
Split Enzyme-Based Biosensors for Structural Characterization of Soluble and Insoluble β-Glucans


Abstract
:1. Introduction
2. Results
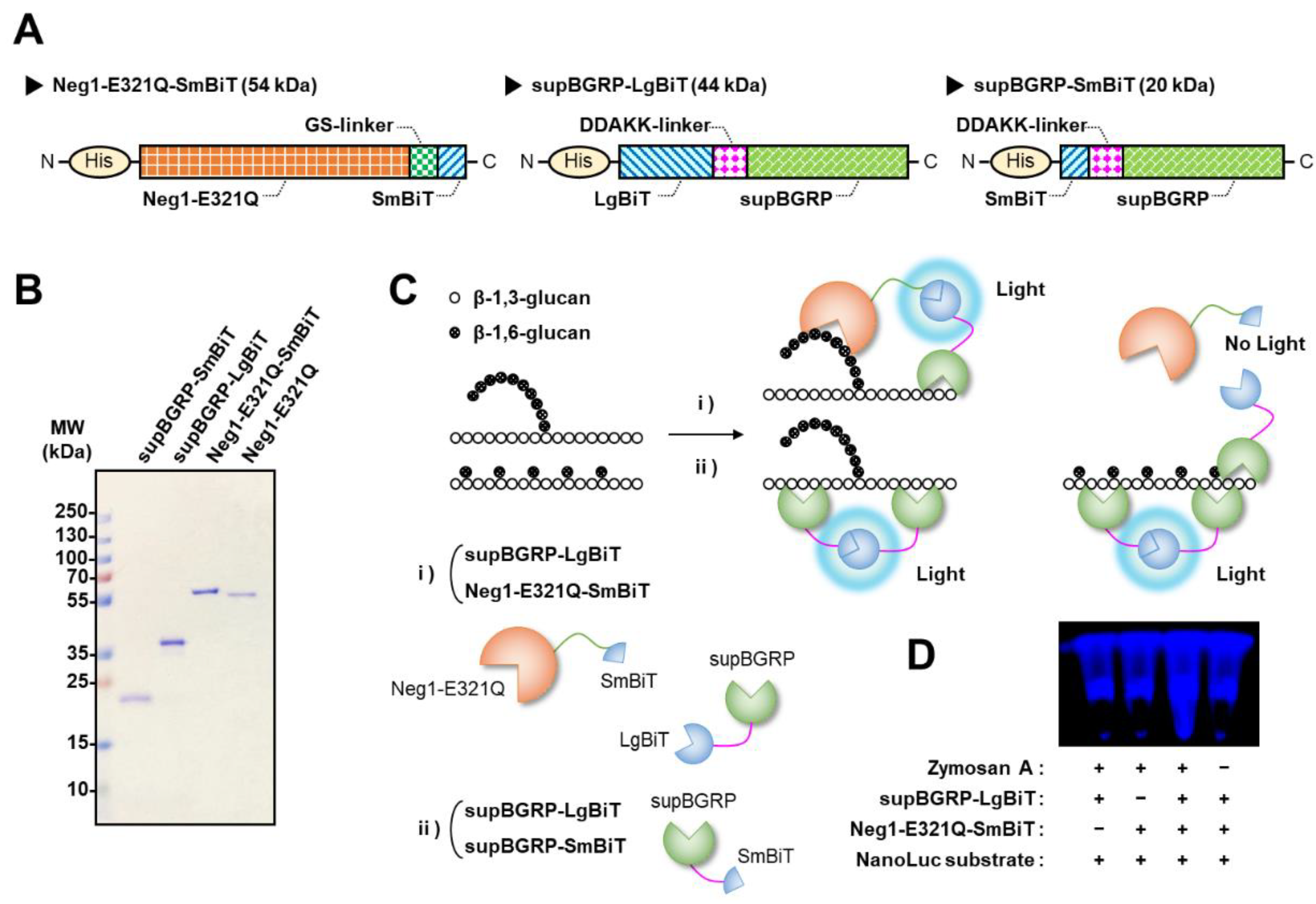
2.1. Protein Engineering for Development of New Split-NanoLuc Complementation Assay for the Structural Characterization of β-Glucan
2.2. Verification of the Reactivity of Split-NanoLuc-Fused Glucan-Binding Proteins to β-Glucans
2.3. Development of the Split-NanoLuc Complementation Assay for β-Glucan
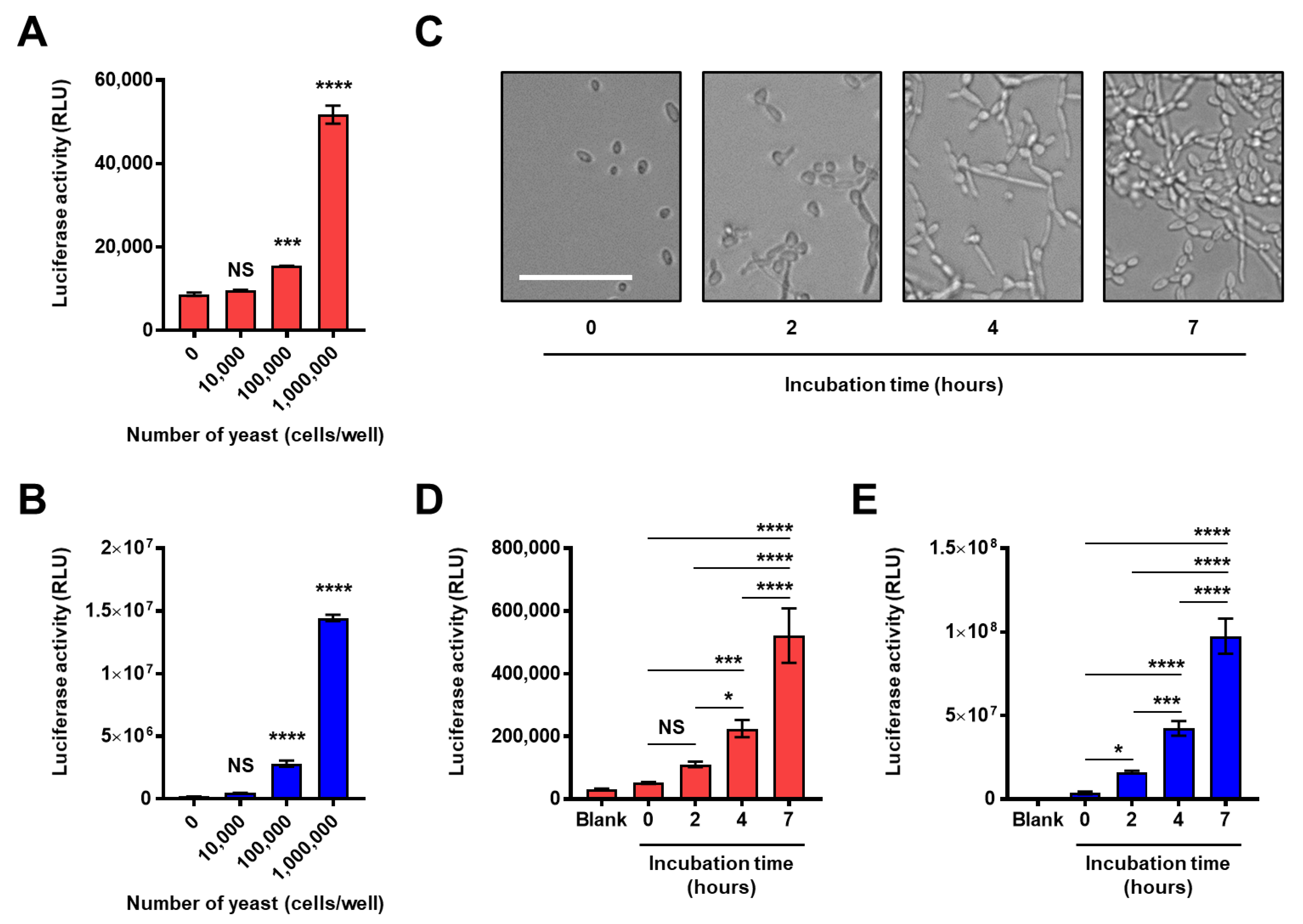
2.4. Application for the Analysis of the Composition and Structure of the Fungal Intact Cell Wall
2.5. Application of the Split-NanoLuc Complementation Assay for Real-Time Monitoring of Structural Changes in β-Glucan
2.6. Comparison of Reactivity between Conventional HRP-Based ELISA and Split-NanoLuc Complementation Assay for Detection of Soluble and Insoluble β-Glucan
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of Mushroom-Derived Polysaccharide Fraction
4.3. Preparation of HKCA and Cell-Wall Solubilized β-Glucan
4.4. Plasmid Construction
4.5. Preparation of Split Enzyme-Fused Glucan-Binding Proteins
4.6. Biotinylation of Neg1-E321Q
4.7. Sandwich ELISA-Like Assay
4.8. Split-NanoLuc Complementation Assay
4.9. Preparation of C. albicans Cells
4.10. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BGRP | β-1,3-glucan recognition protein |
| BSA | bovine serum albumin |
| CSBG | Candida soluble β-glucan |
| ELISA | Enzyme-linked immunosorbent assay |
| GEM | glucan enzymatic method |
| HKCA | heat-killed C. albicans |
| HRP | horseradish peroxidase |
| LAL | Limulus amebocyte lysate |
| PBS | phosphate-buffered saline |
References
- Lipke, P.N.; Ovalle, R. Cell Wall Architecture in Yeast: New Structure and New Challenges. J. Bacteriol. 1998, 180, 3735–3740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Z.; Otaka, K.; Maoka, T.; Hidaka, K.; Ishijima, S.; Oda, M.; Ohnishi, M. Structure of β-Glucan Oligomer from Laminarin and Its Effect on Human Monocytes to Inhibit the Proliferation of U937 Cells. Biosci. Biotechnol. Biochem. 2005, 69, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Read, S.M.; Currie, G.; Bacic, A. Analysis of the structural heterogeneity of laminarin by electrospray-ionisation-mass spectrometry. Carbohydr. Res. 1996, 281, 187–201. [Google Scholar] [CrossRef]
- Kikumoto, S.; Miyajima, T.; Kimura, K.; Okubo, S.; Komatsu, N. Polysaccharide Produced by Schizophyllum commune Part II. J. Agric. Chem. Soc. Jpn. 1971, 45, 162–168. [Google Scholar] [CrossRef] [Green Version]
- Tada, R.; Harada, T.; Nagi-Miura, N.; Adachi, Y.; Nakajima, M.; Yadomae, T.; Ohno, N. NMR characterization of the structure of a β-(1→3)-d-glucan isolate from cultured fruit bodies of Sparassis crispa. Carbohydr. Res. 2007, 342, 2611–2618. [Google Scholar] [CrossRef]
- Adachi, Y.; Ishii, M.; Kanno, T.; Tetsui, J.; Ishibashi, K.; Yamanaka, D.; Miura, N.; Ohno, N. N-Terminal (1→3)-β-D-glucan recognition proteins from insects recognize the difference in ultra-structures of (1→3)-β-D-glucan. Int. J. Mol. Sci. 2019, 20, 3498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamanaka, D.; Takatsu, K.; Kimura, M.; Swamydas, M.; Ohnishi, H.; Umeyama, T.; Oyama, F.; Lionakis, M.S.; Ohno, N. Development of a novel β-1,6-glucan–specific detection system using functionally-modified recombinant endo-β-1,6-glucanase. J. Biol. Chem. 2020, 295, 5362–5376. [Google Scholar] [CrossRef] [Green Version]
- Kanno, T.; Adachi, Y.; Ishibashi, K.-I.; Yamanaka, D.; Ohno, N. Quantification of β-Glucan from Culinary-Medicinal Mushrooms Using Novel Artificial β-Glucan Recognition Protein. Int. J. Med. Mushrooms 2020, 22, 269–276. [Google Scholar] [CrossRef]
- Hall, M.P.; Unch, J.; Binkowski, B.F.; Valley, M.P.; Butler, B.L.; Wood, M.G.; Otto, P.; Zimmerman, K.; Vidugiris, G.; Machleidt, T.; et al. Engineered Luciferase Reporter from a Deep Sea Shrimp Utilizing a Novel Imidazopyrazinone Substrate. ACS Chem. Biol. 2012, 7, 1848–1857. [Google Scholar] [CrossRef]
- Dixon, A.S.; Schwinn, M.K.; Hall, M.P.; Zimmerman, K.; Otto, P.; Lubben, T.H.; Butler, B.L.; Binkowski, B.F.; Machleidt, T.; Kirkland, T.A.; et al. NanoLuc Complementation Reporter Optimized for Accurate Measurement of Protein Interactions in Cells. ACS Chem. Biol. 2016, 11, 400–408. [Google Scholar] [CrossRef]
- Ozawa, T.; Kaihara, A.; Sato, M.; Tachihara, K.; Umezawa, Y. Split Luciferase as an Optical Probe for Detecting Protein−Protein Interactions in Mammalian Cells Based on Protein Splicing. Anal. Chem. 2001, 73, 2516–2521. [Google Scholar] [CrossRef]
- Luker, K.E.; Smith, M.C.P.; Luker, G.D.; Gammon, S.T.; Piwnica-Worms, D. Kinetics of regulated protein-protein interactions revealed with firefly luciferase complementation imaging in cells and living animals. Proc. Natl. Acad. Sci. USA 2004, 101, 12288–12293. [Google Scholar] [CrossRef] [Green Version]
- Paulmurugan, R.; Gambhir, S.S. Monitoring Protein−Protein Interactions Using Split Synthetic Renilla Luciferase Protein-Fragment-Assisted Complementation. Anal. Chem. 2003, 75, 1584–1589. [Google Scholar] [CrossRef] [PubMed]
- Pillemer, L.; Ecker, E. ANTICOMPLEMENTARY FACTOR IN FRESH YEAST. J. Biol. Chem. 1941, 137, 139–142. [Google Scholar] [CrossRef]
- Ikeda, Y.; Adachi, Y.; Ishii, T.; Miura, N.; Tamura, H.; Ohno, N. Dissociation of Toll-Like Receptor 2-Mediated Innate Immune Response to Zymosan by Organic Solvent-Treatment without Loss of Dectin-1 Reactivity. Biol. Pharm. Bull. 2008, 31, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Ohno, N.; Miura, T.; Miura, N.; Adachi, Y.; Yadomae, T. Structure and biological activities of hypochlorite oxidized zymosan. Carbohydr. Polym. 2001, 44, 339–349. [Google Scholar] [CrossRef]
- Ohno, N.; Miura, T.; Miura, N.N.; Chiba, N.; Uchiyama, M.; Adachi, Y.; Yadomae, T. Inflammatory and immunopharmacological activities of meta-periodate oxidized zymosan. Zentralblatt für Bakteriologie 1999, 289, 63–77. [Google Scholar] [CrossRef]
- Bain, J.M.; Louw, J.; Lewis, L.E.; Okai, B.; Walls, C.A.; Ballou, E.R.; Walker, L.A.; Reid, D.; Munro, C.A.; Brown, A.J.P.; et al. Candida albicans Hypha Formation and Mannan Masking of β-Glucan Inhibit Macrophage Phagosome Maturation. mBio 2014, 5, e01874-14. [Google Scholar] [CrossRef] [Green Version]
- Synytsya, A.; Novák, M. Structural diversity of fungal glucans. Carbohydr. Polym. 2013, 92, 792–809. [Google Scholar] [CrossRef] [PubMed]
- Obayashi, T.; Kawai, T.; Yoshida, M.; Mori, T.; Goto, H.; Yasuoka, A.; Shimada, K.; Iwasaki, H.; Teshima, H.; Kohno, S.; et al. Plasma (1→3)-β-D-glucan measurement in diagnosis of invasive deep mycosis and fungal febrile episodes. Lancet 1995, 345, 17–20. [Google Scholar] [CrossRef]
- Yoneda, A.; Kurokawa, T. A sensitive sandwich ELISA to measure (1→3)-β-D-glucan levels in blood. J. Immunol. Methods 2011, 365, 158–165. [Google Scholar]
- Tanaka, S.; Aketagawa, J.; Takahashi, S.; Shibata, Y.; Tsumuraya, Y.; Hashimoto, Y. Activation of a limulus coagulation factor G by (1→3)-β-D-glucans. Carbohydr. Res. 1991, 218, 167–174. [Google Scholar] [CrossRef]
- Sander, I.; Fleischer, C.; Borowitzki, G.; Br\ufcning, T.; Raulf-Heimsoth, M. Development of a two-site enzyme immunoassay based on monoclonal antibodies to measure airborne exposure to (1→3)-β-d-glucan. J. Immunol. Methods 2008, 337, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Graham, L.M.; Tsoni, S.V.; Willment, J.A.; Williams, D.L.; Taylor, P.R.; Gordon, S.; Dennehy, K.; Brown, G.D. Soluble Dectin-1 as a tool to detect β-glucans. J. Immunol. Methods 2006, 314, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Sunamura, E.; Iwasaki, M.; Shiina, S.; Kitahara, S.; Yotani, T.; Manabe, M.; Miyazaki, O. A novel enzyme immunoassay for the measurement of plasma (1→3)-β-D-glucan levels. J. Immunol. Methods 2020, 487, 112872. [Google Scholar] [CrossRef] [PubMed]
- Milton, D.K.; Alwis, K.U.; Fisette, L.; Muilenberg, M. Enzyme-linked immunosorbent assay specific for (1→6) branched, (1→3)-β-D-glucan detection in environmental samples. Appl. Environ. Microbiol. 2001, 67, 5420–5424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danielson, M.E.; Dauth, R.; Elmasry, N.A.; Langeslay, R.R.; Magee, A.S.; Will, P.M. Enzymatic Method To Measure β-1,3-β-1,6-Glucan Content in Extracts and Formulated Products (GEM Assay). J. Agric. Food Chem. 2010, 58, 10305–10308. [Google Scholar] [CrossRef]
- Ide, M.; Okumura, M.; Koizumi, K.; Kumagai, M.; Yoshida, I.; Yoshida, M.; Mishima, T.; Nakamura, M. Novel Method to Quantify β-Glucan in Processed Foods: Sodium Hypochlorite Extracting and Enzymatic Digesting (SEED) Assay. J. Agric. Food Chem. 2018, 66, 1033–1038. [Google Scholar] [CrossRef]
- Brown, G.D.; Gordon, S. A new receptor for β-glucans. Nature 2001, 413, 36–37. [Google Scholar] [CrossRef]
- Brown, G.D.; Gordon, S. Fungal β-Glucans and Mammalian Immunity. Immunity 2003, 19, 311–315. [Google Scholar] [CrossRef] [Green Version]
- Adachi, Y.; Ohno, N.; Ohsawa, M.; Oikawa, S.; Yadomae, T. Change of biological activities of (1→3)-β-D-glucan from Grifola frondosa upon molecular weight reduction by heat treatment. Chem. Pharm. Bull. 1990, 38, 477–481. [Google Scholar] [CrossRef] [Green Version]
- Volman, J.J.; Ramakers, J.D.; Plat, J. Dietary modulation of immune function by β-glucans. Physiol. Behav. 2008, 94, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.S.; Passos, C.P.; Madureira, P.; Vilanova, M.; Coimbra, M.A. Structure–function relationships of immunostimulatory polysaccharides: A review. Carbohydr. Polym. 2015, 132, 378–396. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, D.; Ishibashi, K.-I.; Adachi, Y.; Ohno, N. Quantification of Mushroom-Derived Soluble β-1,6-Glucan Using the Function-Modified Recombinant β-1,6-Glucanase. Int. J. Med. Mushrooms 2020, 22, 855–868. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, D.; Tada, R.; Adachi, Y.; Ishibashi, K.-I.; Motoi, M.; Iwakura, Y.; Ohno, N. Agaricus brasiliensis-derived β-glucans exert immunoenhancing effects via a dectin-1-dependent pathway. Int. Immunopharmacol. 2012, 14, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Ohno, N.; Uchiyama, M.; Tsuzuki, A.; Tokunaka, K.; Miura, N.N.; Adachi, Y.; Aizawa, M.W.; Tamura, H.; Tanaka, S.; Yadomae, T. Solubilization of yeast cell-wall β-(1→3)-d-glucan by sodium hypochlorite oxidation and dimethyl sulfoxide extraction. Carbohydr. Res. 1999, 316, 161–172. [Google Scholar] [CrossRef]
- Oyama, S.; Yamagata, Y.; Abe, K.; Nakajima, T. Cloning and Expression of an Endo-1,6-β-D-glucanase Gene (neg1) fromNeurospora crassa. Biosci. Biotechnol. Biochem. 2002, 66, 1378–1381. [Google Scholar] [CrossRef] [Green Version]
- Temple, M.J.; Cuskin, F.; Baslé, A.; Hickey, N.; Speciale, G.; Williams, S.J.; Gilbert, H.J.; Lowe, E.C. A Bacteroidetes locus dedicated to fungal 1,6-β-glucan degradation: Unique substrate conformation drives specificity of the key endo-1,6-β-glucanase. J. Biol. Chem. 2017, 292, 10639–10650. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Structure | Source | Elemental Composition (%) | ||
|---|---|---|---|---|---|
| C | H | N | |||
| Zymosan A | β-1,6-/β-1,3-glucan | Saccharomyces cerevisiae | 43.16 | 6.92 | 1.75 |
| Curdlan | Linear β-1,3-glucan | Alcaligenes faecalis | 39.67 | 6.75 | 0.03 |
| Pustulan | β-1,6-glucan (slight β-1,3-glucan) | Lasallia pustulata | 39.20 | 6.91 | 0.04 |
| Scleroglucan | β-1,6-/β-1,3-glucan | Sclerotium rolfsii | 40.42 | 6.66 | 0.84 |
| Paramylon | Linear β-1,3-glucan | Euglena gracilis | 41.88 | 6.59 | 0.00 |
| Pachyman | β-1,6-/β-1,3-glucan | Wolfiporia extensa | 40.24 | 6.82 | 0.08 |
| Laminarin | mono-β-1,6-/β-1,3-glucan | Laminaria digitata | 39.34 | 6.75 | 0.00 |
| Yeast BG | β-1,6-/β-1,3-glucan | Yeast | 42.99 | 6.93 | 1.83 |
| Barley BG | β-1,3-/β-1,4-glucan | Barley | 40.13 | 6.66 | 0.03 |
| AgCAS | β-1,3-/β-1,6-glucan, others | Agaricus brasiliensis | - | - | - |
| HKCA | β-1,3-/β-1,6-glucan, others | Candida albicans | - | - | - |
| Chitin | β-1,4-poly-N-acetyl-d-glucosamine | Crab shell | - | - | - |
| Dextran | α-1,4-/α-1,6-glucan | Leuconostoc mesenteroides | - | - | - |
| Xylan | β-1,4-xylan backbone, others | Corn core | - | - | - |
| Mannan | α-1,6-/α-1,2-, α-1,3-mannan | Saccharomyces cerevisiae | - | - | - |
| Depleted zymosan | β-1,6-/β-1,3-glucan (insoluble) | Saccharomyces cerevisiae | - | - | - |
| CSBG | β-1,6-/β-1,3-glucan (soluble) | Candida albicans | - | - | - |
| Primer/Fragment | Sequence |
|---|---|
| pColdI-n361-F | 5’-TAGGTAATCTCTGCTTAAAAGCACAG-3’ |
| NEG1-FS-R | 5’-GCCGCCGCTGCCGCCGCCGCTGCCGCCCGCCCCTGCAGCCGG-3’ |
| GS-SmBiT-SS | 5’-GGCGGCAGCGGCGGCGTGACCGGCTACCGGCTGTTCGAGGAGATTCTGTAGGTAATCTCTGCT-3’ |
| GS-SmBiT-AS | 5’-AGCAGAGATTACCTACAGAATCTCCTCGAACAGCCGGTAGCCGGTCACGCCGCCGCTGCCGCC-3’ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamanaka, D.; Kurita, S.; Hanayama, Y.; Adachi, Y. Split Enzyme-Based Biosensors for Structural Characterization of Soluble and Insoluble β-Glucans. Int. J. Mol. Sci. 2021, 22, 1576. https://doi.org/10.3390/ijms22041576
Yamanaka D, Kurita S, Hanayama Y, Adachi Y. Split Enzyme-Based Biosensors for Structural Characterization of Soluble and Insoluble β-Glucans. International Journal of Molecular Sciences. 2021; 22(4):1576. https://doi.org/10.3390/ijms22041576
Chicago/Turabian StyleYamanaka, Daisuke, Suzuka Kurita, Yuka Hanayama, and Yoshiyuki Adachi. 2021. "Split Enzyme-Based Biosensors for Structural Characterization of Soluble and Insoluble β-Glucans" International Journal of Molecular Sciences 22, no. 4: 1576. https://doi.org/10.3390/ijms22041576
APA StyleYamanaka, D., Kurita, S., Hanayama, Y., & Adachi, Y. (2021). Split Enzyme-Based Biosensors for Structural Characterization of Soluble and Insoluble β-Glucans. International Journal of Molecular Sciences, 22(4), 1576. https://doi.org/10.3390/ijms22041576