
An Intermediate Concentration of Calcium with Antioxidant Supplement in Culture Medium Enhances Proliferation and Decreases the Aging of Bone Marrow Mesenchymal Stem Cells

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Results
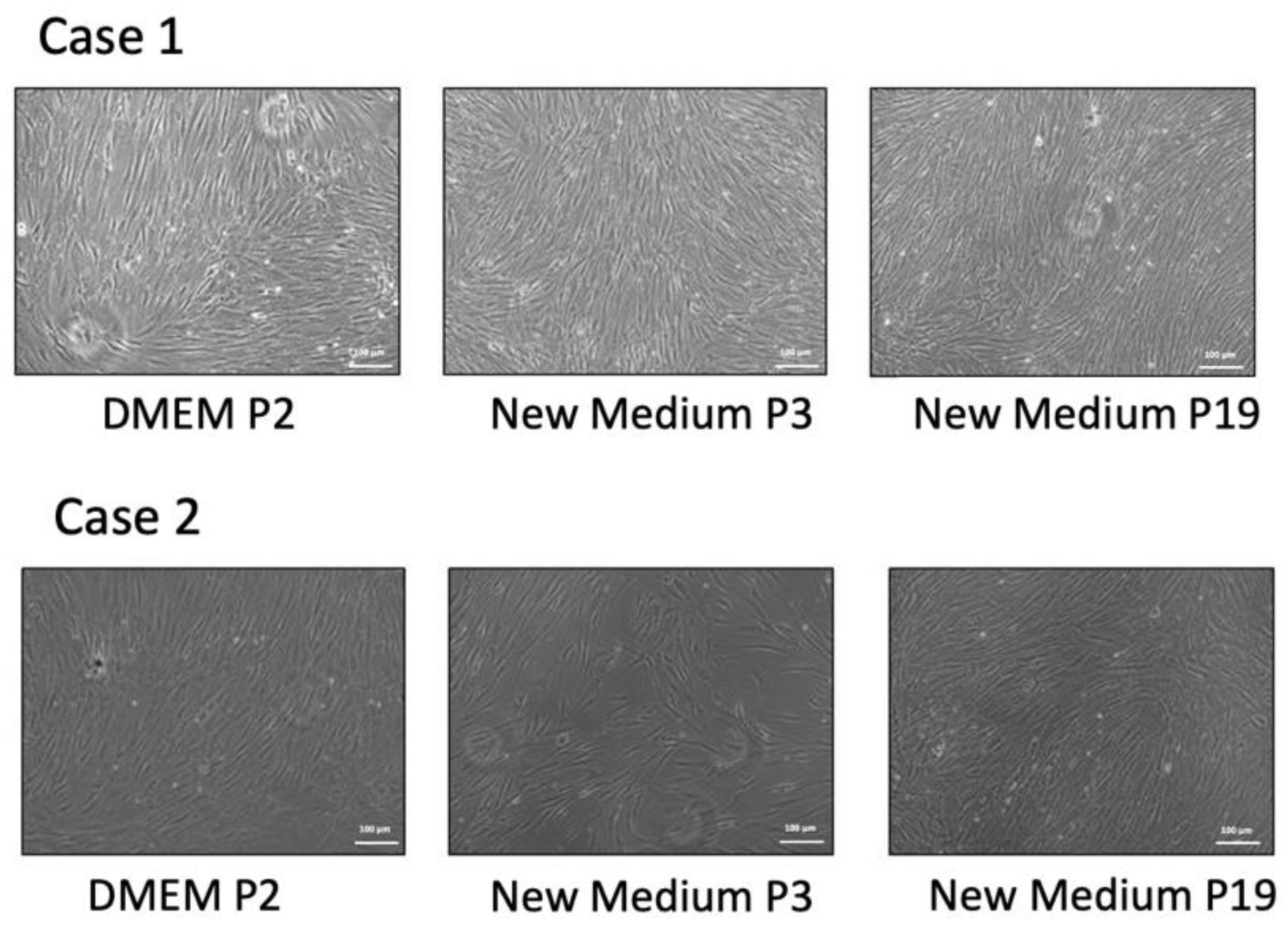
2.1. Isolation of Human Bone Marrow Stem Cells
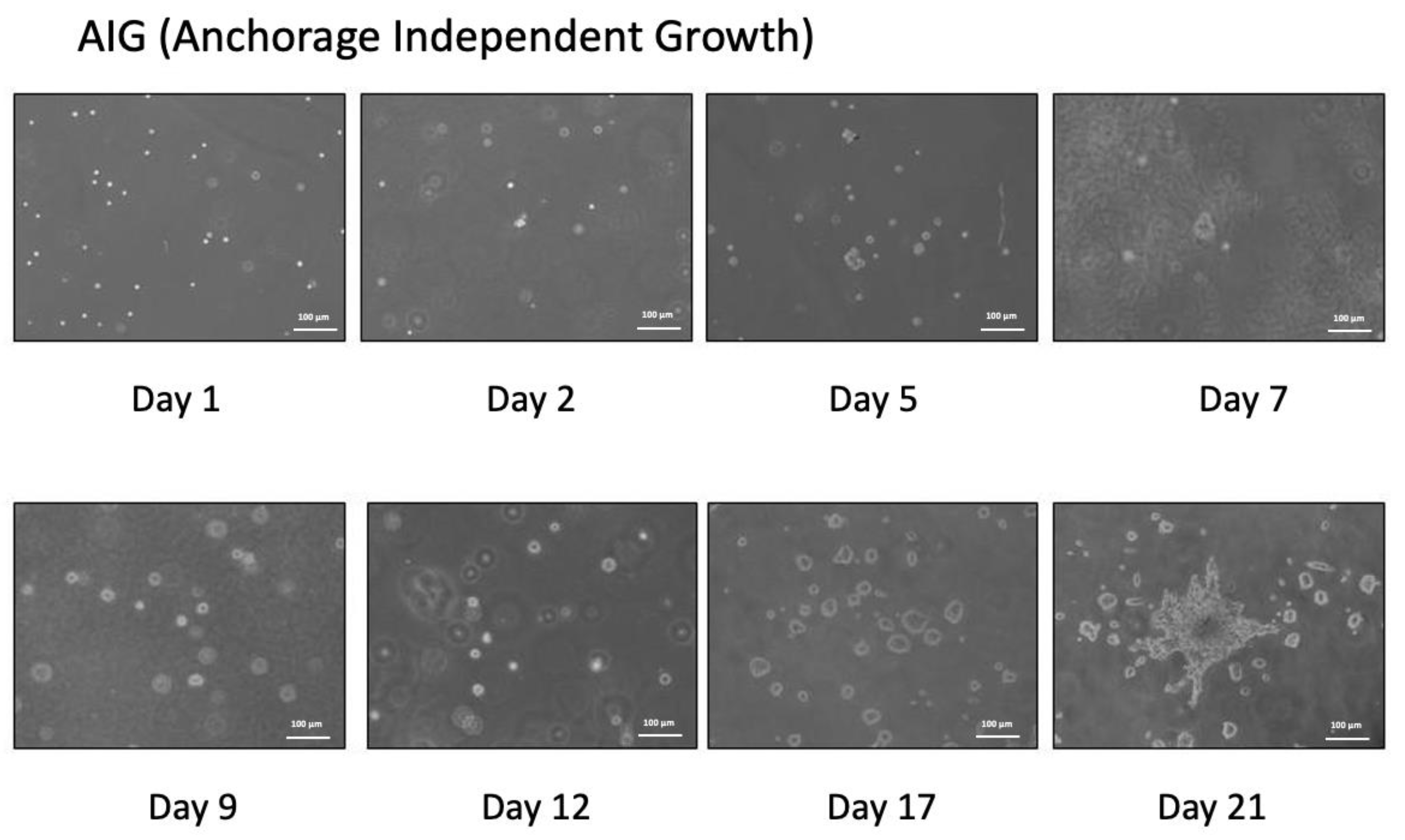
2.2. Anchorage Independent Growth
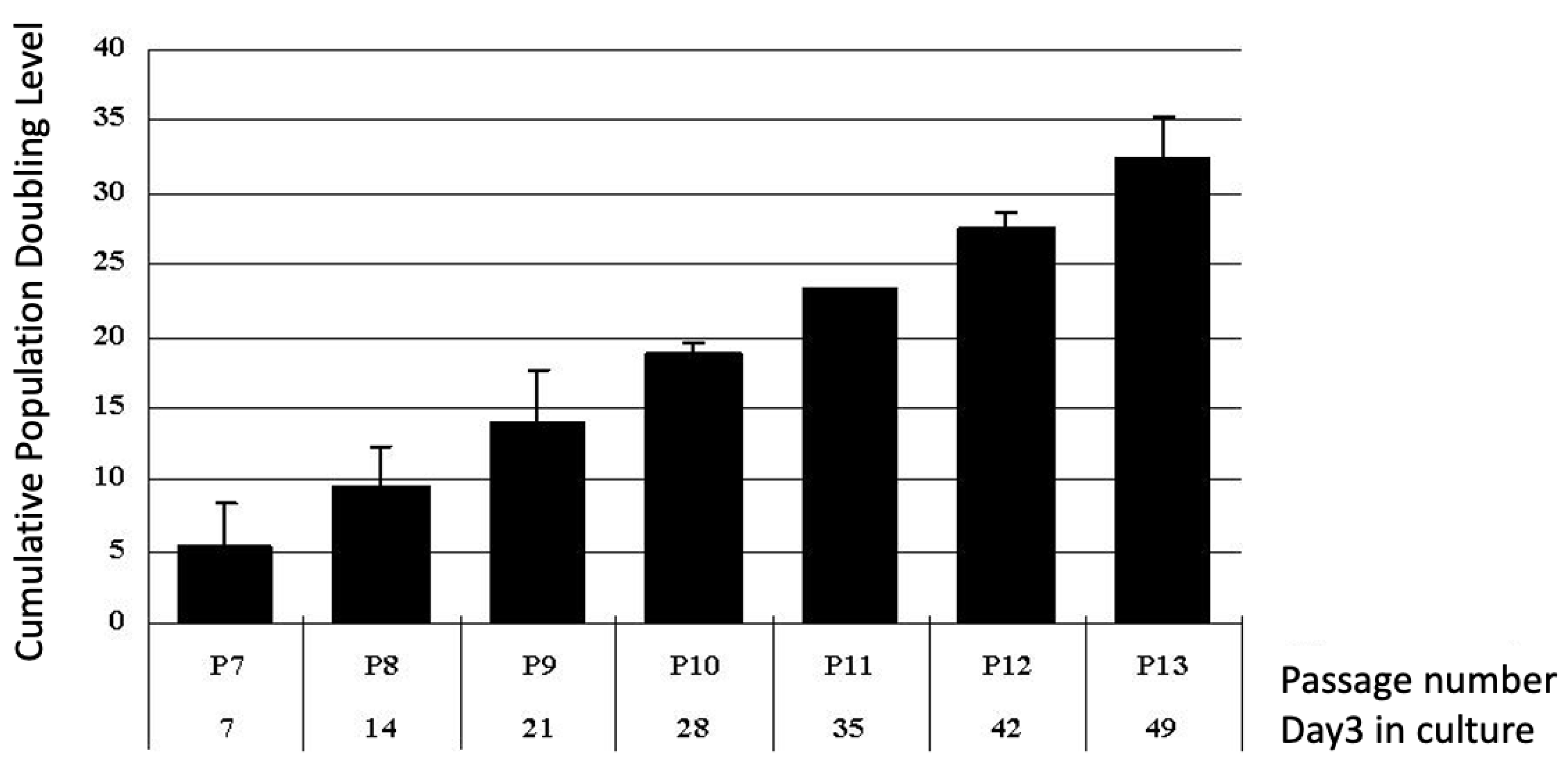
2.3. Proliferation Potential of Putative Human Bone Marrow Stem Cells
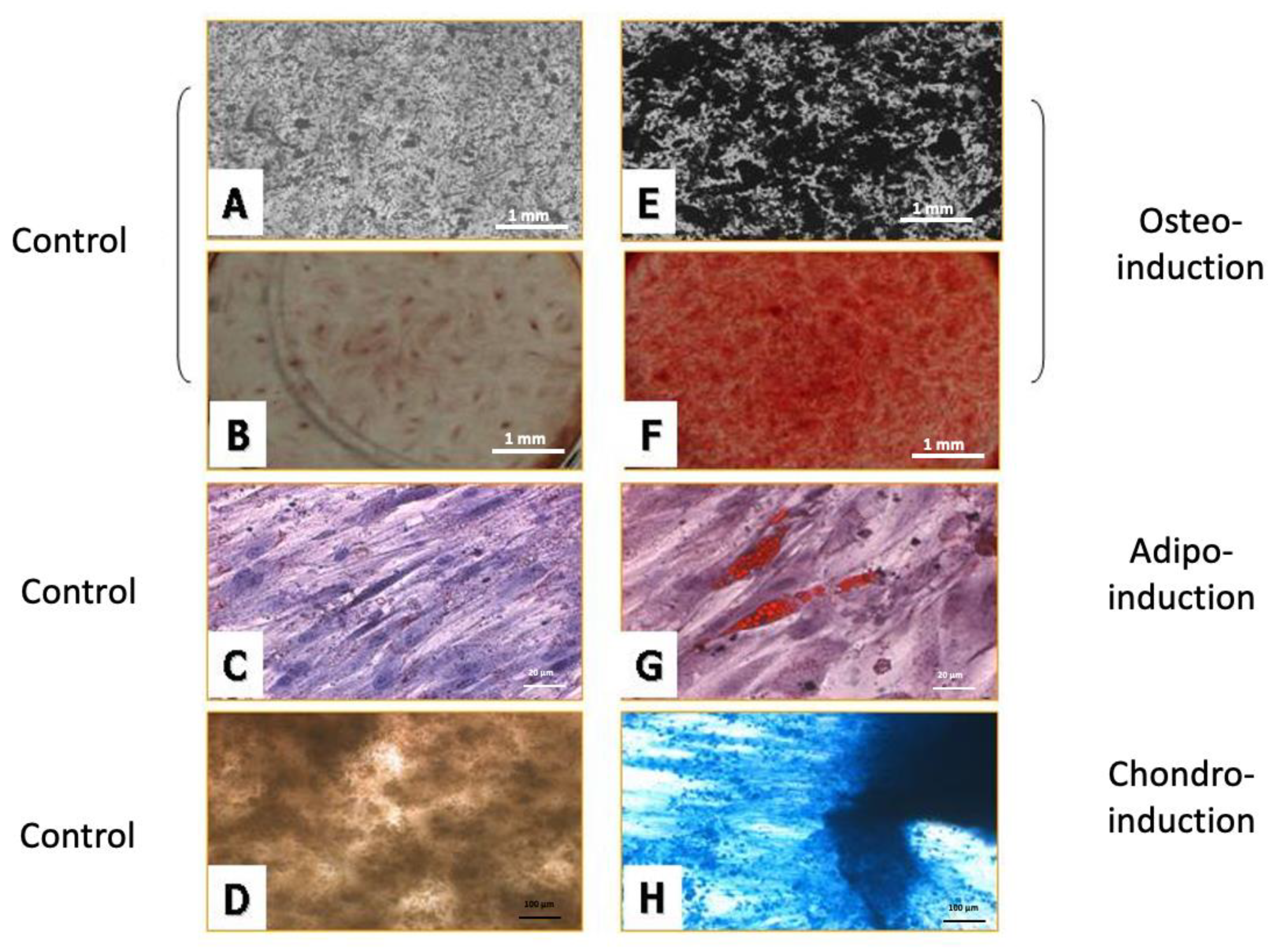
2.4. Induction of Multilineage Stem Cell Differentiation
2.4.1. Osteogenic Differentiation
2.4.2. Adipose Differentiation
2.4.3. Chondrogenic Differentiation
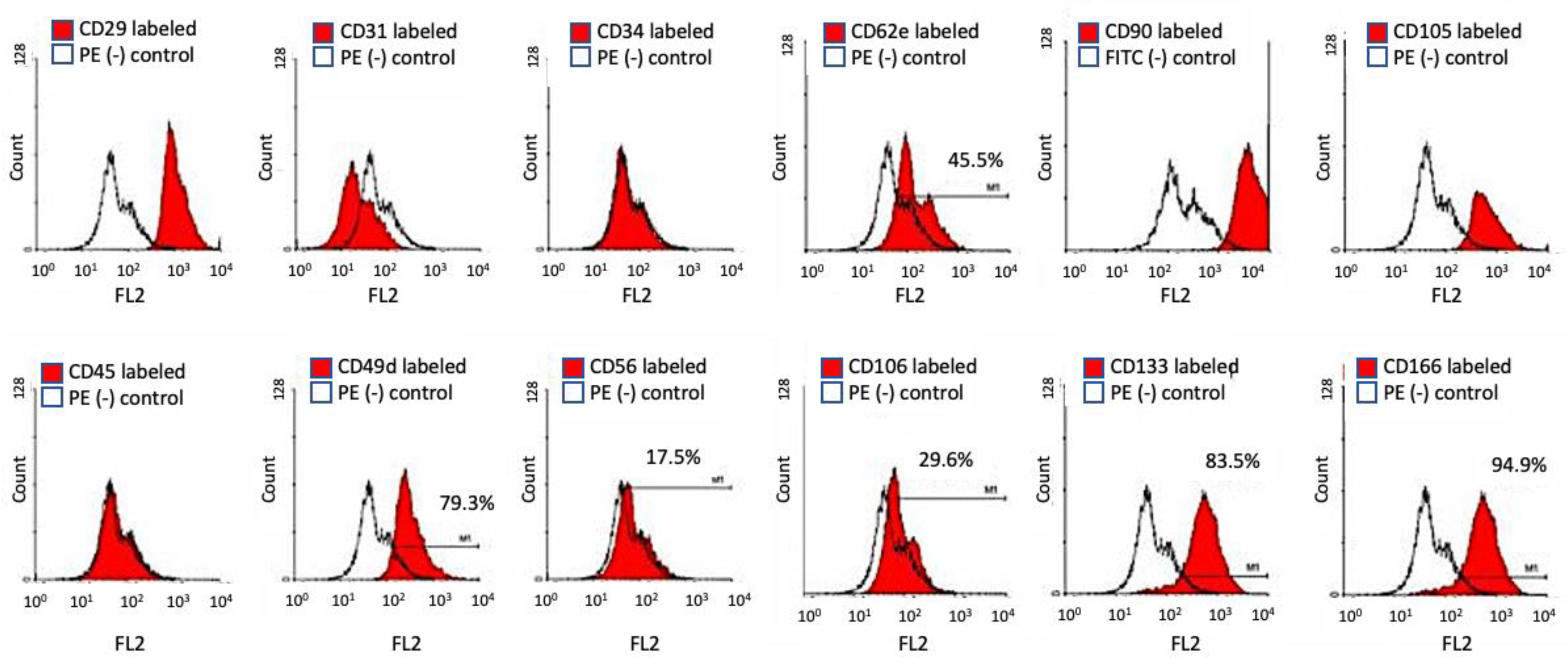
2.5. Flow Cytometry and Cell Surface Marker for Stem Cell Specificity
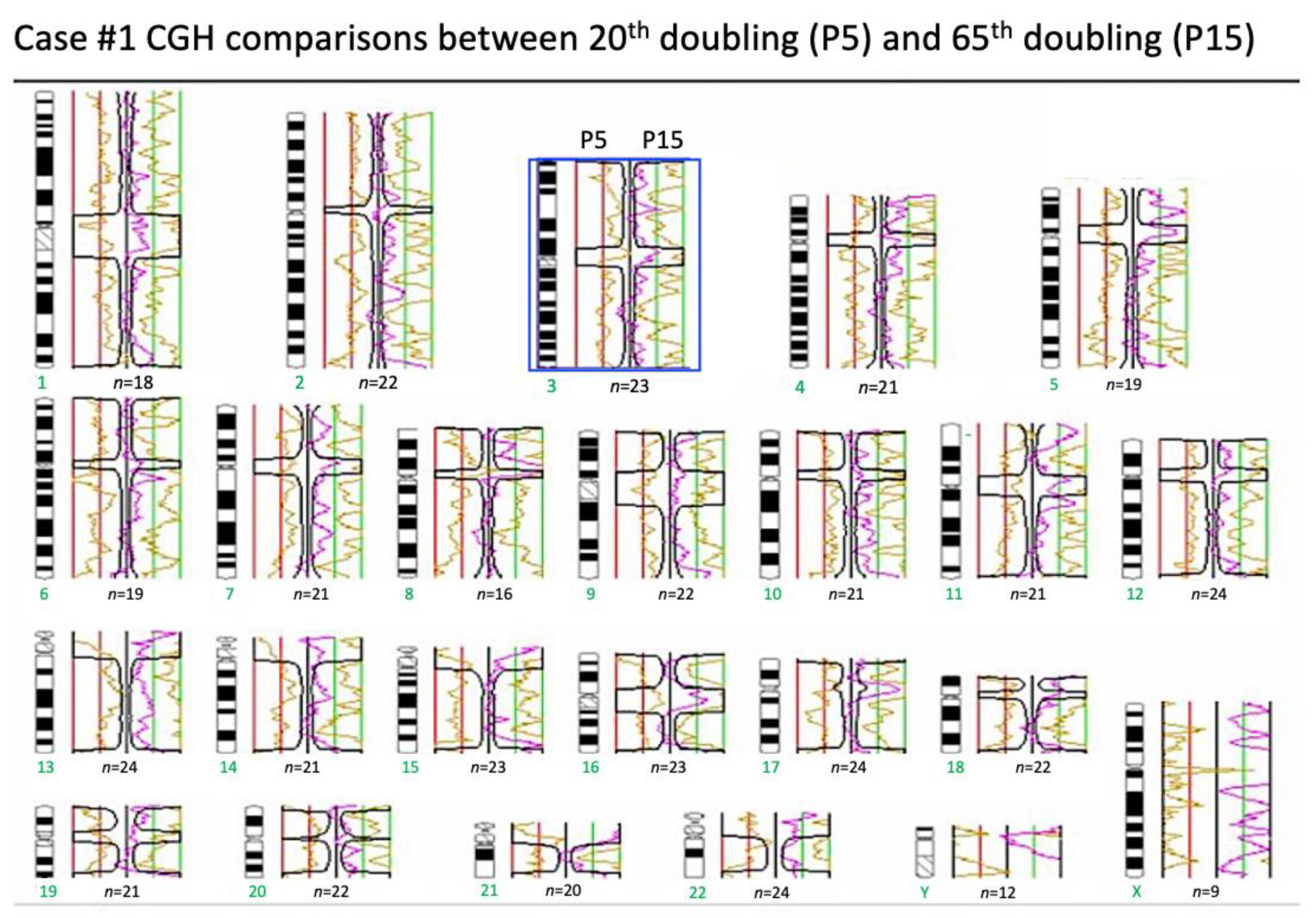
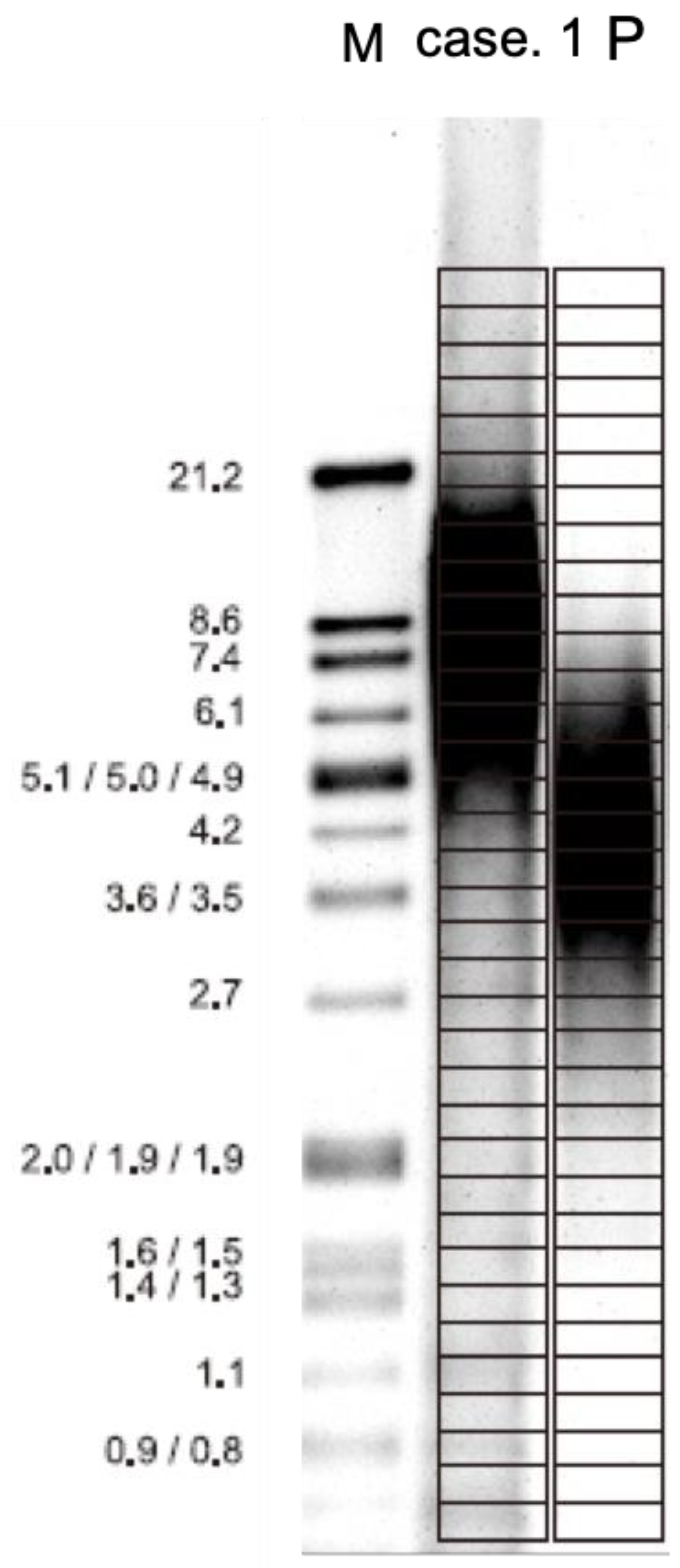
2.6. Array Comparative Genomic Hybridization (Array-CGH)
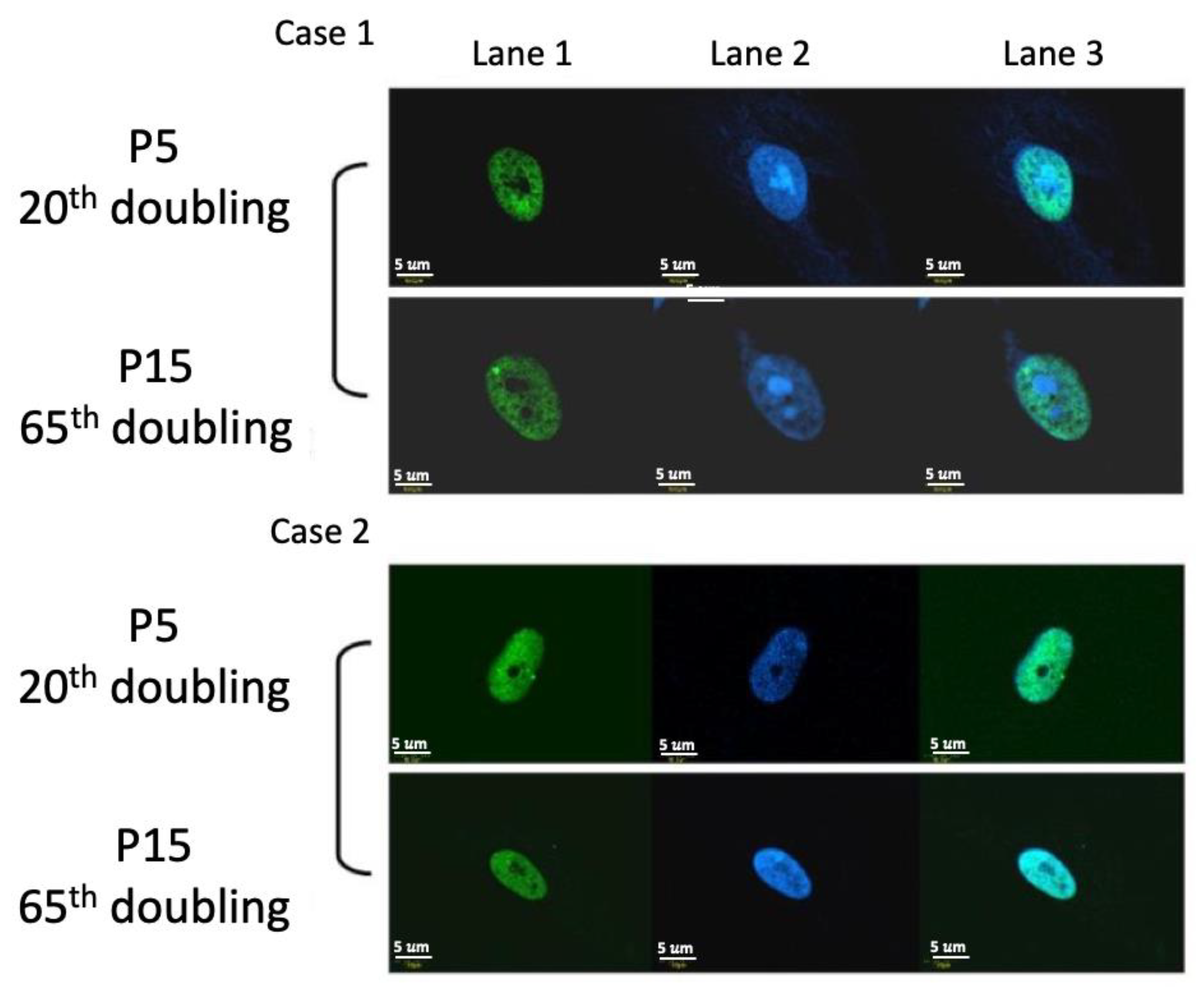
2.7. DNA Damage Protein 53BP1 Immunostaining
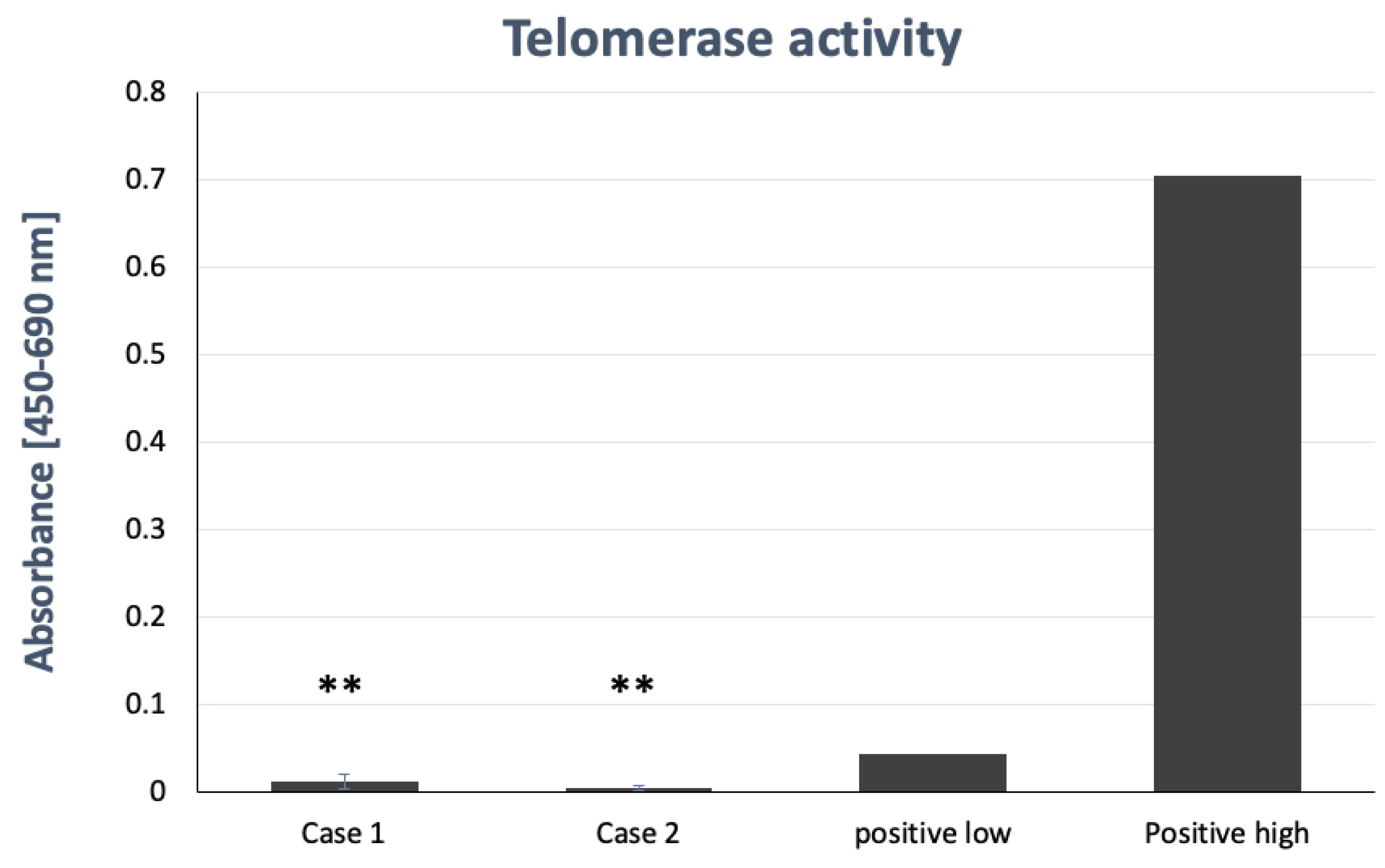
2.8. Telomerase Activity Assay
2.9. Telomere Length Assay
3. Discussion
4. Materials and Methods
4.1. Isolation of Human Bone Marrow Stroma Cells
4.2. Cumulative Population Doubling Level (cpdl)
4.3. Anchorage Independent Growth (AIG)
4.4. Multilineage Differentiation (Adipogenesis, Chondrogenesis, and Osteogenesis)
4.4.1. Osteogenic Differentiation
4.4.2. Chondrogenic Differentiation
4.4.3. Adipogenic Differentiation
4.5. Flow Cytometry and Cell Surface Marker for Stem Cell Specificity
4.6. Array Comparative Genomic Hybridization (Array-CGH)
4.7. DNA Damage Protein 53BP1 Immunostaining
4.8. Telomerase Activity Assay
4.9. Telomere Length Assay
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, H.T.; Lee, M.J.; Chen, C.H.; Chuang, S.C.; Chang, L.F.; Ho, M.L.; Hung, S.H.; Fu, Y.C.; Wang, Y.H.; Wang, H.I.; et al. Proliferation and differentiation potential of human adipose-derived mesenchymal stem cells isolated from elderly patients with osteoporotic fractures. J. Cell Mol. Med. 2012, 16, 582–593. [Google Scholar] [CrossRef]
- Saler, M.; Caliogna, L.; Botta, L.; Benazzo, F.; Riva, F.; Gastaldi, G. hASC and DFAT, Multipotent Stem Cells for Regenerative Medicine: A Comparison of Their Potential Differentiation In Vitro. Int. J. Mol. Sci. 2017, 18, 2699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, F.; Maioli, M.; Leonardi, E.; Olivi, E.; Pasquinelli, G.; Valente, S.; Mendez, A.J.; Ricordi, C.; Raffaini, M.; Tremolada, C.; et al. A new nonenzymatic method and device to obtain a fat tissue derivative highly enriched in pericyte-like elements by mild mechanical forces from human lipoaspirates. Cell Transpl. 2013, 22, 2063–2077. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, L.; Zhang, Y.; Huang, L.; Shi, Q. Mesenchymal stem cells—A promising strategy for treating knee osteoarthritis. Bone Joint Res. 2020, 9, 719–728. [Google Scholar] [CrossRef]
- Ruth, K. Stem Cells: Scientific Progress and Future Research Directions; National Institutes of Health: Bethesda, MD, USA, 2001; pp. 23–42.
- Zhang, K.; Ikeda, Y.; Kasugai, S.; Ikeda, M. Extended Culture Conditions for Multipotent Bone Marrow-Derived Mesenchymal Stem Cells. Kokubyo Gakkai Zasshi 2016, 83, 13–24. [Google Scholar]
- Bianco, P.; Gehron Robey, P. Marrow stromal stem cells. J. Clin. Investig. 2000, 105, 1663–1668. [Google Scholar] [CrossRef] [Green Version]
- Andrzejewska, A.; Lukomska, B.; Janowski, M. Concise Review: Mesenchymal Stem Cells: From Roots to Boost. Stem Cells 2019, 37, 855–864. [Google Scholar] [CrossRef] [Green Version]
- Friedenstein, A.J.; Petrakova, K.V.; Kurolesova, A.I.; Frolova, G.P. Heterotopic of bone marrow.Analysis of precursor cells for osteogenic and hematopoietic tissues. Transplantation 1968, 6, 230–247. [Google Scholar] [CrossRef]
- Neth, P.; Ciccarella, M.; Egea, V.; Hoelters, J.; Jochum, M.; Ries, C. Wnt signaling regulates the invasion capacity of human mesenchymal stem cells. Stem Cells 2006, 24, 1892–1903. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Chen, S.; Jiang, Q.; Pei, M. Determinants of stem cell lineage differentiation toward chondrogenesis versus adipogenesis. Cell Mol. Life Sci. 2019, 76, 1653–1680. [Google Scholar] [CrossRef] [PubMed]
- Cossu, G.; Mavilio, F. Myogenic stem cells for the therapy of primary myopathies: Wishful thinking or therapeutic perspective? J. Clin. Investig. 2000, 105, 1669–1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camernik, K.; Barlic, A.; Drobnic, M.; Marc, J.; Jeras, M.; Zupan, J. Mesenchymal Stem Cells in the Musculoskeletal System: From Animal Models to Human Tissue Regeneration? Stem Cell Rev. Rep. 2018, 14, 346–369. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Banno, K.; Kunitomi, H.; Tominaga, E.; Aoki, D. Current state and outlook for drug repositioning anticipated in the field of ovarian cancer. J. Gynecol. Oncol. 2019, 30, e10. [Google Scholar] [CrossRef]
- Baghaei, K.; Hashemi, S.M.; Tokhanbigli, S.; Asadi Rad, A.; Assadzadeh-Aghdaei, H.; Sharifian, A.; Zali, M.R. Isolation, differentiation, and characterization of mesenchymal stem cells from human bone marrow. Gastroenterol. Hepatol. Bed. Bench. 2017, 10, 208–213. [Google Scholar]
- Bruder, S.P.; Jaiswal, N.; Haynesworth, S.E. Growth kinetics, self-renewal, and the osteogenic potential of purified human mesenchymal stem cells during extensive subcultivation and following cryopreservation. J. Cell Biochem. 1997, 64, 278–294. [Google Scholar] [CrossRef]
- Yu, Y. Application of Stem Cell Technology in Antiaging and Aging-Related Diseases. Adv. Exp. Med. Biol. 2018, 1086, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Slack, J.M. Stem cells in epithelial tissues. Science 2000, 287, 1431–1433. [Google Scholar] [CrossRef]
- Chang, J.K.; Ho, M.L.; Yeh, C.H.; Chen, C.H.; Wang, G.J. Osteogenic Gene Expression Decreases in Stromal Cells of Patients with Osteonecrosis. Clin. Orthop. Relat. Res. 2006, 453, 286–292. [Google Scholar] [CrossRef]
- Caplan, A.I.; Bruder, S.P. Mesenchymal stem cells: Building blocks for molecular medicine in the 21st century. Trends Mol. Med. 2001, 7, 259–264. [Google Scholar] [CrossRef]
- Lin, T.M.; Tsai, J.L.; Lin, S.D.; Lai, C.S.; Chang, C.C. Accelerated growth and prolonged lifespan of adipose tissue-derived human mesenchymal stem cells in a medium using reduced calcium and antioxidants. Stem Cells Dev. 2005, 14, 92–102. [Google Scholar] [CrossRef]
- Minguell, J.J.; Erices, A.; Conget, P. Mesenchymal stem cells. Exp. Biol. Med. (Maywood) 2001, 226, 507–520. [Google Scholar] [CrossRef] [PubMed]
- Rubin, C.S.; Hirsch, A.; Fung, C.; Rosen, O.M. Development of hormone receptors and hormonal responsiveness in vitro. Insulin receptors and insulin sensitivity in the preadipocyte and adipocyte forms of 3T3-L1 cells. J. Biol. Chem. 1978, 253, 7570–7578. [Google Scholar] [CrossRef]
- Schaffler, A.; Buchler, C. Concise review: Adipose tissue-derived stromal cells--basic and clinical implications for novel cell-based therapies. Stem Cells 2007, 25, 818–827. [Google Scholar] [CrossRef]
- Moon, G.J.; Cho, Y.H.; Kim, D.H.; Sung, J.H.; Son, J.P.; Kim, S.; Cha, J.M.; Bang, O.Y. Serum-mediated Activation of Bone Marrow-derived Mesenchymal Stem Cells in Ischemic Stroke Patients: A Novel Preconditioning Method. Cell Transplant. 2018, 27, 485–500. [Google Scholar] [CrossRef] [Green Version]
- Tangtrongsup, S.; Kisiday, J.D. Differential Effects of the Antioxidants N-Acetylcysteine and Pyrrolidine Dithiocarbamate on Mesenchymal Stem Cell Chondrogenesis. Cell. Mol. Bioeng. 2019, 12, 153–163. [Google Scholar] [CrossRef]
- Popov, A.; Scotchford, C.; Grant, D.; Sottile, V. Impact of Serum Source on Human Mesenchymal Stem Cell Osteogenic Differentiation in Culture. Int. J. Mol. Sci. 2019, 20, 5051. [Google Scholar] [CrossRef] [Green Version]
- Diduch, D.R.; Coe, M.R.; Joyner, C.; Owen, M.E.; Balian, G. Two cell lines from bone marrow that differ in terms of collagen synthesis, osteogenic characteristics, and matrix mineralization. J. Bone Joint Surg. Am. 1993, 75, 92–105. [Google Scholar] [CrossRef]
- Smith, J.; Ladi, E.; Mayer-Proschel, M.; Noble, M. Redox state is a central modulator of the balance between self-renewal and differentiation in a dividing glial precursor cell. Proc. Natl. Acad. Sci. USA 2000, 97, 10032–10037. [Google Scholar] [CrossRef] [Green Version]
- De Flora, S.; Izzotti, A.; D’Agostini, F.; Balansky, R.M. Mechanisms of N-acetylcysteine in the prevention of DNA damage and cancer, with special reference to smoking-related end-points. Carcinogenesis 2001, 22, 999–1013. [Google Scholar] [CrossRef] [Green Version]
- Hata, R.; Senoo, H. L-ascorbic acid 2-phosphate stimulates collagen accumulation, cell proliferation, and formation of a three-dimensional tissuelike substance by skin fibroblasts. J. Cell. Physiol. 1989, 138, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Chepda, T.; Cadau, M.; Girin, P.; Frey, J.; Chamson, A. Monitoring of ascorbate at a constant rate in cell culture: Effect on cell growth. In Vitro Cell. Dev. Biol. Anim. 2001, 37, 26–30. [Google Scholar] [CrossRef]
- Chen, S.T.; Kang, L.; Wang, C.Z.; Huang, P.J.; Huang, H.T.; Lin, S.Y.; Chou, S.H.; Lu, C.C.; Shen, P.C.; Lin, Y.S.; et al. (−)-Epigallocatechin-3-Gallate Decreases Osteoclastogenesis via Modulation of RANKL and Osteoprotegrin. Molecules 2019, 24, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, O.K.; Kuo, T.K.; Chen, W.M.; Lee, K.D.; Hsieh, S.L.; Chen, T.H. Isolation of multipotent mesenchymal stem cells from umbilical cord blood. Blood 2004, 103, 1669–1675. [Google Scholar] [CrossRef] [Green Version]
- Berthiaume, M.J.; Raynauld, J.P.; Martel-Pelletier, J.; Labonte, F.; Beaudoin, G.; Bloch, D.A.; Choquette, D.; Haraoui, B.; Altman, R.D.; Hochberg, M.; et al. Meniscal tear and extrusion are strongly associated with progression of symptomatic knee osteoarthritis as assessed by quantitative magnetic resonance imaging. Ann. Rheum. Dis. 2005, 64, 556–563. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.H.; Lai, C.H.; Hong, Y.K.; Lu, J.M.; Lin, S.Y.; Lee, T.C.; Chang, L.Y.; Ho, M.L.; Conway, E.M.; Wu, H.L.; et al. Thrombomodulin Functional Domains Support Osteoblast Differentiation and Bone Healing in Diabetes in Mice. J. Bone Miner. Res. 2020, 35, 1812–1823. [Google Scholar] [CrossRef]
- Shao, P.L.; Wu, S.C.; Lin, Z.Y.; Ho, M.L.; Chen, C.H.; Wang, C.Z. Alpha-5 Integrin Mediates Simvastatin-Induced Osteogenesis of Bone Marrow Mesenchymal Stem Cells. Int. J. Mol. Sci. 2019, 20, 506. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.Y.; Kang, L.; Wang, C.Z.; Huang, H.H.; Cheng, T.L.; Huang, H.T.; Lee, M.J.; Lin, Y.S.; Ho, M.L.; Wang, G.J.; et al. (−)-Epigallocatechin-3-Gallate (EGCG) Enhances Osteogenic Differentiation of Human Bone Marrow Mesenchymal Stem Cells. Molecules 2018, 23, 3221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.H.; Chen, C.Y.; Chou, L.Y.; Chen, C.H.; Kang, L.; Wang, C.Z. Enhancement of Bone Marrow-Derived Mesenchymal Stem Cell Osteogenesis and New Bone Formation in Rats by Obtusilactone A. Int. J. Mol. Sci. 2017, 18, 2422. [Google Scholar] [CrossRef] [Green Version]
- Tai, I.C.; Wang, Y.H.; Chen, C.H.; Chuang, S.C.; Chang, J.K.; Ho, M.L. Simvastatin enhances Rho/actin/cell rigidity pathway contributing to mesenchymal stem cells’ osteogenic differentiation. Int. J. Nanomed. 2015, 10, 5881–5894. [Google Scholar] [CrossRef] [Green Version]
- Chuang, S.C.; Chen, C.H.; Fu, Y.C.; Tai, I.C.; Li, C.J.; Chang, L.F.; Ho, M.L.; Chang, J.K. Estrogen receptor mediates simvastatin-stimulated osteogenic effects in bone marrow mesenchymal stem cells. Biochem. Pharmacol. 2015, 98, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Teong, B.; Kuo, S.M.; Chen, C.H.; Chen, Y.K.; Cheng, Z.J.; Huang, H.H. Characterization and human osteoblastic proliferation- and differentiation-stimulatory effects of phosphatidylcholine liposomes-encapsulated propranolol hydrochloride. Biomed. Mater. Eng. 2014, 24, 1875–1887. [Google Scholar] [CrossRef]
- Fu, Y.C.; Lin, C.C.; Chang, J.K.; Chen, C.H.; Tai, I.C.; Wang, G.J.; Ho, M.L. A novel single pulsed electromagnetic field stimulates osteogenesis of bone marrow mesenchymal stem cells and bone repair. PLoS ONE 2014, 9, e91581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.J.; Chen, H.T.; Ho, M.L.; Chen, C.H.; Chuang, S.C.; Huang, S.C.; Fu, Y.C.; Wang, G.J.; Kang, L.; Chang, J.K. PPARgamma silencing enhances osteogenic differentiation of human adipose-derived mesenchymal stem cells. J. Cell Mol. Med. 2013, 17, 1188–1193. [Google Scholar] [CrossRef]
- Hung, S.H.; Yeh, C.H.; Huang, H.T.; Wu, P.; Ho, M.L.; Chen, C.H.; Wang, C.; Chao, D.; Wang, G.J. Pioglitazone and dexamethasone induce adipogenesis in D1 bone marrow stromal cell line, but not through the peroxisome proliferator-activated receptor-gamma pathway. Life Sci. 2008, 82, 561–569. [Google Scholar] [CrossRef]
- Yeh, C.H.; Chang, J.K.; Ho, M.L.; Chen, C.H.; Wang, G.J. Different differentiation of stroma cells from patients with osteonecrosis: A pilot study. Clin. Orthop. Relat. Res. 2009, 467, 2159–2167. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.H.; Ho, M.L.; Chang, J.K.; Hung, S.H.; Wang, G.J. Green tea catechin enhances osteogenesis in a bone marrow mesenchymal stem cell line. Osteoporos. Int. 2005, 16, 2039–2045. [Google Scholar] [CrossRef]
- Lin, Y.H.; Kang, L.; Feng, W.H.; Cheng, T.L.; Tsai, W.C.; Huang, H.T.; Lee, H.C.; Chen, C.H. Effects of Lipids and Lipoproteins on Mesenchymal Stem Cells Used in Cardiac Tissue Regeneration. Int. J. Mol. Sci. 2020, 21, 4770. [Google Scholar] [CrossRef]
- Wang, F.; Tarkkonen, K.; Nieminen-Pihala, V.; Nagano, K.; Majidi, R.A.; Puolakkainen, T.; Rummukainen, P.; Lehto, J.; Roivainen, A.; Zhang, F.P.; et al. Mesenchymal Cell-Derived Juxtacrine Wnt1 Signaling Regulates Osteoblast Activity and Osteoclast Differentiation. J. Bone Miner. Res. 2019, 34, 1129–1142. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Hu, Z.; Wu, J.; Mei, Y.; Zhang, Q.; Zhang, H.; Miao, D.; Sun, W. Sirt1 Promotes Osteogenic Differentiation and Increases Alveolar Bone Mass via Bmi1 Activation in Mice. J. Bone Miner. Res. 2019, 34, 1169–1181. [Google Scholar] [CrossRef]
- Wang, K.; Le, L.; Chun, B.M.; Tiede-Lewis, L.M.; Shiflett, L.A.; Prideaux, M.; Campos, R.S.; Veno, P.A.; Xie, Y.; Dusevich, V.; et al. A Novel Osteogenic Cell Line That Differentiates Into GFP-Tagged Osteocytes and Forms Mineral with a Bone-Like Lacunocanalicular Structure. J. Bone Miner. Res. 2019, 34, 979–995. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Liu, J.; Gan, Y.; Dai, K.; Zhao, J.; Huang, M.; Huang, Y.; Zhuang, Y.; Zhang, X. High-Dose TGF-beta1 Impairs Mesenchymal Stem Cell-Mediated Bone Regeneration via Bmp2 Inhibition. J. Bone Miner. Res. 2020, 35, 167–180. [Google Scholar] [CrossRef]
- Antich-Rossello, M.; Forteza-Genestra, M.A.; Calvo, J.; Gaya, A.; Monjo, M.; Ramis, J.M. Platelet-derived extracellular vesicles promote osteoinduction of mesenchymal stromal cells. Bone Joint Res. 2020, 9, 667–674. [Google Scholar] [CrossRef]
- Hefka Blahnova, V.; Dankova, J.; Rampichova, M.; Filova, E. Combinations of growth factors for human mesenchymal stem cell proliferation and osteogenic differentiation. Bone Joint Res. 2020, 9, 412–420. [Google Scholar] [CrossRef]
- Osagie-Clouard, L.; Sanghani-Kerai, A.; Coathup, M.; Meeson, R.; Briggs, T.; Blunn, G. The influence of parathyroid hormone 1-34 on the osteogenic characteristics of adipose- and bone-marrow-derived mesenchymal stem cells from juvenile and ovarectomized rats. Bone Joint Res. 2019, 8, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [Green Version]
- Denker, A.E.; Nicoll, S.B.; Tuan, R.S. Formation of cartilage-like spheroids by micromass cultures of murine C3H10T1/2 cells upon treatment with transforming growth factor-beta 1. Differentiation 1995, 59, 25–34. [Google Scholar] [CrossRef]
- Leboy, P.S.; Vaias, L.; Uschmann, B.; Golub, E.; Adams, S.L.; Pacifici, M. Ascorbic acid induces alkaline phosphatase, type X collagen, and calcium deposition in cultured chick chondrocytes. J. Biol. Chem. 1989, 264, 17281–17286. [Google Scholar] [CrossRef]
- Wu, S.C.; Chen, C.H.; Wang, J.Y.; Lin, Y.S.; Chang, J.K.; Ho, M.L. Hyaluronan size alters chondrogenesis of adipose-derived stem cells via the CD44/ERK/SOX-9 pathway. Acta Biomater. 2018, 66, 224–237. [Google Scholar] [CrossRef]
- Wang, C.Z.; Eswaramoorthy, R.; Lin, T.H.; Chen, C.H.; Fu, Y.C.; Wang, C.K.; Wu, S.C.; Wang, G.J.; Chang, J.K.; Ho, M.L. Enhancement of chondrogenesis of adipose-derived stem cells in HA-PNIPAAm-CL hydrogel for cartilage regeneration in rabbits. Sci. Rep. 2018, 8, 10526. [Google Scholar] [CrossRef] [Green Version]
- Teong, B.; Wu, S.C.; Chang, C.M.; Chen, J.W.; Chen, H.T.; Chen, C.H.; Chang, J.K.; Ho, M.L. The stiffness of a crosslinked hyaluronan hydrogel affects its chondro-induction activity on hADSCs. J. Biomed. Mater. Res. B Appl. Biomater. 2018, 106, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.H.; Wu, S.C.; Chen, C.H.; Wang, G.J.; Chang, J.K.; Ho, M.L. Parathyroid hormone 1-34 reduces dexamethasone-induced terminal differentiation in human articular chondrocytes. Toxicology 2016, 368–369, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.C.; Chen, C.H.; Chang, J.K.; Fu, Y.C.; Wang, C.K.; Eswaramoorthy, R.; Lin, Y.S.; Wang, Y.H.; Lin, S.Y.; Wang, G.J.; et al. Hyaluronan initiates chondrogenesis mainly via CD44 in human adipose-derived stem cells. J. Appl. Physiol. (1985) 2013, 114, 1610–1618. [Google Scholar] [CrossRef]
- Liu, M.; Alharbi, M.; Graves, D.; Yang, S. IFT80 Is Required for Fracture Healing Through Controlling the Regulation of TGF-beta Signaling in Chondrocyte Differentiation and Function. J. Bone Miner. Res. 2020, 35, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Arai, A.; Udagawa, N.; Zhao, L.; Nishida, D.; Murakami, K.; Hiraga, T.; Takao-Kawabata, R.; Matsuo, K.; Komori, T.; et al. Parathyroid Hormone Shifts Cell Fate of a Leptin Receptor-Marked Stromal Population from Adipogenic to Osteoblastic Lineage. J. Bone Miner Res. 2019, 34, 1952–1963. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| h-ASC | BM-MSC | Osteoblast-Like Cells | BM-MSC Bone Media P6 | BM-MSC P6 New Media | BM-MSC P13 New Media | BM-MSC P15 New Media | |
|---|---|---|---|---|---|---|---|
| CD 29 | + | + | + | + | + | + | + |
| CD 31 | − | − | + | − | − | − | − |
| CD 34 | +/− | − | + | − | − | − | − |
| CD 44 | + | + | + | + | |||
| CD 45 | − | − | − | − | − | − | − |
| CD 56 | − | − | + | +/− | − | − | − |
| CD 62e | − | − | + | +/− | − | − | − |
| CD 90 | + | + | + | + | + | + | |
| CD105 | + | + | + | + | + | + | |
| CD106 | − | − | − | − | − | − | − |
| CD133 | − | − | +/− | +/− | − | − | |
| CD166 | + | + | + | +/− | +/− | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.-D.; Chuang, S.-C.; Cheng, T.-L.; Lee, M.-J.; Chen, H.-T.; Lin, S.-Y.; Huang, H.-T.; Ho, C.-J.; Lin, Y.-S.; Kang, L.; et al. An Intermediate Concentration of Calcium with Antioxidant Supplement in Culture Medium Enhances Proliferation and Decreases the Aging of Bone Marrow Mesenchymal Stem Cells. Int. J. Mol. Sci. 2021, 22, 2095. https://doi.org/10.3390/ijms22042095
Yang C-D, Chuang S-C, Cheng T-L, Lee M-J, Chen H-T, Lin S-Y, Huang H-T, Ho C-J, Lin Y-S, Kang L, et al. An Intermediate Concentration of Calcium with Antioxidant Supplement in Culture Medium Enhances Proliferation and Decreases the Aging of Bone Marrow Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2021; 22(4):2095. https://doi.org/10.3390/ijms22042095
Chicago/Turabian StyleYang, Chung-Da, Shu-Chun Chuang, Tsung-Lin Cheng, Mon-Juan Lee, Hui-Ting Chen, Sung-Yen Lin, Hsuan-Ti Huang, Cheng-Jung Ho, Yi-Shan Lin, Lin Kang, and et al. 2021. "An Intermediate Concentration of Calcium with Antioxidant Supplement in Culture Medium Enhances Proliferation and Decreases the Aging of Bone Marrow Mesenchymal Stem Cells" International Journal of Molecular Sciences 22, no. 4: 2095. https://doi.org/10.3390/ijms22042095
APA StyleYang, C. -D., Chuang, S. -C., Cheng, T. -L., Lee, M. -J., Chen, H. -T., Lin, S. -Y., Huang, H. -T., Ho, C. -J., Lin, Y. -S., Kang, L., Ho, M. -L., Chang, J. -K., & Chen, C. -H. (2021). An Intermediate Concentration of Calcium with Antioxidant Supplement in Culture Medium Enhances Proliferation and Decreases the Aging of Bone Marrow Mesenchymal Stem Cells. International Journal of Molecular Sciences, 22(4), 2095. https://doi.org/10.3390/ijms22042095