
The Cutinase Bdo_10846 Play an Important Role in the Virulence of Botryosphaeria dothidea and in Inducing the Wart Symptom on Apple Plant


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
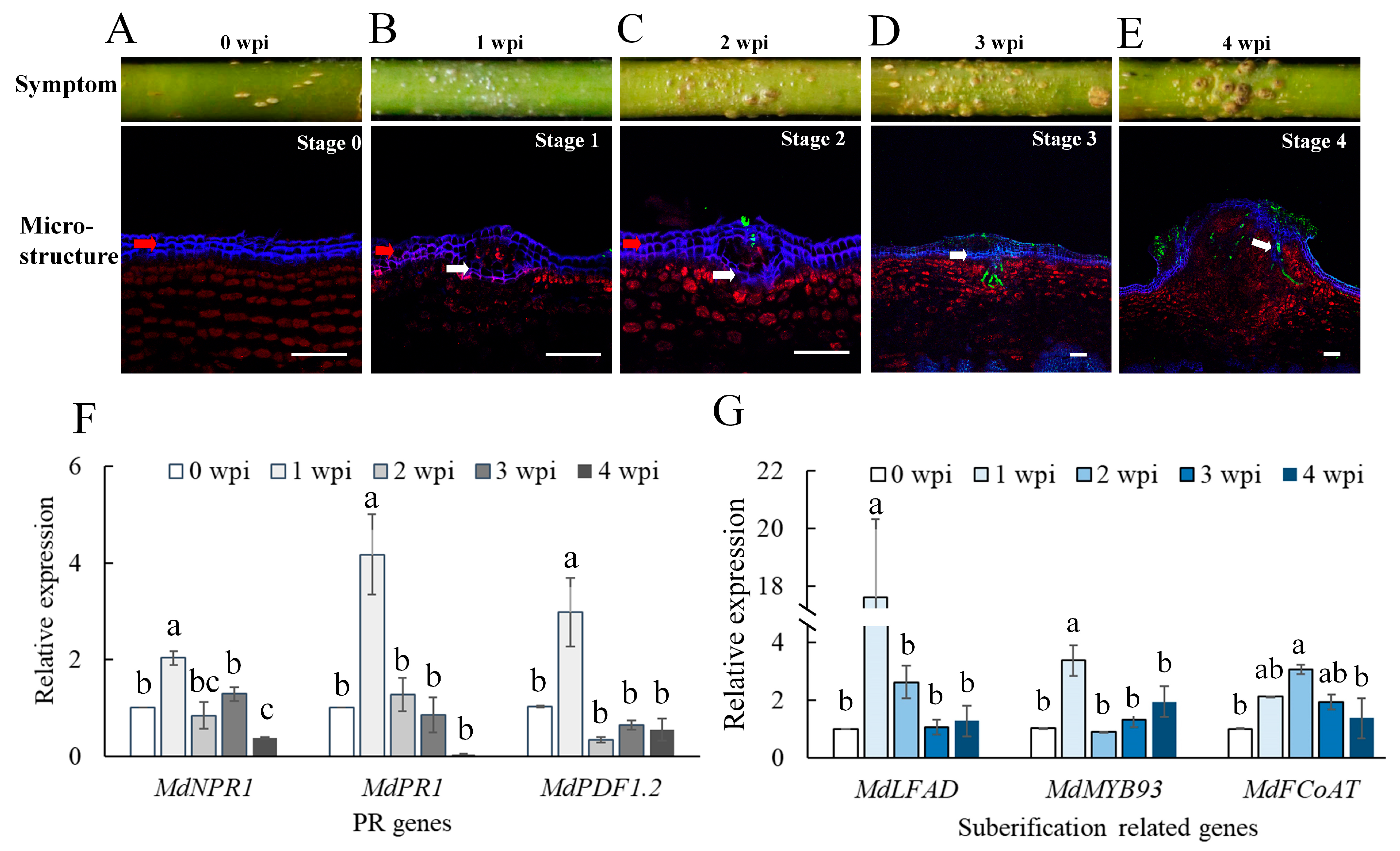
2.1. Invasion of B. dothidea Stimulated the Plant Defense Response and Wart Formation
2.2. Cutinase Genes in B. dothidea Were Upregulated during Infection of Apple Shoots
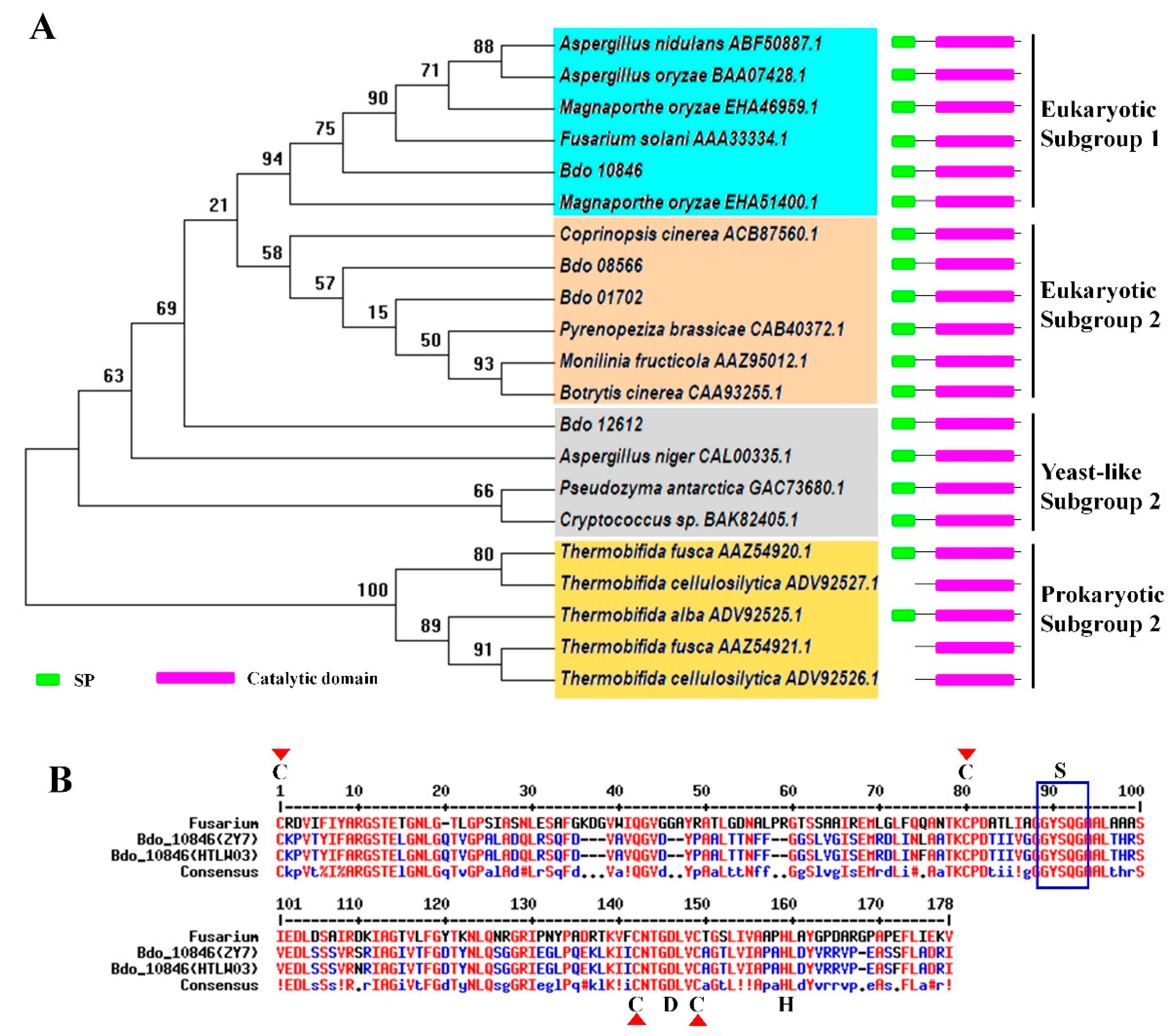
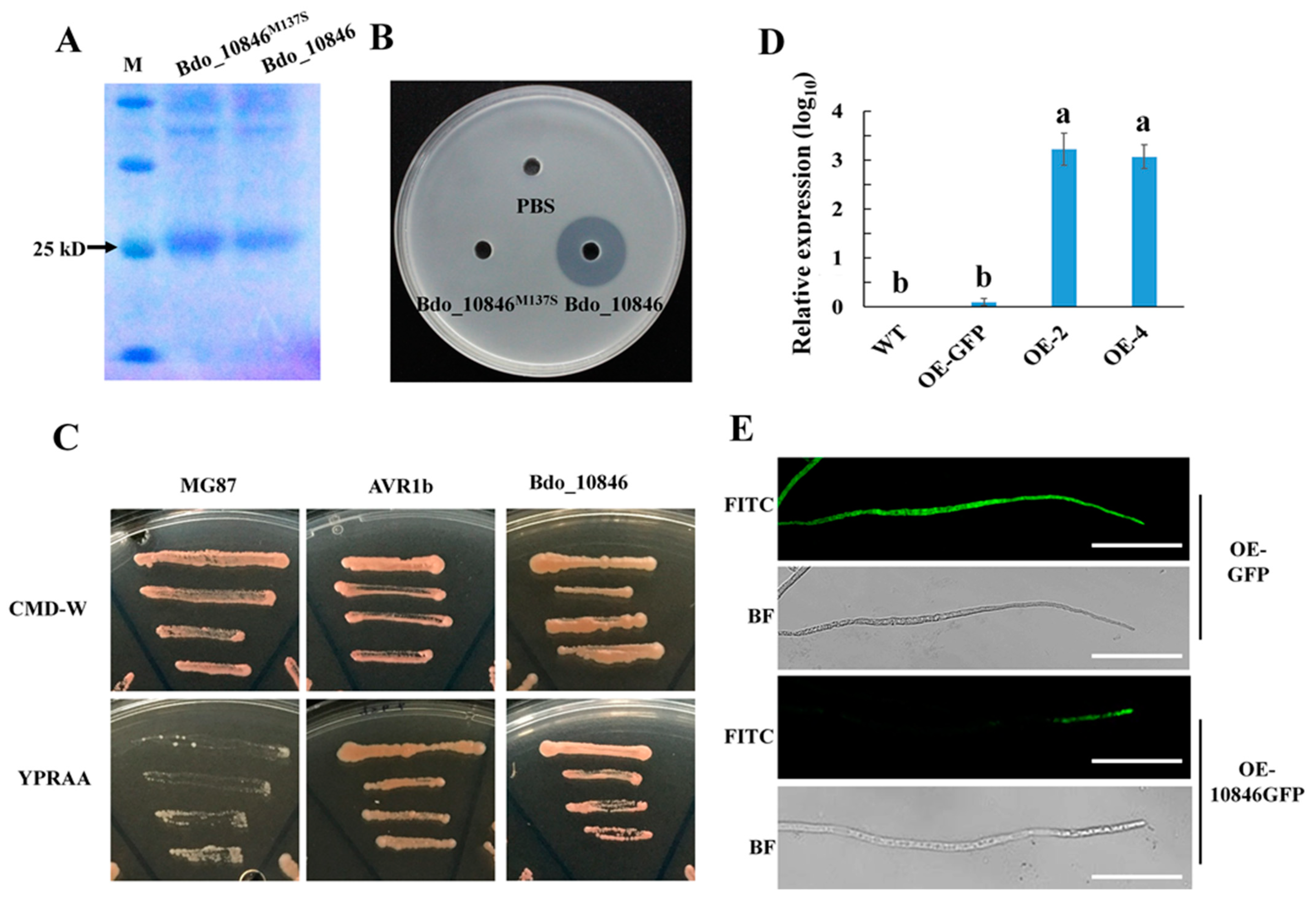
2.3. Enzyme Activity, Signal Peptide Examination and Localization of Bdo_10846
2.4. Deleting Bdo_10846 Reduced the B. dothidea Cutinase Activity
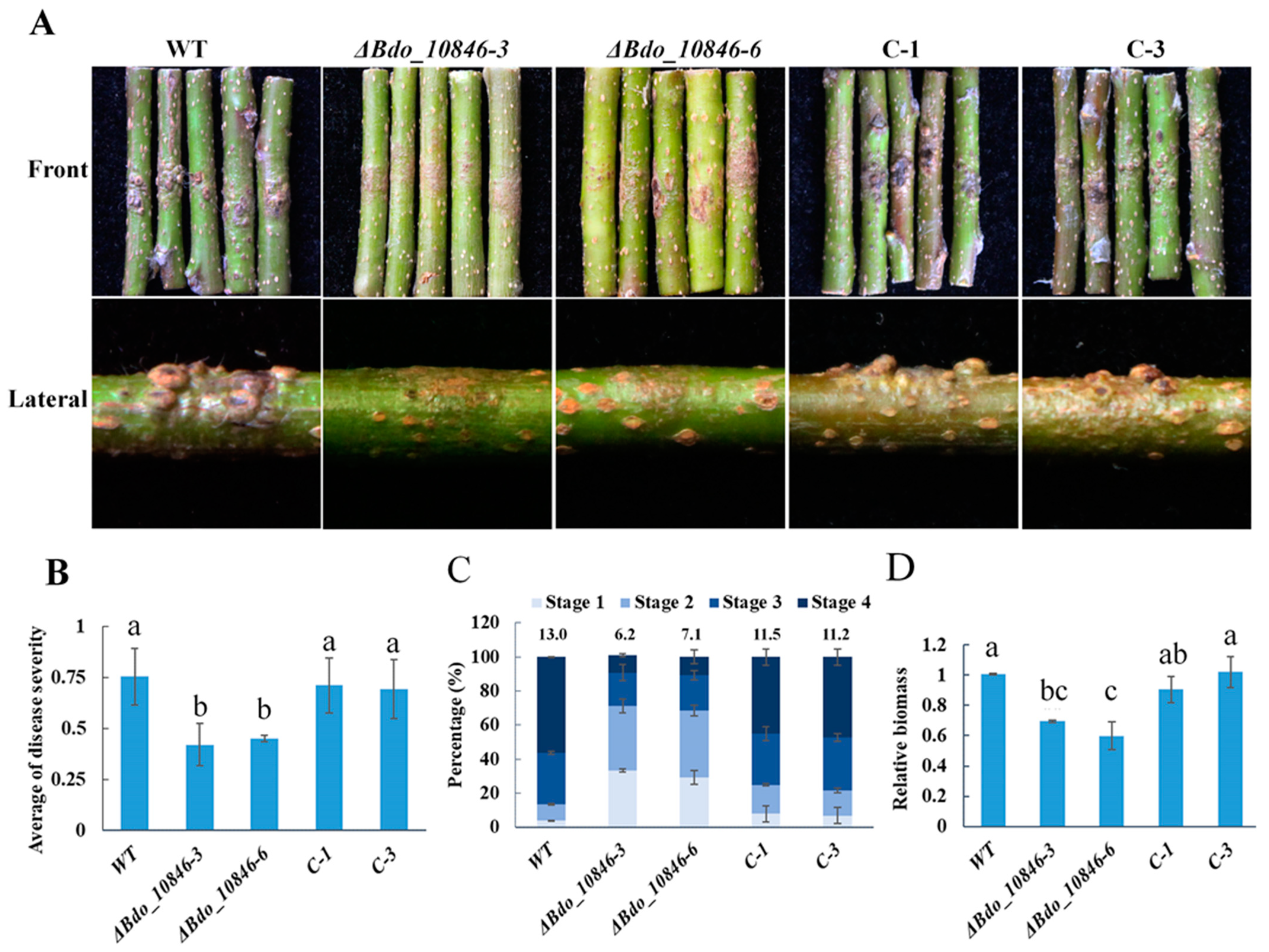
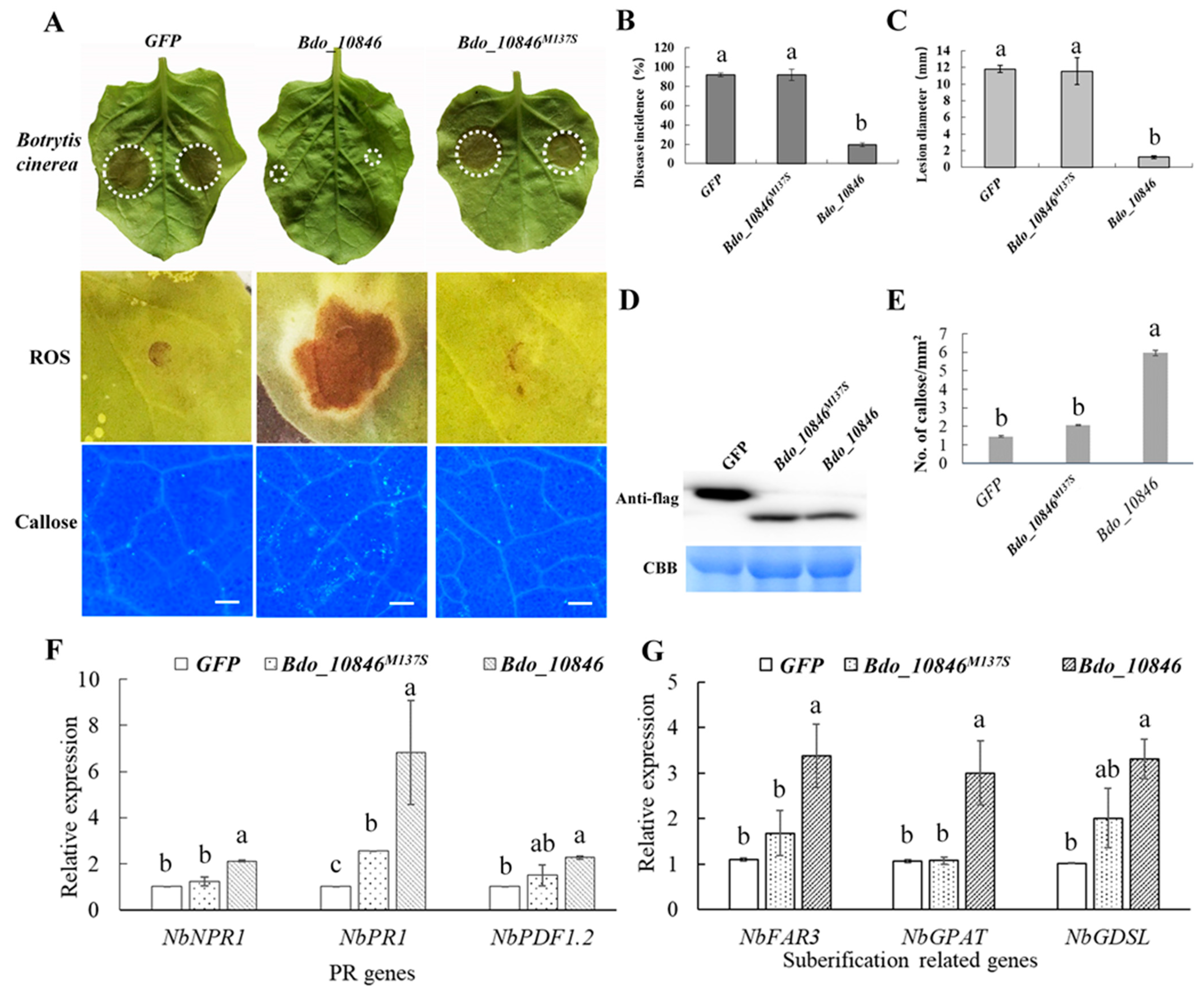
2.5. Bdo_10846 Is Required for the Full Virulence of B. dothidea
2.6. Bdo_10846 Can Trigger Plant Defense Responses
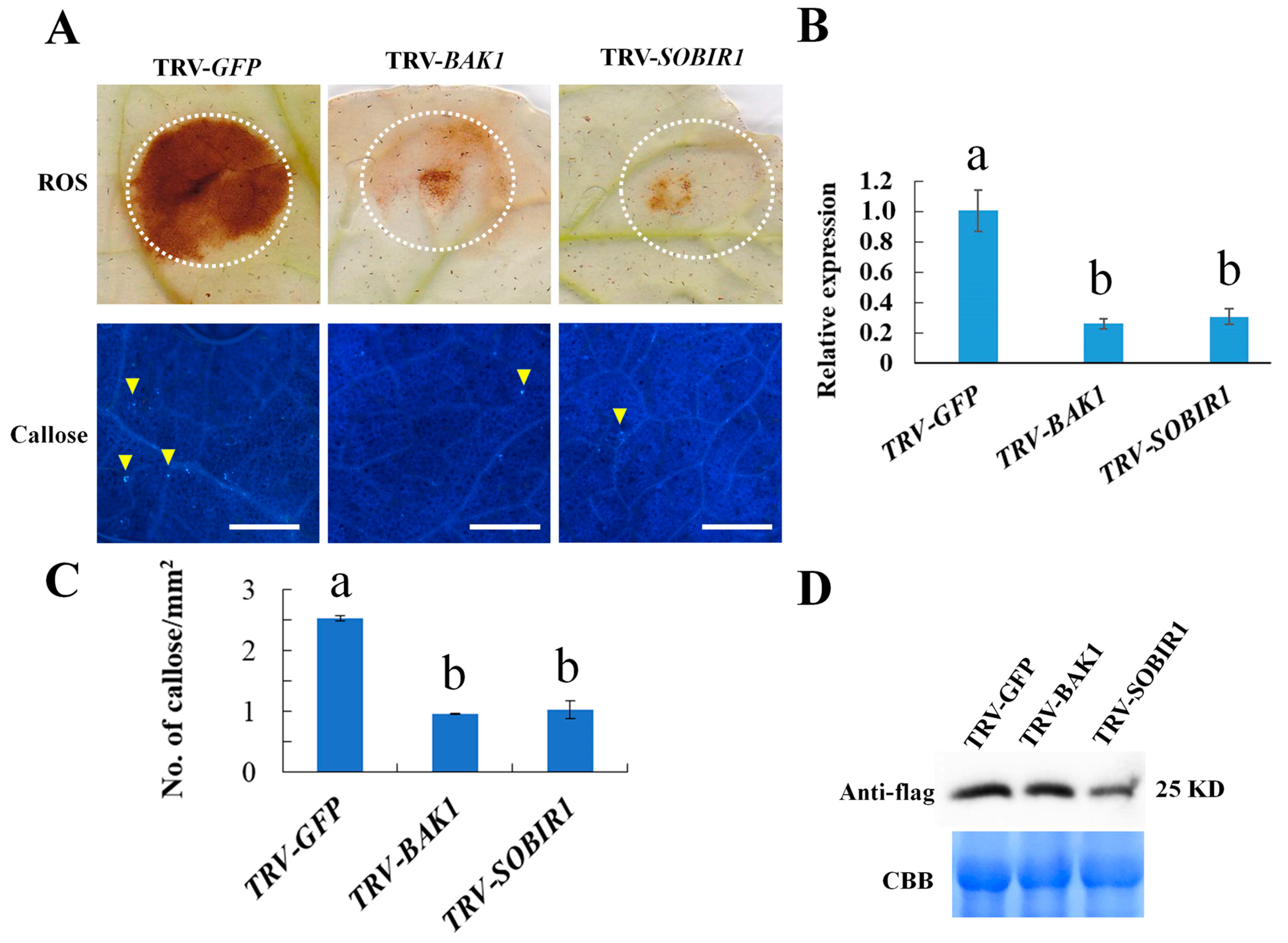
2.7. NbBAK1 and NbSOBIR1 Are Required for Bdo_10846 Induced Defense Responses
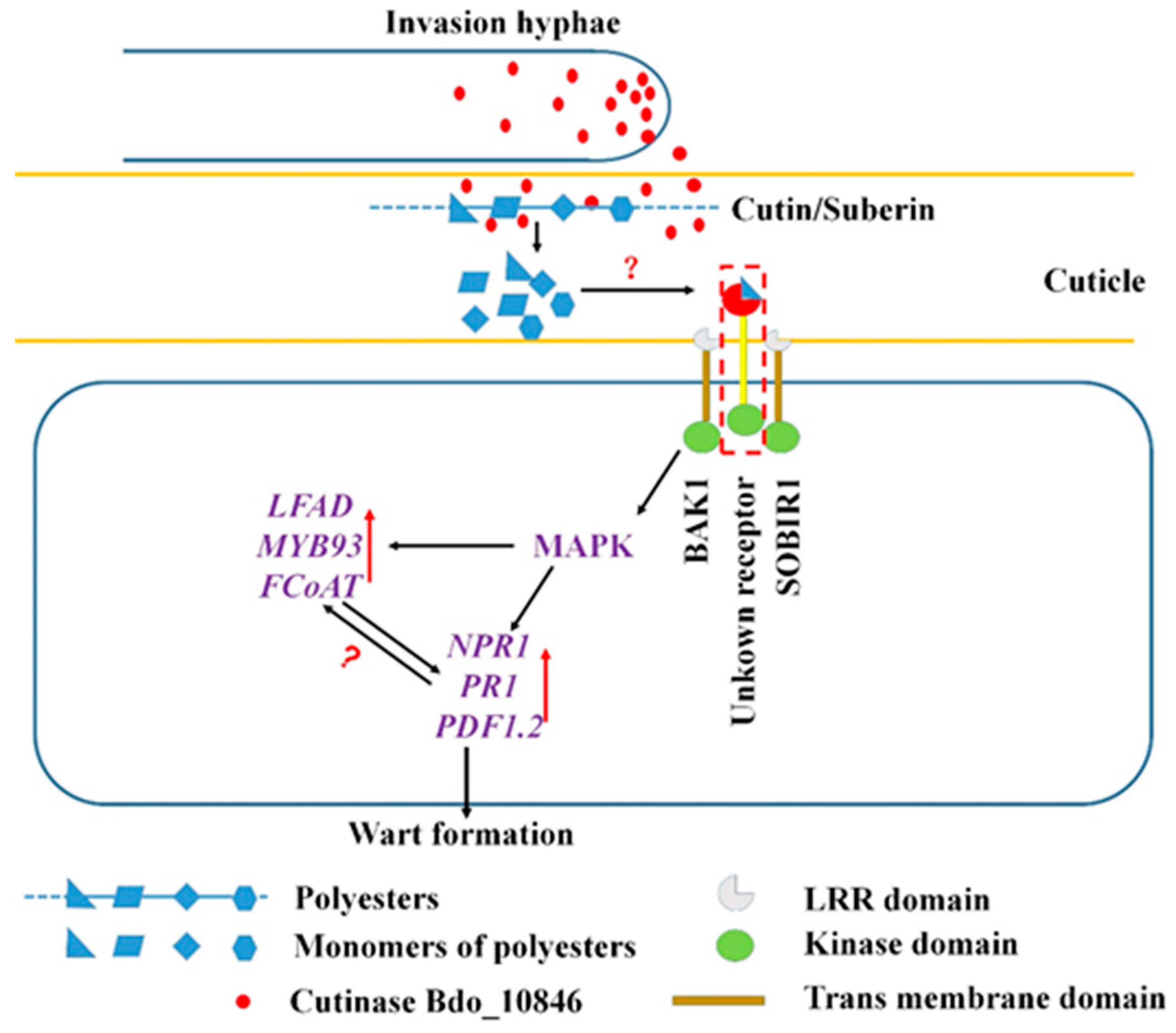
3. Discussion
4. Materials and Methods
4.1. Isolates Used and Culture Conditions
4.2. Analysis of Cutinase Protein and Gene Sequences
4.3. RNA Extraction and qRT-PCR
4.4. Plasmid Construction
4.5. Yeast Secretion Trap Assay
4.6. Pathogenicity Test
4.7. Histochemical Analysis
4.8. Prokaryotic Expression of Bdo_10846
4.9. Cutinase Activity Assay
4.10. Virus-Induced Gene Silencing (VIGS)
4.11. Transient Expression Assay
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marsberg, A.; Kemler, M.; Jami, F.; Nagel, J.H.; Postma-Smidt, A.; Naidoo, S.; Wingfield, M.J.; Crous, P.W.; Spatafora, J.; Hesse, C.N. Botryosphaeria dothidea: A talent pathogen of global importance to woody plant health. Mol. Plant Pathol. 2017, 18, 477–488. [Google Scholar] [CrossRef]
- Milholland, R.D. Occurrence of a New Race of Botryosphaeria corticis on Highbush and Rabbiteye Blueberry. Plant Dis. 1984, 68, 522–523. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Rumbos, I.C.; Alves, A.; Correia, A. Morphology and phylogeny of Botryosphaeria dothidea causing fruit rot of olives. Mycopathology 2005, 159, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Pitt, W.M.; Sosnowski, M.R.; Huang, R.; Qiu, Y.; Steel, C.C.; Savocchia, S. Evaluation of Fungicides for the Management of Botryosphaeria Canker of Grapevines. Plant Dis. 2012, 96, 1303–1308. [Google Scholar] [CrossRef]
- Yu, L.; Chen, X.L.; Gao, L.L.; Chen, H.R.; Huang, Q. First Report of Botryosphaeria dothidea Causing Canker and Shoot Blight of Eucalyptus in China. Plant Dis. 2009, 93, 764. [Google Scholar] [CrossRef]
- Tang, W.; Ding, Z.; Zhou, Z.Q.; Wang, Y.Z.; Guo, L.Y. Phylogenetic and Pathogenic Analyses Show That the Causal Agent of Apple Ring Rot in China Is Botryosphaeria dothidea. Plant Dis. 2012, 96, 486–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bargel, H.; Cerman, Z.; Koch, K.; Neinhuis, C. Evans Review No. 3: Structure–function relationships of the plant cuticle and cuticular waxes—A smart material. Funct. Plant Biol. 2006, 33, 893–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heredia, A. Biophysical and biochemical characteristics of cutin, a plant barrier biopolymer. Biochim. Biophys. Acta 2003, 1620, 1–7. [Google Scholar] [CrossRef]
- Pollard, M.; Beisson, F.; Li, Y.; Ohlrogge, J.B. Building lipid barriers: Biosynthesis of cutin and suberin. Trends Plant Sci. 2008, 13, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Franke, R.; Briesen, I.; Wojciechowski, T.; Faust, A.; Yephremov, A.; Nawrath, C.; Schreiber, L. Apoplastic polyesters in Arabidopsis surface tissues—A typical suberin and a particular cutin. Phytochemistry 2005, 66, 2643–2658. [Google Scholar] [CrossRef] [Green Version]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; Debono, A.; Durrett, T.P.; et al. Acyl-Lipid Metabolism. Arab. Book 2010, 8, e0133. [Google Scholar] [CrossRef] [Green Version]
- Molina, I.; Bonaventure, G.; Ohlrogge, J.; Pollard, M. The lipid polyester composition of Arabidopsis thaliana and Brassica napus seeds. Phytochemistry 2006, 67, 2597–2610. [Google Scholar] [CrossRef]
- Dean, B.B.; Kolattukudy, P.E. Synthesis of Suberin during Wound-healing in Jade Leaves, Tomato Fruit, and Bean Pods. Plant Physiol. 1976, 58, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Lashbrooke, J.; Cohen, H.; Levy-Samocha, D.; Tzfadia, O.; Panizel, I.; Zeisler, V.; Massalha, H.; Stern, A.; Trainotti, L.; Schreiber, L.; et al. MYB107 and MYB9 Homologs Regulate Suberin Deposition in Angiosperms. Plant Cell 2016, 28, 2097–2116. [Google Scholar] [CrossRef] [Green Version]
- Legay, S.; Guerriero, G.; André, C.; Guignard, C.; Cocco, E.; Charton, S.; Boutry, M.; Rowland, O.; Hausman, J.-F. MdMyb93 is a regulator of suberin deposition in russeted apple fruit skins. New Phytol. 2016, 212, 977–991. [Google Scholar] [CrossRef]
- Legay, S.; Guerriero, G.; Deleruelle, A.; Lateur, M.; Evers, D.; André, C.M.; Hausman, J.-F. Apple russeting as seen through the RNA-seq lens: Strong alterations in the exocarp cell wall. Plant Mol. Biol. 2015, 88, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Verdaguer, R.; Soler, M.; Serra, O.; Garrote, A.; Fernández, S.; Company-Arumí, D.; Anticó, E.; Molinas, M.; Figueras, M. Silencing of the potato StNAC103 gene enhances the accumulation of suberin polyester and associated wax in tuber skin. J. Exp. Bot. 2016, 67, 5415–5427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Su, L.; Chen, J.; Wu, J. Cutinase: Characteristics, preparation, and application. Biotechnol. Adv. 2013, 31, 1754–1767. [Google Scholar] [CrossRef] [PubMed]
- Petersen, S.B.; Drabløs, F.; Petersen, M.T.N.; Petersen, E.I. [3] Identification of important motifs in protein sequences: Program MULTIM and its applications to lipase-related sequences. Methods Enzymol. 1997, 284, 61–85. [Google Scholar] [CrossRef]
- Davies, K.; De Lorono, I.; Foster, S.; Li, D.; Johnstone, K.; Ashby, A. Evidence for a role of cutinase in pathogenicity of Pyrenopeziza brassicae on brassicas. Physiol. Mol. Plant Pathol. 2000, 57, 63–75. [Google Scholar] [CrossRef]
- Kwon, M.-A.; Kim, H.S.; Yang, T.H.; Song, B.K.; Song, J.K. High-level expression and characterization of Fusarium solani cutinase in Pichia pastoris. Protein Expr. Purif. 2009, 68, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Wanga, G.-Y.; Michailides, T.J.; Hammock, B.D.; Leec, Y.-M.; Bostock, R.M. Affinity Purification and Characterization of a Cutinase from the Fungal Plant Pathogen Monilinia fructicola (Wint.) Honey. Arch. Biochem. Biophys. 2000, 382, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Sirakova, T.; Rogers, L.; Ettinger, W.F.; Kolattukudy, P.E. Regulation of constitutively expressed and induced cu-tinase genes by different zinc finger transcription factors in Fusarium solani f. sp. pisi (Nectria haematococca). J Biol. Chem. 2002, 277, 7905–7912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Chen, J.; Li, D.-W.; Zheng, L.; Huang, J. CglCUT1 gene required for cutinase activity and pathogenicity of Colletotrichum gloeosporioides causing anthracnose of Camellia oleifera. Eur. J. Plant Pathol. 2016, 147, 103–114. [Google Scholar] [CrossRef]
- Lee, M.-H.; Chiu, C.-M.; Roubtsova, T.; Chou, C.-M.; Bostock, R.M. Overexpression of a Redox-Regulated Cutinase Gene, MfCUT1, Increases Virulence of the Brown Rot Pathogen Monilinia fructicola on Prunus spp. Mol. Plant-Microbe Interactions 2010, 23, 176–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Hou, J.; Wang, Y.; Jin, Y.; Borth, W.; Zhao, F.; Liu, Z.; Hu, J.; Zuo, Y. Genome-wide identification, classification and expression analysis in fungal–plant interactions of cutinase gene family and functional analysis of a putative ClCUT7 in Curvularia lunata. Mol. Genet. Genom. 2016, 291, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Skamnioti, P.; Gurr, S.J. Magnaporthe grisea cutinase2 eediates appressorium differentiation and host penetration and is required for full virulence. Plant Cell. 2007, 19, 2674–2689. [Google Scholar] [CrossRef] [Green Version]
- Auyong, A.S.M.; Ford, R.; Taylor, P.W.J. The role of cutinase and its impact on pathogenicity of Colletotrichum truncatum. J. Plant Pathol. Microbe 2016, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nat. Cell Biol. 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Hou, S.; Wang, X.; Chen, D.; Yang, X.; Wang, M.; Turrà, D.; Di Pietro, A.; Zhang, W. The Secreted Peptide PIP1 Amplifies Immunity through Receptor-Like Kinase 7. PLOS Pathog. 2014, 10, e1004331. [Google Scholar] [CrossRef] [Green Version]
- Hou, S.; Yang, Y.; Wu, D.; Zhang, C. Plant immunity: Evolutinary insights from PBS1, Pto, and RIN4. Plant Signal. Behav. 2014, 6, 794–799. [Google Scholar] [CrossRef] [Green Version]
- Ranf, S. Sensing of molecular patterns through cell surface immune receptors. Curr. Opin. Plant Biol. 2017, 38, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C. Plant pattern-recognition receptors. Trends Immunol. 2014, 35, 345–351. [Google Scholar] [CrossRef]
- Chassot, C.; Nawrath, C.; Métraux, J.-P. Cuticular defects lead to full immunity to a major plant pathogen. Plant J. 2007, 49, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Gui, Y.-J.; Zhang, W.-Q.; Zhang, D.-D.; Zhou, L.; Short, D.P.G.; Wang, J.; Ma, X.-F.; Li, T.-G.; Kong, Z.-Q.; Wang, B.-L.; et al. A Verticillium dahliae Extracellular Cutinase Modulates Plant Immune Responses. Mol. Plant-Microbe Interactions 2018, 31, 260–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Liang, X.; Gleason, M.L.; Zhang, R.; Sun, G. Comparative genomics of Botryosphaeria dothidea and B. kuwatsukai, causal agents of apple ring rot, reveals both species expansion of pathogenicity-related genes and variations in virulence gene content during speciation. IMA Fungus 2018, 9, 243–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.Y.; Zhao, W.S.; Chen, Z.; Xing, Q.K.; Hang, Q.G.; Chethana, K.W.T.; Xue, M.F.; Xu, J.P.; Phillips, A.J.L.; Wang, Y.; et al. Comparative genome and transcriptome analyses reveal adaptations to opportunistic infections in woody plant degrading pathogens of Botryosphaeriaceae. DNA Res. 2018, 25, 87–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Z. Study of the infection process of Botryosphaeria dothidea on apple shoots and identification of a mycovirus infect B. dothidea. Ph.D. Thesis, China Agricultural University, Beijing, China, 2018. (In Chinese). [Google Scholar]
- Dong, B.-Z.; Guo, L.-Y. An efficient gene disruption method for the woody plant pathogen Botryosphaeria dothidea. BMC Biotechnol. 2020, 20, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Chassot, C.; Nawrath, C.; Métraux, J.P. The cuticle: Not only a barrier for plant defence: A novel defence syndrome in plants with cuticular defects. Plant Signal. Behav. 2008, 3, 142–144. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.-J.; Wang, S.-X.; Liang, Y.-N.; Wen, S.-H.; Dong, B.-Z.; Ding, Z.; Guo, L.-Y.; Zhu, X.-Q. Candidate Effectors from Botryosphaeria dothidea Suppress Plant Immunity and Contribute to Virulence. Int. J. Mol. Sci. 2021, 22, 552. [Google Scholar] [CrossRef]
- Campilho, A.; Nieminen, K.; Ragni, L. The development of the periderm: The final frontier between a plant and its envi-ronment. Curr. Opin. Plant Bio. 2020, 53, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Castro-Ochoa, D.; Peña-Montes, C.; González-Canto, A.; Alva-Gasca, A.; Esquivel-Bautista, R.; Navarro-Ocaña, A.; Farrés, A. ANCUT2, an Extracellular Cutinase from Aspergillus nidulans Induced by Olive Oil. Appl. Biochem. Biotechnol. 2012, 166, 1275–1290. [Google Scholar] [CrossRef]
- Ben Ali, Y.; Verger, R.; Abousalham, A. Lipases or Esterases: Does It Really Matter? Toward a New Bio-Physico-Chemical Classification. Methods Mol. Biol. 2012, 861, 31–51. [Google Scholar] [CrossRef]
- Sharma, R.; Chisti, Y.; Banerjee, U.C. Production, purification, characterization, and applications of lipases. Biotechnol. Adv. 2001, 19, 627–662. [Google Scholar] [CrossRef] [Green Version]
- Skamnioti, P.; Gurr, S.J. Cutinase and hydrophobin interplay. Plant Signal. Behav. 2008, 3, 248–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Wu, Q.; Cao, S.; Zhao, T.; Chen, L.; Zhuang, P.; Zhou, X.; Gao, Z. A novel protein elicitor (SsCut) from Sclerotinia sclerotiorum induces multiple defense responses in plants. Plant Mol. Biol. 2014, 86, 495–511. [Google Scholar] [CrossRef] [PubMed]
- Liebrand, T.W.; Burg, H.A.V.D.; Joosten, M.H. Two for all: Receptor-associated kinases SOBIR1 and BAK1. Trends Plant Sci. 2014, 19, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Postel, S.; Küfner, I.; Beuter, C.; Mazzotta, S.; Schwedt, A.; Borlotti, A.; Halter, T.; Kemmerling, B.; Nürnberger, T. The multifunctional leucine-rich repeat receptor kinase BAK1 is implicated in Arabidopsis development and immunity. Eur. J. Cell Biol. 2010, 89, 169–174. [Google Scholar] [CrossRef]
- Blanc, C.; Coluccia, F.; L’Haridon, F.; Torres, M.; Ortiz-Berrocal, M.; Stahl, E.; Reymond, P.; Schreiber, L.; Nawrath, C.; Métraux, J.-P.; et al. The Cuticle Mutant eca2 Modifies Plant Defense Responses to Biotrophic and Necrotrophic Pathogens and Herbivory Insects. Mol. Plant-Microbe Interactions 2018, 31, 344–355. [Google Scholar] [CrossRef] [Green Version]
- Mantas, S.; Davidsson, P.r.R.; Ville, P.; Tarja, K.; Martin, B.; Nina, S.; Pekka, H.; Palva, E.T. Peroxidase-generated apo-plastic ROS impair cuticle integrity and contribute to DAMP-elicited defenses. Front. Plant Sci. 2016, 7, 1945. [Google Scholar]
- Kauss, H.; Fauth, M.; Merten, A.; Jeblick, W. Cucumber Hypocotyls Respond to Cutin Monomers via Both an Inducible and a Constitutive H2O2-Generating System. Plant Physiol. 1999, 120, 1175–1182. [Google Scholar] [CrossRef] [Green Version]
- Schweizer, P.; Felix, G.; Buchala, A.; Müller, C.; Métraux, J.-P. Perception of free cutin monomers by plant cells. Plant J. 1996, 10, 331–341. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, H.; Wang, C.; Xu, J.-R. Correction: Comparative analysis of fungal genomes reveals different plant cell wall degrading capacity in fungi. BMC Genom. 2014, 15, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhang, X.; Liu, L.; Xiang, M.; Wang, W.; Sun, X.; Che, Y.; Guo, L.; Liu, G.; Guo, L.; et al. Genomic and transcriptomic analysis of the endophytic fungus Pestalotiopsis fici reveals its lifestyle and high potential for synthesis of natural products. BMC Genom. 2015, 16, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Copes, W.E.; Hendrix, F.F. Effect of temperature on sporulation of Botryosphaeria dothidea, B. obtusa, and B. rhodina. Plant Dis. 2004, 88, 292–296. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Hamari, Z.; Han, K.H.; Seo, J.A.; Reyes-Domínguez, Y.; Scazzocchio, C. Double-joint PCR: A PCR-based mo-lecular tool for gene manipulations in filamentous fungi. Fungal Genet. Biol. 2004, 41, 973–981. [Google Scholar] [CrossRef]
- A Jacobs, K.; A Collins-Racie, L.; Colbert, M.; Duckett, M.; Golden-Fleet, M.; Kelleher, K.; Kriz, R.; LaVallie, E.R.; Merberg, D.; Spaulding, V.; et al. A genetic selection for isolating cDNAs encoding secreted proteins. Gene 1997, 198, 289–296. [Google Scholar] [CrossRef]
- Liu, S.-Y.; Chen, J.-Y.; Wang, J.-L.; Li, L.; Xiao, H.-L.; Adam, S.M.; Dai, X.-F. Molecular characterization and functional analysis of a specific secreted protein from highly virulent defoliating Verticillium dahliae. Gene 2013, 529, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.; Zhang, H.T.; Zhou, Z.Q.; Wang, L.; Yan, Z.L.; Wang, S.R. Evaluation of resistance to apple ring rot in Malus germplasms (In Chinese). Acta Hort. 2018, 44, 1559–1568. [Google Scholar]
- Zhang, Y.; Wang, X.; Rong, W.; Yan, Z.; Li, Z.; Wu, L.; Zhang, G.; Ma, Z. Histochemical Analyses Reveal That Stronger Intrinsic Defenses in Gossypium barbadense Than in G. hirsutum Are Associated With Resistance to Verticillium dahliae. Mol. Plant-Microbe Interactions 2017, 30, 984–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inch, S.; Ploetz, R.; Held, B.; Blanchette, R. Histological and anatomical responses in avocado, Persea americana, induced by the vascular wilt pathogen, Raffaelea lauricola. Bot. 2012, 90, 627–635. [Google Scholar] [CrossRef] [Green Version]
- Thomas, R.; Fang, X.; Ranathunge, K.; Anderson, T.R.; Peterson, C.A.; Bernards, M.A. Soybean root suberin: Anatomical distribution, chemical compsiton, and relationship to partial resisitance to Phytophtora sojae. Plant Physiol. 2007, 144, 299–311. [Google Scholar] [CrossRef] [Green Version]
- Rao, T.; Lubin, J.W.; Armstrong, G.S.; Tucey, T.M.; Lundblad, V.; Wuttke, D.S. Structure of Est3 reveals a bimodal surface with differential roles in telomere replication. Proc. Natl. Acad. Sci. USA 2013, 111, 214–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayward, A.; Padmanabhan, M.; Dinesh-Kumar, S.P. Virus-Induced Gene Silencing in Nicotiana benthamiana and Other Plant Species. Methods Mol. Biol. 2011, 678, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Piisilä, M.; A Keceli, M.; Brader, G.; Jakobson, L.; Jõesaar, I.; Sipari, N.; Kollist, H.; Palva, E.T.; Kariola, T. The F-box protein MAX2 contributes to resistance to bacterial phytopathogens in Arabidopsis thaliana. BMC Plant Biol. 2015, 15, 53. [Google Scholar] [CrossRef] [Green Version]
- Schenk, S.T.; Hernández-Reyes, C.; Samans, B.; Stein, E.; Neumann, C.; Schikora, M.; Reichelt, M.; Mithöfer, A.; Becker, A.; Kogel, K.-H.; et al. N-Acyl-Homoserine Lactone Primes Plants for Cell Wall Reinforcement and Induces Resistance to Bacterial Pathogens via the Salicylic Acid/Oxylipin Pathway. Plant Cell 2014, 26, 2708–2723. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, B.-Z.; Zhu, X.-Q.; Fan, J.; Guo, L.-Y. The Cutinase Bdo_10846 Play an Important Role in the Virulence of Botryosphaeria dothidea and in Inducing the Wart Symptom on Apple Plant. Int. J. Mol. Sci. 2021, 22, 1910. https://doi.org/10.3390/ijms22041910
Dong B-Z, Zhu X-Q, Fan J, Guo L-Y. The Cutinase Bdo_10846 Play an Important Role in the Virulence of Botryosphaeria dothidea and in Inducing the Wart Symptom on Apple Plant. International Journal of Molecular Sciences. 2021; 22(4):1910. https://doi.org/10.3390/ijms22041910
Chicago/Turabian StyleDong, Bao-Zhu, Xiao-Qiong Zhu, Jun Fan, and Li-Yun Guo. 2021. "The Cutinase Bdo_10846 Play an Important Role in the Virulence of Botryosphaeria dothidea and in Inducing the Wart Symptom on Apple Plant" International Journal of Molecular Sciences 22, no. 4: 1910. https://doi.org/10.3390/ijms22041910
APA StyleDong, B. -Z., Zhu, X. -Q., Fan, J., & Guo, L. -Y. (2021). The Cutinase Bdo_10846 Play an Important Role in the Virulence of Botryosphaeria dothidea and in Inducing the Wart Symptom on Apple Plant. International Journal of Molecular Sciences, 22(4), 1910. https://doi.org/10.3390/ijms22041910