
Spermatozoa Transcriptional Response and Alterations in PL Proteins Properties after Exposure of Mytilus galloprovincialis to Mercury

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
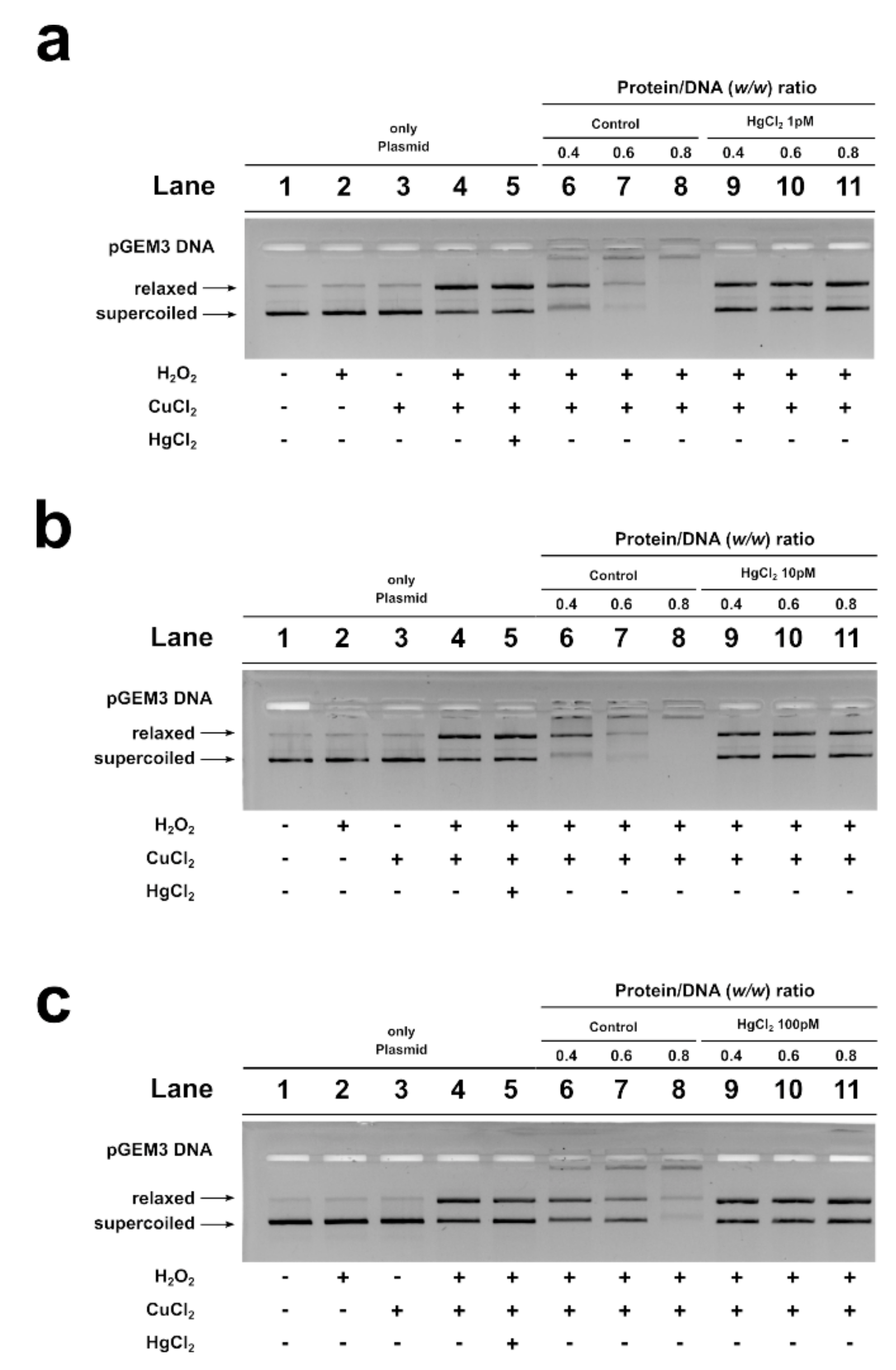
2. Results
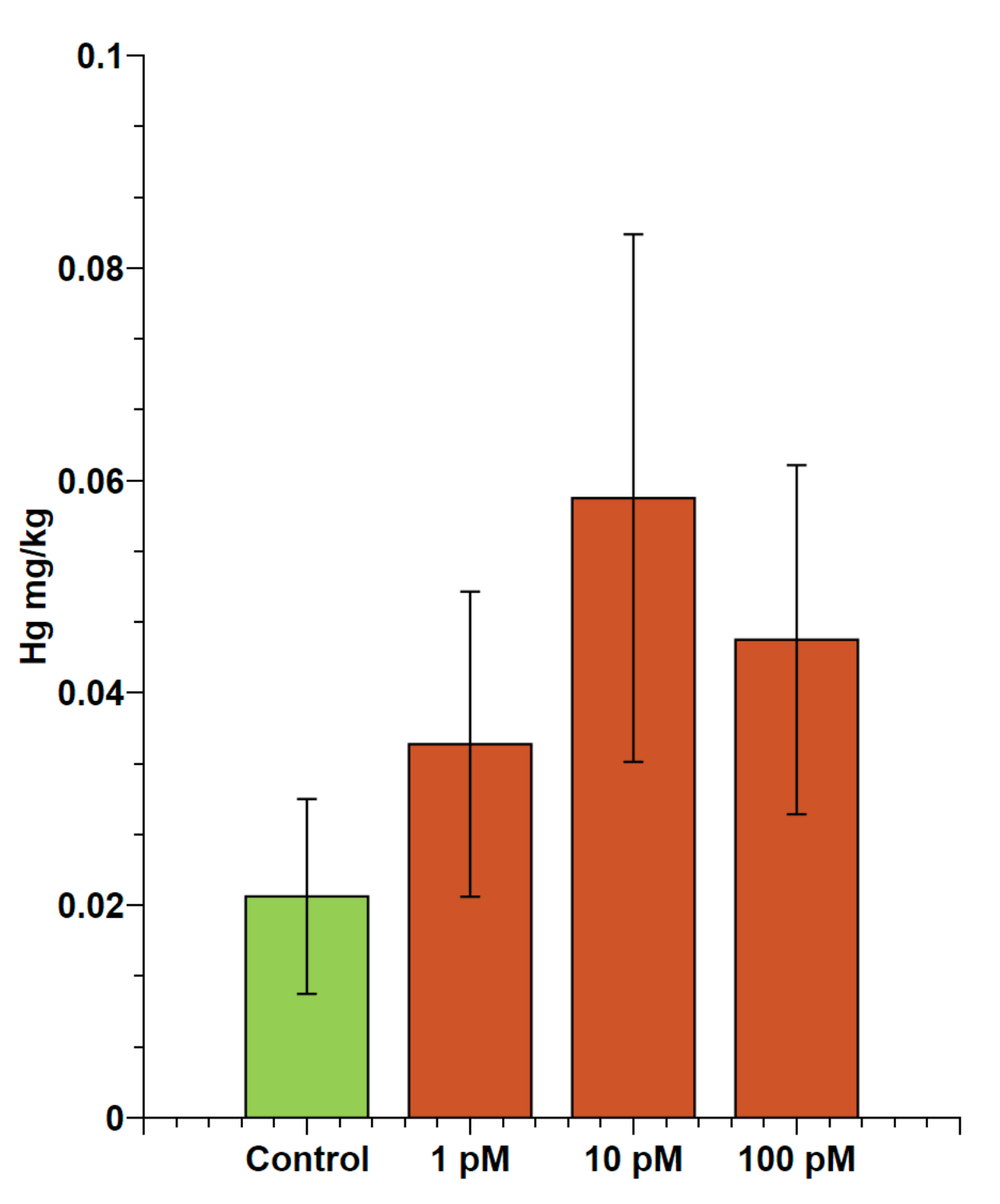
2.1. Evaluation of Accumulation of Mercury in Male Gonads
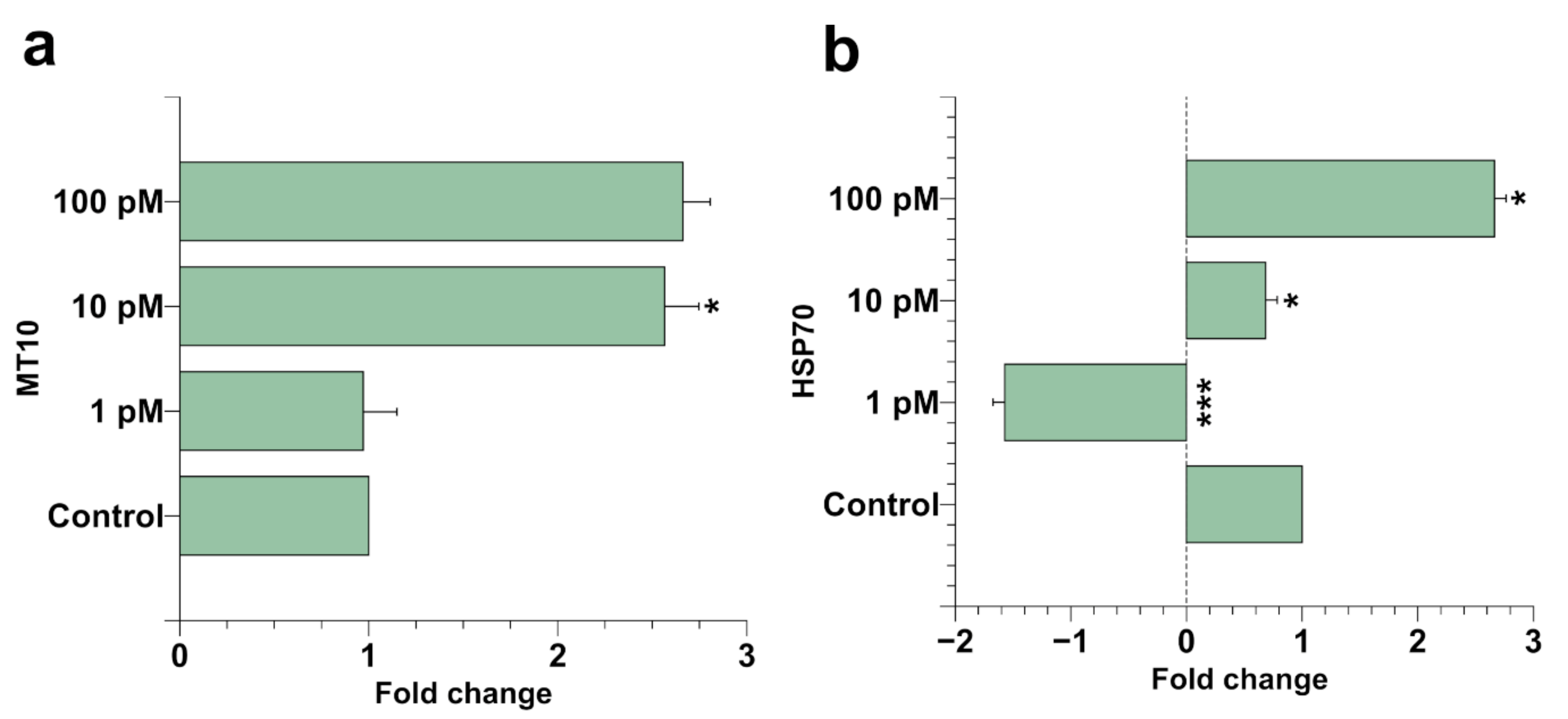
2.2. Spermatozoa Gene Expression
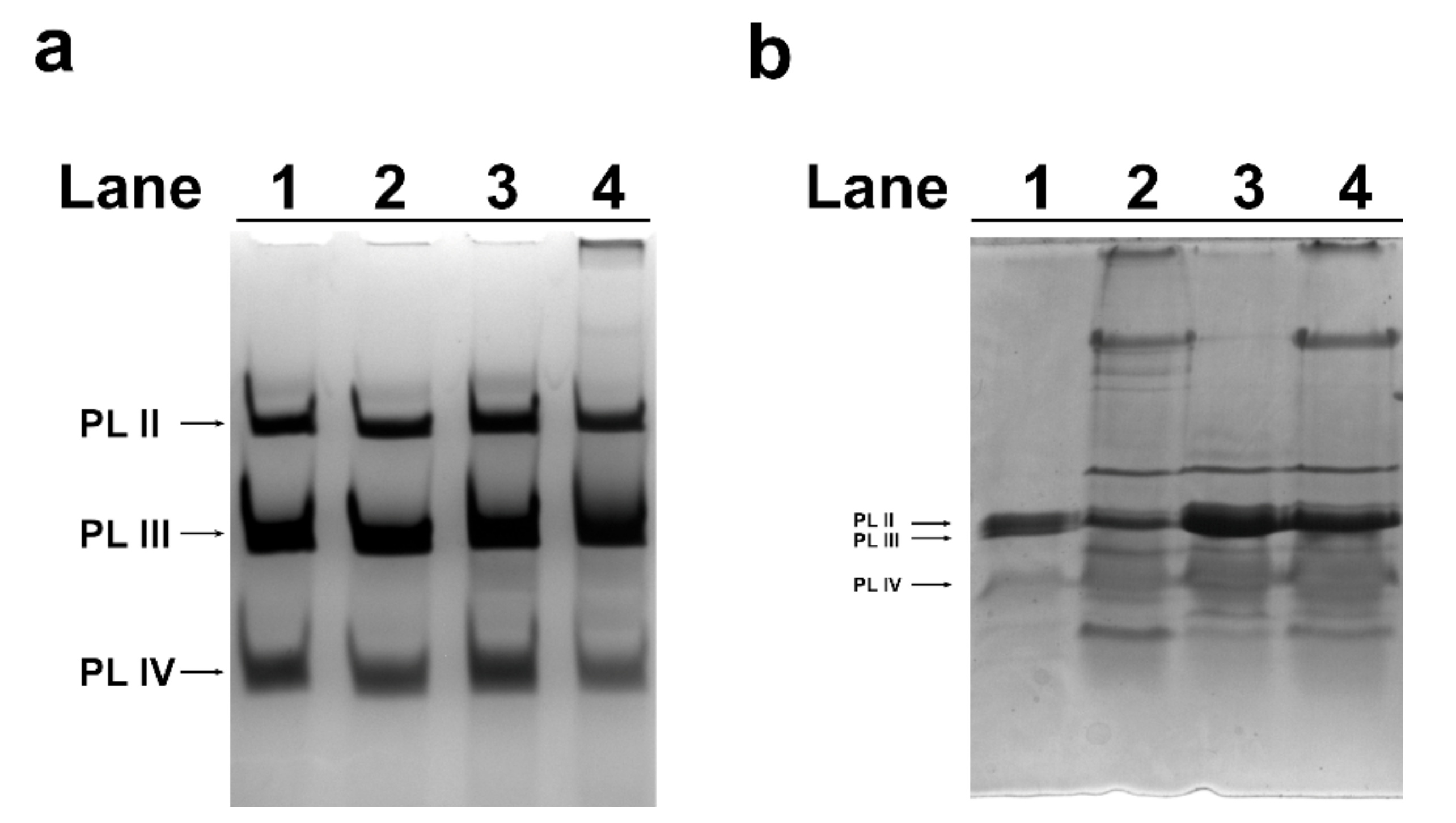
2.3. Acid-Urea (AU)-PAGE of M. galloprovincialis PL-Proteins
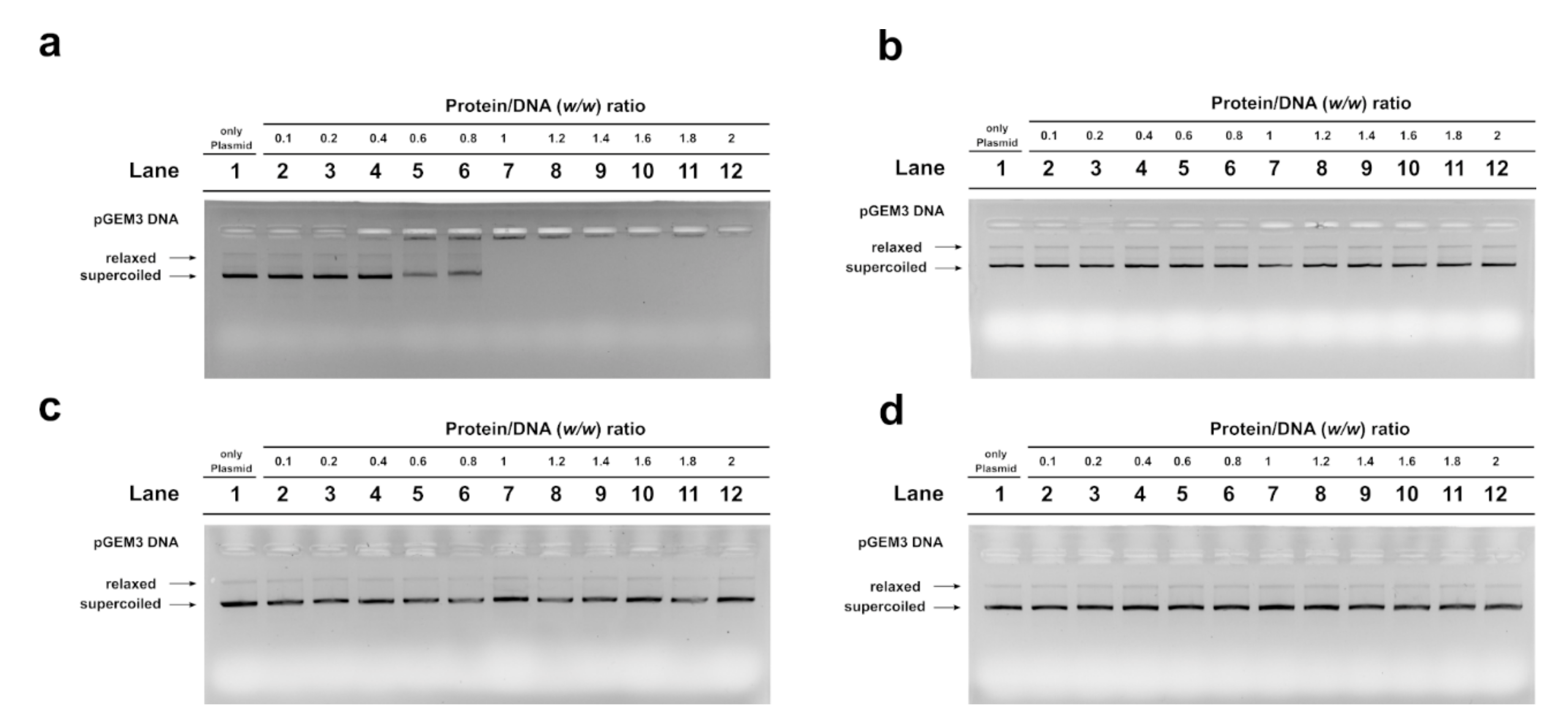
2.4. Electrophoretic Mobility Shift Assay (EMSA) Assay
2.5. DNA Protection Analysis
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Evaluation of Accumulation of Mercury in Male Mussel Gonads
4.3. Mussels Sampling and HgCl2 Exposure
4.4. Spermatozoa Sampling and Processing
4.5. RNA Extraction and RT-qPCR
4.6. PL Proteins from M. galloprovincialis Spermatozoa Extraction and Analyses
4.7. Plasmid DNA Preparation and Analysis
4.8. Analysis of the Effect of M. galloprovincialis PL Proteins on DNA Electrophoretic Mobility
4.9. DNA Protection Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ASW | Artificial sea water |
| AU-PAGE | Acetic acid-urea polyacrylamide gel electrophoresis |
| SDS-PAGE | Sodium dodecyl sulphate–polyacrylamide gel electrophoresis |
| EDTA | Ethylenediaminetetraacetic acid |
| EMSA | Electrophoretic mobility shift assay |
| ICP-MS | Inductively coupled plasma–mass spectrometry |
| PCA | Perchloric acid |
| PL | Protamine-like |
| qPCR | Quantitative polymerase chain reaction |
| RT-qPCR | Reverse transcript quantitative polymerase chain reaction |
| SNBP | Sperm nuclear basic protein |
References
- Zebral, Y.D.; da Silva Fonseca, J.; Marques, J.A.; Bianchini, A. Carbonic Anhydrase as a Biomarker of Global and Local Impacts: Insights from Calcifying Animals. Int. J. Mol. Sci. 2019, 20, 3092. [Google Scholar] [CrossRef] [Green Version]
- Depledge, M.H.; Aagaard, A.; Györkös, P. Assessment of Trace Metal Toxicity Using Molecular, Physiological and Behavioural Biomarkers. Mar. Pollut. Bull. 1995, 31, 19–27. [Google Scholar] [CrossRef]
- Piscopo, M.; Notariale, R.; Rabbito, D.; Ausió, J.; Olanrewaju, O.S.; Guerriero, G. Mytilus galloprovincialis (Lamarck, 1819) Spermatozoa: Hsp70 Expression and Protamine-like Protein Property Studies. Environ. Sci. Pollut. Res. Int. 2018, 25, 12957–12966. [Google Scholar] [CrossRef] [PubMed]
- Piscopo, M.; Trifuoggi, M.; Notariale, R.; Labar, S.; Troisi, J.; Giarra, A.; Rabbito, D.; Puoti, R.; de Benedictis, D.; Brundo, M.V.; et al. Protamine-like Proteins’ Analysis as an Emerging Biotechnique for Cadmium Impact Assessment on Male Mollusk Mytilus galloprovincialis (Lamarck 1819). Acta Biochim. Pol. 2018, 65, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Piscopo, M.; Ricciardiello, M.; Palumbo, G.; Troisi, J. Selectivity of Metal Bioaccumulation and Its Relationship with Glutathione S-Transferase Levels in Gonadal and Gill Tissues of Mytilus galloprovincialis Exposed to Ni (II), Cu (II) and Cd (II). Rend. Fis. Acc. Lincei 2016, 27, 737–748. [Google Scholar] [CrossRef] [Green Version]
- Rittschof, D.; McClellan-Green, P. Molluscs as Multidisciplinary Models in Environment Toxicology. Mar. Pollut. Bull. 2005, 50, 369–373. [Google Scholar] [CrossRef]
- Piscopo, M. Seasonal Dependence of Cadmium Molecular Effects on Mytilus galloprovincialis (Lamarck, 1819) Protamine-like Protein Properties. Mol. Reprod. Dev. 2019, 86, 1418–1429. [Google Scholar] [CrossRef]
- His, E.; Beiras, R.; Seaman, M.N.L. The Assessment of Marine Pollution—Bioassays with Bivalve Embryos and Larvae. In Advances in Marine Biology; Southward, A.J., Tyler, P.A., Young, C.M., Eds.; Academic Press: Cambridge, MA, USA, 1999; Volume 37, pp. 1–178. [Google Scholar]
- Wessel, N.; Rousseau, S.; Caisey, X.; Quiniou, F.; Akcha, F. Investigating the Relationship between Embryotoxic and Genotoxic Effects of Benzo[a]Pyrene, 17α-Ethinylestradiol and Endosulfan on Crassostrea gigas Embryos. Aquat. Toxicol. 2007, 85, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Bellas, J. Comparative Toxicity of Alternative Antifouling Biocides on Embryos and Larvae of Marine Invertebrates. Sci. Total Environ. 2006, 367, 573–585. [Google Scholar] [CrossRef]
- Connor, P.M. Acute Toxicity of Heavy Metals to Some Marine Larvae. Mar. Pollut. Bull. 1972, 3, 190–192. [Google Scholar] [CrossRef]
- Beiras, R.; His, E. Effects of Dissolved Mercury on Embryo-Genesis, Survival, Growth and Metamorphosis of Crassostrea gigas Oyster Larvae. Mar. Ecol. Prog. Ser. 1994, 113, 95–103. [Google Scholar] [CrossRef]
- Piscopo, M.; Notariale, R.; Tortora, F.; Lettieri, G.; Palumbo, G.; Manna, C. Novel Insights into Mercury Effects on Hemoglobin and Membrane Proteins in Human Erythrocytes. Molecules 2020, 25, 3278. [Google Scholar] [CrossRef] [PubMed]
- Cinnirella, S.; Bruno, D.E.; Pirrone, N.; Horvat, M.; Živković, I.; Evers, D.C.; Johnson, S.; Sunderland, E.M. Mercury Concentrations in Biota in the Mediterranean Sea, a Compilation of 40 Years of Surveys. Sci. Data 2019, 6, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mergler, D.; Anderson, H.A.; Chan, L.H.M.; Mahaffey, K.R.; Murray, M.; Sakamoto, M.; Stern, A.H. Panel on Health Risks and Toxicological Effects of Methylmercury Methylmercury Exposure and Health Effects in Humans: A Worldwide Concern. Ambio 2007, 36, 3–11. [Google Scholar] [CrossRef]
- Scheuhammer, A.M.; Meyer, M.W.; Sandheinrich, M.B.; Murray, M.W. Effects of Environmental Methylmercury on the Health of Wild Birds, Mammals, and Fish. Ambio 2007, 36, 12–18. [Google Scholar] [CrossRef]
- Beiras, R.; Albentosa, M. Inhibition of Embryo Development of the Commercial Bivalves Ruditapes decussatus and Mytilus galloprovincialis by Trace Metals; Implications for the Implementation of Seawater Quality Criteria. Aquaculture 2004, 230, 205–213. [Google Scholar] [CrossRef]
- Fathallah, S.; Medhioub, M.N.; Medhioub, A.; Kraiem, M.M. Toxicity of Hg, Cu and Zn on Early Developmental Stages of the European Clam (Ruditapes decussatus) with Potential Application in Marine Water Quality Assessment. Environ. Monit. Assess. 2010, 171, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Vergílio, C.S.; Moreira, R.V.; Carvalho, C.E.V.; Melo, E.J.T. Effects of in Vitro Exposure to Mercury on Male Gonads and Sperm Structure of the Tropical Fish Tuvira Gymnotus carapo (L.). J. Fish Dis. 2014, 37, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Lahnsteiner, F.; Mansour, N.; Berger, B. The Effect of Inorganic and Organic Pollutants on Sperm Motility of Some Freshwater Teleosts. J. Fish Biol. 2004, 65, 1283–1297. [Google Scholar] [CrossRef]
- Varotto, L.; Domeneghetti, S.; Rosani, U.; Manfrin, C.; Cajaraville, M.P.; Raccanelli, S.; Pallavicini, A.; Venier, P. DNA Damage and Transcriptional Changes in the Gills of Mytilus galloprovincialis Exposed to Nanomolar Doses of Combined Metal Salts (Cd, Cu, Hg). PLoS ONE 2013, 8, e54602. [Google Scholar] [CrossRef] [Green Version]
- Geffard, O.; His, E.; Budzinski, H.; Seaman, M.; Garrigues, P. Biological quality of seawater evaluated in situ with embryo-larval test of Crassostrea gigas and Mytilus galloprovincialis. C. R. Acad. Sci. III 2001, 324, 1149–1155. [Google Scholar] [CrossRef] [Green Version]
- Beiras, R.; His, E. Effects of Dissolved Mercury on Embryogenesis, Survival and Growth of Mytilus galloprovincialis Mussel Larvae. Mar. Ecol. Prog. Ser. 1995, 126, 185–189. [Google Scholar] [CrossRef]
- Lettieri, G.; Marra, F.; Moriello, C.; Prisco, M.; Notari, T.; Trifuoggi, M.; Giarra, A.; Bosco, L.; Montano, L.; Piscopo, M. Molecular Alterations in Spermatozoa of a Family Case Living in the Land of Fires. A First Look at Possible Transgenerational Effects of Pollutants. Int. J. Mol. Sci. 2020, 21, 6710. [Google Scholar] [CrossRef] [PubMed]
- Vassalli, Q.A.; Caccavale, F.; Avagnano, S.; Murolo, A.; Guerriero, G.; Fucci, L.; Ausió, J.; Piscopo, M. New Insights into Protamine-like Component Organization in Mytilus galloprovincialis’ Sperm Chromatin. DNA Cell Biol. 2015, 34, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Basile, A.; Loppi, S.; Piscopo, M.; Paoli, L.; Vannini, A.; Monaci, F.; Sorbo, S.; Lentini, M.; Esposito, S. The Biological Response Chain to Pollution: A Case Study from the “Italian Triangle of Death” Assessed with the Liverwort Lunularia Cruciata. Environ. Sci. Pollut. Res. Int. 2017, 24, 26185–26193. [Google Scholar] [CrossRef]
- Bonanno, G.; Orlando-Bonaca, M. Perspectives on Using Marine Species as Bioindicators of Plastic Pollution. Mar. Pollut. Bull. 2018, 137, 209–221. [Google Scholar] [CrossRef]
- Antioxidant Activity and Ultrastructural Alterations in the Biosensor Lemna minor L. Exposed in Bags in Sarno River (South Italy). Available online: https://www.iris.unina.it/handle/11588/694126?mode=full.367 (accessed on 19 December 2020).
- Maresca, V.; Fusaro, L.; Sorbo, S.; Siciliano, A.; Loppi, S.; Paoli, L.; Monaci, F.; Karam, E.A.; Piscopo, M.; Guida, M.; et al. Functional and Structural Biomarkers to Monitor Heavy Metal Pollution of One of the Most Contaminated Freshwater Sites in Southern Europe. Ecotoxicol. Environ. Saf. 2018, 163, 665–673. [Google Scholar] [CrossRef]
- Maresca, V.; Lettieri, G.; Sorbo, S.; Piscopo, M.; Basile, A. Biological Responses to Cadmium Stress in Liverwort Conocephalum conicum (Marchantiales). Int. J. Mol. Sci. 2020, 21, 6485. [Google Scholar] [CrossRef]
- Lettieri, G.; Mollo, V.; Ambrosino, A.; Caccavale, F.; Troisi, J.; Febbraio, F.; Piscopo, M. Molecular Effects of Copper on the Reproductive System of Mytilus galloprovincialis. Mol. Reprod. Dev. 2019, 86, 1357–1368. [Google Scholar] [CrossRef]
- De Guglielmo, V.; Puoti, R.; Notariale, R.; Maresca, V.; Ausió, J.; Troisi, J.; Verrillo, M.; Basile, A.; Febbraio, F.; Piscopo, M. Alterations in the Properties of Sperm Protamine-like II Protein after Exposure of Mytilus galloprovincialis (Lamarck 1819) to Sub-Toxic Doses of Cadmium. Ecotoxicol. Environ. Saf. 2019, 169, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Dellali, M.; Gnassia Barelli, M.; Romeo, M.; Aissa, P. The Use of Acetylcholinesterase Activity in Ruditapes decussatus and Mytilus galloprovincialis in the Biomonitoring of Bizerta Lagoon. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 130, 227–235. [Google Scholar] [CrossRef]
- Smaoui-Damak, W.; Hamza-Chaffai, A.; Bebianno, M.J.; Amiard, J.C. Variation of Metallothioneins in Gills of the Clam Ruditapes decussatus from the Gulf of Gabès (Tunisia). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2004, 139, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Köhler, K.; Riisgård, H.U. Formation of Metallothioneins in Relation to Accumulation of Cadmium in the Common Mussel Mytilus Edulis. Mar. Biol. 1982, 66, 53–58. [Google Scholar] [CrossRef]
- Ceratto, N.; Dondero, F.; van de Loo, J.-W.; Burlando, B.; Viarengo, A. Cloning and Sequencing of a Novel Metallothionein Gene in Mytilus galloprovincialis Lam. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2002, 131, 217–222. [Google Scholar] [CrossRef]
- Baršyt≐, D.; White, K.N.; Lovejoy, D.A. Cloning and Characterization of Metallothionein CDNAs in the Mussel Mytilus edulis L. Digestive Gland. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1999, 122, 287–296. [Google Scholar] [CrossRef]
- Dondero, F.; Piacentini, L.; Banni, M.; Rebelo, M.; Burlando, B.; Viarengo, A. Quantitative PCR Analysis of Two Molluscan Metallothionein Genes Unveils Differential Expression and Regulation. Gene 2005, 345, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Benhamed, S.; Guardiola, F.A.; Martínez, S.; Martínez-Sánchez, M.J.; Pérez-Sirvent, C.; Mars, M.; Esteban, M.A. Exposure of the Gilthead Seabream (Sparus Aurata) to Sediments Contaminated with Heavy Metals down-Regulates the Gene Expression of Stress Biomarkers. Toxicol. Rep. 2016, 3, 364–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haap, T.; Köhler, H.-R. Cadmium Tolerance in Seven Daphnia Magna Clones Is Associated with Reduced Hsp70 Baseline Levels and Induction. Aquat. Toxicol. 2009, 94, 131–137. [Google Scholar] [CrossRef]
- Singer, C.; Zimmermann, S.; Sures, B. Induction of Heat Shock Proteins (Hsp70) in the Zebra Mussel (Dreissena polymorpha) Following Exposure to Platinum Group Metals (Platinum, Palladium and Rhodium): Comparison with Lead and Cadmium Exposures. Aquat. Toxicol. 2005, 75, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Fioretti, F.M.; Febbraio, F.; Carbone, A.; Branno, M.; Carratore, V.; Fucci, L.; Ausió, J.; Piscopo, M. A Sperm Nuclear Basic Protein from the Sperm of the Marine Worm Chaetopterus Variopedatus with Sequence Similarity to the Arginine-Rich C-Termini of Chordate Protamine-Likes. DNA Cell Biol. 2012, 31, 1392–1402. [Google Scholar] [CrossRef]
- Piscopo, M.; Tomei, L.; De Petrocellis, L.; Geraci, G. Anion-Mediated Lysine-Arginine Interaction. Evidence in Chaetopterus Variopedatus Sperm Protamine. FEBS Lett. 1993, 334, 125–127. [Google Scholar] [CrossRef] [Green Version]
- Piscopo, M.; De Petrocellis, L.; Conte, M.; Pulcrano, G.; Geraci, G. On the Possibility That H1 Histone Interaction with DNA Occurs through Phosphates Connecting Lysine and Arginine Side Chain Groups. Acta Biochim. Pol. 2006, 53, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Piscopo, M.; Conte, M.; Di Paola, F.; Conforti, S.; Rana, G.; De Petrocellis, L.; Fucci, L.; Geraci, G. Relevance of Arginines in the Mode of Binding of H1 Histones to DNA. DNA Cell Biol. 2010, 29, 339–347. [Google Scholar] [CrossRef]
- Salvati, D.; Conforti, S.; Conte, M.; Matassa, D.S.; Fucci, L.; Piscopo, M. Self-Association of Chaetopterus Variopedatus Sperm Histone H1-like. Relevance of Arginine Content and Possible Physiological Role. Acta Biochim. Pol. 2008, 55, 701–706. [Google Scholar] [CrossRef]
- Lettieri, G.; D’Agostino, G.; Mele, E.; Cardito, C.; Esposito, R.; Cimmino, A.; Giarra, A.; Trifuoggi, M.; Raimondo, S.; Notari, T.; et al. Discovery of the Involvement in DNA Oxidative Damage of Human Sperm Nuclear Basic Proteins of Healthy Young Men Living in Polluted Areas. Int. J. Mol. Sci. 2020, 21, 4198. [Google Scholar] [CrossRef] [PubMed]
- BS EN 13805:2014. Foodstuffs. Determination of Trace Elements. Pressure Digestion; D09 A0E4; NSAI: Dublin, Ireland, 2014. [Google Scholar]
- Lettieri, G.; Maione, M.; Ranauda, M.A.; Mele, E.; Piscopo, M. Molecular Effects on Spermatozoa of Mytilus galloprovincialis Exposed to Hyposaline Conditions. Mol. Reprod. Dev. 2019, 86, 650–660. [Google Scholar] [CrossRef]
- Rave, N.; Crkvenjakov, R.; Boedtker, H. Identification of Procollagen MRNAs Transferred to Diazobenzyloxymethyl Paper from Formaldehyde Agarose Gels. Nucleic Acids Res. 1979, 6, 3559–3567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Notariale, R.; Basile, A.; Montana, E.; Romano, N.C.; Cacciapuoti, M.G.; Aliberti, F.; Gesuele, R.; De Ruberto, F.; Sorbo, S.; Tenore, G.C.; et al. Protamine-like Proteins Have Bactericidal Activity. The First Evidence in Mytilus galloprovincialis. Acta Biochim. Pol. 2018, 65, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Piscopo, M.; Trifuoggi, M.; Scarano, C.; Gori, C.; Giarra, A.; Febbraio, F. Relevance of Arginine Residues in Cu(II)-Induced DNA Breakage and Proteinase K Resistance of H1 Histones. Sci. Rep. 2018, 8, 7414. [Google Scholar] [CrossRef] [Green Version]
- Carbone, A.; Fioretti, F.M.; Fucci, L.; Ausió, J.; Piscopo, M. High Efficiency Method to Obtain Supercoiled DNA with a Commercial Plasmid Purification Kit. Acta Biochim. Pol. 2012, 59, 275–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | F-Primer | F-Primer Length | R-Primer | R-Primer Length | Accession Number |
|---|---|---|---|---|---|
| 18S | GCCACACGAGATTGAGCAAT | 20 | CTCGCGCTTACTGGGAATTC | 20 | L244B9 |
| hsp70 | CGCGATGCCAAACTAGACAA | 20 | TCACCTGACAAAATGGCTGC | 20 | AY861684 |
| mt10 | GCCTGCACCTTGTAACTGTAT | 21 | CTGTACACCCTGCTTCACAC | 20 | AY566248 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lettieri, G.; Notariale, R.; Ambrosino, A.; Di Bonito, A.; Giarra, A.; Trifuoggi, M.; Manna, C.; Piscopo, M. Spermatozoa Transcriptional Response and Alterations in PL Proteins Properties after Exposure of Mytilus galloprovincialis to Mercury. Int. J. Mol. Sci. 2021, 22, 1618. https://doi.org/10.3390/ijms22041618
Lettieri G, Notariale R, Ambrosino A, Di Bonito A, Giarra A, Trifuoggi M, Manna C, Piscopo M. Spermatozoa Transcriptional Response and Alterations in PL Proteins Properties after Exposure of Mytilus galloprovincialis to Mercury. International Journal of Molecular Sciences. 2021; 22(4):1618. https://doi.org/10.3390/ijms22041618
Chicago/Turabian StyleLettieri, Gennaro, Rosaria Notariale, Alessia Ambrosino, Alfredo Di Bonito, Antonella Giarra, Marco Trifuoggi, Caterina Manna, and Marina Piscopo. 2021. "Spermatozoa Transcriptional Response and Alterations in PL Proteins Properties after Exposure of Mytilus galloprovincialis to Mercury" International Journal of Molecular Sciences 22, no. 4: 1618. https://doi.org/10.3390/ijms22041618
APA StyleLettieri, G., Notariale, R., Ambrosino, A., Di Bonito, A., Giarra, A., Trifuoggi, M., Manna, C., & Piscopo, M. (2021). Spermatozoa Transcriptional Response and Alterations in PL Proteins Properties after Exposure of Mytilus galloprovincialis to Mercury. International Journal of Molecular Sciences, 22(4), 1618. https://doi.org/10.3390/ijms22041618