
AP2M1 Supports TGF-β Signals to Promote Collagen Expression by Inhibiting Caveolin Expression

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
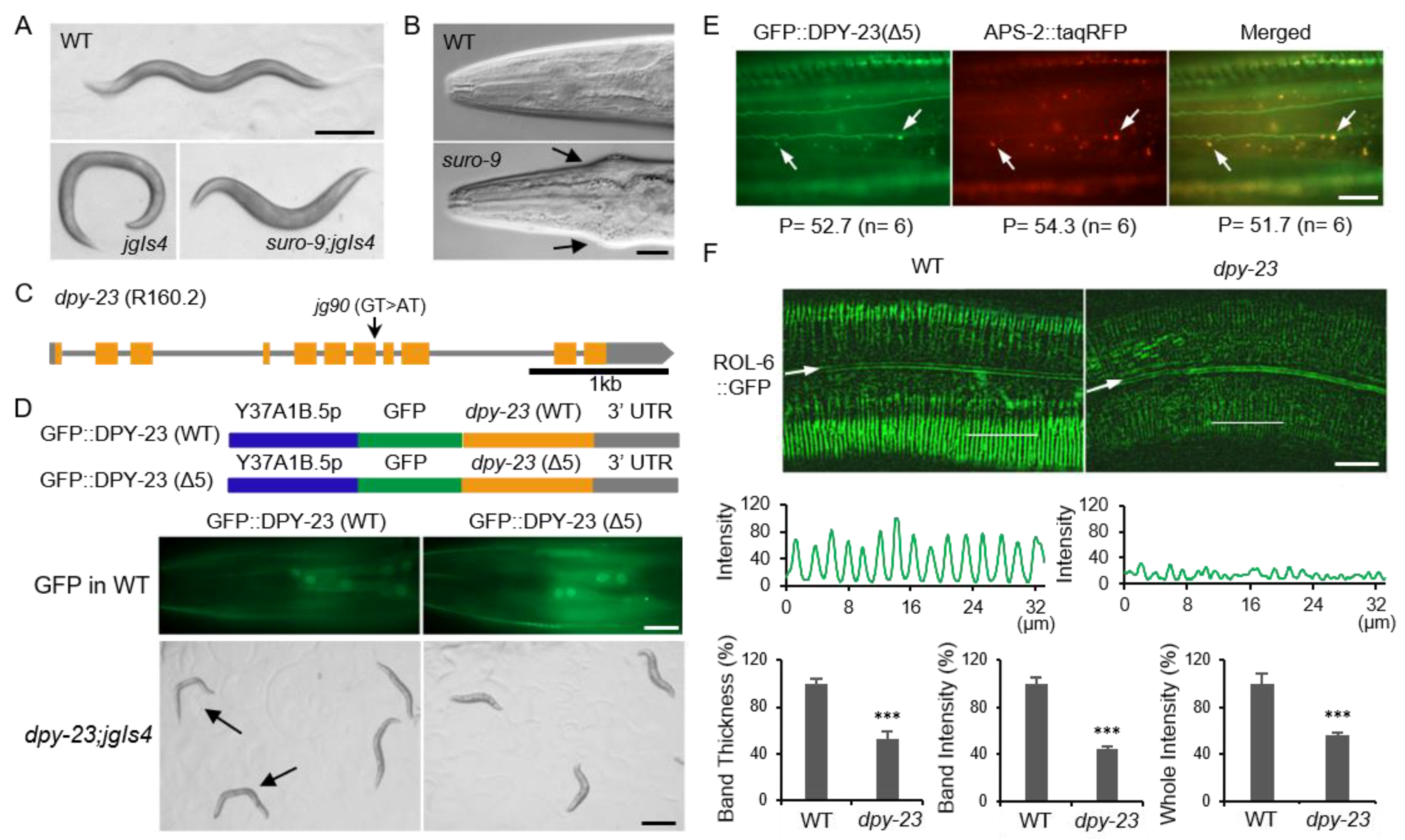
2.1. DPY-23/AP2M1 is Required for the Normal Cuticle
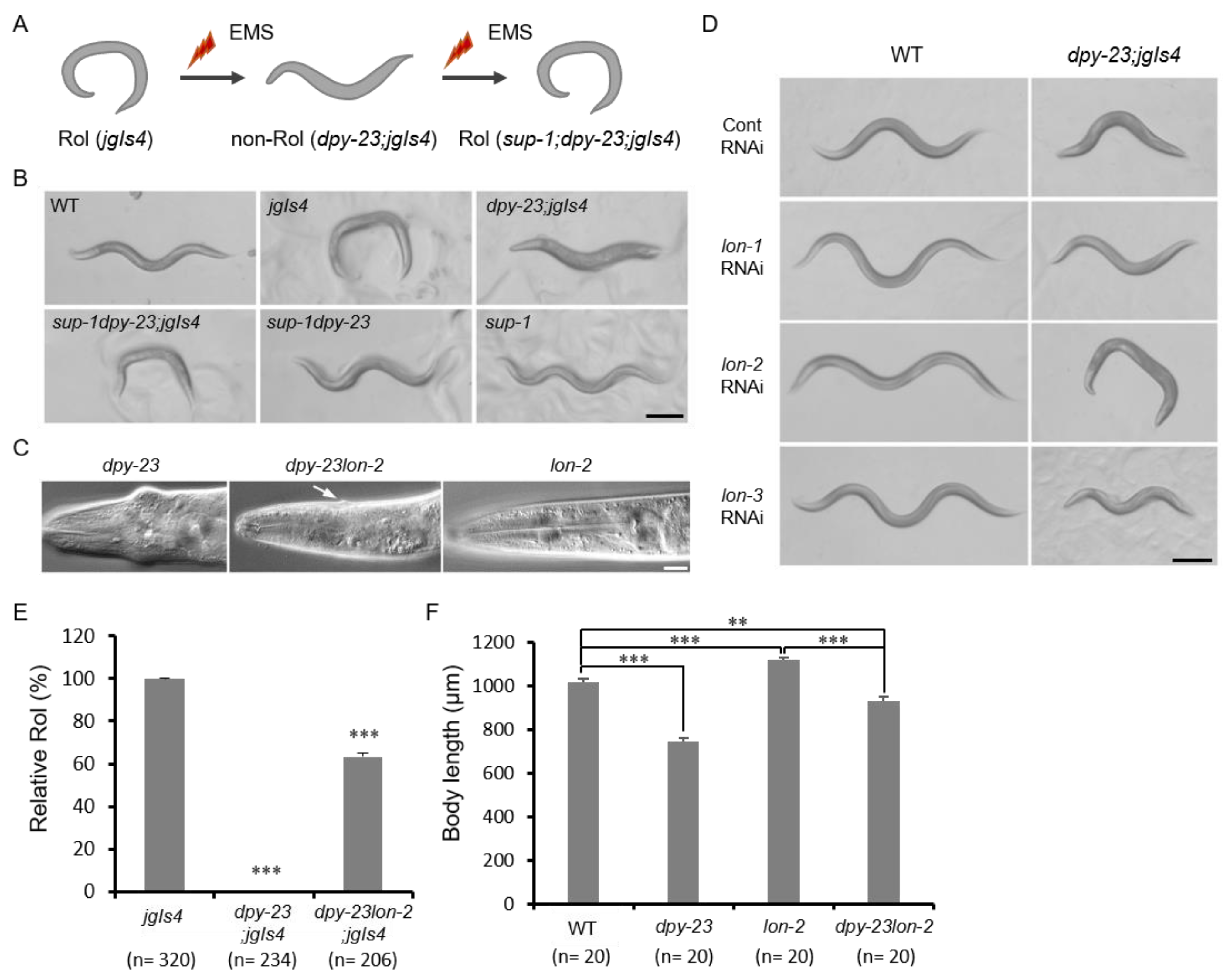
2.2. The Lon-2 Mutation Suppressed the Dpy-23 Mutant Phenotype
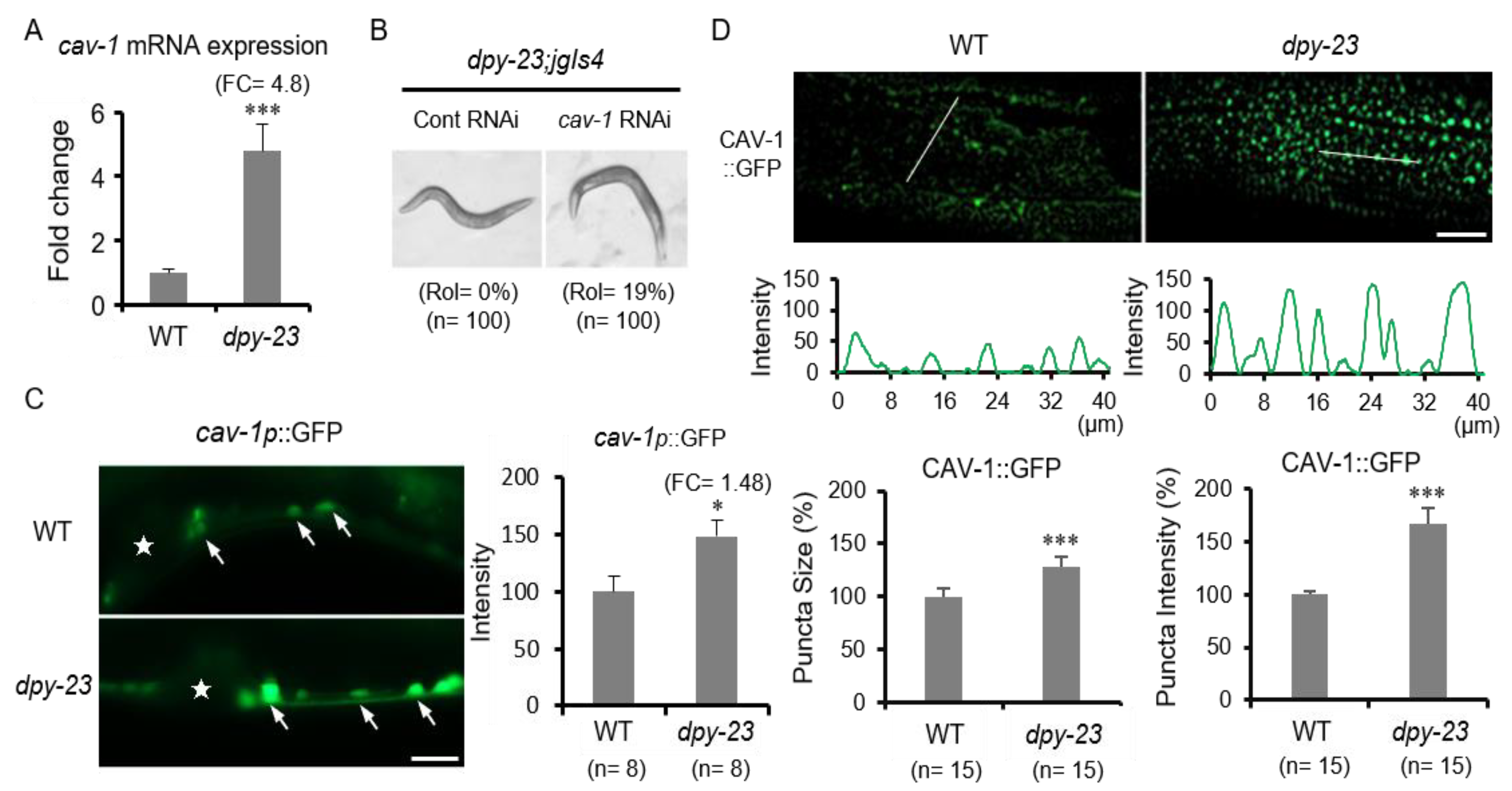
2.3. Caveolin Is Upregulated in the Dpy-23 Mutant
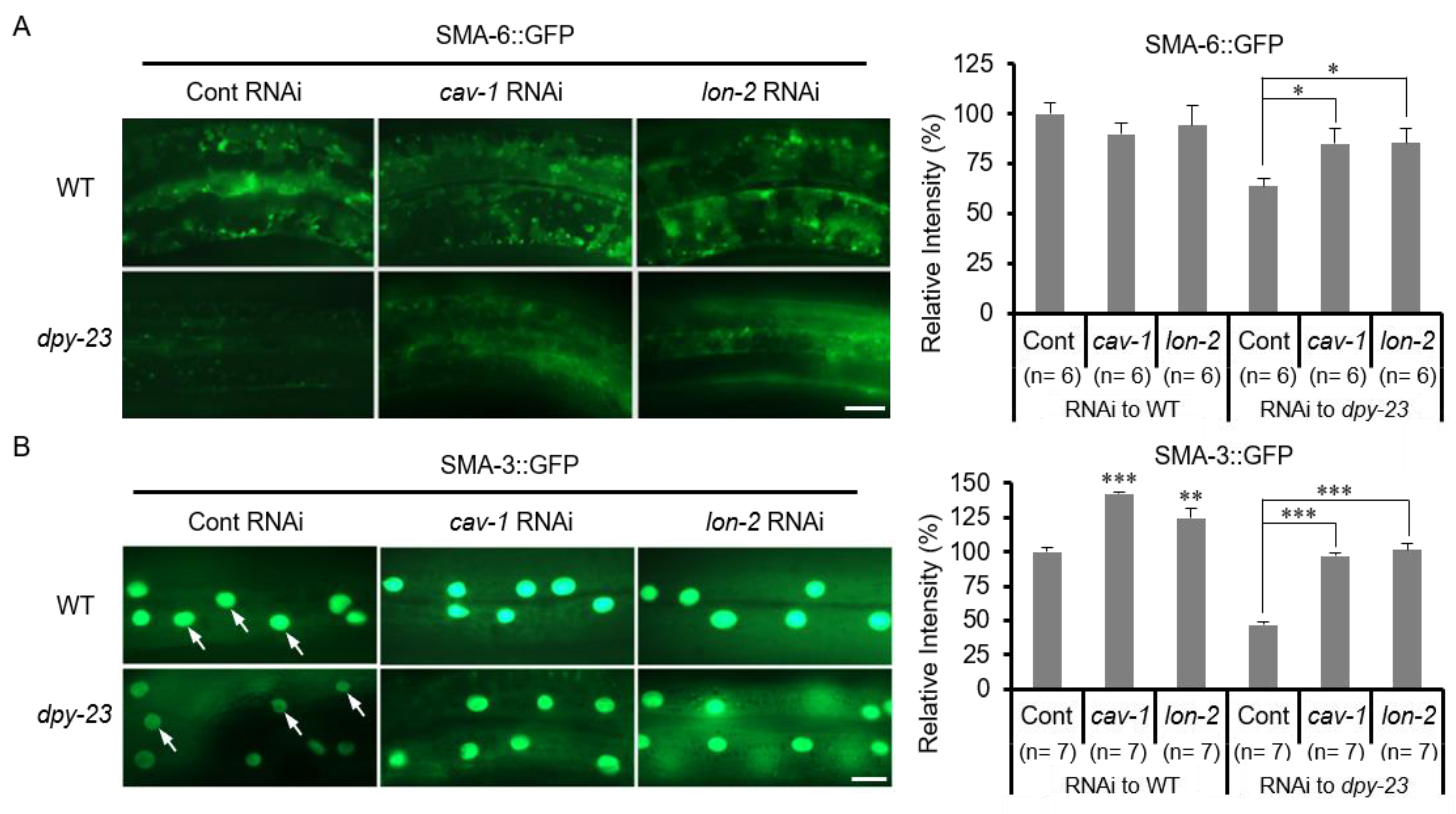
2.4. TGF-β Signaling Was Reduced in the Dpy-23 Mutant
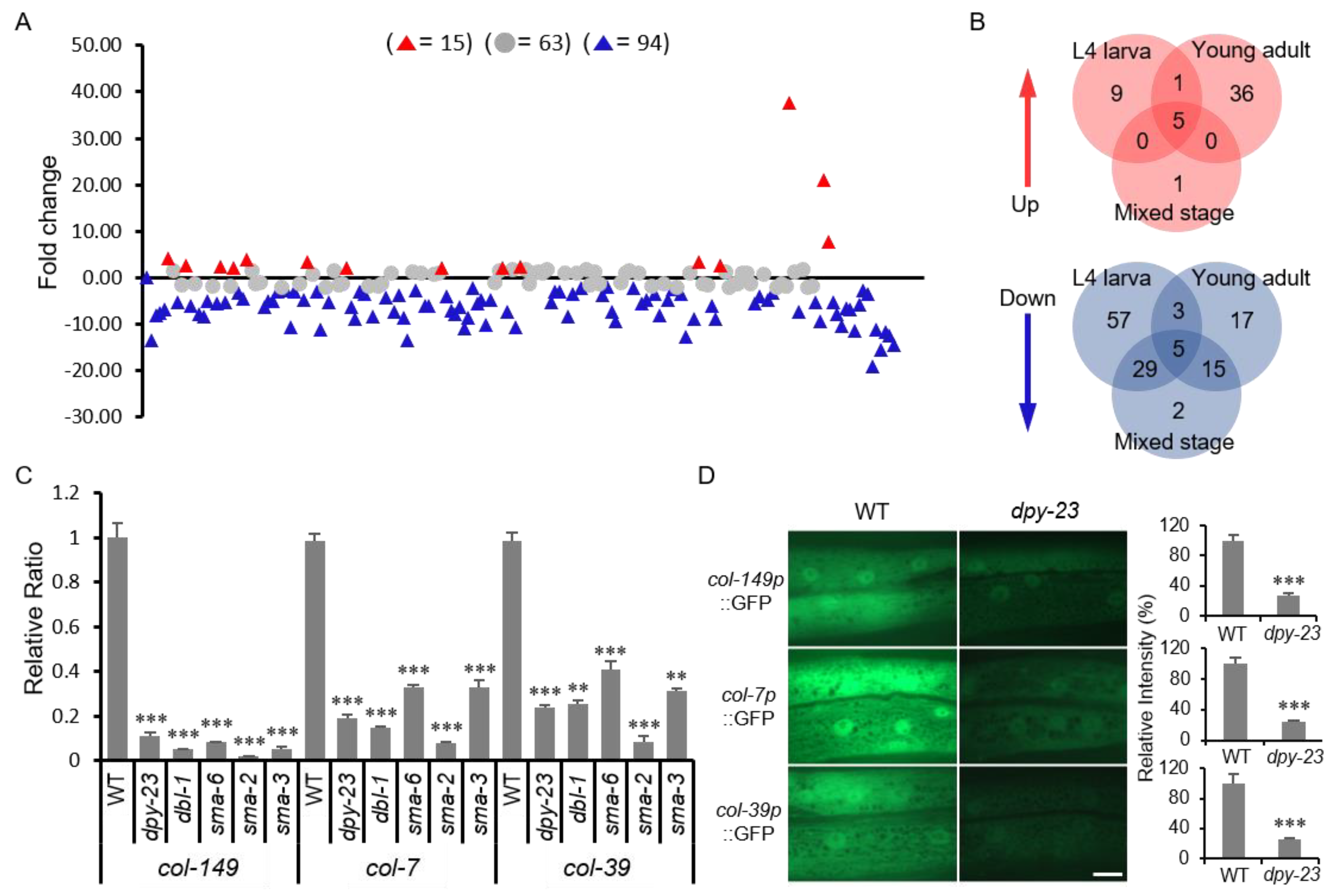
2.5. DPY-23 is Required for the Expression of Several Collagen Genes
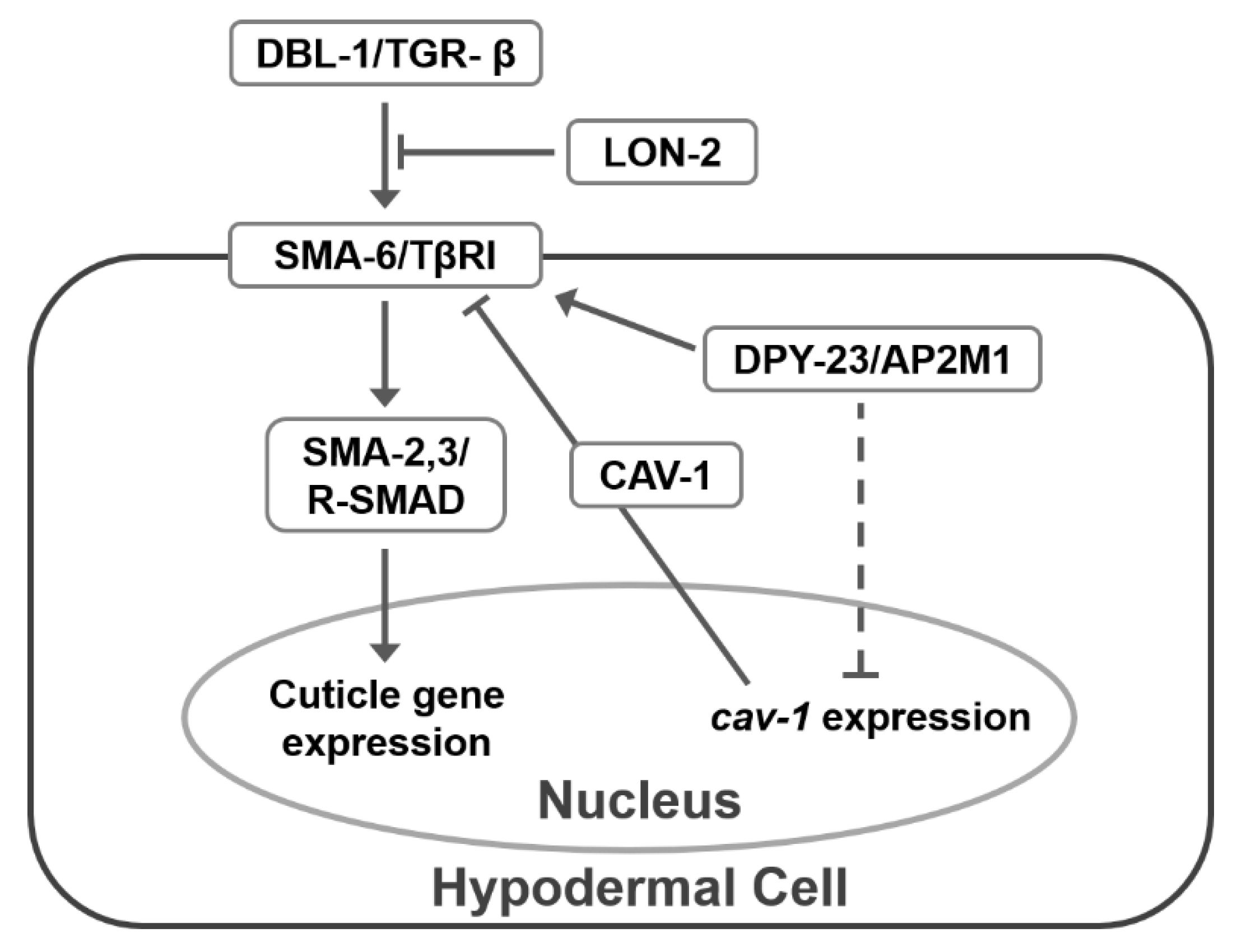
3. Discussion
4. Materials and Methods
4.1. C. elegans Culture and Strains
4.2. Mutagenesis and Mutant Cloning
4.3. Fluorescent Protein Fusion Reporter Constructs and Microinjection
4.4. RNA Extraction and RT-PCR
4.5. RNA Interference (RNAi)
4.6. RNA-seq Analysis
4.7. Microscopy and Statistics
4.8. Rol Ratio and Body Length Measurement
4.9. Quantitative Real-Time PCR (qRT-PCR)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AP-2 | Clathrin-associated protein complex II |
| ECM | Extracellular matrix |
| Dpy | dumpy |
| Lon | Long |
| Sma | Small |
| TGF-β | Tumor growth factor-beta |
| TβR | TGF-β receptor |
| CDE | clathrin-dependent endocytosis |
| CIE | clathrin-independent endocytosis |
| qRT-PCR | quantitative reverse transcription PCR |
References
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [Green Version]
- Karamanos, N.K.; ProQuest. Extracellular Matrix: Pathobiology and Signaling. eBOOK 2012. [Google Scholar] [CrossRef]
- Ricard-Blum, S. The collagen family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taffoni, C.; Pujol, N. Mechanisms of innate immunity in C. elegans epidermis. Tissue Barriers 2015, 3, e1078432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, A.P.; Johnstone, I.L. The cuticle. WormBook Online Rev. C. elegans Biol. 2007, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massague, J. TGF-beta signal transduction. Annu. Rev. Biochem. 1998, 67, 753–791. [Google Scholar] [CrossRef] [PubMed]
- Branton, M.H.; Kopp, J.B. TGF-beta and fibrosis. Microbes Infect. 1999, 1, 1349–1365. [Google Scholar] [CrossRef]
- Cutroneo, K.R.; White, S.L.; Phan, S.H.; Ehrlich, H.P. Therapies for bleomycin induced lung fibrosis through regulation of TGF-beta1 induced collagen gene expression. J. Cell. Physiol. 2007, 211, 585–589. [Google Scholar] [CrossRef]
- ten Dijke, P.; Hill, C.S. New insights into TGF-beta-Smad signalling. Trends Biochem. Sci. 2004, 29, 265–273. [Google Scholar] [CrossRef]
- Savage-Dunn, C.; Maduzia, L.L.; Zimmerman, C.M.; Roberts, A.F.; Cohen, S.; Tokarz, R.; Padgett, R.W. Genetic screen for small body size mutants in C. elegans reveals many TGFbeta pathway components. Genesis 2003, 35, 239–247. [Google Scholar] [CrossRef]
- Gumienny, T.L.; Savage-Dunn, C. TGF-beta signaling in C. elegans. WormBook 2013, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Gumienny, T.L.; MacNeil, L.T.; Wang, H.; de Bono, M.; Wrana, J.L.; Padgett, R.W. Glypican LON-2 is a conserved negative regulator of BMP-like signaling in Caenorhabditis elegans. Curr. Biol. 2007, 17, 159–164. [Google Scholar] [CrossRef] [Green Version]
- Di Guglielmo, G.M.; Le Roy, C.; Goodfellow, A.F.; Wrana, J.L. Distinct endocytic pathways regulate TGF-beta receptor signalling and turnover. Nat. Cell Biol. 2003, 5, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Del Galdo, F.; Lisanti, M.P.; Jimenez, S.A. Caveolin-1, transforming growth factor-beta receptor internalization, and the pathogenesis of systemic sclerosis. Curr. Opin. Rheumatol. 2008, 20, 713–719. [Google Scholar] [CrossRef]
- Chen, Y.G. Endocytic regulation of TGF-beta signaling. Cell Res. 2009, 19, 58–70. [Google Scholar] [CrossRef] [PubMed]
- Tzavlaki, K.; Moustakas, A. TGF-beta Signaling. Biomolecules 2020, 10, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, H.; Choudhury, A.; Pagano, R.E.; Leof, E.B. Ligand-dependent and -independent transforming growth factor-beta receptor recycling regulated by clathrin-mediated endocytosis and Rab11. Mol. Biol. Cell 2004, 15, 4166–4178. [Google Scholar] [CrossRef]
- Yao, D.; Ehrlich, M.; Henis, Y.I.; Leof, E.B. Transforming growth factor-beta receptors interact with AP2 by direct binding to beta2 subunit. Mol. Biol. Cell 2002, 13, 4001–4012. [Google Scholar] [CrossRef] [Green Version]
- Razani, B.; Zhang, X.L.; Bitzer, M.; von Gersdorff, G.; Bottinger, E.P.; Lisanti, M.P. Caveolin-1 regulates transforming growth factor (TGF)-beta/SMAD signaling through an interaction with the TGF-beta type I receptor. J. Biol. Chem. 2001, 276, 6727–6738. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.H.; Kim, D.H.; Nam, H.W.; Park, S.Y.; Shim, J.; Cho, J.W. Tyrosylprotein sulfotransferase regulates collagen secretion in Caenorhabditis elegans. Mol. Cells 2010, 29, 413–418. [Google Scholar] [CrossRef]
- Kim, T.H.; Kim, Y.J.; Cho, J.W.; Shim, J. A novel zinc-carboxypeptidase SURO-1 regulates cuticle formation and body morphogenesis in Caenorhabditis elegans. FEBS Lett. 2011, 585, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Beacham, G.M.; Partlow, E.A.; Lange, J.J.; Hollopeter, G. NECAPs are negative regulators of the AP2 clathrin adaptor complex. Elife 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Boehm, M.; Bonifacino, J.S. Genetic analyses of adaptin function from yeast to mammals. Gene 2002, 286, 175–186. [Google Scholar] [CrossRef]
- McKay, S.J.; Johnsen, R.; Khattra, J.; Asano, J.; Baillie, D.L.; Chan, S.; Dube, N.; Fang, L.; Goszczynski, B.; Ha, E.; et al. Gene expression profiling of cells, tissues, and developmental stages of the nematode C. elegans. Cold Spring Harb. Symp. Quant. Biol. 2003, 68, 159–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taneja-Bageshwar, S.; Gumienny, T.L. Two functional domains in C. elegans glypican LON-2 can independently inhibit BMP-like signaling. Dev. Biol. 2012, 371, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Lakdawala, M.F.; Madhu, B.; Faure, L.; Vora, M.; Padgett, R.W.; Gumienny, T.L. Genetic interactions between the DBL-1/BMP-like pathway and dpy body size-associated genes in Caenorhabditis elegans. Mol. Biol. Cell 2019, 30, 3151–3160. [Google Scholar] [CrossRef]
- Root, K.T.; Plucinsky, S.M.; Glover, K.J. Recent progress in the topology, structure, and oligomerization of caveolin: A building block of caveolae. Curr. Top. Membr. 2015, 75, 305–336. [Google Scholar] [CrossRef]
- Tang, Z.; Okamoto, T.; Boontrakulpoontawee, P.; Katada, T.; Otsuka, A.J.; Lisanti, M.P. Identification, sequence, and expression of an invertebrate caveolin gene family from the nematode Caenorhabditis elegans. Implications for the molecular evolution of mammalian caveolin genes. J. Biol. Chem. 1997, 272, 2437–2445. [Google Scholar] [CrossRef] [Green Version]
- Parker, S.; Walker, D.S.; Ly, S.; Baylis, H.A. Caveolin-2 is required for apical lipid trafficking and suppresses basolateral recycling defects in the intestine of Caenorhabditis elegans. Mol. Biol. Cell 2009, 20, 1763–1771. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Sato, M.; Audhya, A.; Oegema, K.; Schweinsberg, P.; Grant, B.D. Dynamic regulation of caveolin-1 trafficking in the germ line and embryo of Caenorhabditis elegans. Mol. Biol. Cell 2006, 17, 3085–3094. [Google Scholar] [CrossRef] [Green Version]
- Savage-Dunn, C.; Tokarz, R.; Wang, H.; Cohen, S.; Giannikas, C.; Padgett, R.W. SMA-3 smad has specific and critical functions in DBL-1/SMA-6 TGFbeta-related signaling. Dev. Biol. 2000, 223, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, S.; Morita, K.; Mochii, M.; Ueno, N. Hypodermal expression of Caenorhabditis elegans TGF-beta type I receptor SMA-6 is essential for the growth and maintenance of body length. Dev. Biol. 2001, 240, 32–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caja, L.; Dituri, F.; Mancarella, S.; Caballero-Diaz, D.; Moustakas, A.; Giannelli, G.; Fabregat, I. TGF-beta and the Tissue Microenvironment: Relevance in Fibrosis and Cancer. Int. J. Mol. Sci. 2018, 19, 1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, C.L.; Baum, P.D.; Gu, M.; Jorgensen, E.M.; Clark, S.G.; Garriga, G. C. elegans AP-2 and retromer control Wnt signaling by regulating mig-14/Wntless. Dev. Cell 2008, 14, 132–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saied-Santiago, K.; Townley, R.A.; Attonito, J.D.; da Cunha, D.S.; Diaz-Balzac, C.A.; Tecle, E.; Bulow, H.E. Coordination of Heparan Sulfate Proteoglycans with Wnt Signaling To Control Cellular Migrations and Positioning in Caenorhabditis elegans. Genetics 2017, 206, 1951–1967. [Google Scholar] [CrossRef] [Green Version]
- Gleason, R.J.; Akintobi, A.M.; Grant, B.D.; Padgett, R.W. BMP signaling requires retromer-dependent recycling of the type I receptor. Proc. Natl. Acad. Sci. USA 2014, 111, 2578–2583. [Google Scholar] [CrossRef] [Green Version]
- Madaan, U.; Yzeiraj, E.; Meade, M.; Clark, J.F.; Rushlow, C.A.; Savage-Dunn, C. BMP Signaling Determines Body Size via Transcriptional Regulation of Collagen Genes in Caenorhabditis elegans. Genetics 2018, 210, 1355–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakdawala, M.F.; Gumienny, T. Loss of dpy-2 and dpy-9 has stage-specific effects on DBL-1 pathway signaling. MicroPubl. Biol. 2019, 2019. [Google Scholar] [CrossRef]
- Zhao, B.; Wang, Q.; Du, J.; Luo, S.; Xia, J.; Chen, Y.G. PICK1 promotes caveolin-dependent degradation of TGF-beta type I receptor. Cell Res. 2012, 22, 1467–1478. [Google Scholar] [CrossRef] [Green Version]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Bigelow, H.; Doitsidou, M.; Sarin, S.; Hobert, O. MAQGene: Software to facilitate C. elegans mutant genome sequence analysis. Nat. Methods 2009, 6, 549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamath, R.S.; Martinez-Campos, M.; Zipperlen, P.; Fraser, A.G.; Ahringer, J. Effectiveness of specific RNA-mediated interference through ingested double-stranded RNA in Caenorhabditis elegans. Genome Biol. 2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Lim, G.-E.; Kim, Y.-N.; Koo, H.-S.; Shim, J. AP2M1 Supports TGF-β Signals to Promote Collagen Expression by Inhibiting Caveolin Expression. Int. J. Mol. Sci. 2021, 22, 1639. https://doi.org/10.3390/ijms22041639
Lee S, Lim G-E, Kim Y-N, Koo H-S, Shim J. AP2M1 Supports TGF-β Signals to Promote Collagen Expression by Inhibiting Caveolin Expression. International Journal of Molecular Sciences. 2021; 22(4):1639. https://doi.org/10.3390/ijms22041639
Chicago/Turabian StyleLee, Saerom, Ga-Eun Lim, Yong-Nyun Kim, Hyeon-Sook Koo, and Jaegal Shim. 2021. "AP2M1 Supports TGF-β Signals to Promote Collagen Expression by Inhibiting Caveolin Expression" International Journal of Molecular Sciences 22, no. 4: 1639. https://doi.org/10.3390/ijms22041639
APA StyleLee, S., Lim, G. -E., Kim, Y. -N., Koo, H. -S., & Shim, J. (2021). AP2M1 Supports TGF-β Signals to Promote Collagen Expression by Inhibiting Caveolin Expression. International Journal of Molecular Sciences, 22(4), 1639. https://doi.org/10.3390/ijms22041639