
Molecular Control of Oil Metabolism in the Endosperm of Seeds


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
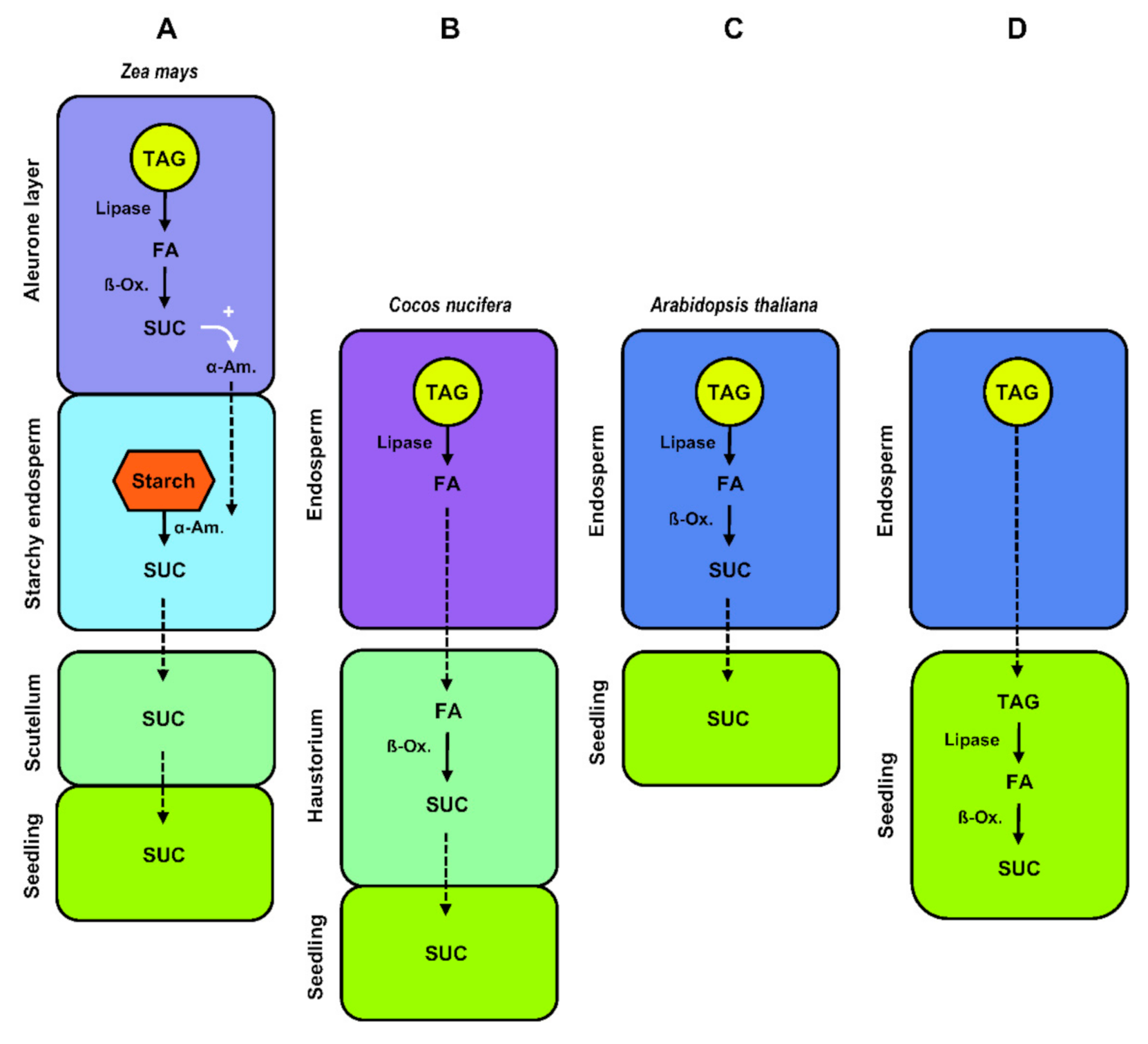
:1. Introduction: Seed Development and Oil Storage
2. Oils Stored in the Endosperm Usually Differ from That Stored in the Embryo
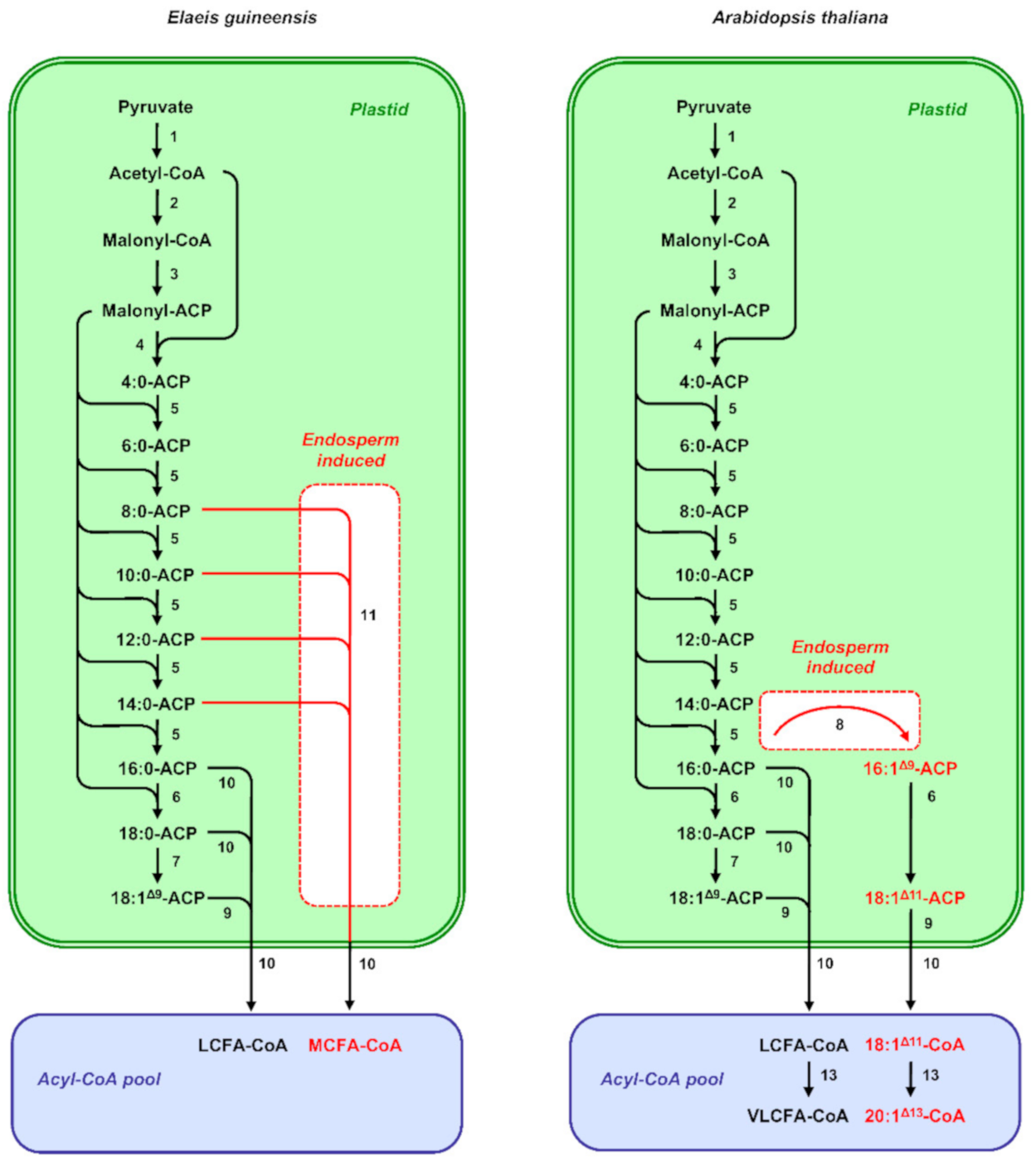
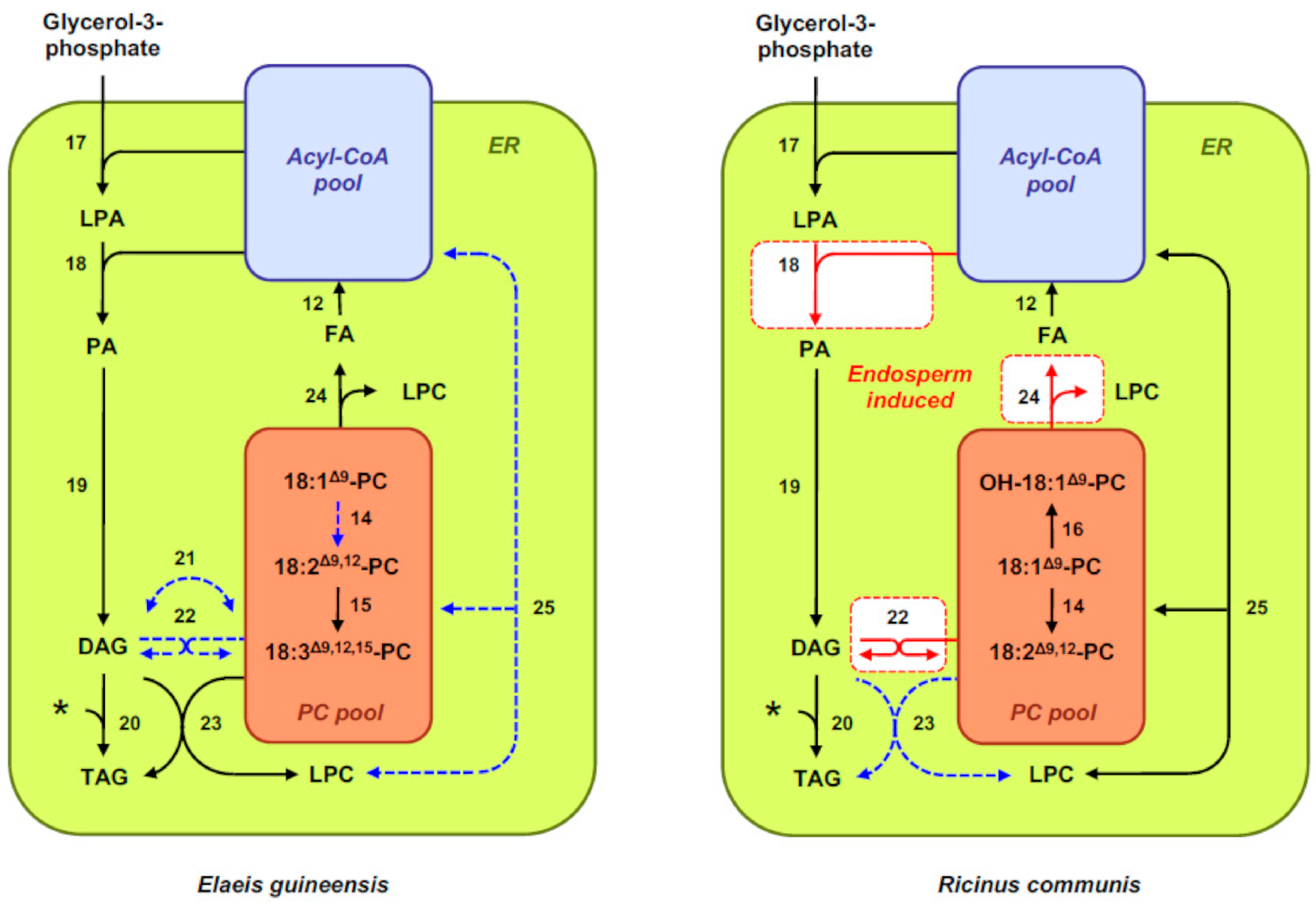
2.1. Fatty Acid and Oil Biosynthesis in Seeds
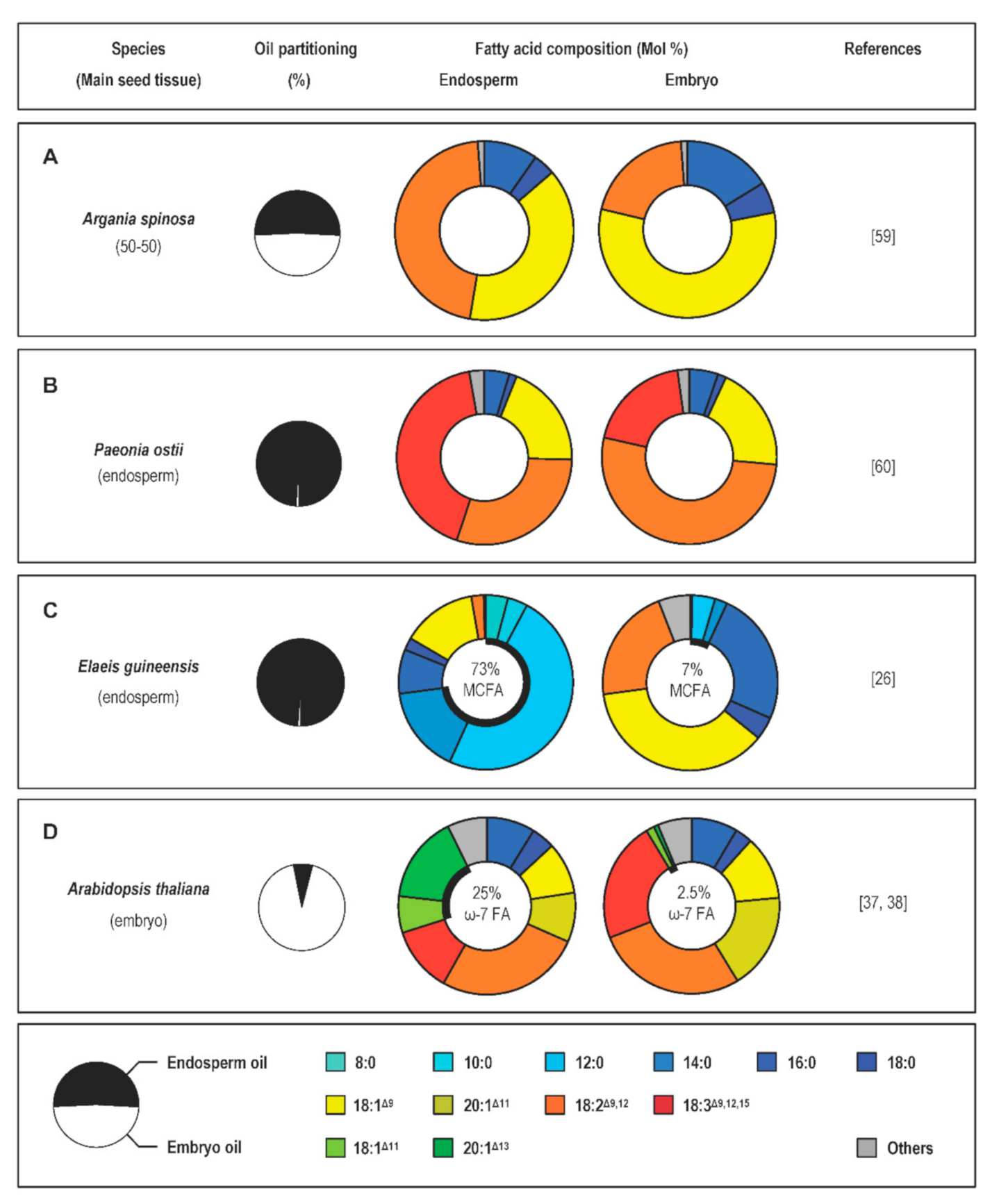
2.2. Contrasted Oil Contents in the Different Tissues of the Seed
3. Regulation of Oil Metabolism in the Endosperm of Maturing Seeds
3.1. Importance of Developmental Regulations
3.2. Transcriptional Control of Fatty-Acid Composition
4. Biological Functions of Oils Stored in the Endosperm
4.1. Promoting Seed Dispersion?
4.2. Driving Water Uptake within the Germinating Seed
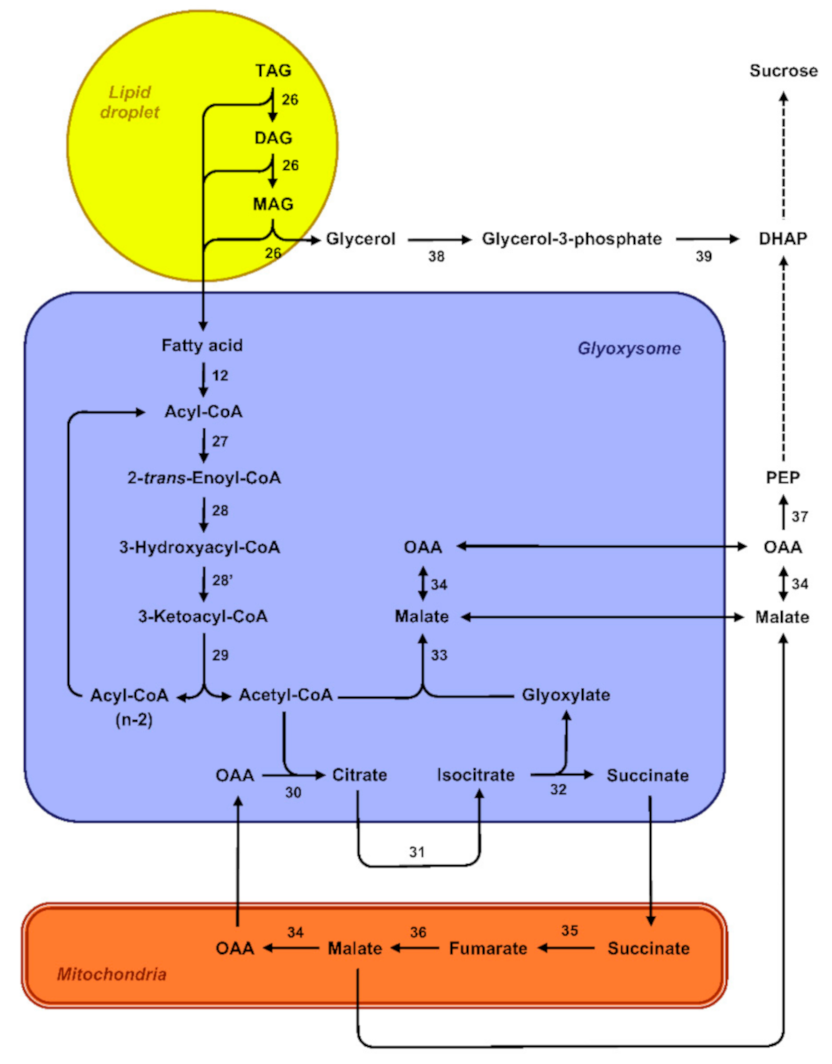
4.3. A Source of Carbon for the Seedling
4.4. Mobilization of Storage Lipids and ROS Signaling
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baud, S.; Dubreucq, B.; Miquel, M.; Rochat, C.; Lepiniec, L. Storage reserve accumulation in Arabidopsis: Metabolic and developmental control of seed filling. Arab. Book 2008, 6, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, W.E. Organismal duplication, inclusive fitness theory, and altruism: Understanding the evolution of endosperm and the angiosperm reproductive syndrome. Proc. Natl. Acad. Sci. USA 1995, 92, 3913–3917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geeta, R. The origin and maintenance of nuclear endosperm: Viewing development through a phylogenetic lens. Proc. Biol. Sci. 2003, 270, 29–35. [Google Scholar] [CrossRef]
- Ingram, G.C. Family life at close quarters: Communication and constraint in angiosperm seed development. Protoplasma 2010, 247, 195–214. [Google Scholar] [CrossRef]
- Friedman, W.E.; Floyd, S.K. Perspective: The origin of flowering plants and their reproductive biology—A tale of two phylogenies. Evolution 2001, 55, 217–231. [Google Scholar]
- Linkies, A.; Graeber, K.; Knight, C.; Leubner-Metzger, G. The evolution of seeds. New Phytol. 2010, 186, 817–831. [Google Scholar] [CrossRef]
- Williams, J.H.; Friedman, W.E. Identification of diploid endosperm in an early angiosperm lineage. Nature 2002, 415, 522–526. [Google Scholar] [CrossRef]
- Forbis, T.A.; Floyd, S.K.; de Queiroz, A. The evolution of embryo size in angiosperms and other seed plants: Implications for the evolution of seed dormancy. Evolution 2002, 56, 2112–2125. [Google Scholar] [CrossRef]
- Baroux, C.; Spillane, C.; Grossniklaus, U. Evolutionary origins of the endosperm in flowering plants. Genome Biol. 2002, 3, reviews1026. [Google Scholar] [CrossRef]
- Joët, T.; Laffargue, A.; Salmona, J.; Doulbeau, S.; Descroix, F.; Bertrand, B.; de Kochko, A.; Dussert, S. Metabolic pathways in tropical dicotyledonous albuminous seeds: Coffea Arabica as a case study. New Phytol. 2009, 182, 146–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanduri, P.; Sharma, R.; Bhat, V.; Tandon, R. Isolation, expression and evolution of fertilization independent endosperm homologs in Podostemaceae. J. Plant Res. 2016, 129, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Bethke, P.C.; Libourel, I.G.L.; Aoyama, N.; Chung, Y.-Y.; Still, D.W.; Jones, R.L. The Arabidopsis aleurone layer responds to nitric oxide, gibberellin, and abscisic acid and is sufficient and necessary for seed dormancy. Plant Physiol. 2007, 143, 1173–1188. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.A.; Herman, E.M. Characterization and functional biology of the soybean aleurone layer. BMC Plant Biol. 2018, 18, 354. [Google Scholar] [CrossRef]
- Shih, M.-D.; Lin, J.-S.; Fang, M.-J.; Tsai, Y.-C.; Hsing, Y.-I. Dye- and fluorescence-based assay to characterize symplastic and apoplastic trafficking in soybean (Glycine max L.) endosperm. Bot. Stud. 2019, 60, 24. [Google Scholar] [CrossRef]
- Liu, J.; Wu, X.; Yao, X.; Yu, R.; Larkin, P.J.; Liu, C.-M. Mutations in the DNA demethylase OsROS1 result in a thickened aleurone and improved nutritional value in rice grains. Proc. Natl. Acad. Sci. USA 2018, 115, 11327–11332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jestin, L.; Ravel, C.; Auroy, S.; Laubin, B.; Perretant, M.-R.; Pont, C.; Charmet, G. Inheritance of the number and thickness of cell layers in barley aleurone tissue (Hordeum vulgare L.): An approach using F2-F3 progeny. Theor. Appl. Genet. 2008, 116, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Berger, F. Endosperm: The crossroad of seed development. Curr. Opin. Plant Biol. 2003, 6, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Olsen, O.-A. Nuclear endosperm development in cereals and Arabidopsis thaliana. Plant Cell 2004, 16, S214–S227. [Google Scholar] [CrossRef] [Green Version]
- Gómez, E.; Royo, J.; Guo, Y.; Thompson, R.; Hueros, G. Establishment of cereal endosperm expression domains: Identification and properties of a maize transfer cell-specific transcription factor. ZmMRP Plant Cell 2002, 14, 599–610. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, Z. Differentiation mechanism and function of the cereal aleurone cells and hormone effects on them. Plant Cell Rep. 2014, 33, 1779–1787. [Google Scholar] [CrossRef]
- Young, T.E.; Gallie, D.R. Programmed cell death during endosperm development. Plant Mol. Biol. 2000, 44, 283–301. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Carbajosa, J.; Carbonero, P. Seed maturation: Developing an intrusive phase to accomplish a quiescent state. Int. J. Dev. Biol. 2005, 49, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Baud, S. Seeds as oil factories. Plant Reprod. 2018, 31, 213–235. [Google Scholar] [CrossRef]
- Baud, S.; Lepiniec, L. Physiological and developmental regulation of seed oil production. Prog. Lipid Res. 2010, 49, 235–249. [Google Scholar] [CrossRef]
- Dussert, S.; Guerin, C.; Andersson, M.; Joët, T.; Tranbarger, T.J.; Pizot, M.; Sarah, G.; Omore, A.; Durand-Gasselin, T.; Morcillo, F. Comparative transcriptome analysis of three oil palm fruit and seed tissues that differ in oil content and fatty acid composition. Plant Physiol. 2013, 162, 1337–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, K.B.; Cullerne, D.P.; El Tahchy, A.; Rolland, V.; Blanchard, C.L.; Wood, C.C.; Singh, S.P.; Petrie, J.R. Identification of genes involved in lipid biosynthesis through de novo transcriptome assembly from Cocos nucifera developing endosperm. Plant Cell Physiol. 2019, 60, 945–960. [Google Scholar] [CrossRef] [PubMed]
- Eastmond, P.J.; Jones, R.L. Hormonal regulation of gluconeogenesis in cereal aleurone is strongly cultivar-dependent and gibberellin action involves SLENDER1 but not GAMYB. Plant J. 2005, 44, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wang, Z. The cereal starch endosperm development and its relationship with other endosperm tissues and embryo. Protoplasma 2015, 252, 33–40. [Google Scholar] [CrossRef]
- Shewry, P.R.; Wan, Y.; Hawkesford, M.J.; Tosi, P. Spatial distribution of functional components in the starchy endosperm of wheat grains. J. Cereal Sci. 2020, 91, 102869. [Google Scholar] [CrossRef]
- Harwood, J.L. Recent advances in the biosynthesis of plant fatty acids. Biochim. Biophys. Acta 1996, 1301, 7–56. [Google Scholar] [CrossRef]
- Troncoso-Ponce, M.A.; Nikovics, K.; Marchive, C.; Lepiniec, L.; Baud, S. New insights on the organization and regulation of the fatty acid biosynthetic network in the model higher plant Arabidopsis thaliana. Biochimie 2016, 120, 3–8. [Google Scholar] [CrossRef]
- Shanklin, J.; Cahoon, E.B. Desaturation and related modifications of fatty acids. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 611–641. [Google Scholar] [CrossRef] [Green Version]
- Shanklin, J.; Somerville, C. Stearoyl-acyl-carrier-protein desaturase from higher plants is structurally unrelated to the animal and fungal homologs. Proc. Natl. Acad. Sci. USA 1991, 88, 2510–2514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazaz, S.; Barthole, G.; Domergue, F.; Ettaki, H.; To, A.; Vasselon, D.; de Vos, D.; Belcram, K.; Lepiniec, L.; Baud, S. Differential activation of partially redundant Δ9 stearoyl-ACP desaturase genes is critical for omega-9 monounsaturated fatty acid biosynthesis during seed development in Arabidopsis. Plant Cell 2020, 32, 3613–3637. [Google Scholar] [CrossRef] [PubMed]
- Guy, E.J.; Whittle, E.; Moche, M.; Lengqvist, J.; Lindqvist, Y.; Shanklin, J. Remote control of regioselectivity in acyl-acyl carrier protein-desaturase. Proc. Natl. Acad. Sci. USA 2011, 108, 16594–16599. [Google Scholar] [CrossRef] [Green Version]
- Bryant, F.M.; Munoz-Azcarate, O.; Kelly, A.A.; Beaudoin, F.; Kurup, S.; Eastmond, P.J. ACYL-ACYLCARRIER PROTEIN DESATURASE2 and 3 are responsible for making omega-7 fatty acids in the Arabidopsis aleurone. Plant Physiol. 2016, 172, 154–162. [Google Scholar] [CrossRef]
- Troncoso-Ponce, M.A.; Barthole, G.; Tremblais, G.; To, A.; Miquel, M.; Lepiniec, L.; Baud, S. Transcriptional activation of two delta-9 palmitoyl-ACP desaturase genes by MYB115 and MYB118 is critical for biosynthesis of omega-7 monounsaturated fatty acids in the endosperm of Arabidopsis seeds. Plant Cell 2016, 28, 2666–2682. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.; Davies, H.M.; Voelker, T.A. Palmitoyl-acyl carrier protein (ACP) thioesterase and the evolutionary origin of plant acyl-ACP thioesterases. Plant Cell 1995, 7, 359–371. [Google Scholar]
- Pollard, M.R.; Anderson, L.; Fan, C.; Hawkins, D.J.; Davies, H.M. A specific acyl-ACP thioesterase implicated in medium-chain fatty acid production in immature cotyledons of Umbellularia californica. Arch. Biochem. Biophys. 1991, 284, 306–312. [Google Scholar] [CrossRef]
- Voelker, T.; Kinney, A.J. Variations in the biosynthesis of seed-storage lipids. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 335–361. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Snapp, A.R.; Lu, C. Identification of three genes encoding microsomal oleate desaturases (FAD2) from the oilseed crop Camelina sativa. Plant Physiol. Biochem. 2011, 49, 223–229. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, C.M.; Baker, D.; Bennett, G.; Clarke, J.; Bancroft, I. Two high linolenic mutants of Arabidopsis thaliana contain megabase-scale genome duplications encompassing the FAD3 locus. Plant J. 2011, 68, 912–918. [Google Scholar] [CrossRef]
- Horn, P.J.; Liu, J.; Cocuron, J.-C.; McGlew, K.; Thrower, N.A.; Larson, M.; Lu, C.; Alonso, A.P.; Ohlrogge, J. Identification of multiple lipid genes with modifications in expression and sequence associated with the evolution of hydroxyl fatty acid accumulation in Physaria fendleri. Plant J. 2016, 86, 323–348. [Google Scholar] [CrossRef] [PubMed]
- Bates, P.D.; Durrett, T.P.; Ohlrogge, J.B.; Pollard, M. Analysis of acyl fluxes through multiple pathways of triacylglycerol synthesis in developing soybean embryos. Plant Physiol. 2009, 150, 55–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shockey, J.; Regmi, A.; Cotton, K.; Adhikari, N.; Browse, J.; Bates, P.D. Identification of Arabidopsis GPAT9 (At5g60620) as an essential gene involved in triacylglycerol biosynthesis. Plant Physiol. 2016, 170, 163–179. [Google Scholar] [CrossRef] [Green Version]
- Maisonneuve, S.; Bessoule, J.-J.; Lessire, R.; Delseny, M.; Roscoe, T.J. Expression of rapeseed microsomal lysophosphatidic acid acyltransferase isozymes enhances seed oil content in Arabidopsis. Plant Physiol. 2010, 152, 670–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eastmond, P.J.; Quettier, A.-L.; Kroon, J.T.M.; Craddock, C.; Adams, N.; Slabas, A.R. PHOSPHATIDIC ACID PHOSPHOHYDROLASE 1 and 2 regulate phospholipid synthesis at the endoplasmic reticulum in Arabidopsis. Plant Cell 2010, 22, 2796–2811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, P.D. Understanding the control of acyl flux through the lipid metabolic network of plant oil biosynthesis. Biochim. Biophys. Acta 2016, 1861, 1214–1225. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Xin, Z.; Ren, Z.; Miquel, M.; Browse, J. An enzyme regulating triacylglycerol composition is encoded by the ROD1 gene of Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 18837–18842. [Google Scholar] [CrossRef] [Green Version]
- Bates, P.D.; Stymne, S.; Ohlrogge, J. Biochemical pathways in seed oil synthesis. Curr. Opin. Plant Biol. 2013, 16, 358–364. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Shen, W.; Kazachkov, M.; Chen, G.; Chen, Q.; Carlsson, A.S.; Stymne, S.; Weselake, R.; Zou, J. Metabolic interactions between the Lands cycle and the Kennedy pathway of glycerolipid synthesis in Arabidopsis developing seeds. Plant Cell 2012, 24, 4652–4669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, A.H.C. Oil bodies and oleosins in seeds. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 177–200. [Google Scholar] [CrossRef]
- Jolivet, P.; Roux, E.; D’Andrea, S.; Davanture, M.; Negroni, L.; Zivy, M.; Chardot, T. Protein composition of oil bodies in Arabidopsis thaliana ecotype WS. Plant Physiol. Biochem. 2004, 42, 501–509. [Google Scholar] [CrossRef]
- Miquel, M.; Trigui, G.; d’Andréa, S.; Kelemen, Z.; Baud, S.; Berger, A.; Deruyffelaere, C.; Trubuil, A.; Lepiniec, L.; Dubreucq, B. Specialization of oleosins in oil body dynamics during seed development in Arabidopsis seeds. Plant Physiol. 2014, 164, 1866–1878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aitzetmüller, K.; Matthäus, B.; Friedrich, H. A new database for seed oil fatty acids—The database SOFA. Eur. J. Lipid Sci. Technol. 2003, 105, 92–103. [Google Scholar] [CrossRef]
- Matthäus, B. The database seed oil fatty acids (SOFA) is back on the internet! Eur. J. Lipid Sci. Technol. 2012, 114, 701–702. [Google Scholar] [CrossRef]
- Ohlrogge, J.; Thrower, N.; Mhaske, V.; Stymne, S.; Baxter, M.; Yang, W.; Liu, J.; Shaw, K.; Shorrosh, B.; Zhang, M.; et al. PlantFAdb: A resource for exploring hundreds of plant fatty acid structures synthesized by thousands of plants and their phylogenetic relationship. Plant J. 2018, 96, 1299–1308. [Google Scholar] [CrossRef] [Green Version]
- Errouane, K.; Doulbeau, S.; Vaissayre, V.; Leblanc, O.; Collin, M.; Kaid-Harche, M.; Dussert, S. The embryo and the endosperm contribute equally to argan seed oil yield but confer distinct lipid features to argan oil. Food Chem. 2015, 181, 270–276. [Google Scholar] [CrossRef]
- Yu, S.; Du, S.; Yuan, J.; Hu, Y. Fatty acid profile in the seeds and seed tissues of Paeonia L. species as new oil plant resources. Sci. Rep. 2016, 6, 26944. [Google Scholar] [CrossRef] [Green Version]
- López-Villalobos, A.; Dodds, P.F.; Hornung, R. Changes in fatty acid composition during development of tissues of coconut (Cocos nucifera L.) embryos in the intact nut and in vitro. J. Exp. Bot. 2001, 52, 933–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Xia, W.; Mason, A.S.; Cao, Z.; Fan, H.; Zhang, B.; Zhang, J.; Ma, Z.; Peng, M.; Huang, D. Genetic control of fatty acid composition in coconut (Cocos nucifera), African oil palm (Elaeis guineensis), and date palm (Phoenix dactylifera). Planta 2019, 249, 333–350. [Google Scholar] [CrossRef]
- Wu, Y.; Li, R.; Hildebrand, D.F. Biosynthesis and metabolic engineering of palmitoleate production, an important contributor to human health and sustainable industry. Prog. Lipid Res. 2012, 51, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Ettaki, H.; Troncoso-Ponce, M.A.; To, A.; Barthole, G.; Lepiniec, L.; Baud, S. Overexpression of MYB115, AAD2, or AAD3 in Arabidopsis thaliana seeds yields contrasting omega-7 contents. PLoS ONE 2018, 13, e0192156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penfield, S.; Rylott, E.L.; Gilday, A.D.; Graham, S.; Larson, T.R.; Graham, I.A. Reserve mobilization in the Arabidopsis endosperm fuels hypocotyl elongation in the dark, is dependent of abscisic acid, and requires PHOSPHOENOLPYRUVATE CARBOXYKINASE. Plant Cell 2004, 16, 2705–2718. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Beisson, F.; Ohlrogge, J. Oil content of Arabidopsis seeds: The influence of seed anatomy, light and plant-to-plant variation. Phytochemistry 2006, 67, 904–915. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Park, H.; Koster, K.L.; Cahoon, R.E.; Nguyen, H.T.M.; Shanklin, J.; Clemente, T.E.; Cahoon, E.B. Redirection of metabolic flux for higher levels of omega-7 monounsaturated fatty acid accumulation in camelina seeds. Plant Biotechnol. J. 2015, 13, 38–50. [Google Scholar] [CrossRef]
- Lu, S.; Sturtevant, D.; Aziz, M.; Jin, C.; Li, Q.; Chapman, K.D.; Guo, L. Spatial analysis of lipid metabolites and expressed genes reveals tissue-specific heterogeneity of lipid metabolism in high- and low-oil Brassica napus L. seeds. Plant J. 2018, 94, 915–932. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Ramos, L.C.; Tango, J.S.; Savi, A.; Leal, N.R. Variability for oil and fatty acid composition in castorbean varieties. J. Am. Oil Chem. Soc. 1984, 61, 1841–1843. [Google Scholar] [CrossRef]
- Sturtevant, D.; Romsdahl, T.B.; Yu, X.-H.; Burks, D.J.; Azad, R.K.; Shanklin, J.; Chapman, K.D. Tissue-specific differences in metabolites and transcripts contribute to the heterogeneity of ricinoleic acid accumulation in Ricinus communis L. (castor) seeds. Metabolomics 2019, 16, 6. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.P.; Kroon, J.T.M.; Swarbreck, D.; Febrer, M.; Larson, T.R.; Graham, I.A.; Caccamo, M.; Slabas, A.R. Tissue-specific whole transcriptome sequencing in castor, directed at understanding triacylglycerol lipid biosynthetic pathways. PLoS ONE 2012, 7, e30100. [Google Scholar] [CrossRef] [Green Version]
- Burgal, J.; Shockey, J.; Lu, C.; Dyer, J.; Larson, T.; Graham, I.; Browse, J. Metabolic engineering of hydroxyl fatty acid production in plants: RcDGAT2 drives dramatic increases in ricinoleate levels in seed oil. Plant Biotechnol. J. 2008, 6, 819–831. [Google Scholar] [CrossRef] [Green Version]
- Price, P.B.; Parsons, J. Distribution of lipids in embryonic axis, bran-endosperm, and hull fractions oh hulless barley and hulless oat grain. J. Agric. Food Chem. 1979, 27, 813–815. [Google Scholar] [CrossRef]
- Banaś, A.; Dębski, H.; Banaś, W.; Heneen, W.K.; Dahlqvist, A.; Bafor, M.; Gummeson, P.-O.; Marttila, S.; Ekman, Å.; Carlsson, A.S.; et al. Lipids in grain tissues of oat (Avena sativa): Differences in content, time of deposition, and fatty acid composition. J. Exp. Bot. 2007, 58, 2463–2470. [Google Scholar] [CrossRef]
- Leprince, O.; Aelst, A.C.; van Pritchard, H.W.; Murphy, D.J. Oleosins prevent oil-body coalescence during seed imbibition as suggested by a low-temperature scanning electron microscope study if dessication-tolerent and -sensitive oilseeds. Planta 1998, 204, 109–119. [Google Scholar] [CrossRef]
- Siloto, R.M.P.; Findlay, K.; López-Villalobos, A.; Yeung, E.C.; Nykiforuk, C.L.; Moloney, M.M. The accumulation of oleosins determines the size of seed oil bodies in Arabidopsis. Plant Cell 2006, 18, 1961–1974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heneen, W.K.; Karlsson, G.; Brismar, K.; Gummeson, P.-O.; Marttila, S.; Leonova, S.; Carlsson, A.S.; Bafor, M.; Banas, A.; Mattsson, B.; et al. Fusion of oil bodies in endosperm of oat grains. Planta 2008, 228, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Tschiersch, H.; Liebsch, G.; Borisjuk, L.; Stangelmayer, A.; Rolletschek, H. An imaging method for oxygen distribution, respiration and photosynthesis at a microscopic level of resolution. New Phytol. 2012, 196, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Rolletschek, H.; Weber, H.; Borisjuk, L. Energy status and its control on embryogenesis of legumes. Embryo photosynthesis contributes to oxygen supply and is coupled to biosynthetix fluxes. Plant Physiol. 2003, 132, 1196–1206. [Google Scholar] [CrossRef] [Green Version]
- Borisjuk, L.; Rolletschek, H. The oxygen status of the developing seed. New Phytol. 2009, 182, 17–30. [Google Scholar] [CrossRef]
- Borisjuk, L.; Nguyen, T.H.; Neuberger, T.; Rutten, T.; Tschiersch, H.; Claus, B.; Feussner, I.; Webb, A.G.; Jakob, P.; Weber, H.; et al. Gradients of lipid storage, photosynthesis and plastid differentiation in developing soybean seeds. New Phytol. 2005, 167, 761–776. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.K.; Ohlrogge, J.B.; Shachar-Hill, Y. The role of light in soybean seed filling metabolism. Plant J. 2009, 58, 220–234. [Google Scholar] [CrossRef] [PubMed]
- Borisjuk, L.; Neuberger, T.; Schwender, J.; Heinzel, N.; Sunderhaus, S.; Fuchs, J.; Hay, J.O.; Tschiersch, H.; Braun, H.-P.; Denolf, P.; et al. Seed architecture shapes embryo metabolism in oilseed rape. Plant Cell 2013, 25, 1625–1640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sturtevant, D.; Dueñas, M.E.; Lee, Y.-J.; Chapman, K.D. Three-dimensional visualization of membrane phospholipid distributions in Arabidopsis thaliana seeds: A spatial perspective of molecular heterogeneity. Biochim. Biophys. Acta 2017, 1862, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodfield, H.K.; Sturtevant, D.; Borisjuk, L.; Munz, E.; Gushina, I.A.; Chapman, K.; Harwood, J.L. Spatial and temporal mapping of key lipid species in Brassica napus seeds. Plant Physiol. 2017, 173, 1998–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troncoso-Ponce, M.A.; Kilaru, A.; Cao, X.; Durrett, T.P.; Fan, J.; Jensen, J.K.; Thrower, N.A.; Pauly, M.; Wilkerson, C.; Ohlrogge, J.B. Comparative deep transcriptional profiling of four developing oilseeds. Plant J. 2011, 68, 1014–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, K.; Yi, C.; Tian, D.; Sangha, J.S.; Hong, Y.; Yin, Z. Expression of fatty acid and lipid biosynthetic genes in developing endosperm of Jatropha curcas. Biotechnol. Biofuels 2012, 5, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barthole, G.; To, A.; Marchive, C.; Brunaud, V.; Soubigou-Taconnat, L.; Berger, N.; Dubreucq, B.; Lepiniec, L.; Baud, S. MYB118 represses endosperm maturation in seeds of Arabidopsis. Plant Cell 2014, 26, 3515–3537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sood, A.; Chauhan, R.S. Regulation of FA and TAG biosynthesis pathway genes in endosperms and embryos of high and low oil content genotypes of Jatropha curcas L. Plant Physiol. Biochem. 2015, 94, 253–267. [Google Scholar] [CrossRef]
- Baud, S.; Mendoza, M.S.; To, A.; Harscoët, E.; Lepiniec, L.; Dubreucq, B. WRINKLED1 specifies the regulatory action of LEAFY COTYLEDON2 towards fatty acid metabolism during seed maturation in Arabidopsis. Plant J. 2007, 50, 825–838. [Google Scholar] [CrossRef]
- Baud, S.; Lepiniec, L. Regulation of de novo fatty acid synthesis in maturing oilseeds of Arabidopsis. Plant Physiol. Biochem. 2009, 47, 448–455. [Google Scholar] [CrossRef]
- Barthole, G.; Lepiniec, L.; Rogowsky, P.M.; Baud, S. Controlling lipid accumulation in cereal grains. Plant Sci. 2012, 185-186, 33–39. [Google Scholar] [CrossRef]
- Roscoe, T.T.; Guilleminot, J.; Bessoule, J.-J.; Berger, F.; Devic, M. Complementation of seed maturation phenotypes by ectopic expression of ABSISIC ACID INSENSITIVE3, FUSCA3 and LEAFY COTYLEDON2 in Arabidopsis. Plant Cell Physiol. 2015, 56, 1215–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroj, T.; Savino, G.; Valon, C.; Giraudat, J.; Parcy, F. Regulation of storage protein gene expression in Arabidopsis. Development 2003, 130, 6065–6073. [Google Scholar] [CrossRef] [Green Version]
- Mendes, A.; Kelly, A.A.; van Erp, H.; Shaw, E.; Powers, S.J.; Kurup, S.; Eastmond, P.J. bZIP67 regulates the omega-3 fatty acid content of Arabidopsis seed oil by activating fatty acid desaturase. Plant Cell 2013, 25, 3104–3116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatihi, A.; Boulard, C.; Bouyer, D.; Baud, S.; Dubreucq, B.; Lepiniec, L. Deciphering and modifying LAFL transcriptional regulatory network in seed for improving yield and quality of storage compounds. Plant Sci. 2016, 250, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Jing, F.; Cantu, D.C.; Tvaruzkova, J.; Chipman, J.P.; Nikolau, B.J.; Yandeau-Nelson, M.D.; Reilly, P.J. Phylogenetic and experimental characterization of an acyl-ACP thioesterase family reveals significant diversity in enzymatic specificity and activity. BMC Biochem. 2011, 12, 44. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Gao, L.; Sun, R.; Yu, T.; Liang, Y.; Li, D.; Zheng, Y. Seed-specific expression of an acyl-acyl carrier protein thioesterase CnFatB3 from coconut (Cocos nucifera L.) increases the accumulation of medium-chain fatty acids in transgenic Arabidopsis seeds. Sci. Hortic. 2017, 223, 5–9. [Google Scholar] [CrossRef]
- Liang, Y.; Yuan, Y.; Liu, T.; Mao, W.; Zheng, Y.; Li, D. Identification and computational annotation of genes differentially expressed in pulp development of Cocos nucifera L. by suppression subtractive hybridization. BMC Plant Biol. 2014, 14, 205. [Google Scholar] [CrossRef] [Green Version]
- Guerin, C.; Serret, J.; Montúfar, R.; Vaissayre, V.; Bastos-Siqueira, A.; Durand-Gasselin, T.; Tregear, J.; Morcillo, F.; Dussert, S. Palm seed and fruit lipid composition: Phylogenetic and ecological perspectives. Ann. Bot. 2020, 125, 157–172. [Google Scholar] [CrossRef]
- Aymé, L.; Jolivet, P.; Nicaud, J.-M.; Chardot, T. Molecular characterization of the Elaeis guineensis medium-chain fatty acid diacylglycerol acyltransferase DGAT1-1 by heterologous expression in Yarrowia lipolytica. PLoS ONE 2015, 10, e0143113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, K.B.; Taylor, M.C.; Cullerne, D.P.; Blanchard, C.L.; Wood, C.C.; Singh, S.P.; Petrie, J.R. A reconfigured Kennedy pathway which promotes efficient accumulation of medium-chain fatty acids in leaf oils. Plant Biotechnol. J. 2017, 15, 1397–1408. [Google Scholar] [CrossRef] [PubMed]
- Milar, A.A.; Smith, M.A.; Kunst, L. All fatty acids are not equal: Discrimination in plant membrane lipids. Trends Plant Sci. 2000, 5, 95–101. [Google Scholar] [CrossRef]
- Van de Loo, F.J.; Broun, P.; Turner, S.; Somerville, C. An oleate 12-hydroxylase from Ricinus communis L. is a fatty acyl desaturase homolog. Proc. Natl. Acad. Sci. USA 1995, 92, 6743–6747. [Google Scholar] [CrossRef] [Green Version]
- Bayon, S.; Chen, G.; Weselake, R.J.; Browse, J. A small phospholipase A2-alpha from castor catalyzes the removal of hydroxyl fatty acids from phosphatidylcholine in transgenic Arabidopsis seeds. Plant Physiol. 2015, 167, 1259–1270. [Google Scholar] [CrossRef] [Green Version]
- Arrayo-Caro, J.M.; Chileh, T.; Kazachkov, M.; Zou, J.; Alonso, D.L.; Garcia-Maroto, F. The multigene family of lysophosphatidate (LPAT)-related enzymes in Ricinus communis: Cloning and molecular characterization of two LPAT genes that are expressed in castor seeds. Plant Sci. 2013, 199-200, 29–40. [Google Scholar] [CrossRef]
- Hu, Z.; Ren, Z.; Lu, C. The phosphatidylcholine diacylglycerol cholinephosphotransferase is required for efficient hydroxyl fatty acid accumulation in transgenic Arabidopsis. Plant Physiol. 2012, 158, 1944–1954. [Google Scholar] [CrossRef] [Green Version]
- Onstein, R.E.; Baker, W.J.; Couvreur, T.L.P.; Faurby, S.; Herrera-Alsina, K.; Svenning, J.-C.; Kissling, W.D. To adapt or go extinct? The fate of megafaunal palm fruits under past global change. Proc. R. Soc. B 2018, 285, 20180882. [Google Scholar] [CrossRef] [Green Version]
- Bosque, C.; Ramirez, R.; Rodriguez, D. The diet of the oilbird in Venezuela. Ornitol. Neotrop. 1995, 6, 67–80. [Google Scholar]
- Acevedo-Quintero, J.F.; Zamora-Abrego, J.G. Role of mammals on seed dispersal and predation processes of Mauritia flexuosa (Arecaceae) in the Colombian Amazon. Rev. Biol. Trop. 2016, 64, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Ross, J.H.E.; Sanchez, J.; Millan, F.; Murphy, D.J. Differential presence of oleosins in oleogenic seed and mesocarp tissues in olive (Olea europaea) and avocado (Persea americana). Plant Sci. 1993, 93, 203–210. [Google Scholar] [CrossRef]
- Gurr, M.I.; Harwood, J.L.; Frayn, K.N.; Murphy, D.J.; Michel, R.H. Lipids: Biochemistry, Biotechnology and Health, 6th ed.; Wiley-Blackwell: Oxford, UK, 2016; pp. 1–448. [Google Scholar]
- Leonova, S.; Grimberg, Å.; Marttila, S.; Stymbe, S.; Carlsson, A.S. Mobilization of lipid reserves during germination of oat (Avena sativa L.), a cereal rich in endosperm oil. J. Exp. Bot. 2010, 11, 3089–3099. [Google Scholar] [CrossRef] [Green Version]
- Orłowski, G.; Czarnecka, J. Granivory of birds and seed dispersal: Viable seeds of Amaranthus retroflexus recovered from the droppings of the grey partridge Perdix perdix. Pol. J. Ecol. 2009, 57, 191–196. [Google Scholar]
- Kurek, P.; Holeksa, J. Grains in the diets of medium-sized carnivores—A case of diplochory? Pol. J. Ecol. 2015, 63, 1–9. [Google Scholar] [CrossRef]
- Manz, B.; Müller, K.; Kucera, B.; Volke, F.; Leubner-Metzger, G. Water uptake and distribution in germinating tobacco seeds investigated in vivo by nuclear magnetic resonance imaging. Plant Physiol. 2005, 138, 1538–1551. [Google Scholar] [CrossRef] [Green Version]
- Steinbrecher, T.; Leubner-Metzger, G. The biomechanics of seed germination. J. Exp. Bot. 2017, 68, 765–783. [Google Scholar] [CrossRef] [PubMed]
- Munz, E.; Rolletschek, H.; Oeltze-Jafra, S.; Fuchs, J.; Guendel, A.; Neuberger, T.; Ortleb, S.; Jakob, P.M.; Borisjuk, L. A functional imaging study of germinating oilseed rape seed. New Phytol. 2017, 216, 1181–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paszkiewicz, G.; Gualberto, J.M.; Benamar, A.; Macherel, D.; Logan, D.C. Arabidopsis seed mitochondria are bioenergetically active immediately upon imbibition and specialize via biogenesis in preparation for autotrophic growth. Plant Cell 2017, 29, 109–128. [Google Scholar] [CrossRef] [PubMed]
- Schneider, H.; Manz, B.; Westhoff, M.; Mimietz, S.; Szimtenings, M.; Neuberger, T.; Faber, C.; Krohne, G.; Haase, A.; Volke, F.; et al. The impact of lipid distribution, composition and mobility on xylem water refilling of the resurrection plant Myrothamnus flabellifolia. New Phytol. 2003, 159, 487–505. [Google Scholar] [CrossRef] [Green Version]
- Westoby, M.; Jurado, E.; Leishman, M. Comparative evolutionary ecology of seed size. Trends Ecol. Evol. 1992, 7, 368–372. [Google Scholar] [CrossRef]
- Sanyal, A.; Lenoir, J.; O’Neill, C.; Dubois, F.; Decocq, G. Intraspecific and interspecific adaptive latitudinal cline in Brassicaceae seed oil traits. Am. J. Bot. 2018, 105, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Kornberg, H.L.; Beevers, H. A mechanism of conversion of fat to carbohydrate in castor beans. Nature 1957, 180, 35–36. [Google Scholar] [CrossRef] [PubMed]
- Penfield, S.; Graham, S.; Graham, I.A. Storage reserve mobilization in germinating oilseeds: Arabidopsis as a model system. Biochem. Soc. Trans. 2005, 33, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Masclaux-Daubresse, C.; D’Andrea, S.; Bouchez, I.; Cacas, J.-L. Reserve lipids and plant autophagy. J. Exp. Bot. 2020, 71, 2854–2861. [Google Scholar] [CrossRef]
- Han, B.; Xu, H.; Feng, Y.; Xu, W.; Cui, Q.; Liu, A. Genomic characterization and expressional profiles of autophagy-related genes (ATGs) in oilseed crop castor bean (Ricinus communis L.). Int. J. Mol. Sci. 2020, 21, 562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deruyffelaere, C.; Bouchez, I.; Morin, H.; Guillot, A.; Miquel, M.; Froissard, M.; Chardot, T.; D’Andréa, S. Ubiquitin-mediated proteasomal degradation of oleosins is involved in oil body mobilization during post-germinative seedling growth in Arabidopsis. Plant Cell Physiol. 2015, 56, 1374–1387. [Google Scholar] [CrossRef] [Green Version]
- Eastmond, P.J. SUGAR-DEPENDENT1 encodes a patatin domain triacylglycerol lipase that initiates storage oil breakdown in germinating Arabidopsis seeds. Plant Cell 2006, 18, 665–675. [Google Scholar] [CrossRef] [Green Version]
- Graham, I.A. Seed storage oil mobilization. Annu. Rev. Plant Biol. 2008, 59, 115–142. [Google Scholar] [CrossRef]
- Beevers, H. Microbodies in higher plants. Annu. Rev. Plant Physiol. 1979, 30, 159–193. [Google Scholar] [CrossRef]
- Baker, A.; Graham, I.A.; Holdsworth, M.; Smith, S.M.; Theodoulou, F.L. Chewing the fat: β-oxidation in signaling and development. Trends Plant Sci. 2006, 11, 124–132. [Google Scholar] [CrossRef]
- De Bellis, L.; Luvisi, A.; Alpi, A. Aconitase: to be or not to be inside plant glyoxysomes, that is the question. Biology 2020, 9, 162. [Google Scholar] [CrossRef] [PubMed]
- Hutton, D.; Stumpf, P.K. Characterization of the ß-oxidation systems from maturing and germinating castor bean seeds. Plant Physiol. 1969, 44, 508–516. [Google Scholar] [CrossRef] [Green Version]
- Huang, A.H.C.; Bevers, H. Developmental changes in endosperm of germinating castor bean independent of embryonic axis. Plant Physiol. 1974, 54, 277–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, M.; Beevers, H. Subcellular distribution of gluconeogenetic enzymes in germinating castor bean endosperm. Plant Physiol. 1979, 64, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Penfield, S.; Li, Y.; Gilday, A.D.; Graham, S.; Graham, I.A. Arabidopsis ABA INSENSITIVE4 regulates lipid mobilization in the embryo and reveals repression of seed germination by the endosperm. Plant Cell 2006, 18, 1887–1899. [Google Scholar] [CrossRef] [Green Version]
- Eklund, D.M.; Edqvist, J. Localization of nonspecific lipid transfer proteins correlate with programmed cell death responses during endosperm degradation in Euphorbia lagascae seedlings. Plant Physiol. 2003, 132, 1249–1259. [Google Scholar] [CrossRef] [Green Version]
- Kader, J.C. Lipid-transfer proteins in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 627–654. [Google Scholar] [CrossRef] [Green Version]
- Salminen, T.A.; Blomqvist, K.; Edqvist, J. Lipid transfer proteins: classification, nomenclature, structure, and function. Planta 2016, 244, 971–997. [Google Scholar] [CrossRef] [Green Version]
- Edqvist, J.; Farbos, I. Characterization of germination-specific lipid transfer proteins from Euphorbia lagascae. Planta 2002, 215, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Mazzottini-dos-Santos, H.C.; Ribeiro, L.M.; Oliveira, D.M.T. Roles of the haustorium and endosperm during the development of seedlings of Acrocomia aculeata (Arecaceae): dynamics of reserve mobilization and accumulation. Protoplasma 2017, 254, 1563–1578. [Google Scholar] [CrossRef]
- DeMason, D.A. Growth parameters in the cotyledon of date seedlings. Bot. Gaz. 1984, 145, 176–183. [Google Scholar] [CrossRef]
- Sugimuma, Y.; Murakami, T. Structure and functions of the haustorium in germinating coconut palm seed. Jpn. Agric. Res. Q. 1990, 24, 1–14. [Google Scholar]
- Dias, D.S.; Ribeiro, L.M.; Lopes, P.S.N.; Melo, G.A.; Müller, M.; Munné-Bosch, S. Haustorium-endosperm relationships and the integration between developmental pathways during reserve mobilization in Butia capitata (Arecaceae) seeds. Ann. Bot. 2018, 122, 267–277. [Google Scholar] [CrossRef] [PubMed]
- DeMason, D.A. Histochemical and ultrastuctural changes in the haustorium of date (Phoenix dactylifera L.). Protoplasma 1985, 126, 168–177. [Google Scholar] [CrossRef]
- Oo, K.C.; Stumpf, P.K. Some enzymatic activities in the germinating oil palm (Elaeis guineensis) seedling. Plant Physiol. 1983, 73, 1028–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alang, Z.C.; Moir, G.F.J.; Jones, L.H. Composition, degradation and utilization of endosperm during germination in the oil palm (Elaeis guineensis Jacq.). Ann. Bot. 1988, 61, 261–268. [Google Scholar] [CrossRef]
- Cui, J.; Lamade, E.; Tcherkez, G. Seed germination in oil palm (Elaeis guineensis Jacq.): A review of metabolic pathways and control mechanisms. Int. J. Mol. Sci. 2020, 21, 4227. [Google Scholar] [CrossRef]
- Oo, K.C.; Stumpf, P.K. The metabolism of the germinating oil palm (Elaeis guineensis) seedling. Plant Physiol. 1983, 73, 1033–1037. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, N.C.C.; Lopes, P.S.N.; Ribeiro, L.M.; Mercadante-Simões, M.O.; Oliveira, L.A.A.; Silvério, F.O. Seed structure, germination, and reserve mobilization in Butia capitata (Arecaceae). Trees 2013, 27, 1633–1645. [Google Scholar] [CrossRef]
- Boatman, S.G.; Crombie, M. Fat metabolism in the West African palm oil (Elaeis guineensis). J. Exp. Bot. 1958, 9, 52–74. [Google Scholar] [CrossRef]
- Clarke, N.A.; Wilkinson, M.C.; Laidman, D.L. Lipid metabolism in germinating cereals. In Lipids in Cereal Technology; Barnes, P.J., Ed.; Academic Press: Cambridge, MA, USA, 1983; pp. 57–92. [Google Scholar]
- Doig, R.I.; Colborne, A.J.; Morris, G.; Laidman, D.L. The induction of glyoxysomal enzyme activities in the aleurone cells of germinating wheat. J. Exp. Bot. 1975, 26, 387–398. [Google Scholar] [CrossRef]
- Holtman, W.L.; Heistek, J.C.; Mattern, K.A.; Bakhuizen, R.; Douma, A.C. Beta-oxidation of fatty acids is linked to the glyoxylate cycle in the aleurone but not in the embryo of germinating barley. Plant Sci. 1994, 99, 43–53. [Google Scholar] [CrossRef]
- Jensen, S.A.; Heltved, F. Visualization of enzyme activity in germinating cereal seeds using a lipase sensitive fluorochrome. Carlsberg Res. Commun. 1982, 47, 297–303. [Google Scholar] [CrossRef] [Green Version]
- Ekstrand, B.; Gangby, I.; Åkesson, G. Lipase activity in oats-distribution, pH dependence, and heat inactivation. Chemistry 1992, 69, 379–381. [Google Scholar]
- Aoki, N.; Scofield, G.N.; Wang, X.-D.; Offler, C.E.; Patrick, J.W.; Furbank, R.T. Pathway of sugar transport in germinating wheat seeds. Plant Physiol. 2006, 141, 1255–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arc, E.; Sechet, J.; Corbineau, F.; Rajjou, L.; Marion-Poll, A. ABA crosstalk with ethylene and nitric oxide in seed dormancy and germination. Front. Plant Sci. 2013, 4, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekkers, B.J.W.; Pearce, S.; van Bolderen-Veldkamp, R.P.; Marshall, A.; Widera, P.; Gilbert, J.; Drost, H.-G.; Bassel, G.W.; Müller, K.; King, J.R.; et al. Transcriptional dynamics of two seed compartments with opposing roles in Arabidopsis seed germination. Plant Physiol. 2013, 163, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Nonogaki, H. Seed dormancy and germination—Emerging mechanisms and new hypotheses. Front. Plant Sci. 2014, 5, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, C.; Northcote, D.H. The action of exogenous gibberellic acid on protein and mRNA in germinating castor bean seeds. Planta 1982, 154, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Newman, J.C.; Briggs, D.E. Glyceride metabolism and gluconeogenesis in barley endosperm. Phytochemistry 1976, 15, 1453–1458. [Google Scholar] [CrossRef]
- Arnalte, M.-E.; Cornejo, M.-J.; Bush, D.S.; Jones, R.L. Gibberellic acid stimulates lipid metabolism in barley aleurone protoplasts. Plant Sci. 1991, 77, 223–232. [Google Scholar] [CrossRef]
- Bethke, P.C.; Fath, A.; Jones, R.L. Regulation of viability and cell death by hormones in cereal aleurones. J. Plant Physiol. 2001, 158, 429–438. [Google Scholar] [CrossRef]
- Shu, K.; Zhang, H.; Wang, S.; Chen, M.; Wu, Y.; Tang, S.; Liu, C.; Feng, Y.; Cao, X.; Xie, Q. ABI4 regulates primary seed dormancy by regulating the biogenesis of abscisic acid and gibberellins in arabidopsis. PLoS Genet. 2013, 9, e1003577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailly, C. Active oxygen species and antioxydants in seed biology. Seed Sci. Res. 2004, 14, 93–107. [Google Scholar] [CrossRef]
- Feussner, I.; Wasternack, C.; Kindl, H.; Kuhn, H. Lipoxygenase-catalyzed oxygenation of storage lipids is implicated in lipid mobilization during germination. Proc. Natl. Acad. Sci. USA 1995, 92, 11849–11853. [Google Scholar] [CrossRef] [Green Version]
- Bailly, C. The signaling role of ROS in the regulation of seed germination and dormancy. Biochem. J. 2019, 20, 3019–3032. [Google Scholar] [CrossRef]
- Bailly, C.; El-Maarouf-Bouteau, H.; Corbineau, F. From intracellular signaling networks to cell death: the dual role of reactive oxygen species in seed physiology. Comptes Rendus Biol. 2008, 331, 806–814. [Google Scholar] [CrossRef]
- Chen, H.; Ruan, J.; Chu, P.; Fu, W.; Liang, Z.; Li, Y.; Tong, J.; Xiao, L.; Liu, J.; Li, C.; et al. AtPER1 enhances primary seed dormancy and reduces seed germination by suppressing the ABA catabolism and GA biosynthesis in Arabidopsis seeds. Plant J. 2020, 101, 310–323. [Google Scholar] [CrossRef]
- Palma, K.; Kermode, A. Metabolism of hydrogen peroxide during reserve mobilization and programmed cell death of barley (Hordeum vulgare L.) aleurone layer cells. Free Radic. Biol. Med. 2003, 35, 1261–1270. [Google Scholar] [CrossRef]
- Yan, D.; Duermeyer, L.; Leoveanu, C.; Nambara, E. The functions of the endosperm during seed germination. Plant Cell Physiol. 2014, 55, 1521–1533. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miray, R.; Kazaz, S.; To, A.; Baud, S. Molecular Control of Oil Metabolism in the Endosperm of Seeds. Int. J. Mol. Sci. 2021, 22, 1621. https://doi.org/10.3390/ijms22041621
Miray R, Kazaz S, To A, Baud S. Molecular Control of Oil Metabolism in the Endosperm of Seeds. International Journal of Molecular Sciences. 2021; 22(4):1621. https://doi.org/10.3390/ijms22041621
Chicago/Turabian StyleMiray, Romane, Sami Kazaz, Alexandra To, and Sébastien Baud. 2021. "Molecular Control of Oil Metabolism in the Endosperm of Seeds" International Journal of Molecular Sciences 22, no. 4: 1621. https://doi.org/10.3390/ijms22041621
APA StyleMiray, R., Kazaz, S., To, A., & Baud, S. (2021). Molecular Control of Oil Metabolism in the Endosperm of Seeds. International Journal of Molecular Sciences, 22(4), 1621. https://doi.org/10.3390/ijms22041621