
Physiology and Physical Chemistry of Bile Acids




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Synthesis in the Liver and Storage in the Gallbladder: BAs in Cholesterol Metabolism
3. From the Gallbladder to the Intestine: Lipid Solubilization and Absorption
4. Antimicrobial Activity
5. Functionalized BAs in Medicine
6. BA-Based Polymers
7. BA Lipid Mixtures
8. BA Polymer Mixtures
9. Self-Assembly of Natural and Chemically Modified BAs
10. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BA | Bile acid |
| CA | Cholic acid |
| CDCA | Chenodeoxycholic acid |
| GCA | Glycocholic acid |
| TCA | Taurocholic acid |
| TCDCA | Taurochenodeoxycholic acid |
| GCDCA | Glycochenodeoxycholic acid |
| DCA | Deoxycholic acid |
| LCA | Lithocholic acid |
| CYP7A1 | Cholesterol 7α-hydroxylase |
| CYP27 | Sterol 27-hydroxylase |
| CY8B1 | 12α-hydroxylase |
| CYP7B1 | Oxoysterol 7α-hydroxylase |
| BAD | Bile acid derivative |
| PNIPAM | Poly(N-isopropyl acryl amide) |
| PAMPTMA | Poly((3-acrylamido-propyl)-trimethylammonium chloride |
| GDCA | Glycodeoxycholic acid |
| TDCA | Taurodeoxycholic acid |
References
- Thomas, C.; Pellicciari, R.; Pruzanski, M.; Auwerx, J.; Schoonjans, K. Targeting Bile-Acid Signalling for Metabolic Diseases. Nat. Rev. Drug Discov. 2008, 7, 678–693. [Google Scholar] [CrossRef]
- Barbier, O.; Trottier, J.; Kaeding, J.; Caron, P.; Verreault, M. Lipid-Activated Transcription Factors Control Bile Acid Glucuronidation. Mol. Cell. Biochem. 2009, 326, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Kuhajda, K.; Kandrac, J.; Kevresan, S.; Mikov, M.; Fawcett, J.P. Structure and Origin of Bile Acids: An Overview. Eur. J. Drug Metab. Pharm. 2006, 31, 135–143. [Google Scholar] [CrossRef]
- Madenci, D.; Egelhaaf, S.U. Self-Assembly in Aqueous Bile Salt Solution. Curr. Opin. Colloid Interface Sci. 2010, 15, 109–115. [Google Scholar] [CrossRef]
- Galantini, L.; di Gregorio, M.C.; Gubitosi, M.; Travaglini, L.; Vázquez, J.; Jover, A.; Meijide, F.; Soto, V.H.; Pavel, N.V. Bile Salts and Derivatives: Rigid Unconventional Amphiphiles as Dispersants, Carriers and Superstructure Building Blocks. Curr. Opin. Colloid Interface Sci. 2015, 20, 170–182. [Google Scholar] [CrossRef]
- di Gregorio, M.C.; Travaglini, L.; Del Giudice, A.; Cautela, J.; Pavel, N.V.; Galantini, L. Bile Salts: Natural Surfactants and Precursors of a Broad Family of Complex Amphiphiles. Langmuir 2019, 35, 6803–6821. [Google Scholar] [CrossRef]
- Quinn, R.A.; Melnik, A.V.; Vrbanac, A.; Fu, T.; Patras, K.A.; Christy, M.P.; Bodai, Z.; Belda-Ferre, P.; Tripathi, A.; Chung, L.K.; et al. Global Chemical Effects of the Microbiome Include New Bile-Acid Conjugations. Nature 2020, 579, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.S.; Magnusson, B.M.; Burczynski, F.J.; Weiss, M. Enterohepatic Circulation. Clin. Pharm. 2002, 41, 751–790. [Google Scholar] [CrossRef] [PubMed]
- Russell, D.W. Fifty Years of Advances in Bile Acid Synthesis and Metabolism. J. Lipid Res. 2009, 50, 120–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodwin, B.; Jones, S.A.; Price, R.R.; Watson, M.A.; McKee, D.D.; Moore, L.B.; Galardi, C.; Wilson, J.G.; Lewis, M.C.; Roth, M.E.; et al. A Regulatory Cascade of the Nuclear Receptors FXR, SHP-1, and LRH-1 Represses Bile Acid Biosynthesis. Mol. Cell 2000, 6, 517–526. [Google Scholar] [CrossRef]
- Lu, T.T.; Makishima, M.; Repa, J.J.; Schoonjans, K.; Kerr, T.A.; Auwerx, J.; Mangelsdorf, D.J. Molecular Basis for Feedback Regulation of Bile Acid Synthesis by Nuclear Receptos. Mol. Cell 2000, 6, 507–515. [Google Scholar] [CrossRef]
- Denson, L.A.; Sturm, E.; Echevarria, W.; Zimmerman, T.L.; Makishima, M.; Mangelsdorf, D.J.; Karpen, S.J. The Orphan Nuclear Receptor, Shp, Mediates Bile Acid-Induced Inhibition of the Rat Bile Acid Transporter, Ntcp. Gastroenterology 2001, 121, 140–147. [Google Scholar] [CrossRef]
- Kim, I.; Ahn, S.H.; Inagaki, T.; Choi, M.; Ito, S.; Guo, G.L.; Kliewer, S.A.; Gonzalez, F.J. Differential Regulation of Bile Acid Homeostasis by the Farnesoid X Receptor in Liver and Intestine. J. Lipid Res. 2007, 48, 2664–2672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, B.; Wang, L.; Chiang, J.Y.; Zhang, Y.; Klaassen, C.D.; Guo, G.L. Mechanism of Tissue-Specific Farnesoid X Recptor in Suppressing the Expression of Genes in Bile-Acid Synthesis in Mice. Hepatology 2012, 56, 1034–1043. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Francl, J.M.; Boehme, S.; Chiang, J.Y.L. Regulation of Cholesterol and Bile Acid Homeostasis by the Cholesterol 7α-Hydroxylase/Steroid Response Element-Binding Protein 2/MicroRNA-33a Axis in Mice. Hepatology 2013, 58, 1111–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meissner, M.; Wolters, H.; de Boer, R.A.; Havinga, R.; Boverhof, R.; Bloks, V.W.; Kuipers, F.; Groen, A.K. Bile Acid Sequestration Normalizes Plasma Cholesterol and Reduces Atherosclerosis in Hypercholesterolemic Mice. No Additional Effect of Physical Activity. Atherosclerosis 2013, 228, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Mazidi, M.; Rezaie, P.; Karimi, E.; Kengne, A.P. The effects of bile acid sequestrants on lipid profile and blood glucose concentrations: A systematic review and meta-analysis of randomized controlled trials. Int. J. Cardiol. 2017, 227, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, F.; Bloks, V.W.; Groen, A.K. Beyond intestinal soap—Bile acids in metabolic control. Nat. Rev. Endocrinol. 2014, 10, 488–498. [Google Scholar] [CrossRef]
- Hou, R.; Goldberg, A.C. Lowering Low-Density Lipoprotein Cholesterol:Statins, Ezetimibe, Bile Acid Sequestrants, and Combinations: Comparative Efficacy and Safety. Endocrinol. Metab. Clin. N. Am. 2009, 38, 79–97. [Google Scholar] [CrossRef] [PubMed]
- Sehayek, E.; Ono, J.G.; Shefer, S.; Nguyen, L.B.; Wang, N.; Batta, A.K.; Salen, G.; Smith, J.D.; Tall, A.R.; Breslow, J.L. Biliary Cholesterol Excretion: A Novel Mechanism That Regulates Dietary Cholesterol Absorption. Proc. Natl. Acad. Sci. USA 1998, 95, 10194–10199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandeville, W.; Goldberg, D. The Sequestration of Bile Acids, a Non-Absorbed Method for Cholesterol Reduction: A Review. Curr. Pharm. Des. 1997, 3, 15–28. [Google Scholar]
- Camilleri, M.; Gores, G.J. Therapeutic Targeting of Bile Acids. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 55905, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Lundasen, T.; Galman, C.; Angelini, B.; Rudling, M. Circulating Intestinal Fibroblast Growth Factor 19 Has a Pronounced Diurnal Variation and Modulates Hepatic Bile Acid Synthesis in Man. J. Intern. Med. 2006, 260, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Walters, J.R.F.; Tasleem, A.L.I.M.; Omer, O.S.; Brydon, W.G.; Dew, T.; Roux, C.W.L.E. A New Mechanism for Bile Acid Diarrhea: Defective Feedback Inhibition of Bile Acid Biosynthesis. Clin. Gastroenterol. Hepatol. 2009, 7, 1189–1194. [Google Scholar] [CrossRef] [Green Version]
- Johnston, I.; Nolan, J.; Pattni, S.S.; Walters, J.R.F. New Insights into Bile Acid Malabsorption. Curr. Gastroenterol. Rep. 2011, 13, 418–425. [Google Scholar] [CrossRef]
- Walters, J.R.F. Bile Acid Diarrhoea and FGF19: New Views. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, A.F.; Mysels, K.J. Bile Salts as Biological Surfactants. Colloids Surf. 1988, 30, 145–173. [Google Scholar] [CrossRef]
- Huval, C.C.; Holmes-Farley, S.R.; Mandeville, W.H.; Sacchiero, R.; Dhal, P.K. Syntheses of Hydrophobically Modified Cationic Hydrogels by Copolymerization of Alkyl Substituted Diallylamine Monomers and Their Use as Bile Acid Sequestrants. Eur. Polym. J. 2004, 40, 693–701. [Google Scholar] [CrossRef]
- Wilcox, C.; Turner, J.; Green, J. Systematic Review: The Management of Chronic Diarrhoea Due to Bile Acid Malabsorption. Aliment. Pharm. Ther. 2014, 39, 923–939. [Google Scholar] [CrossRef]
- Hansen, M.; Sonne, D.P.; Knop, F.K. Bile Acid Sequestrants: Glucose-Lowering Mechanisms and Efficacy in Type 2 Diabetes. Curr. Diab. Rep. 2014, 14, 482. [Google Scholar] [CrossRef]
- Seetharam, P.; Rodrigues, G. Short Bowel Syndrome: A Review of Management Options. Saudi J. Gastroenterol. 2011, 17, 229–235. [Google Scholar] [PubMed]
- Emmett, M.; Guirl, M.J.; Santa Ana, C.A.; Porter, J.L.; Neimark, S.; Hofmann, A.F.; Fordtran, J.S. Conjugated Bile Acid Replacement Therapy Reduces Urinary Oxalate Excretion in Short Bowel Syndrome. Am. J. Kidney Dis. 2003, 41, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Gruy-Kapral, C.; Little, K.H.; Fordtran, J.S.; Meziere, T.L.; Hagey, L.R.; Hofmann, A.F. Conjugated Bile ACid Replacement Theraphy for Short-Bowel Syndrome. Gastroenterology 1999, 116, 15–21. [Google Scholar] [CrossRef]
- Van de Heijning, B.J.M.; Stolk, M.F.J.; Van Erpecum, K.J.; Renooij, W.; Groen, A.K.; VanBerge-Henegouwen, G.P. Bile Salt-Induced Cholesterol Crystal Formation from Model Bile Vesicles: A Time Course Study. J. Lipid Res. 1994, 35, 1002–1011. [Google Scholar] [CrossRef]
- Tamesue, N.; Juniper, K. Concentrations of Bile Salts at the Critical Micellar Concentration of Human Gall Bladder Bile. Gastroenterology 1967, 52, 473–479. [Google Scholar] [CrossRef]
- Higuchi, W.I.; Tzeng, C.S.; Chang, S.J.; Chiang, H.J.; Liu, C.L. Estimation of Cholesterol Solubilization by a Mixed Micelle Binding Model in Aqueous Tauroursodeoxycholate:Lecithin:Cholesterol Solutions. J. Pharm. Sci. 2008, 97, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Reynier, M.O.; Montet, J.C.; Gerolami, A.; Marteau, C.; Crotte, C.; Montet, A.M.; Mathieu, S. Comparative Effects of Cholic, Chenodeoxycholic, and Ursodeoxycholic Acids on Micellar Solubilization and Intestinal Absorption of Cholesterol. J. Lipid Res. 1981, 22, 467–473. [Google Scholar] [CrossRef]
- Rudling, M.; Laskar, A.; Straniero, S. Gallbladder Bile Supersaturated with Cholesterol in Gallstone Patients Preferentially Develops from Shortage of Bile Acids. J. Lipid Res. 2019, 60, 498–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carey, M.C.; Small, D.M. The Physical Chemistry of Cholesterol Solubility in Bile. Relationship to Gallstone Formation and Dissolution in Man. J. Clin. Investig. 1978, 61, 998–1026. [Google Scholar] [CrossRef] [Green Version]
- Coreta-Gomes, F.M.; Vaz, W.L.C.; Wasielewski, E.; Geraldes, C.F.G.; Moreno, M.J. Quantification of Cholesterol Solubilized in Dietary Micelles: Dependence on Human Bile Salt Variability and the Presence of Dietary Food Ingredients. Langmuir 2016, 32, 4564–4574. [Google Scholar] [CrossRef]
- Portincasa, P. Therapy of Gallstone Disease: What It Was, What It Is, What It Will Be. World J. Gastrointest. Pharm. Ther. 2012, 3, 7–20. [Google Scholar] [CrossRef]
- Jang, S.I.; Fang, S.; Kim, K.P.; Ko, Y.; Kim, H.; Oh, J.; Hong, G.Y.; Lee, S.Y.; Kim, J.M.; Noh, I.; et al. Combination Treatment with N-3 Polyunsaturated Fatty Acids and Ursodeoxycholic Acid Dissolves Cholesterol Gallstones in Mice. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, J.L. Bile Formation and Secretion. Compr. Physiol. 2013, 3, 1035–1078. [Google Scholar] [PubMed] [Green Version]
- Funabashi, M.; Grove, T.L.; Wang, M.; Varma, Y.; McFadden, M.E.; Brown, L.C.; Guo, C.; Higginbottom, S.; Almo, S.C.; Fischbach, M.A. A Metabolic Pathway for Bile Acid Dehydroxylation by the Gut Microbiome. Nature 2020, 582, 566–570. [Google Scholar] [CrossRef] [PubMed]
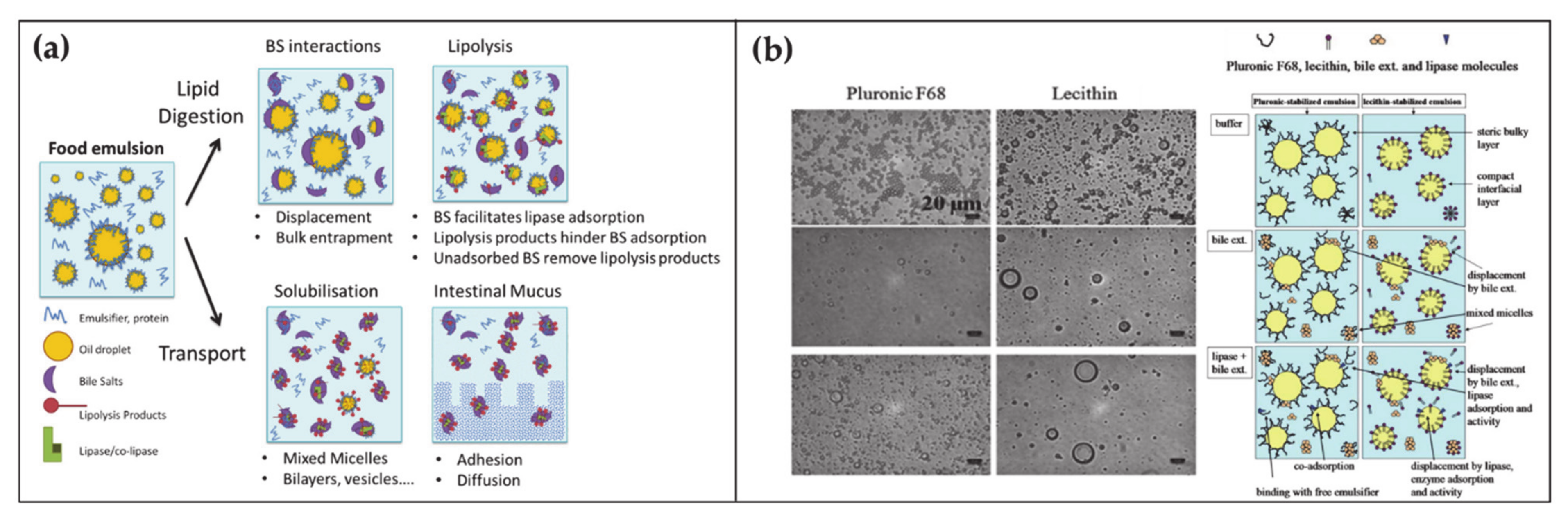
- Maldonado-Valderrama, J.; Wilde, P.; MacIerzanka, A.; MacKie, A. The Role of Bile Salts in Digestion. Adv. Colloid Interface Sci. 2011, 165, 36–46. [Google Scholar] [CrossRef]
- Macierzanka, A.; Torcello-Gómez, A.; Jungnickel, C.; Maldonado-Valderrama, J. Bile Salts in Digestion and Transport of Lipids. Adv. Colloid Interface Sci. 2019, 274, 102045. [Google Scholar] [CrossRef]
- Naso, J.N.; Bellesi, F.A.; Pizones Ruiz-Henestrosa, V.M.; Pilosof, A.M.R. Studies on the Interactions between Bile Salts and Food Emulsifiers under in Vitro Duodenal Digestion Conditions to Evaluate Their Bile Salt Binding Potential. Colloids Surf. B. 2019, 174, 493–500. [Google Scholar] [CrossRef]
- Cremers, C.M.; Knoefler, D.; Vitvitsky, V.; Banerjee, R.; Jakob, U. Bile Salts Act as Effective Protein-Unfolding Agents and Instigators of Disulfide Stress in Vivo. Proc. Natl. Acad. Sci. USA 2014, 111, E1610–E1619. [Google Scholar] [CrossRef] [Green Version]
- Pabois, O.; Lorenz, C.D.; Harvey, R.D.; Grillo, I.; Grundy, M.M.L.; Wilde, P.J.; Gerelli, Y.; Dreiss, C.A. Molecular Insights into the Behaviour of Bile Salts at Interfaces: A Key to Their Role in Lipid Digestion. J. Colloid Interface Sci. 2019, 556, 266–277. [Google Scholar] [CrossRef]
- Moghimipour, E.; Ameri, A.; Handali, S. Absorption-Enhancing Effects of Bile Salts. Molecules 2015, 20, 14451–14473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naumann, S.; Schweiggert-Weisz, U.; Eglmeier, J.; Haller, D.; Eisner, P. In Vitro Interactions of Dietary Fibre Enriched Food Ingredients with Primary and Secondary Bile Acids. Nutrients 2019, 11, 1424. [Google Scholar] [CrossRef] [Green Version]
- Shokry, D.S.; Waters, L.J.; Parkes, G.M.B.; Mitchell, J.C.; Snowden, M.J. Formation of a Bile Salt-Drug Hydrogel to Predict Human Intestinal Absorption. J. Pharm. Sci. 2019, 108, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Torcello-Gómez, A.; Maldonado-Valderrama, J.; Martín-Rodríguez, A.; McClements, D.J. Physicochemical Properties and Digestibility of Emulsified Lipids in Simulated Intestinal Fluids: Influence of Interfacial Characteristics. Soft Matter 2011, 7, 6167–6177. [Google Scholar] [CrossRef]
- Hofmann, A.F.; Eckmann, L. How Bile Acids Confer Gut Mucosal Protection against Bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 4333–4334. [Google Scholar] [CrossRef] [Green Version]
- Begley, M.; Gahan, C.G.M.; Hill, C. The Interaction between Bacteria and Bile. FEMS Microbiol. Rev. 2005, 29, 625–651. [Google Scholar] [CrossRef] [Green Version]
- Pazzi, P.; Puviani, A.C.; Libera, M.D.; Guerra, G.; Ricci, D.; Gullini, S.; Ottolenghi, C. Bile Salt-Induced Cytotoxicity and Ursodeoxycholate Cytoprotection: In-Vitro Study in Perifused Rat Hepatocytes. Eur. J. Gastroenterol. Hepatol. 1997, 9, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.Y.; Shaffer, E.A.; Costerton, J.W. Antibacterial Activity of Bile Salts against Common Biliary Pathogens—Effects of Hydrophobicity of the Molecule and in the Presence of Phospholipids. Dig. Dis. Sci. 1993, 38, 2104–2112. [Google Scholar] [CrossRef] [PubMed]
- Leverrier, P.; Dimova, D.; Pichereau, V.; Auffray, Y.; Boyaval, P.; Jan, G. Susceptibility and Adaptive Response to Bile Salts in Propionibacterium Freudenreichii: Physiological and Proteomic Anlysis. Appl. Environ. Microbiol. 2003, 69, 3809–3818. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, T.; Mori, M. Influence of Various Bile Salts on β-Glucuronidase Activity of Intestinal Bacteria. Lett. Appl. Microbiol. 1997, 25, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Coleman, R.; Lowe, P.J.; Billington, D. Membrane Lipid Composition and Susceptibility to Bile Salt Damage. BBA Biomembr. 1980, 599, 294–300. [Google Scholar] [CrossRef]
- Kociubinski, G.; Gómez Zavaglia, A.; Pérez, P.F.; Disalvo, E.A.; De Antoni, G.L. Effect of Bile Components on the Surface Properties of Bifidobacteria. J. Dairy Res. 2002, 69, 293–302. [Google Scholar] [CrossRef]
- Tian, Y.; Gui, W.; Koo, I.; Smith, P.B.; Allman, E.L.; Nichols, R.G.; Rimal, B.; Cai, J.; Liu, Q.; Patterson, A.D. The Microbiome Modulating Activity of Bile Acids. Gut Microbes 2020, 11, 979–996. [Google Scholar] [CrossRef]
- Small, D.M.; Lilly, H.S.; Hamilton, J.A.; Cabral, D.J. Transbilayer Movement of Bile Acids in Model Membranes. Biochemistry 1987, 26, 1801–1804. [Google Scholar]
- Inagaki, T.; Moschetta, A.; Lee, Y.K.; Peng, L.; Zhao, G.; Downes, M.; Yu, R.T.; Shelton, J.M.; Richardson, J.A.; Repa, J.J.; et al. Regulation of Antibacterial Defense in the Small Intestine by the Nuclear Bile Acid Receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 3920–3925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.D.; Myers, C.J.; Harris, S.C.; Kakiyama, G.; Lee, I.K.; Yun, B.S.; Matsuzaki, K.; Furukawa, M.; Min, H.K.; Bajaj, J.S.; et al. Bile Acid 7α-Dehydroxylating Gut Bacteria Secrete Antibiotics That Inhibit Clostridium Difficile: Role of Secondary Bile Acids. Cell Chem. Biol. 2019, 26, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Han, M.; Heinrich, B.; Fu, Q.; Zhang, Q.; Sandhu, M.; Agdashian, D.; Terabe, M.; Berzofsky, J.A.; Fako, V.; et al. Gut Microbiome–Mediated Bile Acid Metabolism Regulates Liver Cancer via NKT Cells. Science 2018, 360, eaan5931. [Google Scholar] [CrossRef] [Green Version]
- Mao, J.; Chen, X.; Wang, C.; Li, W.; Li, J. Effects and Mechanism of the Bile Acid (Farnesoid X) Receptor on the Wnt/β-Catenin Signaling Pathway in Colon Cancer. Oncol. Lett. 2020, 20, 337–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Ciaula, A.; Wang, D.Q.H.; Molina, E.M.; Baccetto, R.L.; Calamita, G.; Palmieri, V.O.; Portincasa, P. Bile Acids and Cancer: Direct and Environmental-Dependent Effects. Ann. Hepatol. 2017, 16, S87–S105. [Google Scholar] [CrossRef] [PubMed]
- Pavlović, N.; Goločorbin-Kon, S.; Danić, M.; Stanimirov, B.; Al-Salami, H.; Stankov, K.; Mikov, M. Bile Acids and Their Derivatives as Potential Modifiers of Drug Release and Pharmacokinetic Profiles. Front. Pharm. 2018, 9, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Q.H.; Carey, M.C. Therapeutic Uses of Animal Biles in Traditional Chinese Medicine: An Ethnopharmacological, Biophysical Chemical and Medicinal Review. World J. Gastroenterol. 2014, 20, 9952–9975. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, A.A.; Beach, A.; Davies, G.F.; Harkness, T.A.A.; LeBlanc, A.; Titorenko, V.I. Lithocholic Bile Acid Selectively Kills Neuroblastoma Cells, While Sparing Normal Neuronal Cells. Oncotarget 2011, 2, 761–782. [Google Scholar] [CrossRef] [Green Version]
- Luu, T.H.; Bard, J.M.; Carbonnelle, D.; Chaillou, C.; Huvelin, J.M.; Bobin-Dubigeon, C.; Nazih, H. Lithocholic Bile Acid Inhibits Lipogenesis and Induces Apoptosis in Breast Cancer Cells. Cell. Oncol. 2018, 41, 13–24. [Google Scholar] [CrossRef]
- Goldberg, A.A.; Titorenko, V.I.; Beach, A.; Thomas Sanderson, J. Bile Acids Induce Apoptosis Selectively in Androgen-Dependent and -Independent Prostate Cancer Cells. PeerJ 2013, 1, e122. [Google Scholar] [CrossRef]
- Trah, J.; Arand, J.; Oh, J.; Pagerols-Raluy, L.; Trochimiuk, M.; Appl, B.; Heidelbach, H.; Vincent, D.; Saleem, M.A.; Reinshagen, K.; et al. Lithocholic Bile Acid Induces Apoptosis in Human Nephroblastoma Cells: A Non-Selective Treatment Option. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Goossens, J.F.; Bailly, C. Ursodeoxycholic Acid and Cancer: From Chemoprevention to Chemotherapy. Pharm. Ther. 2019, 203, 107396. [Google Scholar] [CrossRef]
- Blanchet, M.; Brunel, J.M. Bile Acid Derivatives: From Old Molecules to a New Potent Therapeutic Use: An Overview. Curr. Med. 2018, 25, 3613–3636. [Google Scholar] [CrossRef] [PubMed]
- Suh, H.; Jung, E.J.; Kim, T.H.; Lee, H.Y.; Park, Y.H.; Kim, K.W. Anti-angiogenic activity of ursodeoxycholic acid and its derivatives. Cancer Lett. 1997, 113, 117–122. [Google Scholar] [CrossRef]
- Liu, H.; Qin, C.K.; Han, G.Q.; Xu, H.W.; Ren, W.H.; Qin, C.Y. Synthetic chenodeoxycholic acid derivative, HS-1200, induces apoptosis of human hepatoma cells via a mitochondrial pathway. Cancer Lett. 2008, 270, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Park, S.E.; Lee, S.W.; Hossain, M.A.; Kim, M.Y.; Kim, M.N.; Ahn, E.Y.; Park, Y.C.; Suh, H.; Kim, G.Y.; Choi, Y.H.; et al. A chenodeoxycholic derivative, HS-1200, induces apoptosis and cell cycle modulation via Egr-1 gene expression control on human hepatoma cells. Cancer Lett. 2008, 270, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Im, E.; Choi, Y.H.; Paik, K.J.; Suh, H.; Jin, Y.; Kim, K.W.; Yoo, Y.H.; Kim, N.D. Novel bile acid derivatives induce apoptosis via a p53-independent pathway in human breast carcinoma cells. Cancer Lett. 2001, 163, 83–93. [Google Scholar] [CrossRef]
- Im, E.-O.; Lee, S.; Suh, H.; Kim, K.-W.; Bae, Y.T.; Kim, N.D. A novel ursodeoxycholic acid derivative induces apoptosis in human MCF-7 breast cancer cells. Pharm. Pharmacol. Commun. 1999, 5, 293–298. [Google Scholar] [CrossRef]
- Kim, N.D.; Im, E.-O.; Hyun Yoo, Y.; Hyun Choi, Y. Modulation of the cell cycle and induction of apoptosis in human cancer cells by synthetic bile acids. Curr. Cancer Drug Targets 2006, 6, 681–689. [Google Scholar] [PubMed]
- Choi, Y.H.; Im, E.O.; Suh, H.; Jin, Y.; Yoo, Y.H.; Kim, N.D. Apoptosis and modulation of cell cycle control by synthetic derivatives of ursodeoxycholic acid and chenodeoxycholic acid in human prostate cancer cells. Cancer Lett. 2003, 199, 157–167. [Google Scholar] [CrossRef]
- Park, S.; Choi, H.J.; Yee, S.B.; Chung, H.Y.; Suh, H.; Choi, Y.H.; Yoo, Y.H.; Kim, N.D. Synthetic bile acid derivatives inhibit cell proliferation and induce apoptosis in HT-29 human colon cancer cells. Int. J. Oncol. 2004, 25, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Brossard, D.; El Kihal, L.; Clément, M.; Sebbahi, W.; Khalid, M.; Roussakis, C.; Rault, S. Synthesis of bile acid derivatives and in vitro cytotoxic activity with pro-apoptotic process on multiple myeloma (KMS-11), glioblastoma multiforme (GBM), and colonic carcinoma (HCT-116) human cell lines. Eur. J. Med. Chem. 2010, 45, 2912–2918. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Kim, Y.S.; Lee, G.Y.; Nam, J.O.; Lee, S.K.; Park, R.W.; Kim, S.Y.; Kim, I.S.; Byun, Y. Antiangiogenic effect of bile acid acylated heparin derivative. Pharm. Res. 2006, 24, 176–185. [Google Scholar] [CrossRef]
- Park, K.; Lee, S.K.; Son, D.H.; Park, S.A.; Kim, K.; Chang, H.W.; Jeong, E.J.; Park, R.W.; Kim, I.S.; Kwon, I.C.; et al. The attenuation of experimental lung metastasis by a bile acid acylated-heparin derivative. Biomaterials 2007, 28, 2667–2676. [Google Scholar] [CrossRef]
- Singh, M.; Singh, A.; Kundu, S.; Bansal, S.; Bajaj, A. Deciphering the role of charge, hydration, and hydrophobicity for cytotoxic activities and membrane interactions of bile acid based facial amphiphiles. Biochim. Biophys. Acta Biomembr. 2013, 1828, 1926–1937. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, D.S.; Shilpa Anantaraju, H.; Sriram, D.; Yogees-wari, P.; Nanjegowda, S.H.; Mallu, P.; Sakhuja, R. Synthesis, characterization and biological evaluation of bile acid-aromatic/heteroaromatic amides linked via amino acids as anti-cancer agents. Steroids 2016, 107, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, E.; Chinaglia, N.; Capobianco, M.L.; Marchetti, P.; Huang, T.E.; Weng, H.C.; Guh, J.H.; Hsu, L.C.; Perrone, D.; Navacchia, M.L. Dihydroartemisinin–Bile Acid Hybridization as an Effective Approach to Enhance Dihydroartemisinin Anticancer Activity. ChemMedChem 2019, 14, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Deng, Y.; Hsu, J.; Leu, W.; Marchesi, E.; Perrone, D.; Hsu, L. Evaluation of the Anticancer Activity of a Bile Acid-Dihydroartemisinin Hybrid in Hepatocellular Carcinoma Cells. 2020, 11, 1–14. Front. Pharmacol. 2020, 11, 1–14. [Google Scholar]
- Xiao, L.; Yu, E.; Yue, H.; Li, Q. Enhanced Liver Targeting of Camptothecin via Conjugation with Deoxycholic Acid. Molecules 2019, 24, 1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navacchia, M.L.; Marchesi, E.; Mari, L.; Chinaglia, N.; Gallerani, E.; Gavioli, R.; Capobianco, M.L.; Perrone, D. Rational Design of Nucleoside-Bile Acid Conjugates Incorporating a Triazole Moiety for Anticancer Evaluation and SAR Exploration. Molecules 2017, 22, 1710. [Google Scholar] [CrossRef] [Green Version]
- Seroka, B.; Łotowski, Z.; Hryniewicka, A.; Rárová, L.; Sicinski, R.R.; Tomkiel, A.M.; Morzycki, J.W. Synthesis of New Cisplatin Derivatives from Bile Acids. Molecules 2020, 25, 655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navacchia, M.L.; Fraix, A.; Chinaglia, N.; Gallerani, E.; Perrone, D.; Cardile, V.; Graziano, A.C.E.; Capobianco, M.L.; Sortino, S. NO Photoreleaser-Deoxyadenosine and -Bile Acid Derivative Bioconjugates as Novel Potential Photochemotherapeutics. ACS Med. Chem. Lett. 2016, 7, 939–943. [Google Scholar] [CrossRef] [Green Version]
- Salomatina, O.V.; Popadyuk, I.I.; Zakharenko, A.L.; Olga, D.; Chepanova, A.A.; Dyrkheeva, N.S.; Komarova, N.I.; Reynisson, J.; Anarbaev, R.O.; Salakhutdinov, N.F.; et al. Deoxycholic Acid as a Molecular Scaffold for Tyrosyl-DNA Phosphodiesterase 1 Inhibition: A Synthesis, Structure-Activity Relationship and Molecular Modeling Study. Steroids 2020, 165, 108771. [Google Scholar] [CrossRef]
- Salomatina, O.V.; Popadyuk, I.I.; Zakharenko, A.L.; Zakharova, O.D.; Fadeev, D.S.; Komarova, N.I.; Reynisson, J.; Arabshahi, H.J.; Chand, R.; Volcho, K.P.; et al. Novel Semisynthetic Derivatives of Bile Acids as Effective Tyrosyl-DNA Phosphodiesterase 1 Inhibitors. Molecules 2018, 23, 679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schölmerich, J.; Becher, M.-S.; Schmidt, K.; Schubert, R.; Kremer, B.; Feldhaus, S.; Gerok, W. Influence of Hydroxylation and Conjugation of Bile Salts on Their Membrane-Damaging Properties-Studies on Isolated Hepatocytes and Lipid Membrane Vesicles. Hepatology 1984, 4, 661–666. [Google Scholar]
- Sannasiddappa, T.H.; Lund, P.A.; Clarke, S.R. In Vitro Antibacterial Activity of Unconjugated and Conjugated Bile Salts on Staphylococcus Aureus. Front. Microbiol. 2017, 8, 1581. [Google Scholar] [CrossRef] [Green Version]
- Neves, A.R.; Almeida, J.R.; Carvalhal, F.; Câmara, A.; Pereira, S.; Antunes, J.; Vasconcelos, V.; Pinto, M.; Silva, E.R.; Sousa, E.; et al. Overcoming Environmental Problems of Biocides: Synthetic Bile Acid Derivatives as a Sustainable Alternative. Ecotoxicol. Environ. Saf. 2020, 187, 109812. [Google Scholar] [CrossRef]
- Guan, Q.; Li, C.; Schmidt, E.J.; Boswell, J.S.; Walsh, J.P.; Allman, G.W.; Savage, P.B. Preparation and Characterization of Cholic Acid-Derived Antimicrobial Agents with Controlled Stabilities. Org. Lett. 2000, 2, 2837–2840. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.; Guan, Q.; Walsh, J.P.; Boswell, J.S.; Winter, T.W.; Winter, E.S.; Boyd, S.S.; Li, C.; Savage, P.B. Correlation of the Antibacterial Activities of Cationic Peptide Antibiotics and Cationic Steroid Antibiotics. J. Med. Chem. 2002, 45, 663–669. [Google Scholar] [CrossRef]
- Savage, P.B.; Li, C.; Taotafa, U.; Ding, B.; Guan, Q. Antibacterial Properties of Cationic Steroid Antibiotics. FEMS Microbiol. Lett. 2002, 217, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Sha, Y.; Jui, M.S.; Lamm, M.E.; Ma, Y.; Tang, C. Facial Amphiphilicity-Induced Self-Assembly (FAISA) of Amphiphilic Copolymers. Macromolecules 2019, 52, 9526–9535. [Google Scholar] [CrossRef]
- Rahman, M.A.; Jui, M.S.; Bam, M.; Cha, Y.; Luat, E.; Alabresm, A.; Nagarkatti, M.; Decho, A.W.; Tang, C. Facial Amphiphilicity-Induced Polymer Nanostructures for Antimicrobial Applications. ACS Appl. Mater. Interfaces 2020, 12, 21221–21230. [Google Scholar] [CrossRef]
- Rahman, M.A.; Bam, M.; Luat, E.; Jui, M.S.; Ganewatta, M.S.; Shokfai, T.; Nagarkatti, M.; Decho, A.W.; Tang, C. Macromolecular-Clustered Facial Amphiphilic Antimicrobials. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kramer, W.; Glombik, H. Bile Acid Reabsorption Inhibitors (BARI): Novel Hypolipidemic Drugs. Curr. Med. Chem. 2006, 13, 997–1016. [Google Scholar] [CrossRef] [PubMed]
- Tolle-Sander, S.; Lentz, K.A.; Maeda, D.Y.; Coop, A.; Polli, J.E. Increased Acyclovir Oral Bioavailability via a Bile Acid Conjugate. Mol. Pharm. 2004, 1, 40–48. [Google Scholar] [CrossRef]
- Jha, S.K.; Chung, J.Y.; Pangeni, R.; Choi, H.S.; Subedi, L.; Kweon, S.; Choi, J.U.; Byun, Y.; Kim, Y.H.; Park, J.W. Enhanced Antitumor Efficacy of Bile Acid-Lipid Complex-Anchored Docetaxel Nanoemulsion via Oral Metronomic Scheduling. J. Control. Release 2020, 328, 368–394. [Google Scholar] [CrossRef] [PubMed]
- Sievänen, E. Exploitation of Bile Acid Transport Systems in Prodrug Design. Molecules 2007, 12, 1859–1889. [Google Scholar] [CrossRef] [Green Version]
- Kramer, W. Transporters, Trojan Horses and Therapeutics: Suitability of Bile Acid and Peptide Transporters for Drug Delivery. Biol. Chem. 2011, 392, 77–94. [Google Scholar] [CrossRef]
- Subramanian, S.; Iles, T.; Ikramuddin, S.; Steer, C.J. Merit of an Ursodeoxycholic Acid Clinical Trial in COVID-19 Patients. Vaccines 2020, 8, 320. [Google Scholar] [CrossRef] [PubMed]
- Carino, A.; Moraca, F.; Fiorillo, B.; Marchianò, S.; Sepe, V.; Biagioli, M.; Finamore, C.; Bozza, S.; Francisci, D.; Distrutti, E.; et al. Hijacking SARS-CoV-2/ACE2 Receptor Interaction by Natural and Semi-Synthetic Steroidal Agents Acting on Functional Pockets on the Receptor Binding Domain. Front. Chem. 2020, 8, 1–15. [Google Scholar] [CrossRef]
- Zhao, Y.; Cho, H.; Widanapathirana, L.; Zhang, S. Conformationally Controlled Oligocholate Membrane Transporters: Learning through Water Play. Acc. Chem. Res. 2013, 28, 2763–2772. [Google Scholar] [CrossRef] [Green Version]
- Le Dévédec, F.; Fuentealba, D.; Strandman, S.; Bohne, C.; Zhu, X.X. Aggregation Behavior of Pegylated Bile Acid Derivatives. Langmuir 2012, 28, 13431–13440. [Google Scholar] [CrossRef] [PubMed]
- Giguère, G.; Zhu, X.X. Functional Star Polymers with a Cholic Acid Core and Their Thermosensitive Properties. Biomacromolecules 2010, 11, 201–206. [Google Scholar] [CrossRef]
- Despa, F.; Luo, J.T.; Li, J.; Duan, Y.; Lam, K.S. Cholic Acid Micelles—Controlling the Size of the Aqueous Cavity by PEGylation. Phys. Chem. Chem. Phys. 2010, 12, 1589–1594. [Google Scholar] [CrossRef] [PubMed]
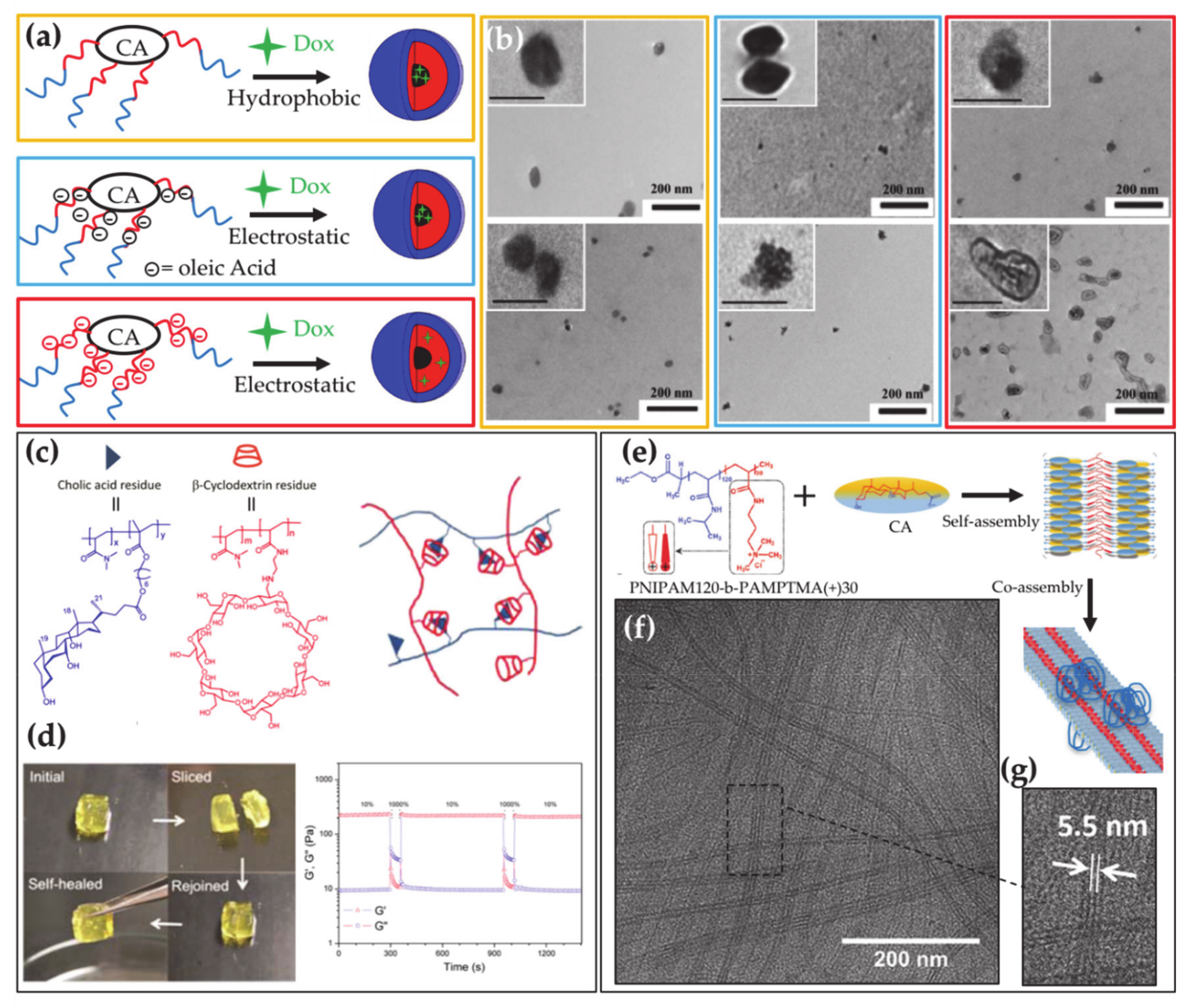
- Cunningham, A.J.; Robinson, M.; Banquy, X.; Leblond, J.; Zhu, X.X. Bile Acid-Based Drug Delivery Systems for Enhanced Doxorubicin Encapsulation: Comparing Hydrophobic and Ionic Interactions in Drug Loading and Release. Mol. Pharm. 2018, 15, 1266–1276. [Google Scholar] [CrossRef]
- Pal, S.; Ghosh Roy, S.; De, P. Synthesis via RAFT Polymerization of Thermo- and PH-Responsive Random Copolymers Containing Cholic Acid Moieties and Their Self-Assembly in Water. Polym. Chem. 2014, 5, 1275–1284. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.G.; Zhu, X.X. Self-Healing Supramolecular Hydrogel Made of Polymers Bearing Cholic Acid and β-Cyclodextrin Pendants. Chem. Mater. 2015, 27, 387–393. [Google Scholar] [CrossRef]
- Shao, Y.; Jia, Y.G.; Shi, C.; Luo, J.; Zhu, X.X. Block and Random Copolymers Bearing Cholic Acid and Oligo(Ethylene Glycol) Pendant Groups: Aggregation, Thermosensitivity, and Drug Loading. Biomacromolecules 2014, 15, 1837–1844. [Google Scholar] [CrossRef]
- Du, J.; Tang, Y.; Lewis, A.L.; Armes, S.P. PH-Sensitive Vesicles Based on a Biocompatible Zwitterionic Diblock Copolymer. J. Am. Chem. Soc. 2005, 127, 17982–17983. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Qi, J.; Yao, P. Amphiphilic Cholic-Acid-Modified Dextran Sulfate and Its Application for the Controlled Delivery of Superoxide Dismutase. Macromol. Biosci. 2012, 12, 515–524. [Google Scholar] [CrossRef]
- Ahlheirn, M.; Hallensleben, M.L. Kondensationspolymerisation von Gallensäuren Herrn. 1988, 9, 299–302. Macromol. Rapid Commun. 1988; 9, 299–302. [Google Scholar]
- Shao, Y.; Lavigueur, C.; Zhu, X.X. Multishape Memory Effect of Norbornene-Based Copolymers with Cholic Acid Pendant Groups. Macromolecules 2012, 45, 1924–1930. [Google Scholar] [CrossRef]
- Zhang, K.; Jia, Y.G.; Tsai, I.H.; Strandman, S.; Ren, L.; Hong, L.; Zhang, G.; Guan, Y.; Zhang, Y.; Zhu, X.X. “Bitter-Sweet” Polymeric Micelles Formed by Block Copolymers from Glucosamine and Cholic Acid. Biomacromolecules 2017, 18, 778–786. [Google Scholar] [CrossRef]
- Nichifor, M.; Stanciu, M.C.; Doroftei, F. Self-Assembly of Dextran–b–Deoxycholic Acid Polyester Copolymers: Copolymer Composition and Self-Assembly Procedure Tune the Aggregate Size and Morphology. Carbohydr. Polym. 2021, 252, 117147. [Google Scholar] [CrossRef] [PubMed]
- Stojančević, M.; Pavlović, N.; Goločorbin-Kon, S.; Mikov, M. Application of Bile Acids in Drug Formulation and Delivery. Front. Life Sci. 2013, 7, 112–122. [Google Scholar] [CrossRef]
- Damgé, C.P.; Maincent, P. Nanoparticles Strategies for the Oral Delivery of Insulin. Expert Opin. Drug Deliv. 2008, 5, 45–68. [Google Scholar] [CrossRef]
- Al-Mahallawi, A.M.; Abdelbary, A.A.; Aburahma, M.H. Investigating the Potential of Employing Bilosomes as a Novel Vesicular Carrier for Transdermal Delivery of Tenoxicam. Int. J. Pharm. 2015, 485, 329–340. [Google Scholar] [CrossRef]
- Mendonça, P.V.; Serra, A.C.; Silva, C.L.; Simões, S.; Coelho, J.F.J. Polymeric Bile Acid Sequestrants—Synthesis Using Conventional Methods and New Approaches Based on “Controlled”/Living Radical Polymerization. Prog. Polym. Sci. 2013, 38, 445–461. [Google Scholar] [CrossRef] [Green Version]
- Heřmánková, E.; Žák, A.; Poláková, L.; Hobzová, R.; Hromádka, R.; Širc, J. Polymeric Bile Acid Sequestrants: Review of Design, in Vitro Binding Activities, and Hypocholesterolemic Effects. Eur. J. Med. Chem. 2018, 144, 300–317. [Google Scholar] [CrossRef] [PubMed]
- Cameron, N.S.; Eisenberg, A.; Brown, G.R. Amphiphilic Block Copolymers as Bile Acid Sorbents: 1. Synthesis of Polystyrene-b-Poly(N,N,N-Trimethylammoniumethylene Acrylamide Chloride). Biomacromolecules 2002, 3, 116–123. [Google Scholar] [CrossRef]
- Cameron, N.S.; Eisenberg, A.; Brown, G.R. Amphiphilic Block Copolymers as Bile Acid Sorbents: 2. Polystyrene-b-Poly(N,N,N-Trimethylammoniumethylene Acrylamide Chloride): Self-Assembly and Application to Serum Cholesterol Reduction. Biomacromolecules 2002, 3, 124–132. [Google Scholar]
- Bayati, S.; Galantini, L.; Knudsen, K.D.; Schillén, K. Effects of Bile Salt Sodium Glycodeoxycholate on the Self-Assembly of PEO-PPO-PEO Triblock Copolymer P123 in Aqueous Solution. Langmuir 2015, 31, 13519–13527. [Google Scholar] [CrossRef] [PubMed]
- Bayati, S.; Anderberg Haglund, C.; Pavel, N.V.; Galantini, L.; Schillén, K. Interaction between Bile Salt Sodium Glycodeoxycholate and PEO-PPO-PEO Triblock Copolymers in Aqueous Solution. RSC Adv. 2016, 6, 69313–69325. [Google Scholar] [CrossRef]
- Patel, V.; Ray, D.; Bahadur, A.; Ma, J.; Aswal, V.K.; Bahadur, P. Pluronic®-Bile Salt Mixed Micelles. Colloids Surf. B. 2018, 166, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Singla, P.; Singh, O.; Chabba, S.; Aswal, V.K.; Mahajan, R.K. Sodium Deoxycholate Mediated Enhanced Solubilization and Stability of Hydrophobic Drug Clozapine in Pluronic Micelles. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2018, 191, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Tasca, E.; Giudice, A. Del; Galantini, L.; Schillén, K.; Giuliani, A.M.; Giustini, M. A Fluorescence Study of the Loading and Time Stability of Doxorubicin in Sodium Cholate/PEO-PPO-PEO Triblock Copolymer Mixed Micelles. J. Colloid Interface Sci. 2019, 540, 593–601. [Google Scholar] [CrossRef]
- Tasca, E.; Andreozzi, P.; Del Giudice, A.; Galantini, L.; Schillén, K.; Giuliani, A.M.; de los Angeles Ramirez, M.; Moya, S.E.; Giustini, M. Poloxamer/Sodium Cholate Co-Formulation for Micellar Encapsulation of Doxorubicin with High Efficiency for Intracellular Delivery: An in-Vitro Bioavailability Study. J. Colloid Interface Sci. 2020, 579, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Schillén, K.; Galantini, L.; Du, G.; Del Giudice, A.; Alfredsson, V.; Carnerup, A.M.; Pavel, N.V.; Masci, G.; Nyström, B. Block Copolymers as Bile Salt Sequestrants: Intriguing Structures Formed in a Mixture of an Oppositely Charged Amphiphilic Block Copolymer and Bile Salt. Phys. Chem. Chem. Phys. 2019, 21, 12518–12529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, G.; Del Giudice, A.; Alfredsson, V.; Carnerup, A.M.; Pavel, N.V.; Loh, W.; Masci, G.; Nyström, B.; Galantini, L.; Schillén, K. Effect of Temperature on the Association Behavior in Aqueous Mixtures of an Oppositely Charged Amphiphilic Block Copolymer and Bile Salt. Polymer 2020, 206, 122871. [Google Scholar] [CrossRef]
- di Gregorio, M.C.; Gubitosi, M.; Travaglini, L.; Pavel, N.V.; Jover, A.; Meijide, F.; Vázquez Tato, J.; Sennato, S.; Schillén, K.; Tranchini, F.; et al. Supramolecular Assembly of a Thermoresponsive Steroidal Surfactant with an Oppositely Charged Thermoresponsive Block Copolymer. Phys. Chem. Chem. Phys. 2017, 19, 1504–1515. [Google Scholar] [CrossRef] [PubMed]
- Pigliacelli, C.; Belton, P.; Wilde, P.; Qi, S. Probing the Molecular Interactions between Pharmaceutical Polymeric Carriers and Bile Salts in Simulated Gastrointestinal Fluids Using NMR Spectroscopy. J. Colloid Interface Sci. 2019, 551, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Carey, M.C.; Small, D.M. Micellar Properties of Dihydroxy and Trihydroxy Bile Salts: Effects of Counterion and Temperature. J. Colloid Interface Sci. 1969, 31, 382–396. [Google Scholar] [CrossRef]
- Li, Y.; Holzwarth, J.F.; Bohne, C. Aggregation Dynamics of Sodium Taurodeoxycholate and Sodium Deoxycholate. Langmuir 2000, 16, 2038–2041. [Google Scholar] [CrossRef]
- Kawamura, H.; Murata, Y.; Yamaguchi, T.; Igimi, H.; Tanaka, M.; Sugihara, G.; Kratohvil, J.P. Spin-Label Studies of Bile Salt Micelles. J. Phys. Chem. 1989, 93, 3321–3326. [Google Scholar] [CrossRef]
- Campanelli, A.R.; de Sanctis, S.C.; Galantini, L.; Giglio, E.; Scaramuzza, L. A Possible Helical Model for Sodium Glycocholate Micellar Aggregates. J. Incl. Phenom. Mol. Recognit. Chem. 1991, 10, 367–377. [Google Scholar] [CrossRef]
- D’Archivio, A.A.; Galantini, L.; Gavuzzo, E.; Giglio, E.; Scaramuzza, L. Possible Models for the Micellar Aggregates of Glycocholate and Taurocholate Salts from Crystal Structures, QELS, and CD Measurements. Langmuir 1996, 12, 4660–4667. [Google Scholar] [CrossRef]
- Galantini, L.; Giglio, E.; Leonelli, A.; Pavel, N.V. An Integrated Study of Small-Angle x-Ray Scattering and Dynamic Light Scattering on Cylindrical Micelles of Sodium Glycodeoxycholate. J. Phys. Chem. B 2004, 108, 3078–3085. [Google Scholar] [CrossRef]
- Galantini, L.; Giampaolo, S.M.; Mannina, L.; Pavel, N.V.; Viel, S. Study of Intermicellar Interactions and Micellar Sizes in Ionic Micelle Solutions by Comparing Collective Diffusion and Self-Diffusion Coefficients. J. Phys. Chem. B 2004, 108, 4799–4805. [Google Scholar] [CrossRef]
- D’Archivio, A.A.; Galantini, L.; Tettamanti, E. Study on Intermicellar Interactions and Micellar Size in Aqueous Solutions of Sodium Taurocholate by Measurements of Collective Diffusion and Self-Diffusion Coefficients. J. Phys. Chem. B 2000, 104, 9255–9259. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, H.; Li, X.; Song, S.; Song, A.; Hao, J. Two Gelation Mechanisms of Deoxycholate with Inorganic Additives: Hydrogen Bonding and Electrostatic Interactions. J. Phys. Chem. B 2016, 120, 6812–6818. [Google Scholar] [CrossRef]
- Li, G.; Hu, Y.; Sui, J.; Song, A.; Hao, J. Hydrogelation and Crystallization of Sodium Deoxycholate Controlled by Organic Acids. Langmuir 2016, 32, 1502–1509. [Google Scholar] [CrossRef] [PubMed]
- McNeel, K.E.; Das, S.; Siraj, N.; Negulescu, I.I.; Warner, I.M. Sodium Deoxycholate Hydrogels: Effects of Modifications on Gelation, Drug Release, and Nanotemplating. J. Phys. Chem. B 2015, 119, 8651–8659. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Feng, L.; Song, A.; Hao, J. Room-Temperature Super Hydrogel as Dye Adsorption Agent. J. Phys. Chem. B 2012, 116, 12850–12856. [Google Scholar] [CrossRef]
- Sun, X.; Xin, X.; Tang, N.; Guo, L.; Wang, L.; Xu, G. Manipulation of the Gel Behavior of Biological Surfactant Sodium Deoxycholate by Amino Acids. J. Phys. Chem. B 2014, 118, 824–832. [Google Scholar] [CrossRef] [PubMed]
- di Gregorio, M.C.; Shimon, L.J.W.; Brumfeld, V.; Houben, L.; Lahav, M.; van der Boom, M.E. Emergence of Chirality and Structural Complexity in Single Crystals at the Molecular and Morphological Levels. Nat. Commun. 2020, 11, 380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salzillo, T.; Giunchi, A.; Masino, M.; Bedoya-Martĺnez, N.; Della Valle, R.G.; Brillante, A.; Girlando, A.; Venuti, E. An Alternative Strategy to Polymorph Recognition at Work: The Emblematic Case of Coronene. Cryst. Growth Des. 2018, 18, 4869–4873. [Google Scholar] [CrossRef]
- Qiao, Y.; Lin, Y.; Wang, Y.; Yang, Z.; Liu, J.; Zhou, J.; Yan, Y.; Huang, J. Metal-Driven Hierarchical Nanohelices. Nano Lett. 2009, 9, 4500–4504. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Wang, Y.; Yang, Z.; Lin, Y.; Huang, J. Self-Templating of Metal-Driven Supramolecular Self-Assembly: A General Approach toward 1D Inorganic Nanotubes. Chem. Mater. 2011, 23, 1182–1187. [Google Scholar] [CrossRef]
- Qiao, Y.; Lin, Y.; Zhang, S.; Huang, J. Lanthanide-Containing Photoluminescent Materials: From Hybrid Hydrogel to Inorganic Nanotubes. Chem. A Eur. J. 2011, 17, 5180–5187. [Google Scholar] [CrossRef] [PubMed]
- Laishram, R.; Bhowmik, S.; Maitra, U. White Light Emitting Soft Materials from Off-the-Shelf Ingredients. J. Mater. Chem. C 2015, 3, 5885–5889. [Google Scholar] [CrossRef]
- Laishram, R.; Maitra, U. Bile Salt-Derived Eu3+ Organogel and Hydrogel: Water-Enhanced Luminescence of Eu3+ in a Gel Matrix. ChemistrySelect 2018, 3, 519–523. [Google Scholar] [CrossRef]
- Wang, H.; Song, S.; Hao, J.; Song, A. Hydrogels Triggered by Metal Ions as Precursors of Network CuS for DNA Detection. Chem. A Eur. J. 2015, 21, 12194–12201. [Google Scholar] [CrossRef]
- Shen, J.; Wang, Y.; Xin, X.; Xu, G.; Li, W.; Wang, L.; Jia, C. Studies on the Gel Behavior and Luminescence Properties of Biological Surfactant Sodium Deoxycholate/Rare-Earth Salts Mixed Systems. J. Colloid Interface Sci. 2014, 431, 82–89. [Google Scholar]
- Cheng, C.Y.; Oh, H.; Wang, T.Y.; Raghavan, S.R.; Tung, S.H. Mixtures of Lecithin and Bile Salt Can Form Highly Viscous Wormlike Micellar Solutions in Water. Langmuir 2014, 30, 10221–10230. [Google Scholar] [CrossRef]
- Markina, A.A.; Ivanov, V.A.; Komarov, P.V.; Khokhlov, A.R.; Tung, S.H. Self-Assembly of Lecithin and Bile Salt in the Presence of Inorganic Salt in Water: Mesoscale Computer Simulation. J. Phys. Chem. B 2017, 121, 7878–7888. [Google Scholar] [CrossRef]
- Cautela, J.; Giustini, M.; Pavel, N.V.; Palazzo, G.; Galantini, L. Wormlike Reverse Micelles in Lecithin/Bile Salt/Water Mixtures in Oil. Colloids Surf. A Phys. Eng. Asp. 2017, 532, 411–419. [Google Scholar] [CrossRef] [Green Version]
- Njauw, C.W.; Cheng, C.Y.; Ivanov, V.A.; Khokhlov, A.R.; Tung, S.H. Molecular Interactions between Lecithin and Bile Salts/Acids in Oils and Their Effects on Reverse Micellization. Langmuir 2013, 29, 3879–3888. [Google Scholar] [CrossRef]
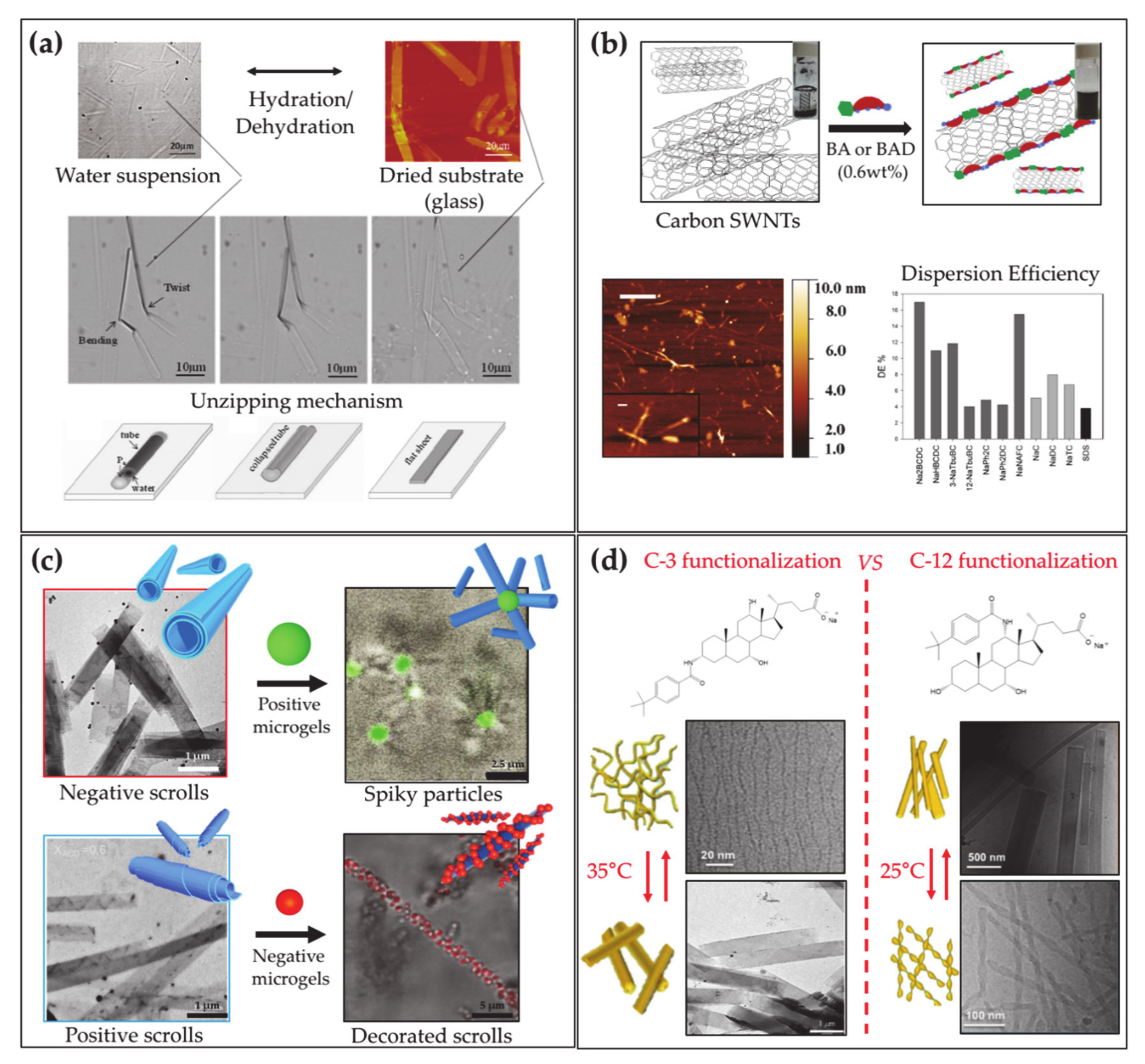
- Jean, B.; Oss-Ronen, L.; Terech, P.; Talmon, Y. Monodisperse Bile-Salt Nanotubes in Water: Kinetics of Formation. Adv. Mater. 2005, 17, 728–731. [Google Scholar] [CrossRef]
- Terech, P.; Velu, S.K.P.; Pernot, P.; Wiegart, L. Salt Effects in the Formation of Self-Assembled Lithocholate Helical Ribbons and Tubes. J. Phys. Chem. B 2012, 116, 11344–11355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zou, J.; Tamhane, K.; Kobzeff, F.F.; Fang, J. Self-Assembly of PH-Switchable Spiral Tubes: Supramolecular Chemical Springs. Small 2010, 6, 217–220. [Google Scholar] [CrossRef]
- Rhodes, S.; Liang, W.; Wang, X.; Reddy, N.R.; Fang, J. Transition from H-Aggregate Nanotubes to J-Aggregate Nanoribbons. J. Phys. Chem. C 2020, 124, 11722–11729. [Google Scholar] [CrossRef]
- Zhang, X.; Bera, T.; Liang, W.; Fang, J. Longitudinal Zipping/Unzipping of Self-Assembled Organic Tubes. J. Phys. Chem. B 2011, 115, 14445–14449. [Google Scholar] [CrossRef] [PubMed]
- D’Alagni, M.; Delfini, M.; Galantini, L.; Giglio, E. A Study of the Interaction of Bilirubin Wtth Sodium Deoxycholate in Aqueous Solutions. J. Phys. Chem. 1992, 96, 10520–10528. [Google Scholar] [CrossRef]
- D’Alagni, M.; D’Archivio, A.A.; Giglio, E.; Scaramuzza, L. Structure of Sodium and Rubidium Taurodeoxycholate Micellar Aggregates and Their Interaction Complexes with Bilirubin-IXα. J. Phys. Chem. 1994, 98, 343–353. [Google Scholar] [CrossRef]
- Meier, A.R.; Yehl, J.B.; Eckenroad, K.W.; Manley, G.A.; Strein, T.G.; Rovnyak, D. Stepwise Aggregation of Cholate and Deoxycholate Dictates the Formation and Loss of Surface-Available Chirally Selective Binding Sites. Langmuir 2018, 34, 6489–6501. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Chauhan, S. Temperature Dependent Micellar Behaviour of Sodium Cholate and Sodium Deoxycholate in the Presence of Ceftriaxone Sodium: A Physicochemical Study. J. Mol. Liq. 2020, 316, 113833. [Google Scholar] [CrossRef]
- Zhang, Q.; Ma, X.; Ward, A.; Hong, W.X.; Jaakola, V.P.; Stevens, R.C.; Finn, M.G.; Chang, G. Designing Facial Amphiphiles for the Stabilization of Integral Membrane Proteins. Angew. Chem. Int. Ed. 2007, 46, 7023–7025. [Google Scholar] [CrossRef]
- Gubitosi, M.; Trillo, J.V.; Alfaro Vargas, A.; Pavel, N.V.; Gazzoli, D.; Sennato, S.; Jover, A.; Meijide, F.; Galantini, L. Characterization of Carbon Nanotube Dispersions in Solutions of Bile Salts and Derivatives Containing Aromatic Substituents. J. Phys. Chem. B 2014, 118, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- di Gregorio, M.C.; Varenik, M.; Gubitosi, M.; Travaglini, L.; Pavel, N.V.; Jover, A.; Meijide, F.; Regev, O.; Galantini, L. Multi Stimuli Response of a Single Surfactant Presenting a Rich Self-Assembly Behavior. RSC Adv. 2015, 5, 37800–37806. [Google Scholar] [CrossRef] [Green Version]
- di Gregorio, M.C.; Pavel, N.V.; Jover, A.; Meijide, F.; Vázquez Tato, J.; Soto Tellini, V.H.; Vargas, A.A.; Regev, O.; Kasavi, Y.; Schillén, K.; et al. PH Sensitive Tubules of a Bile Acid Derivative: A Tubule Opening by Release of Wall Leaves. Phys. Chem. Chem. Phys. 2013, 15, 7560–7566. [Google Scholar] [CrossRef] [PubMed]
- Galantini, L.; Leggio, C.; Jover, A.; Meijide, F.; Pavel, N.V.; Soto Tellini, V.H.; Vázquez Tato, J.; Di Leonardo, R.; Ruocco, G. Kinetics of Formation of Supramolecular Tubules of a Sodium Cholate Derivative. Soft Matter 2009, 5, 3018–3025. [Google Scholar] [CrossRef]
- Vázquez Tato, J.; Meijide, F.; Antelo, A.; Alvarez Alcalde, M.; Jover, A.; Galantini, L.; Pavel, N.V. Supramolecular Structures Generated by a P-Tert-Butylphenylamide Derivative of Deoxycholic Acid. from Planar Sheets to Tubular Structures through Helical Ribbons. Langmuir 2010, 26, 7768–7773. [Google Scholar]
- Meijide, F.; Trillo, J.V.; de Frutos, S.; Galantini, L.; Pavel, N.V.; Soto, V.H.; Jover, A.; Vázquez Tato, J. Formation of Tubules by P-Tert-Butylphenylamide Derivatives of Chenodeoxycholic and Ursodeoxycholic Acids in Aqueous Solution. Steroids 2012, 77, 1205–1211. [Google Scholar] [CrossRef]
- Margulis-Goshen, K.; di Gregorio, M.C.; Pavel, N.V.; Abezgauz, L.; Danino, D.; Vázquez Tato, J.; Soto Tellini, V.H.; Magdassi, S.; Galantini, L. Drug-Loaded Nanoparticles and Supramolecular Nanotubes Formed from a Volatile Microemulsion with Bile Salt Derivatives. Phys. Chem. Chem. Phys. 2013, 15, 6016–6024. [Google Scholar] [CrossRef]
- Trillo, J.V.; Jover, A.; Galantini, L.; Tato, J.V.; Soto, V.H.; Meijide, F.; di Gregorio, M.C.; de Frutos, S. Self-Aggregation Mechanism of a Naphthylamide Cationic Derivative of Cholic Acid. From Fibers to Tubules. RSC Adv. 2013, 4, 5598–5606. [Google Scholar] [CrossRef]
- Travaglini, L.; D’Annibale, A.; Schillén, K.; Olsson, U.; Sennato, S.; Pavel, N.V.; Galantini, L. Amino Acid–Bile Acid Based Molecules: Extremely Narrow Surfactant Nanotubes Formed by a Phenylalanine-Substituted Cholic Acid. Chem. Commun. 2012, 48, 12011–12013. [Google Scholar] [CrossRef]
- Travaglini, L.; D’Annibale, A.; di Gregorio, M.C.; Schillén, K.; Olsson, U.; Sennato, S.; Pavel, N.V.; Galantini, L. Between Peptides and Bile Acids: Self-Assembly of Phenylalanine Substituted Cholic Acids. J. Phys. Chem. B 2013, 117, 9248–9257. [Google Scholar] [CrossRef] [PubMed]
- Travaglini, L.; Gubitosi, M.; di Gregorio, M.C.; Pavel, N.V.; D’Annibale, A.; Giustini, M.; Soto Tellini, V.H.; Vázquez Tato, J.; Obiols-Rabasa, M.; Bayati, S.; et al. On the Self-Assembly of a Tryptophan Labeled Deoxycholic Acid. Phys. Chem. Chem. Phys. 2014, 16, 19492–19504. [Google Scholar] [CrossRef] [PubMed]
- Travaglini, L.; Gubitosi, M.; di Gregorio, M.C.; D’Annibale, A.; Meijide, F.; Giustini, M.; Sennato, S.; Obiols-Rabasa, M.; Schillén, K.; Pavel, N.V.; et al. A Tryptophan-Substituted Cholic Acid: Expanding the Family of Labelled Biomolecules. Colloids Surf. A Phys. Eng. Asp. 2015, 483, 142–149. [Google Scholar] [CrossRef]
- Gubitosi, M.; Travaglini, L.; D’Annibale, A.; Pavel, N.V.; Vázquez Tato, J.; Obiols-Rabasa, M.; Sennato, S.; Olsson, U.; Schillén, K.; Galantini, L. Sugar-Bile Acid-Based Bolaamphiphiles: From Scrolls to Monodisperse Single-Walled Tubules. Langmuir 2014, 30, 6358–6366. [Google Scholar] [CrossRef] [PubMed]
- Gubitosi, M.; D’Annibale, A.; Schillén, K.; Olsson, U.; Pavel, N.V.; Galantini, L. On the Stability of Lithocholate Derivative Supramolecular Tubules. RSC Adv. 2017, 7, 512–517. [Google Scholar] [CrossRef] [Green Version]
- Soto Tellini, V.H.; Jover, A.; Meijide, F.; Vázquez Tato, J.; Galantini, L.; Pavel, N.V. Supramolecular Structures Generated by a P-Tert-Butylphenylamide Derivative of Cholic Acid: From Vesicles to Molecular Tubes. Adv. Mater. 2007, 19, 1752–1756. [Google Scholar] [CrossRef]
- Manghisi, N.; Leggio, C.; Jover, A.; Meijide, F.; Pavel, N.V.; Tellini, V.H.S.; Tato, J.V.; Agostino, R.G.; Galantini, L. Catanionic Tubules with Tunable Charge. Angew. Chem. Int. Ed. 2010, 49, 6604–6607. [Google Scholar] [CrossRef]
- di Gregorio, M.C.; Severoni, E.; Travaglini, L.; Gubitosi, M.; Sennato, S.; Mura, F.; Redondo-Gómez, C.; Jover, A.; Pavel, N.V.; Galantini, L. Bile Acid Derivative-Based Catanionic Mixtures: Versatile Tools for Superficial Charge Modulation of Supramolecular Lamellae and Nanotubes. Phys. Chem. Chem. Phys. 2018, 20, 18957–18968. [Google Scholar] [CrossRef] [PubMed]
- Gubitosi, M.; Travaglini, L.; di Gregorio, M.C.; Pavel, N.V.; Vazquez-Tato, J.; Sennato, S.; Olsson, U.; Schillén, K.; Galantini, L. Tailoring Supramolecular Nanotubes by Bile Salt Based Surfactant Mixtures. Angew. Chem. Int. Ed. 2015, 54, 7018–7021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cautela, J.; Lattanzi, V.; Månsson, L.; Galantini, L.; Crassous, J.J. Sphere–Tubule Superstructures through Supramolecular and Supracolloidal Assembly Pathways. Small 2018, 14, 1803215. [Google Scholar] [CrossRef] [PubMed]
- Gubitosi, M.; Meijide, F.; D’Annibale, A.; Vázquez Tato, J.; Jover, A.; Galantini, L.; Travaglini, L.; di Gregorio, M.C.; Pavel, N.V. Crystal Structure of a Lithium Salt of a Glucosyl Derivative of Lithocholic Acid. Steroids 2016, 113, 87–94. [Google Scholar] [CrossRef]
- Cautela, J.; Severoni, E.; Redondo-Gómez, C.; di Gregorio, M.C.; Del Giudice, A.; Sennato, S.; Angelini, R.; D’Abramo, M.; Schillén, K.; Galantini, L. Substituent Position and Orientation on the Self-Assembly of Steroid Surfactant Isomers. Colloids Surf. B. 2020, 185, 110556. [Google Scholar] [CrossRef]
- di Gregorio, M.C.; Ranjan, P.; Houben, L.; Shimon, L.J.W.; Rechav, K.; Lahav, M.; Van Der Boom, M.E. Metal-Coordination-Induced Fusion Creates Hollow Crystalline Molecular Superstructures. J. Am. Chem. Soc. 2018, 140, 9132–9139. [Google Scholar] [CrossRef] [PubMed]
- Travaglini, L.; di Gregorio, M.C.; Severoni, E.; Annibale, A.D.; Sennato, S.; Tardani, F.; Giustini, M.; Gubitosi, M.; Del, A.; Galantini, L. Deoxycholic Acid and L -Phenylalanine Enrich Their Hydrogel Properties When Combined in a Zwitterionic Derivative. J. Colloid Interface Sci. 2019, 554, 453–462. [Google Scholar] [CrossRef]
- Pore, V.S.; Agalave, S.G.; Pharande, S.G.; Patil, P.A.; Kotmale, A.S. Bile Acid Hydrazides: Gelation, Structural, Physical and Spectroscopic Properties. New J. Chem. 2015, 39, 453–460. [Google Scholar] [CrossRef]
- Travaglini, L.; Giordano, C.; D’Annibale, A.; Gubitosi, M.; di Gregorio, M.C.; Schillén, K.; Stefanucci, A.; Mollica, A.; Pavel, N.V.; Galantini, L. Twisted Nanoribbons from a RGD-Bearing Cholic Acid Derivative. Colloids Surf. B. 2017, 159, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Noponen, V.; Valkonen, A.; Lahtinen, M.; Salo, H.; Sievänen, E. Self-Assembly Properties of Bile Acid Derivatives of L-Cysteine, L-Valine and L-Serine Alkyl Esters. Supramol. Chem. 2013, 25, 133–145. [Google Scholar] [CrossRef]
- Sajisha, V.S.; Maitra, U. Remarkable Isomer-Selective Gelation of Aromatic Solvents by a Polymorph of a Urea-Linked Bile Acid– Amino Acid Conjugate. RSC Adv. 2014, 4, 43167–43171. [Google Scholar] [CrossRef]
- Maity, M.; Maitra, U. Supramolecular Gels from Conjugates of Bile Acids and Amino Acids and Their Applications. Eur. J. Org. Chem. 2017, 2017, 1713–1720. [Google Scholar] [CrossRef]
- di Gregorio, M.C.; Pavel, N.V.; Miragaya, J.; Jover, A.; Meijide, F.; Vázquez Tato, J.; Soto Tellini, V.H.; Galantini, L. Catanionic Gels Based on Cholic Acid Derivatives. Langmuir 2013, 29, 12342–12351. [Google Scholar] [CrossRef]
- Chiang, J.Y.L. Bile Acid Metabolism and Signaling. Compr. Physiol. 2013, 3, 1191–1212. [Google Scholar] [PubMed] [Green Version]
- De Aguiar Vallim, T.Q.; Tarling, E.J.; Edwards, P.A. Pleiotropic Roles of Bile Acids in Metabolism. Cell Metab. 2013, 17, 657–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahlström, A.; Sayin, S.I.; Marschall, H.U.; Bäckhed, F. Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism. Cell Metab. 2016, 24, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zhou, A.; Tang, L.; Lei, Y.; Tang, B.; Zhang, L. Bile Acids: Key Regulators and Novel Treatment Targets for Type 2 Diabetes. J. Diabetes Res. 2020, 2020, 6138438. [Google Scholar] [CrossRef] [PubMed]
- Grant, S.M.; Demorrow, S. Bile Acid Signaling in Neurodegenerative and Neurological Disorders. Int. J. Mol. Sci. 2020, 21, 5982. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
di Gregorio, M.C.; Cautela, J.; Galantini, L. Physiology and Physical Chemistry of Bile Acids. Int. J. Mol. Sci. 2021, 22, 1780. https://doi.org/10.3390/ijms22041780
di Gregorio MC, Cautela J, Galantini L. Physiology and Physical Chemistry of Bile Acids. International Journal of Molecular Sciences. 2021; 22(4):1780. https://doi.org/10.3390/ijms22041780
Chicago/Turabian Styledi Gregorio, Maria Chiara, Jacopo Cautela, and Luciano Galantini. 2021. "Physiology and Physical Chemistry of Bile Acids" International Journal of Molecular Sciences 22, no. 4: 1780. https://doi.org/10.3390/ijms22041780
APA Styledi Gregorio, M. C., Cautela, J., & Galantini, L. (2021). Physiology and Physical Chemistry of Bile Acids. International Journal of Molecular Sciences, 22(4), 1780. https://doi.org/10.3390/ijms22041780