
Neurotrophins Time Point Intervention after Traumatic Brain Injury: From Zebrafish to Human


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Traumatic Brain Injury Incidence
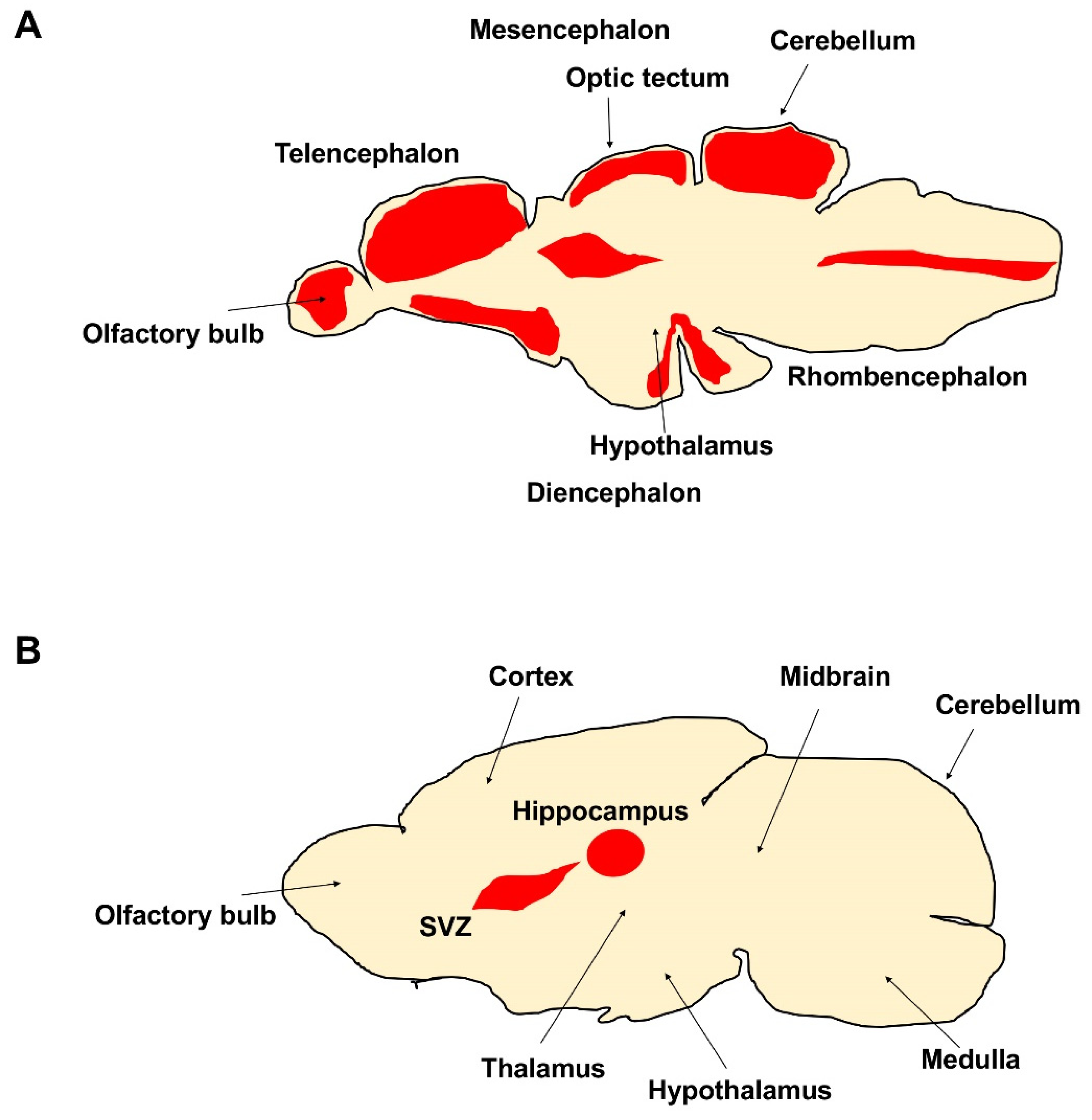
2. Diversity and Homologies in Adult Brains of Zebrafish and Mammals
3. Divergent Adult Neurogenesis in Zebrafish and Mammals
4. Neurotrophins during Vertebrate Evolution
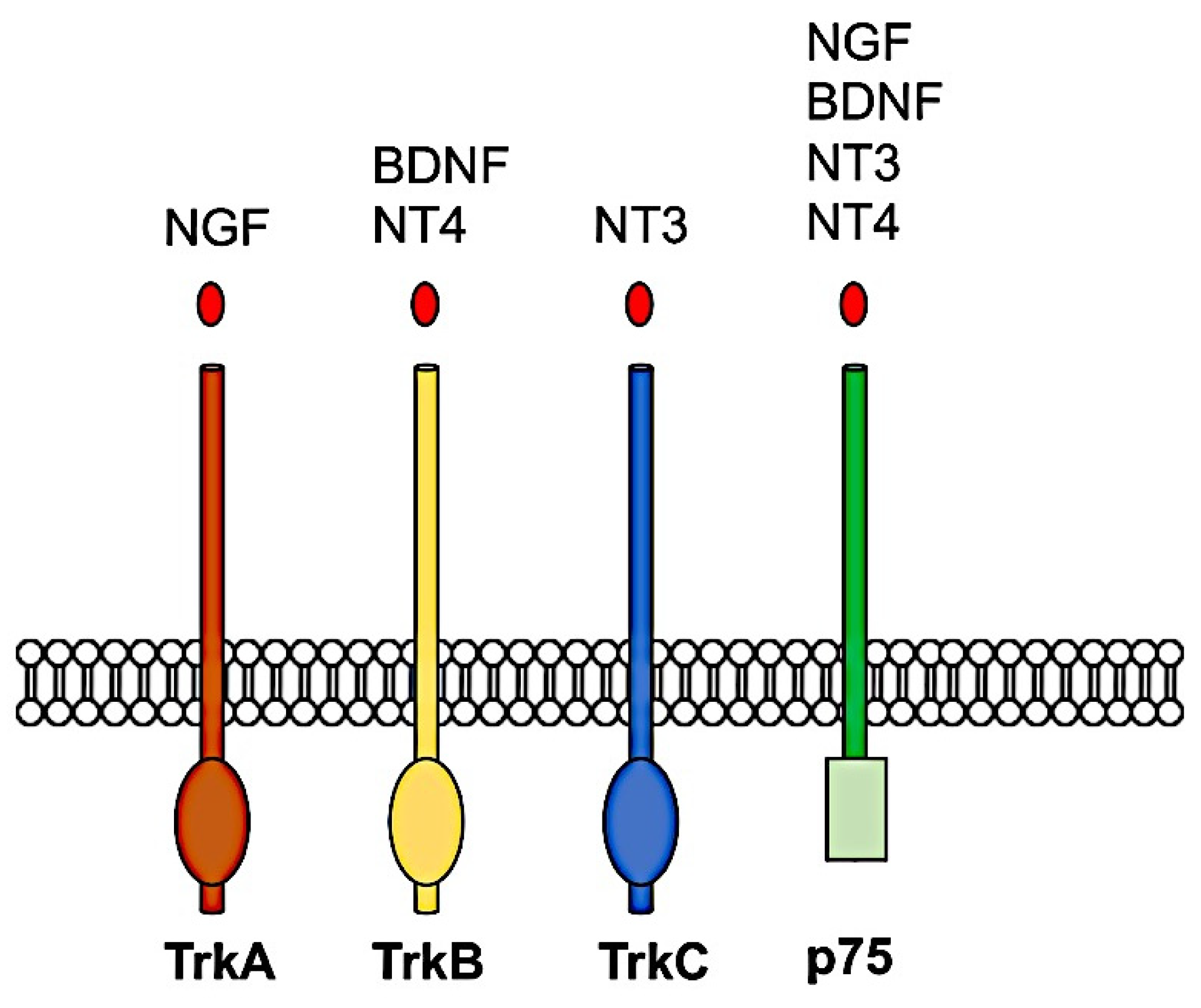
5. Multiple Neurotrophin-Receptor Interactions in the Brain of Mammals after TBI
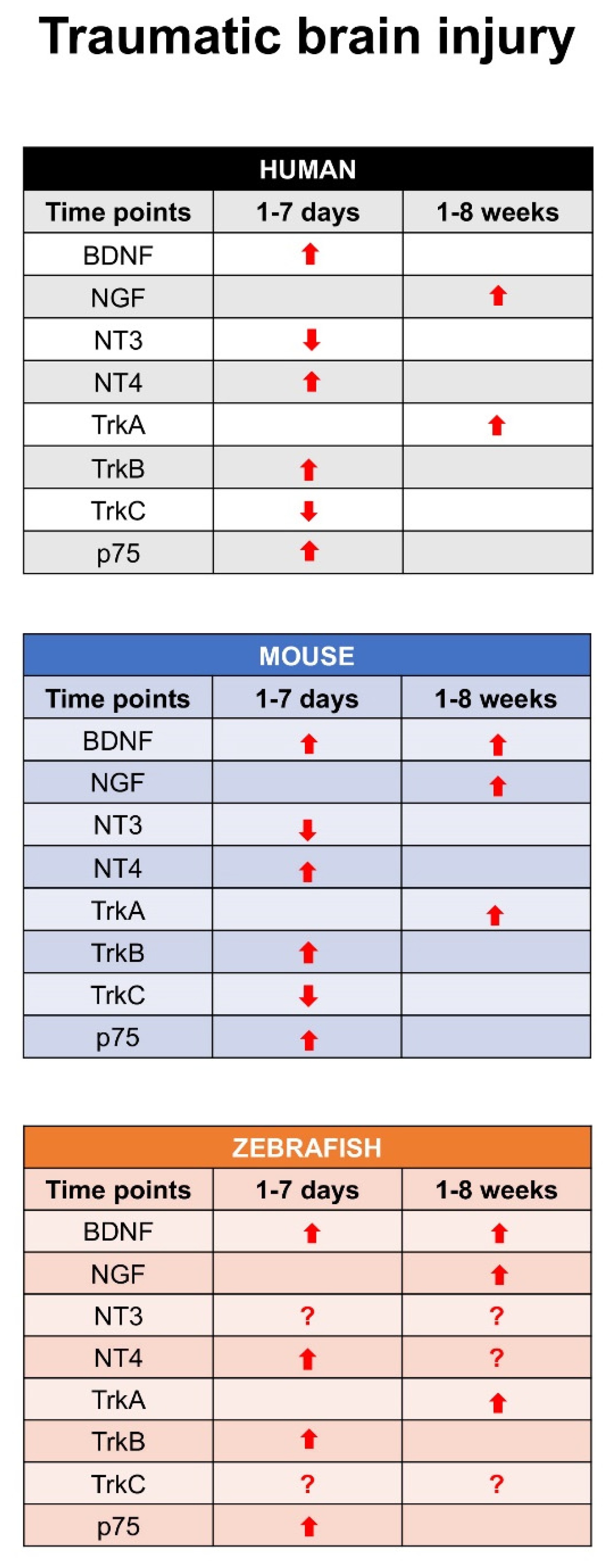
6. Neurotrophins and Their Receptors Expression after TBI in Zebrafish Models
7. The Use of Neurotrophins in Regenerative Medicine: A Promising Therapy
8. Concluding Remarks
9. Materials and Methods
9.1. Literature Search
9.2. Figures
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liao, C.C.; Chiu, W.T.; Yeh, C.C.; Chang, H.C.; Chen, T.L. Risk and outcomes for traumatic brain injury in patients with mental disorders. J. Neurol. Neurosurg. Psychiatry 2012, 83, 1186–1192. [Google Scholar] [CrossRef] [PubMed]
- Ilie, G.; Boak, A.; Adlaf, E.M.; Asbridge, M.; Cusimano, M.D. Prevalence and correlates of traumatic brain injuries among adolescents. JAMA 2013, 309, 2550–2552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cusimano, M.D.; Sharma, B.; Lawrence, D.W.; Ilie, G.; Silverberg, S.; Jones, R. Trends in North American newspaper reporting of brain injury in ice hockey. PLoS ONE 2013, 8, e61865. [Google Scholar] [CrossRef]
- Ilie, G.; Mann, R.E.; Boak, A.; Adlaf, E.M.; Hamilton, H.; Asbridge, M.; Rehm, J.; Cusimano, M.D. Suicidality, bullying and other conduct and mental health correlates of traumatic brain injury in adolescents. PLoS ONE 2014, 9, e94936. [Google Scholar] [CrossRef] [PubMed]
- Wabl, R.; Williamson, C.A.; Pandey, A.S.; Rajajee, V. Long-term and delayed functional recovery in patients with severe cerebrovascular and traumatic brain injury requiring tracheostomy. J. Neurosurg. 2018, 131, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Ilie, G.; Boak, A.; Mann, R.E.; Adlaf, E.M.; Hamilton, H.; Asbridge, M.; Rehm, J.; Cusimano, M.D. Energy Drinks, Alcohol, Sports and Traumatic Brain Injuries among Adolescents. PLoS ONE 2015, 10, e0135860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wullimann, M.F.; Mueller, T.; Distel, M.; Babaryka, A.; Grothe, B.; Koster, R.W. The long adventurous journey of rhombic lip cells in jawed vertebrates: A comparative developmental analysis. Front. Neuroanat. 2011, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Wullimann, M.F.; Mueller, T. Identification and morphogenesis of the eminentia thalami in the zebrafish. J. Comp. Neurol. 2004, 471, 37–48. [Google Scholar] [CrossRef]
- Mueller, T. What is the Thalamus in Zebrafish? Front. Neurosci. 2012, 6, 64. [Google Scholar] [CrossRef] [Green Version]
- Rink, E.; Wullimann, M.F. The teleostean (zebrafish) dopaminergic system ascending to the subpallium (striatum) is located in the basal diencephalon (posterior tuberculum). Brain Res. 2001, 889, 316–330. [Google Scholar] [CrossRef]
- Wullimann, M.F.; Rink, E. Detailed immunohistology of Pax6 protein and tyrosine hydroxylase in the early zebrafish brain suggests role of Pax6 gene in development of dopaminergic diencephalic neurons. Brain Res. Dev. Brain Res. 2001, 131, 173–191. [Google Scholar] [CrossRef]
- Portavella, M.; Vargas, J.P.; Torres, B.; Salas, C. The effects of telencephalic pallial lesions on spatial, temporal, and emotional learning in goldfish. Brain Res. Bull. 2002, 57, 397–399. [Google Scholar] [CrossRef]
- Northcutt, R.G. Connections of the lateral and medial divisions of the goldfish telencephalic pallium. J. Comp. Neurol. 2006, 494, 903–943. [Google Scholar] [CrossRef] [PubMed]
- Mueller, T.; Dong, Z.; Berberoglu, M.A.; Guo, S. The dorsal pallium in zebrafish, Danio rerio (Cyprinidae, Teleostei). Brain Res. 2011, 1381, 95–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wullimann, M.F.; Mueller, T. Teleostean and mammalian forebrains contrasted: Evidence from genes to behavior. J. Comp. Neurol. 2004, 475, 143–162. [Google Scholar] [CrossRef] [PubMed]
- Rink, E.; Wullimann, M.F. Connections of the ventral telencephalon (subpallium) in the zebrafish (Danio rerio). Brain Res. 2004, 1011, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Wullimann, M.F.; Knipp, S. Proliferation pattern changes in the zebrafish brain from embryonic through early postembryonic stages. Anat. Embryol. (Berl.) 2000, 202, 385–400. [Google Scholar] [CrossRef]
- Mueller, T. The conserved bauplan of the teleostean telencephalon. Brain Behav. Evol. 2011, 78, 259–260. [Google Scholar] [CrossRef]
- Mueller, T.; Vernier, P.; Wullimann, M.F. The adult central nervous cholinergic system of a neurogenetic model animal, the zebrafish Danio rerio. Brain Res. 2004, 1011, 156–169. [Google Scholar] [CrossRef] [PubMed]
- Ganz, J.; Kroehne, V.; Freudenreich, D.; Machate, A.; Geffarth, M.; Braasch, I.; Kaslin, J.; Brand, M. Subdivisions of the adult zebrafish pallium based on molecular marker analysis. F1000Research 2014, 3, 308. [Google Scholar] [CrossRef]
- Lindsey, B.W.; di Donato, S.; Kaslin, J.; Tropepe, V. Sensory-specific modulation of adult neurogenesis in sensory structures is associated with the type of stem cell present in the neurogenic niche of the zebrafish brain. Eur. J. Neurosci. 2014, 40, 3591–3607. [Google Scholar] [CrossRef] [PubMed]
- Goldshmit, Y.; Frisca, F.; Pinto, A.R.; Pebay, A.; Tang, J.K.; Siegel, A.L.; Kaslin, J.; Currie, P.D. Fgf2 improves functional recovery-decreasing gliosis and increasing radial glia and neural progenitor cells after spinal cord injury. Brain Behav. 2014, 4, 187–200. [Google Scholar] [CrossRef]
- Diotel, N.; Beil, T.; Strahle, U.; Rastegar, S. Differential expression of id genes and their potential regulator znf238 in zebrafish adult neural progenitor cells and neurons suggests distinct functions in adult neurogenesis. Gene Expr. Patterns 2015, 19, 1–13. [Google Scholar] [CrossRef]
- Diotel, N.; Rodriguez Viales, R.; Armant, O.; Marz, M.; Ferg, M.; Rastegar, S.; Strahle, U. Comprehensive expression map of transcription regulators in the adult zebrafish telencephalon reveals distinct neurogenic niches. J. Comp. Neurol. 2015, 523, 1202–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viales, R.R.; Diotel, N.; Ferg, M.; Armant, O.; Eich, J.; Alunni, A.; Marz, M.; Cuif, L.B.; Rastegar, S.; Strahle, U. The helix-loop-helix protein id1 controls stem cell proliferation during regenerative neurogenesis in the adult zebrafish telencephalon. Stem Cells 2015, 33, 892–903. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, E.; Diotel, N.; Capitaine, C.V.; Maria, R.P.; Gueguen, M.M.; Nasri, A.; Nicolau, J.C.; Kah, O. Steroid modulation of neurogenesis: Focus on radial glial cells in zebrafish. J. Steroid Biochem. Mol. Biol. 2016, 160, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Sundvik, M.; Panula, P. Interactions of the orexin/hypocretin neurones and the histaminergic system. Acta Physiol. (Oxf.) 2015, 213, 321–333. [Google Scholar] [CrossRef]
- Vanhanen, J.; Kinnunen, M.; Nuutinen, S.; Panula, P. Histamine H3 receptor antagonist JNJ-39220675 modulates locomotor responses but not place conditioning by dopaminergic drugs. Psychopharmacology (Berlin) 2015, 232, 1143–1153. [Google Scholar] [CrossRef]
- Semenova, S.A.; Chen, Y.C.; Zhao, X.; Rauvala, H.; Panula, P. The tyrosine hydroxylase 2 (TH2) system in zebrafish brain and stress activation of hypothalamic cells. Histochem. Cell Biol. 2014, 142, 619–633. [Google Scholar] [CrossRef] [PubMed]
- Decker, A.R.; McNeill, M.S.; Lambert, A.M.; Overton, J.D.; Chen, Y.C.; Lorca, R.A.; Johnson, N.A.; Brockerhoff, S.E.; Mohapatra, D.P.; MacArthur, H.; et al. Abnormal differentiation of dopaminergic neurons in zebrafish trpm7 mutant larvae impairs development of the motor pattern. Dev. Biol. 2014, 386, 428–439. [Google Scholar] [CrossRef] [Green Version]
- Hagio, H.; Sato, M.; Yamamoto, N. An ascending visual pathway to the dorsal telencephalon through the optic tectum and nucleus prethalamicus in the yellowfin goby Acanthogobius flavimanus (Temminck & Schlegel, 1845). J. Comp. Neurol. 2018, 526, 1733–1746. [Google Scholar]
- Kawaguchi, M.; Hagio, H.; Yamamoto, N.; Matsumoto, K.; Nakayama, K.; Akazome, Y.; Izumi, H.; Tsuneoka, Y.; Suto, F.; Murakami, Y.; et al. Atlas of the telencephalon based on cytoarchitecture, neurochemical markers, and gene expressions in Rhinogobius flumineus [Mizuno, 1960]. J. Comp. Neurol. 2019, 527, 874–900. [Google Scholar] [CrossRef]
- Kempermann, G.; Gage, F.H. Experience-dependent regulation of adult hippocampal neurogenesis: Effects of long-term stimulation and stimulus withdrawal. Hippocampus 1999, 9, 321–332. [Google Scholar] [CrossRef]
- Clelland, C.D.; Choi, M.; Romberg, C.; Clemenson, G.D., Jr.; Fragniere, A.; Tyers, P.; Jessberger, S.; Saksida, L.M.; Barker, R.A.; Gage, F.H.; et al. A functional role for adult hippocampal neurogenesis in spatial pattern separation. Science 2009, 325, 210–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Praag, H.; Schinder, A.F.; Christie, B.R.; Toni, N.; Palmer, T.D.; Gage, F.H. Functional neurogenesis in the adult hippocampus. Nature 2002, 415, 1030–1034. [Google Scholar] [CrossRef]
- van Praag, H.; Kempermann, G.; Gage, F.H. Running increases cell proliferation and neurogenesis in the adult mouse dentate gyrus. Nat. Neurosci. 1999, 2, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, V.C.; Silla, A.C.; Cazares, H.G.; Verdugo, J.M.G.; Hinojosa, A.Q. Age-related changes in astrocytic and ependymal cells of the subventricular zone. Glia 2014, 62, 790–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, V.C.; Cazares, H.G.; Bonsu, J.M.; Perez, O.G.; Achanta, P.; Wong, J.; Garcia-Verdugo, J.M.; Hinojosa, A.Q. The subventricular zone is able to respond to a demyelinating lesion after localized radiation. Stem Cells 2014, 32, 59–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Tassigny, X.d.A.; Piquer, M.S.S.; Pinedo, U.G.; Pardal, R.; Bonilla, S.; Gonzalez, V.C.; Lopez, I.L.; de la Torre Laviana, F.J.; Verdugo, J.M.G.; Lopez-Barneo, J. Resistance of subventricular neural stem cells to chronic hypoxemia despite structural disorganization of the germinal center and impairment of neuronal and oligodendrocyte survival. Hypoxia (Auckland) 2015, 3, 15–33. [Google Scholar]
- Gonzalez, V.C.; Perez, V.H.; Verdugo, J.M.G. The aged brain: Genesis and fate of residual progenitor cells in the subventricular zone. Front. Cell Neurosci. 2015, 9, 365. [Google Scholar]
- Chaichana, K.L.; Gonzalez, V.C.; Perez, O.G.; Pradilla, G.; Han, J.; Olivi, A.; Brem, H.; Verdugo, J.M.G.; Hinojosa, A.Q. Preservation of glial cytoarchitecture from ex vivo human tumor and non-tumor cerebral cortical explants: A human model to study neurological diseases. J. Neurosci. Methods 2007, 164, 261–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhang, H.; Young, A.G.; Qiu, R.; Argalian, S.; Li, X.; Wu, X.; Lemke, G.; Lu, Q. Transcriptome analysis of neural progenitor cells by a genetic dual reporter strategy. Stem Cells 2011, 29, 1589–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mignone, J.L.; Kukekov, V.; Chiang, A.S.; Steindler, D.; Enikolopov, G. Neural stem and progenitor cells in nestin-GFP transgenic mice. J. Comp. Neurol. 2004, 469, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Etienne, O.; Bery, A.; Roque, T.; Desmaze, C.; Boussin, F.D. Assessing cell cycle progression of neural stem and progenitor cells in the mouse developing brain after genotoxic stress. J. Vis. Exp. 2014, 87, 51029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupo, G.; Gioia, R.; Nisi, P.S.; Biagioni, S.; Cacci, E. Molecular Mechanisms of Neurogenic Aging in the Adult Mouse Subventricular Zone. J. Exp. Neurosci. 2019, 13, 1179069519829040. [Google Scholar] [CrossRef]
- Lupo, G.; Gaetani, S.; Cacci, E.; Biagioni, S.; Negri, R. Molecular Signatures of the Aging Brain: Finding the Links Between Genes and Phenotypes. Neurotherapeutics 2019, 16, 543–553. [Google Scholar] [CrossRef]
- Sorrells, S.F.; Paredes, M.F.; Silla, A.C.; Sandoval, K.; Qi, D.; Kelley, K.W.; James, D.; Mayer, S.; Chang, J.; Auguste, K.I.; et al. Human hippocampal neurogenesis drops sharply in children to undetectable levels in adults. Nature 2018, 555, 377–381. [Google Scholar] [CrossRef]
- Paredes, M.F.; Sorrells, S.F.; Silla, A.C.; Sandoval, K.; Qi, D.; Kelley, K.W.; James, D.; Mayer, S.; Chang, J.; Auguste, K.I.; et al. Does Adult Neurogenesis Persist in the Human Hippocampus? Cell Stem Cell 2018, 23, 780–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calati, R.; Boldrini, M.; Bensassi, I.; Maller, J.J.; Meslin, C.; Wyart, M.; Ritchie, K.; Courtet, P.; Artero, S.; Lopez-Castroman, J. Commentary on “Smaller Hippocampal Volume in Current But Not in Past Depression in Comparison to Healthy Controls: Minor Evidence From an Older Adults Sample”. J. Geriatr. Psychiatry Neurol. 2019, 32, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Boldrini, M.; Galfalvy, H.; Dwork, A.J.; Rosoklija, G.B.; Trencevska-Ivanovska, I.; Pavlovski, G.; Hen, R.; Arango, V.; Mann, J.J. Resilience Is Associated With Larger Dentate Gyrus, While Suicide Decedents With Major Depressive Disorder Have Fewer Granule Neurons. Biol. Psychiatry 2019, 85, 850–862. [Google Scholar] [CrossRef] [PubMed]
- Kizil, C.; Kaslin, J.; Kroehne, V.; Brand, M. Adult neurogenesis and brain regeneration in zebrafish. Dev. Neurobiol. 2012, 72, 429–461. [Google Scholar] [CrossRef] [PubMed]
- Grandel, H.; Kaslin, J.; Ganz, J.; Wenzel, I.; Brand, M. Neural stem cells and neurogenesis in the adult zebrafish brain: Origin, proliferation dynamics, migration and cell fate. Dev. Biol. 2006, 295, 263–277. [Google Scholar] [CrossRef] [Green Version]
- Correa, S.A.; Zupanc, G.K. Re-evaluation of the afferent connections of the pituitary in the weakly electric fish Apteronotus leptorhynchus: An in vitro tract-tracing study. J. Comp. Neurol. 2004, 470, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Zupanc, G.K.; Correa, S.A. Reciprocal neural connections between the central posterior/prepacemaker nucleus and nucleus G in the gymnotiform fish, Apteronotus leptorhynchus. Brain Behav. Evol. 2005, 65, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Zupanc, G.K.; Hinsch, K.; Gage, F.H. Proliferation, migration, neuronal differentiation, and long-term survival of new cells in the adult zebrafish brain. J. Comp. Neurol. 2005, 488, 290–319. [Google Scholar] [CrossRef]
- Kaslin, J.; Ganz, J.; Geffarth, M.; Grandel, H.; Hans, S.; Brand, M. Stem cells in the adult zebrafish cerebellum: Initiation and maintenance of a novel stem cell niche. J. Neurosci. 2009, 29, 6142–6153. [Google Scholar] [CrossRef] [Green Version]
- Hinsch, K.; Zupanc, G.K. Isolation, cultivation, and differentiation of neural stem cells from adult fish brain. J. Neurosci. Methods 2006, 158, 75–88. [Google Scholar] [CrossRef]
- Adolf, B.; Chapouton, P.; Lam, C.S.; Topp, S.; Tannhauser, B.; Strahle, U.; Gotz, M.; Cuif, L.B. Conserved and acquired features of adult neurogenesis in the zebrafish telencephalon. Dev. Biol. 2006, 295, 278–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrini, E.; Mouriec, K.; Anglade, I.; Menuet, A.; Page, Y.L.; Gueguen, M.M.; Marmignon, M.H.; Brion, F.; Pakdel, F.; Kah, O. Identification of aromatase-positive radial glial cells as progenitor cells in the ventricular layer of the forebrain in zebrafish. J. Comp. Neurol. 2007, 501, 150–167. [Google Scholar] [CrossRef]
- Chapouton, P.; Skupien, P.; Hesl, B.; Coolen, M.; Moore, J.C.; Madelaine, R.; Kremmer, E.; Kessler, T.F.; Blader, P.; Lawson, N.D.; et al. Notch activity levels control the balance between quiescence and recruitment of adult neural stem cells. J. Neurosci. 2010, 30, 7961–7974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapouton, P.; Godinho, L. Neurogenesis. Methods Cell Biol. 2010, 100, 73–126. [Google Scholar] [PubMed]
- Chapouton, P.; Adolf, B.; Leucht, C.; Tannhauser, B.; Ryu, S.; Driever, W.; Cuif, L.B. her5 expression reveals a pool of neural stem cells in the adult zebrafish midbrain. Development 2006, 133, 4293–4303. [Google Scholar] [CrossRef] [Green Version]
- Marz, M.; Chapouton, P.; Diotel, N.; Vaillant, C.; Hesl, B.; Takamiya, M.; Lam, C.S.; Kah, O.; Bally-Cuif, L.; Strahle, U. Heterogeneity in progenitor cell subtypes in the ventricular zone of the zebrafish adult telencephalon. Glia 2010, 58, 870–888. [Google Scholar] [CrossRef]
- Wehman, A.M.; Staub, W.; Meyers, J.R.; Raymond, P.A.; Baier, H. Genetic dissection of the zebrafish retinal stem-cell compartment. Dev. Biol. 2005, 281, 53–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- del Rio-Tsonis, K.; Tsonis, P.A. Eye regeneration at the molecular age. Dev. Dyn. 2003, 226, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Wetts, R.; Serbedzija, G.N.; Fraser, S.E. Cell lineage analysis reveals multipotent precursors in the ciliary margin of the frog retina. Dev. Biol. 1989, 136, 254–263. [Google Scholar] [CrossRef]
- Caroleo, M.C.; Costa, N.; Tirassa, P.; Aloe, L. Nerve growth factor produced by activated human monocytes/macrophages is severely affected by ethanol. Alcohol 2004, 34, 107–114. [Google Scholar] [CrossRef]
- Tettamanti, G.; Cattaneo, A.G.; Gornati, R.; de Eguileor, M.; Bernardini, G.; Binelli, G. Phylogenesis of brain-derived neurotrophic factor (BDNF) in vertebrates. Gene 2010, 450, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, M.; Clary, D.O.; Lefcort, F.B.; Reichardt, L.F.; Karten, H.J.; Hallbook, F. Nerve growth factor receptor TrkA is expressed by horizontal and amacrine cells during chicken retinal development. J. Comp. Neurol. 1998, 400, 408–416. [Google Scholar] [CrossRef]
- Hallbook, F.; Lundin, L.G.; Kullander, K. Lampetra fluviatilis neurotrophin homolog, descendant of a neurotrophin ancestor, discloses the early molecular evolution of neurotrophins in the vertebrate subphylum. J. Neurosci. 1998, 18, 8700–8711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibanez, C.F.; Hallbook, F.; Soderstrom, S.; Ebendal, T.; Persson, H. Biological and immunological properties of recombinant human, rat, and chicken nerve growth factors: A comparative study. J. Neurochem. 1991, 57, 1033–1041. [Google Scholar] [CrossRef]
- Hallbook, F.; Ibanez, C.F.; Persson, H. Evolutionary studies of the nerve growth factor family reveal a novel member abundantly expressed in Xenopus ovary. Neuron 1991, 6, 845–858. [Google Scholar] [CrossRef]
- Nilsson, A.S.; Fainzilber, M.; Falck, P.; Ibanez, C.F. Neurotrophin-7: A novel member of the neurotrophin family from the zebrafish. FEBS Lett. 1998, 424, 285–290. [Google Scholar] [CrossRef] [Green Version]
- Prince, V.E.; Moens, C.B.; Kimmel, C.B.; Ho, R.K. Zebrafish hox genes: Expression in the hindbrain region of wild-type and mutants of the segmentation gene, valentino. Development 1998, 125, 393–406. [Google Scholar] [PubMed]
- Vecino, E. Spatiotemporal development of the fish retina: Distribution of calbindin D-28K. Semin. Cell Dev. Biol. 1998, 9, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Vecino, E.; Caminos, E.; Becker, E.; Rudkin, B.B.; Evan, G.I.; Zanca, D.M. Increased levels of TrkA in the regenerating retinal ganglion cells of fish. NeuroReport 1998, 9, 3409–3413. [Google Scholar] [CrossRef] [PubMed]
- Vecino, E.; Caminos, E.; Ugarte, M.; Zanca, D.M.; Osborne, N.N. Immunohistochemical distribution of neurotrophins and their receptors in the rat retina and the effects of ischemia and reperfusion. Gen. Pharmacol. 1998, 30, 305–314. [Google Scholar] [CrossRef]
- Cellerino, A.; Kohler, K. Brain-derived neurotrophic factor/neurotrophin-4 receptor TrkB is localized on ganglion cells and dopaminergic amacrine cells in the vertebrate retina. J. Comp. Neurol. 1997, 386, 149–160. [Google Scholar] [CrossRef]
- Moliner, E.R.; Nauta, W.J. The isodendritic core of the brain stem. J. Comp. Neurol. 1966, 126, 311–335. [Google Scholar] [CrossRef]
- Holtzman, D.M.; Kilbridge, J.; Li, Y.; Cunningham, E.T., Jr.; Lenn, N.J.; Clary, D.O.; Reichardt, L.F.; Mobley, W.C. TrkA expression in the CNS: Evidence for the existence of several novel NGF-responsive CNS neurons. J. Neurosci. 1995, 15, 1567–1576. [Google Scholar] [CrossRef]
- Hempstead, B.L. The many faces of p75NTR. Curr. Opin. Neurobiol. 2002, 12, 260–267. [Google Scholar] [CrossRef]
- Beattie, M.S.; Harrington, A.W.; Lee, R.; Kim, J.Y.; Boyce, S.L.; Longo, F.M.; Bresnahan, J.C.; Hempstead, B.L.; Yoon, S.O. ProNGF induces p75-mediated death of oligodendrocytes following spinal cord injury. Neuron 2002, 36, 375–386. [Google Scholar] [CrossRef] [Green Version]
- Chao, M.V. Dependence receptors: What is the mechanism? Sci. STKE 2003, 2003, PE38. [Google Scholar] [CrossRef]
- Chao, M.V. Retrograde transport redux. Neuron 2003, 39, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Chao, M.V. Neurotrophins and their receptors: A convergence point for many signalling pathways. Nat. Rev. Neurosci. 2003, 4, 299–309. [Google Scholar] [CrossRef]
- Hicks, R.R.; Baldwin, S.A.; Scheff, S.W. Serum extravasation and cytoskeletal alterations following traumatic brain injury in rats. Comparison of lateral fluid percussion and cortical impact models. Mol. Chem. Neuropathol. 1997, 32, 1–16. [Google Scholar] [CrossRef]
- Hicks, R.R.; Numan, S.; Dhillon, H.S.; Prasad, M.R.; Seroogy, K.B. Alterations in BDNF and NT-3 mRNAs in rat hippocampus after experimental brain trauma. Brain Res. Mol. Brain Res. 1997, 48, 401–406. [Google Scholar] [CrossRef]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in neuronal development and function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [Green Version]
- Atwal, J.K.; Massie, B.; Miller, F.D.; Kaplan, D.R. The TrkB-Shc site signals neuronal survival and local axon growth via MEK and P13-kinase. Neuron 2000, 27, 265–277. [Google Scholar] [CrossRef]
- Kaplan, D.; Zirrgiebel, U.; Atwal, J. Center stage for NGF in peripheral (but not central) sensory neuron outgrowth. Neuron 2000, 25, 253–254. [Google Scholar] [CrossRef] [Green Version]
- Reichardt, L.F. Neurotrophin-regulated signalling pathways. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1545–1564. [Google Scholar] [CrossRef] [Green Version]
- Wardle, R.A.; Poo, M.M. Brain-derived neurotrophic factor modulation of GABAergic synapses by postsynaptic regulation of chloride transport. J. Neurosci. 2003, 23, 8722–8732. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Hua, F.; Yuan, L.; Tang, W.; Lu, J.; Yu, S.; Wang, X.; Hu, Y. Potential therapeutic effects of neurotrophins for acute and chronic neurological diseases. BioMed Res. Int. 2014, 2014, 601084. [Google Scholar] [CrossRef] [Green Version]
- Nykjaer, A.; Lee, R.; Teng, K.K.; Jansen, P.; Madsen, P.; Nielsen, M.S.; Jacobsen, C.; Kliemannel, M.; Schwarz, E.; Willnow–, T.E. Sortilin is essential for proNGF-induced neuronal cell death. Nature 2004, 427, 843–848. [Google Scholar] [CrossRef]
- Rostami, E.; Krueger, F.; Plantman, S.; Davidsson, J.; Agoston, D.; Grafman, J.; Risling, M. Alteration in BDNF and its receptors, full-length and truncated TrkB and p75(NTR) following penetrating traumatic brain injury. Brain Res. 2014, 1542, 195–205. [Google Scholar] [CrossRef]
- Bath, K.G.; Akins, M.R.; Lee, F.S. BDNF control of adult SVZ neurogenesis. Dev. Psychobiol. 2012, 54, 578–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucini, C.; D’Angelo, L.; Cacialli, P.; Palladino, A.; de Girolamo, P. BDNF, Brain, and Regeneration: Insights from Zebrafish. Int. J. Mol. Sci. 2018, 19, 3155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cacialli, P.; Palladino, A.; Lucini, C. Role of brain-derived neurotrophic factor during the regenerative response after traumatic brain injury in adult zebrafish. Neural Regen. Res. 2018, 13, 941–944. [Google Scholar]
- Uutela, M.; Lindholm, J.; Louhivuori, V.; Wei, H.; Louhivuori, L.M.; Pertovaara, A.; Akerman, K.; Castren, E.; Castren, M.L. Reduction of BDNF expression in Fmr1 knockout mice worsens cognitive deficits but improves hyperactivity and sensorimotor deficits. Genes Brain Behav. 2012, 11, 513–523. [Google Scholar] [CrossRef]
- Ito, W.; Chehab, M.; Thakur, S.; Li, J.; Morozov, A. BDNF-restricted knockout mice as an animal model for aggression. Genes Brain Behav. 2011, 10, 365–374. [Google Scholar] [CrossRef]
- Rostami, E.; Krueger, F.; Zoubak, S.; dal Monte, O.; Raymont, V.; Pardini, M.; Hodgkinson, C.A.; Goldman, D.; Risling, M.; Grafman, J. BDNF polymorphism predicts general intelligence after penetrating traumatic brain injury. PLoS ONE 2011, 6, e27389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymont, V.; Salazar, A.M.; Krueger, F.; Grafman, J. “Studying injured minds”—The Vietnam head injury study and 40 years of brain injury research. Front. Neurol. 2011, 2, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forbes, C.E.; Poore, J.C.; Barbey, A.K.; Krueger, F.; Solomon, J.; Lipsky, R.H.; Hodgkinson, C.A.; Goldman, D.; Grafman, J. BDNF polymorphism-dependent OFC and DLPFC plasticity differentially moderates implicit and explicit bias. Cereb. Cortex 2012, 22, 2602–2609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Failla, M.D.; Conley, Y.P.; Wagner, A.K. Brain-Derived Neurotrophic Factor (BDNF) in Traumatic Brain Injury-Related Mortality: Interrelationships Between Genetics and Acute Systemic and Central Nervous System BDNF Profiles. Neurorehabil. Neural Repair 2016, 30, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Failla, M.D.; Juengst, S.B.; Arenth, P.M.; Wagner, A.K. Preliminary Associations Between Brain-Derived Neurotrophic Factor, Memory Impairment, Functional Cognition, and Depressive Symptoms Following Severe TBI. Neurorehabil. Neural Repair 2016, 30, 419–430. [Google Scholar] [CrossRef] [Green Version]
- Korley, F.K.; Arrastia, R.D.; Wu, A.H.; Yue, J.K.; Manley, G.T.; Sair, H.I.; van Eyk, J.; Everett, A.D.; Investigators, T.-T.; Okonkwo, D.O.; et al. Circulating Brain-Derived Neurotrophic Factor Has Diagnostic and Prognostic Value in Traumatic Brain Injury. J. Neurotrauma 2016, 33, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Endres, M.; Meisel, A.; Biniszkiewicz, D.; Namura, S.; Prass, K.; Ruscher, K.; Lipski, A.; Jaenisch, R.; Moskowitz, M.A.; Dirnagl, U. DNA methyltransferase contributes to delayed ischemic brain injury. J. Neurosci. 2000, 20, 3175–3181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endres, M.; Fan, G.; Hirt, L.; Fujii, M.; Matsushita, K.; Liu, X.; Jaenisch, R.; Moskowitz, M.A. Ischemic brain damage in mice after selectively modifying BDNF or NT4 gene expression. J. Cereb. Blood Flow Metab. 2000, 20, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Goss, J.R.; Taffe, K.M.; Kochanek, P.M.; de Kosky, S.T. The antioxidant enzymes glutathione peroxidase and catalase increase following traumatic brain injury in the rat. Exp. Neurol. 1997, 146, 291–294. [Google Scholar] [CrossRef]
- Yang, T.; Massa, S.M.; Tran, K.C.; Simmons, D.A.; Rajadas, J.; Zeng, A.Y.; Jang, T.; Carsanaro, S.; Longo, F.M. A small molecule TrkB/TrkC neurotrophin receptor co-activator with distinctive effects on neuronal survival and process outgrowth. Neuropharmacology 2016, 110, 343–361. [Google Scholar] [CrossRef] [Green Version]
- Shetty, A.K.; Rao, M.S.; Hattiangady, B.; Zaman, V.; Shetty, G.A. Hippocampal neurotrophin levels after injury: Relationship to the age of the hippocampus at the time of injury. J. Neurosci. Res. 2004, 78, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Primiani, C.T.; Ryan, V.H.; Rao, J.S.; Cam, M.C.; Ahn, K.; Modi, H.R.; Rapoport, S.I. Coordinated gene expression of neuroinflammatory and cell signaling markers in dorsolateral prefrontal cortex during human brain development and aging. PLoS ONE 2014, 9, e110972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volosin, M.; Trotter, C.; Cragnolini, A.; Kenchappa, R.S.; Light, M.; Hempstead, B.L.; Carter, B.D.; Friedman, W.J. Induction of proneurotrophins and activation of p75NTR-mediated apoptosis via neurotrophin receptor-interacting factor in hippocampal neurons after seizures. J. Neurosci. 2008, 28, 9870–9879. [Google Scholar] [CrossRef] [PubMed]
- Cacialli, P.; Gueguen, M.M.; Coumailleau, P.; D’Angelo, L.; Kah, O.; Lucini, C.; Pellegrini, E. BDNF Expression in Larval and Adult Zebrafish Brain: Distribution and Cell Identification. PLoS ONE 2016, 11, e0158057. [Google Scholar] [CrossRef] [Green Version]
- Cacialli, P.; D’Angelo, L.; Kah, O.; Coumailleau, P.; Gueguen, M.M.; Pellegrini, E.; Lucini, C. Neuronal expression of brain derived neurotrophic factor in the injured telencephalon of adult zebrafish. J. Comp. Neurol. 2018, 526, 569–582. [Google Scholar] [CrossRef]
- Cacialli, P.; D’Angelo, L.; de Girolamo, P.; Avallone, L.; Lucini, C.; Pellegrini, E.; Castaldo, L. Morpho-Functional Features of the Gonads of Danio rerio: The Role of Brain-Derived Neurotrophic Factor. Anat. Rec. (Hoboken) 2018, 301, 140–147. [Google Scholar] [CrossRef] [Green Version]
- Abbate, F.; Guerrera, M.C.; Montalbano, G.; Levanti, M.B.; Germana, G.P.; Navarra, M.; Laura, R.; Vega, J.A.; Ciriaco, E.; Germana, A. Expression and anatomical distribution of TrkB in the encephalon of the adult zebrafish (Danio rerio). Neurosci. Lett. 2014, 563, 66–69. [Google Scholar] [CrossRef]
- Maheras, A.L.; Dix, B.; Carmo OM, S.; Young, A.E.; Gill, V.N.; Sun, J.L.; Booker, A.R.; Thomason, H.A.; Ibrahim, A.E.; Stanislaw, L.; et al. Genetic Pathways of Neuroregeneration in a Novel Mild Traumatic Brain Injury Model in Adult Zebrafish. eNeuro 2018, 5, ENEURO.0208-17.2017. [Google Scholar] [CrossRef] [PubMed]
- Cacialli, P.; Gatta, C.; D’Angelo, L.; Leggieri, A.; Palladino, A.; de Girolamo, P.; Pellegrini, E.; Lucini, C. Nerve growth factor is expressed and stored in central neurons of adult zebrafish. J. Anat. 2019, 235, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Cacialli, P.; Lucini, C. Adult neurogenesis and regeneration in zebrafish brain: Are the neurotrophins involved in? Neural. Regen. Res. 2019, 14, 2067–2068. [Google Scholar]
- Hui, S.P.; Sheng, D.Z.; Sugimoto, K.; Rajal, A.G.; Nakagawa, S.; Hesselson, D.; Kikuchi, K. Zebrafish Regulatory T Cells Mediate Organ-Specific Regenerative Programs. Dev. Cell 2017, 43, 659–672. [Google Scholar] [CrossRef] [Green Version]
- Houlton, J.; Zhou LY, Y.; Barwick, D.; Gowing, E.K.; Clarkson, A.N. Stroke Induces a BDNF-Dependent Improvement in Cognitive Flexibility in Aged Mice. Neural Plast. 2019, 2019, 1460890. [Google Scholar] [CrossRef]
- Baik, J. Neurotrophins and neuropathic pain in patients with traumatic brain injury. Korean J. Pain 2020, 33, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Cragnolini, A.B.; Montenegro, G.; Friedman, W.J.; Masco, D.H. Brain-region specific responses of astrocytes to an in vitro injury and neurotrophins. Mol. Cell Neurosci. 2018, 88, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Houlton, J.; Abumaria, N.; Hinkley SF, R.; Clarkson, A.N. Therapeutic Potential of Neurotrophins for Repair After Brain Injury: A Helping Hand From Biomaterials. Front. Neurosci. 2019, 13, 790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouel, F.; Rolland, A.S.; Devedjian, J.C.; Burnouf, T.; Devos, D. Past and Future of Neurotrophic Growth Factors Therapies in ALS: From Single Neurotrophic Growth Factor to Stem Cells and Human Platelet Lysates. Front. Neurol. 2019, 10, 835. [Google Scholar] [CrossRef]
- Jones, K.A.; Zouikr, I.; Patience, M.; Clarkson, A.N.; Isgaard, J.; Johnson, S.J.; Spratt, N.; Nilsson, M.; Walker, F.R. Chronic stress exacerbates neuronal loss associated with secondary neurodegeneration and suppresses microglial-like cells following focal motor cortex ischemia in the mouse. Brain Behav. Immun. 2015, 48, 57–67. [Google Scholar] [CrossRef]
- Hippensteel, J.A.; Anderson, B.J.; Orfila, J.E.; McMurtry, S.A.; Dietz, R.M.; Su, G.; Ford, J.A.; Oshima, K.; Yang, Y.; Zhang, F.; et al. Circulating heparan sulfate fragments mediate septic cognitive dysfunction. J. Clin. Investig. 2019, 129, 1779–1784. [Google Scholar] [CrossRef] [Green Version]
- Aurand, E.R.; Lampe, K.J.; Bjugstad, K.B. Defining and designing polymers and hydrogels for neural tissue engineering. Neurosci. Res. 2012, 72, 199–213. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cacialli, P. Neurotrophins Time Point Intervention after Traumatic Brain Injury: From Zebrafish to Human. Int. J. Mol. Sci. 2021, 22, 1585. https://doi.org/10.3390/ijms22041585
Cacialli P. Neurotrophins Time Point Intervention after Traumatic Brain Injury: From Zebrafish to Human. International Journal of Molecular Sciences. 2021; 22(4):1585. https://doi.org/10.3390/ijms22041585
Chicago/Turabian StyleCacialli, Pietro. 2021. "Neurotrophins Time Point Intervention after Traumatic Brain Injury: From Zebrafish to Human" International Journal of Molecular Sciences 22, no. 4: 1585. https://doi.org/10.3390/ijms22041585
APA StyleCacialli, P. (2021). Neurotrophins Time Point Intervention after Traumatic Brain Injury: From Zebrafish to Human. International Journal of Molecular Sciences, 22(4), 1585. https://doi.org/10.3390/ijms22041585