles and their protective, antioxidative effects in streptozotocin induced diabetic rats. Sci. Technol. Adv. Mater. 2020, 21, 505–514. [Google Scholar] [CrossRef]
- Mulla, N.A.; Otari, S.V.; Bohara, R.A.; Yadav, H.M.; Pawar, S.H. Rapid and size-controlled biosynthesis of cytocompatible selenium nanoparticles by Azadirachta indica leaves extract for antibacterial activity. Mater. Lett. 2020, 264, 127353. [Google Scholar] [CrossRef]
- Alagesan, V.; Venugopal, S. Green Synthesis of Selenium Nanoparticle Using Leaves Extract of Withania somnifera and Its Biological Applications and Photocatalytic Activities. BioNanoScience 2019, 9, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Anu, K.; Singaravelu, G.; Murugan, K.; Benelli, G. Green-Synthesis of Selenium Nanoparticles Using Garlic Cloves (Allium sativum): Biophysical Characterization and Cytotoxicity on Vero Cells. J. Clust. Sci. 2016, 28, 551–563. [Google Scholar] [CrossRef]
- Li, S.; Shen, Y.; Xie, A.; Yu, X.; Zhang, X.; Yang, L.; Li, C. Rapid, room-temperature synthesis of amorphous selenium/protein composites using Capsicum annuum L extract. Nanotechnology 2007, 18, 405101. [Google Scholar] [CrossRef]
- Ramamurthy, C.; Sampath, K.S.; Arunkumar, P.; Kumar, M.S.; Sujatha, V.; Premkumar, K.; Thirunavukkarasu, C. Green synthesis and characterization of selenium nanoparticles and its augmented cytotoxicity with doxorubicin on cancer cells. Bioprocess. Biosyst. Eng. 2013, 36, 1131–1139. [Google Scholar] [CrossRef]
- Vennila, K.; Chitra, L.; Balagurunathan, R.; Palvannan, T. Comparison of biological activities of selenium and silver nanoparticles attached with bioactive phytoconstituents: Green synthesized using Spermacoce hispida extract. Adv. Nat. Sci. Nanosci. Nanotechnol. 2018, 9, 015005. [Google Scholar] [CrossRef]
- Kokila, K.; Elavarasan, N.; Sujatha, V. Diospyros montana leaf extract-mediated synthesis of selenium nanoparticles and their biological applications. New J. Chem. 2017, 41, 7481–7490. [Google Scholar] [CrossRef]
- Rajaee Behbahani, S.; Iranbakhsh, A.; Ebadi, M.; Majd, A.; Ardebili, Z.O. Red elemental selenium nanoparticles mediated substantial variations in growth, tissue differentiation, metabolism, gene transcription, epigenetic cytosine DNA methylation, and callogenesis in bittermelon (Momordica charantia); an in vitro experiment. PLoS ONE 2020, 15, e0235556. [Google Scholar] [CrossRef]
- Cui, D.; Liang, T.; Sun, L.; Meng, L.; Yang, C.; Wang, L.; Liang, T.; Li, Q. Green synthesis of selenium nanoparticles with extract of hawthorn fruit induced HepG2 cells apoptosis. Pharm. Biol. 2018, 56, 528–534. [Google Scholar] [CrossRef] [Green Version]
- Fardsadegh, B.; Vaghari, H.; Mohammad-Jafari, R.; Najian, Y.; Jafarizadeh-Malmiri, H. Biosynthesis, characterization and antimicrobial activities assessment of fabricated selenium nanoparticles using Pelargonium zonale leaf extract. Green Process. Synth. 2019, 8, 191–198. [Google Scholar] [CrossRef]
- Fardsadegh, B.; Jafarizadeh-Malmiri, H. Aloe vera leaf extract mediated green synthesis of selenium nanoparticles and assessment of their in vitro antimicrobial activity against spoilage fungi and pathogenic bacteria strains. Green Process. Synth. 2019, 8, 399–409. [Google Scholar] [CrossRef]
- Hassanien, R.; Abed-Elmageed, A.A.I.; Husein, D.Z. Eco-Friendly Approach to Synthesize Selenium Nanoparticles: Photocatalytic Degradation of Sunset Yellow Azo Dye and Anticancer Activity. Chem. Select 2019, 4, 9018–9026. [Google Scholar] [CrossRef]
- Mellinas, C.; Jimenez, A.; Garrigos, M.D.C. Microwave-Assisted Green Synthesis and Antioxidant Activity of Selenium Nanoparticles Using Theobroma Cacao L. Bean Shell Extract. Molecules 2019, 24, 4048. [Google Scholar] [CrossRef] [Green Version]
- Menon, S.; Devi, K.S.S.; Agarwal, H.; Shanmugam, V.K. Efficacy of Biogenic Selenium Nanoparticles from an Extract of Ginger towards Evaluation on Anti-Microbial and Anti-Oxidant Activities. Colloid Interface Sci. Commun. 2019, 29, 1–8. [Google Scholar] [CrossRef]
- Wang, L.; Li, C.; Huang, Q.; Fu, X. Biofunctionalization of selenium nanoparticles with a polysaccharide from Rosa roxburghii fruit and their protective effect against H2O2-induced apoptosis in INS-1 cells. Food Funct. 2019, 10, 539–553. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, J.; Ding, D.; Zhang, L.; Muehlmann, L.A.; Deng, S.E.; Wang, X.; Li, W.; Zhang, W. Synthesis and antioxidant properties of Lycium barbarum polysaccharides capped selenium nanoparticles using tea extract. Artif. Cell Nanomed. Biotechnol. 2017, 46, 1463–1470. [Google Scholar] [CrossRef]
- Ezhuthupurakkal, P.B.; Polaki, L.R.; Suyavaran, A.; Subastri, A.; Sujatha, V.; Thirunavukkarasu, C. Selenium nanoparticles synthesized in aqueous extract of Allium sativum perturbs the structural integrity of Calf thymus DNA through intercalation and groove binding. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 74, 597–608. [Google Scholar] [CrossRef]
- Tanaka, Y.K.; Takada, S.; Kumagai, K.; Kobayashi, K.; Hokura, A.; Ogra, Y. Elucidation of tellurium biogenic nanoparticles in garlic, Allium sativum, by inductively coupled plasma-mass spectrometry. J. Trace Elem. Med. Biol. 2020, 62, 126628. [Google Scholar] [CrossRef]
- Anan, Y.; Yoshida, M.; Hasegawa, S.; Katai, R.; Tokumoto, M.; Ouerdane, L.; Lobinski, R.; Ogra, Y. Speciation and identification of tellurium-containing metabolites in garlic, Allium sativum. Metallomics 2013, 5, 1215–1224. [Google Scholar] [CrossRef]
- Cowgill, U.M. The Tellurium Content of Vegetation. Biol. Trace Elem. Res. 1988, 17, 43–67. [Google Scholar] [CrossRef]
- Yurkov, V.; Jappé, J.; Verméglio, A. Tellurite Resistance and Reduction by Obligately Aerobic Photosynthetic Bacteria. Appl. Environ. Microbiol. 1996, 62, 4195–4198. [Google Scholar] [CrossRef] [Green Version]
- Sridharan, K.; Ollakkan, M.S.; Philip, R.; Park, T.J. Non-hydrothermal synthesis and optical limiting properties of one-dimensional Se/C, Te/C and Se–Te/C core–shell nanostructures. Carbon 2013, 63, 263–273. [Google Scholar] [CrossRef]
- Fu, S.; Cai, K.; Wu, L.; Han, H. One-step synthesis of high-quality homogenous Te/Se alloy nanorods with various morphologies. CrystEngComm 2015, 17, 3243–3250. [Google Scholar] [CrossRef]
- Baesman, S.M.; Stolz, J.F.; Kulp, T.R.; Oremland, R.S. Enrichment and isolation of Bacillus beveridgei sp. nov., a facultative anaerobic haloalkaliphile from Mono Lake, California, that respires oxyanions of tellurium, selenium, and arsenic. Extremophiles 2009, 13, 695–705. [Google Scholar] [CrossRef]
- Bajaj, M.; Winter, J. Se (IV) triggers faster Te (IV) reduction by soil isolates of heterotrophic aerobic bacteria: Formation of extracellular SeTe nanospheres. Microb. Cell Fact. 2014, 13, 168. [Google Scholar] [CrossRef]
- Thanh, N.T.K.; Green, L.A.W. Functionalisation of nanoparticles for biomedical applications. Nano Today 2010, 5, 213–230. [Google Scholar] [CrossRef]
- Salata, O.V. Applications of nanoparticles in biology and medicine. J. Nanobiotechnol. 2004, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Zamora-Ledezma, C.; Clavijo, C.D.F.; Medina, E.; Sinche, F.; Santiago Vispo, N.; Dahoumane, S.A.; Alexis, F. Biomedical Science to Tackle the COVID-19 Pandemic: Current Status and Future Perspectives. Molecules 2020, 25, 4620. [Google Scholar] [CrossRef]
- Mout, R.; Moyano, D.F.; Rana, S.; Rotello, V.M. Surface functionalization of nanoparticles for nanomedicine. Chem. Soc. Rev. 2012, 41, 2539–2544. [Google Scholar] [CrossRef]
- Soloviev, M. Nanobiotechnology today: Focus on nanoparticles. J. Nanobiotechnol. 2007, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, X.; Xu, T. Elemental selenium at nano size (Nano-Se) as a potential chemopreventive agent with reduced risk of selenium toxicity: Comparison with se-methylselenocysteine in mice. Toxicol. Sci. 2008, 101, 22–31. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, J.; Yu, H. Elemental selenium at nano size possesses lower toxicity without compromising the fundamental effect on selenoenzymes: Comparison with selenomethionine in mice. Free Radic. Biol. Med. 2007, 42, 1524–1533. [Google Scholar] [CrossRef]
- Kheradmand, E.; Rafii, F.; Yazdi, M.H.; Sepahi, A.A.; Shahverdi, A.R.; Oveisi, M.R. The antimicrobial effects of selenium nanoparticle-enriched probiotics and their fermented broth against Candida albicans. DARU J. Pharm. Sci. 2014, 22, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakibaie, M.; Adeli-Sardou, M.; Mohammadi-Khorsand, T.; ZeydabadiNejad, M.; Amirafzali, E.; Amirpour-Rostami, S.; Ameri, A.; Forootanfar, H. Antimicrobial and Antioxidant Activity of the Biologically Synthesized Tellurium Nanorods; A Preliminary In vitro Study. Iran. J. Biotechnol. 2017, 15, 268–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.; Gershwin, M.E. Drugs and autoimmunity -a contemporary review and mechanistic approach. J. Autoimmun. 2010, 34, J266–J275. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Takano, H.; Yanagisawa, R.; Ichinose, T.; Sakurai, M.; Yoshikawa, T. Effects of nano particles on cytokine expression in murine lung in the absence or presence of allergen. Arch. Toxicol. 2006, 80, 614–619. [Google Scholar] [CrossRef]
- Liao, C.; Li, Y.; Tjong, S.C. Bactericidal and Cytotoxic Properties of Silver Nanoparticles. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.S.; Yasser, M.M.; Sholkamy, E.N.; Ali, A.M.; Mehanni, M.M. Anticancer activity of biostabilized selenium nanorods synthesized by Streptomyces bikiniensis strain Ess_amA-1. Int. J. Nanomed. 2015, 10, 3389–3401. [Google Scholar] [CrossRef] [Green Version]
- Filipe, V.; Hawe, A.; Jiskoot, W. Critical evaluation of Nanoparticle Tracking Analysis (NTA) by NanoSight for the measurement of nanoparticles and protein aggregates. Pharm. Res. 2010, 27, 796–810. [Google Scholar] [CrossRef] [Green Version]
- Prasad, K.S.; Vyas, P.; Prajapati, V.; Patel, P.; Selvaraj, K. Biomimetic synthesis of selenium nanoparticles using cell-free extract of Microbacterium sp. ARB05. Micro. Nano Lett. 2012, 7, 1–4. [Google Scholar] [CrossRef]
- Selenius, M.; Rundlo, A.-K.; Olm, E.; Fernandes, A.P.; Björnstedt, M. Selenium and the Selenoprotein Thioredoxin Reductase in the Prevention, Treatment and Diagnostics of Cancer. Antioxid. Redox Sign. 2010, 12, 867–880. [Google Scholar] [CrossRef]
- Bao, P.; Xiao, K.Q.; Wang, H.J.; Xu, H.; Xu, P.P.; Jia, Y.; Haggblom, M.M.; Zhu, Y.G. Characterization and Potential Applications of a Selenium Nanoparticle Producing and Nitrate Reducing Bacterium Bacillus oryziterrae sp. nov. Sci. Rep. 2016, 6, 34054. [Google Scholar] [CrossRef]
- Schrauzer, G.N. Anticarcinogenic effects of selenium. CMLS Cell Mol. Life Sci. 2000, 57, 1864–1873. [Google Scholar] [CrossRef]
- Zeng, H.; Combs, G.F., Jr. Selenium as an anticancer nutrient: Roles in cell proliferation and tumor cell invasion. J. Nutr. Biochem. 2008, 19, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.D.; Cruz, D.M.; Roy, A.K.; Webster, T.J. Synthesis and characterization of PVP-coated tellurium nanorods and their antibacterial and anticancer properties. J. Nanopart. Res. 2018, 20, 254. [Google Scholar] [CrossRef]
- Mohanty, A.; Kathawala, M.H.; Zhang, J.; Chen, W.N.; Chye Loo, J.S.; Kjelleberg, S.; Yang, L.; Cao, B. Biogenic Tellurium Nanorods as a Novel Antivirulence Agent Inhibiting Pyoverdine Production in Pseudomonas aeruginosa. Biotechnol. Bioeng. 2013, 111, 858–865. [Google Scholar] [CrossRef] [PubMed]
- Aydin, N.; Arslan, M.E.; Sonmez Sonmez, E.; Turkez, H. Cytotoxicity analysis of tellurium dioxide nanoparticles on cultured human pulmonary alveolar epithelial and peripheral blood cell cultures. Biomed. Res. India 2017, 28, 3300–3304. [Google Scholar]
- Forootanfar, H.; Amirpour-Rostami, S.; Jafari, M.; Forootanfar, A.; Yousefizadeh, Z.; Shakibaie, M. Microbial-assisted synthesis and evaluation the cytotoxic effect of tellurium nanorods. Mater. Sci. Eng. C Mater. Biol. Appl. 2015, 49, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Zhong, J.; Shen, B.; Gan, T.; Fu, C.; Zhu, Z.; Li, R.; Yang, X. Comparative study of the cytotoxicity of the nanosized and microsized tellurium powders on HeLa cells. Front. Biol. 2013, 8, 444–450. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Localization | Precursor | Concentration (mM) | Incubation Temperature and Time | Size (nm) * | Color and Shape | Z-Potential (mV) | Sample Quantification | Activity/Application | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|
| Staphylococcus carnosus | Intracellular | Na2SeO3 | 1−5 | 37 °C for 72 h | 439–525 | Red Spherical | −26.13 and −20.40 | Cocktail of proteins derived from S. carnosus | Agriculture Future medicine | [109] |
| Bacillus mycoides Stenotrophomonas maltophilia | Cell free extract | Na2SeO3 | 2 | 27 °C for 6 h or 24 h | 160–171 | Spherical | −70 and −80 | C: 73–75% O: 10–11% Se: 9–11% P: 3–5% S: 1% | Antibacterial Antibiofilm | [114] |
| Acinetobacter schindleri Staphylococcus sciuriExiguobacterium acetylicum Enterobacter cloacae | Near the cell membrane | Na2SeO3 | 10–50 | 25 or 37 °C for 24 h | ~100 | Spherical Transformation to nanowires | N/A | Se: 83.9% | Antibacterial | [115] |
| Stenotrophomonas bentonitica | Intracellular Extracellular | Na2SeO3 | 2 | 28 °C for 48 h | 30–400 (~34) | Orange-red Spherical Hexagonal Polygonal Nanowires | N/A | Extracellular flagella-like proteins | Bioremediation, Safety of deep geological repository systems | [74] |
| Shewanella sp. | N/A | Na2SeO3 | 0.01–1.0 | 30 °C for 24 h | 1–20 | Spherical | N/A | N/A | N/A | [116] |
| Bacillus sp. | Intracellular. Associated to cell debris | SeO2 | 1.26 | 30 °C for 24 h | 80–220 | Red Spherical | –16.3 | Se: 100% | Anticancer Antibiofilm Antiparasitic Antioxidant | [117,118,119,120] |
| Azoarcus sp. | Extracellular Associated to cell debris | Na2SeO3 | 1–8 | 30 °C for 24 h | 123 | Orange Spherical | N/A | N/A | Agriculture Bioremediation | [121] |
| Acinetobacter sp. | Intracellular | Na2SeO3 | 0.1–4 | 30 °C for 24 h | ~100 | Red Spherical Rod shaped polygonal | +10 | Proteins Amines Amides | Anticancer | [58] |
| Duganella sp. Agrobacterium sp. | Cell surface Extracellular polymeric substances (EPS)Culture medium | Na2SeO3 Na2SeO4 | 4 g L−1 2 g L−1 | 28 ± 2 °C | 100–220 | Red Spherical | N/A | Proteins | Agriculture | [110] |
| Burkholderia fungorum | Mostly extracellular | Na2SeO3 | 0.5–2 | 27 °C for 96 h | 170–200 | Red-orange Spherical | From −25 to +20 | Proteins | Bioremediation | [122] |
| Comamonas testosteroni | Intracellular: cytoplasm or periplasm | Se(IV) and Se(VI) | 5 | 28 °C for 48 h | 100–200 | Red fine-grained | N/A | Selenium content 100% | Bioremediation | [123] |
| Bacillus subtilis | Extracellular | Selenite | 4 | 48 °C for 48 h | 50–400 | Red Spherical monoclinic that can transform to anisotropic 1D trigonal structure (nanowires) | N/A | Proteins Biopolymers | Biosensing | [101] |
| Alishewanella sp. | Intracellular | Na2SeO3 | 1 | 37 °C for 4 h | 100–220 | Spherical | −28.7 | Proteins Lipids Organic substances Inorganic ions | Bioremediation | [75] |
| Azospirillum brasilense | Intracellular Extracellular | Na2SeO3 | 10 | 31 °C for 24 h | 50–100 | Spherical | −21 to −24 | Proteins Polysaccharides Lipids | N/A | [124] |
| Azospirillum brasilense | Extracellular | Na2SeO3 Na2SeO4 | 1–5 | 30 °C | 400 | Red Spherical | −18 | Proteins Carbohydrates EPS | Bioremediation Biotechnological applications | [125] |
| Pseudomonas aeruginosa | Cell surface | Selenite | 0.25–1.0 | 37 °C for 24–72 h | 47–165 (~96) | Red Spherical | 251.8 | Proteins | Bioremediation | [126] |
| Stenotrophomonas maltophilia | Intracellular Released to the medium | Na2SeO3 | 0.5–5.0 | 27 °C for 24 and 48 h | 160–250 | Spherical | 140 | Proteins Carbohydrates Lipids | Bioremediation | [113] |
| Bacillus cereus | Intracellular | Na2SeO3 | 0.5–1200 | 30 °C for 24 h | 170 | Red Spherical | N/A | N/A | Medicine Veterinary medicine | [127] |
| Zooglea ramigera | Extracellular | Na2SeO3 | 3 | 30 °C for 48 h | 30–150 | Red Spherical Nanorods (trigonal) | N/A | Enzymes Proteins Bacterial material | N/A | [128] |
| Pseudomonas sp. Lysinibacillus Thauera selenatis | N/A | Na2SeO3 | 200 | 30 °C for 40 days | N/A | Red Spherical | N/A | Reduced in the presence of nitrate | Denitrification of mine wastewater | [129] |
| Escherichia coli | Intracellular Extracellular | Na2SeO3 | 1 | N/A | 50–100 | Spherical | N/A | Quinone-mediated | N/A | [97] |
| Acinetobacter sp. | Intracellular | Na2SeO3 | 1 | 37 °C for 24 h | 100 ± 10 | Orange Spherical amorphous | N/A | Lignin peroxidase | N/A | [130] |
| Enterococcus faecalis | Extracellular | Na2SeO3 | 0.19–2.97 | 37 and 42 °C for 24 and 48 h | 29–195 | Red/light red Spherical | N/A | N/A | Antibacterial | [55] |
| Streptomyces minutiscleroticus | Extracellular | Na2SeO3 | 1 | 48–72 h | 100–250 | Red Spherical | N/A | Proteins | Wound ointment Anticancer drug Coating for medical instruments | [131] |
| Streptomyces griseobrunneus | N/A | N/A | N/A | 30 °C | 48–136 | Red Trigonal | N/A | Proteins Enzymes | Photocatalytic | [132] |
| Vibrio natriegens | Intracellular Associated to cell debris | Na2SeO4 Na2SeO3 | 1 | 30 °C for 24 h | 136 ± 31 | Red Spherical | N/A | Proteins | Bioremediation | [133] |
| Staphylococcus aureus Methicillin-resistant Staphylococcus aureus (MRSA) Escherichia coli Pseudomonas aeruginosa | Intracellular Associated to cell debris | Na2SeO3 | 2 | 37 °C for 72 h | 90–150 | Orange-red | N/A | Lipids Proteins | Antimicrobial | [61] |
| Rhodococcus aetherivorans | Extracellular | Na2SeO3 | 0.5–2 | 40 °C for 40 min then cooled to RT | 53–97 | Spherical Nanorods | −13 to −32 | Organic material | N/A | [99] |
| Pseudomonas stutzeri | Intracellular | Na2SeO3 | 2.5 | 28 °C | 100–250 | Reddish Spherical | −19.5 | Proteins Lipids Other organic substances | N/A | [46] |
| Lactobacillus casei | Intracellular | Na2SeO3 | 1.2 | 37 °C for 24 h | 50–80 | Red Spherical | N/A | Polysaccharides Proteins | Antioxidant Anticancer | [134] |
| Streptomyces enissocaesilis | Extracellular | SeO2 | 5 | 30 °C for 72 h | 20–211 | Brown, orange and deep yellow Spherical | −220 | Proteins | Antimicrobial | [135] |
| Pseudomonas stutzeri | N/A | Na2SeO3 | 1–3 | 37 °C for 48 h | 75–200 | Bright red Spherical | −46.2 | Proteins Organic molecules | Antiangiogenic Antiproliferative | [103] |
| Streptomyces sp. | Extracellular | Na2SeO3 | 1 | 28 °C for 72–96 h | 20–150 | Red Spherical | N/A | Free amines Aromatic rings Cysteine residues Amides | Antibacterial Larvicidal Anthelminthic | [136] |
| Lysinibacillus sp. | Extracellular | Na2SeO3 | 1 | 37 °C for 3 days | 130 | Red Spherical | −19.1 to −28.8 | Proteins Polysaccharides Fatty acids | Antibiofilm Antimicrobial | [137] |
| Lactobacillus acidophilus L. plantarum L. rhamnosus | Extracellular | Na2SeO3 | 4 | 35° for 48 h | 20–80 | Red | N/A | Proteins | N/A | [76] |
| Idiomarina sp. | Intracellular | Na2SeO3 | 4 and 8 | 37 °C for 48 h | 35 and 150–350 | Brick red Spherical/Hexagonal | N/A | N/A | Antineoplastic Anticancer | [138] |
| Ralstonia eutropha | Extracellular | Na2SeO4 | 1.5 | 30 °C for 48 h | 40–120 | Red Spherical/Nanorods | −7.7 | N/A | Antibacterial | [139] |
| Pseudomonas stutzeri | Extracellular Cell surface | Na2SeO4 Na2SeO3 | 5 and 11 mM 4 and 9 mM | 34 °C for 7 days | ≤200 | Red Spherical | N/A | N/A | Bioremediation | [84] |
| Enterobacter cloacae | Intracellular Extracellular | Na2SeO3 | 0.5–15 | 37 °C for 8 h | 100–300 | Red Rod-shaped | N/A | Organic material | N/A | [140] |
| Bacillus cereus | Intracellular Extracellular | Na2SeO3 | 0.5–10 | 37 °C for 48 h | 150–200 | Spherical | −46.86 | Proteins | N/A | [56] |
| Stenotrophomonas maltophilia Ochrobactrum sp. | N/A | Na2SeO3 | 0.5 | 27 °C for 24 and 48 h | 357 | Spherical | N/A | Organic compounds | Antimicrobial Antibiofilm | [71] |
| Shewanella oneidensis | Cell surface Extracellular | Selenite | 0.5 | 30 °C for 6–48 h | 20 | Red Spherical | N/A | EPS | N/A | [141] |
| Synechococcus leopoliensis | Intracellular Extracellular | Na2SeO3 | 5 | 35 °C | 254 ± 52 200 ± 37 | Red-brown Fused spheres Elongated rods | N/A | N/A | N/A | [142] |
| Comamonas testosteroni | Extracellular | Na2SeO3 | 0.2–50 | 28 °C for 24 h | 100–200 | Red Round Rod-shaped | N/A | Proteins | Bioremediation | [143] |
| Azospirillum brasilense | Extracellular | Na2SeO3 | 10–50 | 31–32 °C for 24 h | 25–80 | Red-orange Spherical | −21 to −24 | N/A | N/A | [144] |
| Bacillus cereus | Cell surface | Na2SeO3 | 0.25–1.0 | 37 °C for 24–72 h | 50–150 (~93) | Red Rod-shaped | −31.1 ± 4.9 | N/A | Bioremediation | [145] |
| Bacillus sp. | Extracellular | SeO2 | 6.4 | 33 °C for 72 h | 31–335 (~126) | Red-orange Spherical | N/A | Alcohols Phenols Amides Amines Amino acids | Antioxidant | [83] |
| Species | Location | Size (nm) | Shape | Activity/Application | Ref. |
|---|---|---|---|---|---|
| Trichoderma sp. | Extracellular | 20–220 | Spherical Pseudospherical | N/A | [182] |
| Pleurotus ostreatus | Aqueous extract | 7–28 | Spherical | Antioxidant Antimicrobial Anticancer | [185] |
| Penicillium chrysogenum | Cell-free supernatant | 48–50 | Spherical | Antimicrobial Antibiofilm | [186] |
| Phanerochaete chrysosporium | Intracellular Extracellular | 50–600 | Spherical | Bioremediation | [65] |
| Polyporus umbellatus | N/A | 212 ± 23 82 ± 1 | Spherical | Anticancer Antiproliferative | [187] |
| Auricularia auricula-judae | Embedded in triple helix β-(1,3)-D-glucan | 60 | Hollow nanotubes | Acute myeloid leukemia (AML) therapy | [188] |
| Trichoderma atroviride | Culture filtrate (CF) Cell lysate (CL) Cell wall debris (CW) | 60–123 | Spherical | Production of crop plants (tomatoes) Management of plant diseases | [181] |
| Aureobasidium pullulans Mortierella humilis Trichoderma harzianum Phoma glomerata | Extracellular | 48–78 | Spindle-shaped | Bioremediation | [51] |
| Dictyophora indusiata | Intracellular | 89 | Spherical | Anticancer | [189] |
| Catathelasma ventricosum | N/A | 50 | Spherical | Antidiabetic | [190] |
| Aspergillus oryzae | N/A | 55 | Spherical | Antimicrobial | [184] |
| Pyrenochaeta sp. Acremonium strictum Plectosphaerella cucumerina Stagonospora sp. Alternaria alternata Paraconiothyrium sporulosum | Fungal hyphae Intracellular Extracellular | 50–300 | Spherical | N/A | [191] |
| Alternaria alternata | Extracellular | 30–150 | Spherical | N/A | [180] |
| Pleurotus ostreatus Lentinus edodes Ganoderma lucidum Grifola frondosa | Intracellular Extracellular | 50–150 | Spherical | N/A | [192] |
| Lentinula edodes | Intracellular (fungal hyphae) | 180 ± 17 | N/A | N/A | [193] |
| Pleurotus ostreatus Ganoderma lucidum Grifola frondosa | Intracellular Cell-free filtrate | 20–550 | N/A | N/A | [194] |
| Cordyceps sinensis | N/A | 80–125 | Spherical | Antioxidant | [195] |
| Mariannaea sp. | Intracellular Extracellular | 45 213 | Spherical | N/A | [196] |
| Gliocladium roseum | Cell-free filtrate | 20–80 | Spherical | N/A | [197] |
| Plant Species | Part | Metabolites | Shape | Size (nm) | Activity/Application | Ref. |
|---|---|---|---|---|---|---|
| Withania somnifera | Leaves | Flavonoids Phenolics Tannins | Spherical | 40–90 | Antibacterial Antioxidant Anticancer | [240] |
| Psidium guajava | Leaves | N/A | Spherical | 8–20 | Antibacterial | [227] |
| Allium sativum | Cloves | N/A | Spherical | 40–100 | Cytotoxicity | [241] |
| Cassia auriculata | Leaves | N/A | Amorphous | 10–20 | Anti-leukemia | [237] |
| Momordica charantia | Roots and shoots | Terpenoids Phenolics | Spherical | 10–30 | Toxicological studies | [246] |
| Hawthorn fruit | Fruit | N/A | Spherical | 113 | Antitumor | [247] |
| Hibiscus sabdariffa | Leaves | Phenols Alcohols | Spherical Triangular Hexagonal | 20–50 | Antioxidant | [238] |
| Pelargonium zonale | Leaves | N/A | Spherical | 40–60 | Antibacterial Antifungal | [248] |
| Aloe vera | Leaves | Hydroxyls Amides | Spherical | 121–3243 | Antibacterial Antifungal | [249] |
| Emblica officinali | Fruit | Phenolics Flavonoids Tannins | Spherical | 20–60 | Antimicrobial | [228] |
| Moringa oleifera | Leaves | Phenolics Flavones | Spherical | 23–35 | Anticancer | [250] |
| Triticum aestivum | Roots | N/A | Spherical | 140 ± 40 | Biofertilizer | [236] |
| Broccoli | N/A | Carotenes Glucosinolates Polyphenols | Spherical | 50–150 | Antioxidant Anticancer | [108] |
| Diospyros montana | Leaves | Phenolics Flavonoids | Spherical | 4–16 | Antibacterial Anticancer | [245] |
| Ocimum tenuiflorum | Leaves | Polyphenols | Spherical | 15–20 | Inhibition of nephrolithiasis | [183] |
| Theobroma cacao | Shell | Polysaccharides Proteins Phenolics | Spherical Trigonal | 1–3 | N/A | [251] |
| Zingiber officinale | Roots | Flavonoids Terpenoids | Spherical | 100–150 | Antimicrobial Antioxidant | [252] |
| Mucuna pruriens | Seed | Phytochemicals | Spherical Nanorods | 100–120 | Antioxidant Anticancer | [102] |
| Azadirachta indica | Leaves | Polyphenols Flavonoids Proteins | Spherical | 142–168 221–328 | Antibacterial | [239] |
| Vitis vinifera | N/A | Lignin | Spherical | 3–18 | N/A | [32] |
| Clausena dentata | Leaves | Flavonoids Triterpenoids Polyphenols | Spherical | 46–79 | Larvicidal | [229] |
| Spermacoce hispida | Leaves | Polyols Saponins | Rod-shaped | 120 ± 15 | Anti-inflammatory Antibacterial Anticancer | [244] |
| Rosa roxburghii | N/A | Polysaccharide (RTFP-3) | Spherical | 105 | Antioxidant | [253] |
| Lycium barbarum | Berries | Flavonols (catechins) | Spherical Triangular | 83–160 | Antioxidant | [254] |
| Fenugreek | Seeds | Phenol Flavonol | Oval | 50–150 | Anticancer | [243] |
| Allium sativum | Bulbs | Alcohols Phenols | Spherical | 205 | Antioxidant Anticancer | [255] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zambonino, M.C.; Quizhpe, E.M.; Jaramillo, F.E.; Rahman, A.; Santiago Vispo, N.; Jeffryes, C.; Dahoumane, S.A. Green Synthesis of Selenium and Tellurium Nanoparticles: Current Trends, Biological Properties and Biomedical Applications. Int. J. Mol. Sci. 2021, 22, 989. https://doi.org/10.3390/ijms22030989
Zambonino MC, Quizhpe EM, Jaramillo FE, Rahman A, Santiago Vispo N, Jeffryes C, Dahoumane SA. Green Synthesis of Selenium and Tellurium Nanoparticles: Current Trends, Biological Properties and Biomedical Applications. International Journal of Molecular Sciences. 2021; 22(3):989. https://doi.org/10.3390/ijms22030989
Chicago/Turabian StyleZambonino, Marjorie C., Ernesto Mateo Quizhpe, Francisco E. Jaramillo, Ashiqur Rahman, Nelson Santiago Vispo, Clayton Jeffryes, and Si Amar Dahoumane. 2021. "Green Synthesis of Selenium and Tellurium Nanoparticles: Current Trends, Biological Properties and Biomedical Applications" International Journal of Molecular Sciences 22, no. 3: 989. https://doi.org/10.3390/ijms22030989
APA StyleZambonino, M. C., Quizhpe, E. M., Jaramillo, F. E., Rahman, A., Santiago Vispo, N., Jeffryes, C., & Dahoumane, S. A. (2021). Green Synthesis of Selenium and Tellurium Nanoparticles: Current Trends, Biological Properties and Biomedical Applications. International Journal of Molecular Sciences, 22(3), 989. https://doi.org/10.3390/ijms22030989