1. Introduction
Plants dependent on animals in the pollination process evolved different strategies to attract pollinators, thereby increasing reproductive success. The main parts of these strategies are flower traits (the size, shape, color, scent, and nectar) adapted to a given pollinator or their whole group. Pollinator-mediated selection on floral traits is well documented, and adaptation of plants to the most effective pollinators drives the evolution of pollination syndromes [
1]. The flagship example of the unusual diversity of flowers and equally differentiated pollination mechanisms is Orchidaceae, which is one of the biggest families among flowering plants [
2]. About one-third of its representatives deceive pollinators through sexual or food deception [
2,
3,
4]. Other groups of orchids reward pollinators in a different way, producing oils, nectar, resin, wax, and fragrances [
5,
6]. Among rewards offered by orchids, nectar is the most effective [
2,
7,
8]. Fruiting in nectariferous orchids is significantly higher than in nectarless [
2,
8]. Although nectariferous orchids constitute a large part of the family, and the role of nectar in highly effective pollination is indisputable, information on its chemical composition in Orchidaceae is very scarce. Moreover, many data derive from studies using less sensitive methods in comparison to those applied recently. Importantly, more data on nectar chemistry provide results of studies on plants from other families [
9,
10,
11,
12,
13,
14,
15], but they often focus on cultivars and the feeding needs of their pollinators, mainly bees.
Although available data document a great variability of nectar properties at different levels (species, population, and even individual), some patterns are outlined. In flower nectar, three main sugar components dominate, i.e., sucrose, glucose, and fructose, with different ratios between them. Nectar of the majority species is sucrose dominated [
12,
13,
16], but some papers document domination of hexoses over sucrose [
14,
17,
18,
19]. The concentration of sugars also shows a great variation (from about 7–70%, [
20,
21]) and is connected with pollinator types [
9,
10,
22,
23], especially with the adaptation of their mouthparts to use nectar of a given viscosity. For example, bees prefer the highest concentration of sugars in nectar (35% on average), while bats and hawkmoths can suck nectar with a 17–19% concentration of sugars [
23,
24,
25,
26]. In orchids, nectar sugar concentrations range from a few to about 50% [
17,
18,
27,
28,
29,
30]. The preferences of pollinators also concern other components of nectar: amino acids (AAs). They are present in nectar at a lower amount than sugars but play a significant role as a source of nutrition and in attracting pollinators, thereby affecting reproductive success and survival of nectar-feeding animals [
14,
16,
31,
32,
33]. Some authors suggest that taste function is even more important than a nutritive value [
34,
35]. Nectar of plants adapted to pollination by butterflies is characterized by high AA concentration, while those pollinated by birds or flies are characterized by their lower concentration [
34]. In the nectar of different species, some AAs dominate, and others are present in low concentrations or are absent [
17,
28,
36,
37].
Apart from nectar quantity and quality, its accessibility also influences plant-pollinator interaction, thereby affects plant reproductive success. If nectar is secreted inside the corolla or in a spur, it is protected against evaporation and is available for specific, restricted groups of pollinators. On the other hand, exposed nectar may be collected by pollinators representing different morphological and ecological types and is more vulnerable to evaporation and robbery [
38]. Moreover, nectar in flowers with concealed nectaries tends to be dominated by sucrose, while in more open flowers, it is dominated by glucose and fructose [
29,
39].
In papers dedicated to plant-pollinator interaction, the role of flower structure in attracting pollinators was studied more often than nectar properties [
40,
41,
42,
43]. In particular, phenotypic selection and its dependence on the mutual match between pollinator and flower traits are well documented [
44]. This match is one of the most important evolutionary mechanisms [
2,
4] and is an effect of the potential for adaptation to the local partners. Many researchers have shown that pollinators act as selection agents on floral morphology and contribute to plant fitness [
45,
46,
47]. Van der Niet, et al. [
46] stated that when pollinators’ fitness is strongly influenced by an ability to access the reward in flowers of a given species, the adaptation of pollinators to flowers, rather than flowers to pollinators, takes place. In the case of plants, in which flowers are arranged as the inflorescences, floral display (the length of inflorescence and number of flowers) may also contribute to reproductive success. Plants with larger inflorescences often set more fruit, due to attracting more pollinators, which visit more flowers on larger inflorescences [
48,
49,
50,
51,
52]. However, in cases in which larger inflorescences suffer from factors that decrease fitness, such as a higher probability of geitonogamy or intense herbivore activity, smaller inflorescences are favored by natural selection [
52,
53,
54].
Both floral characters and pollinator assemblages vary in space [
19,
41,
55,
56]. Variation of floral traits in the geographic range of plant species is often an adaptation to the locally most-effective pollinators, being an answer for requirements of their specific assemblages present in a given environment [
14,
28,
46,
56,
57]. The shift of floral traits and pollinators assemblages in space translate into differentiated direction and strength of selection and variation of the level of reproductive success [
58].
Reproductive success depends on more than an evolutionary match between plants and pollinators. Environmental factors, both biotic (co-occurring plants) and abiotic (soil resources, weather conditions), in places where populations exist may also importantly shape plant-pollinator interactions. The composition of local pollinators is strictly connected to the diversity of the plant community because more plant species accumulate a wider spectrum of resources for flower-visiting animals [
59]. Plant species richness, blossom cover, and especially the presence of attractive plant species influence assemblages of pollinators and the frequency of their visits [
60,
61]. It seems especially important in the case of generalist plants, which depend on many species in the pollination process. On the one hand, the presence of other flowering plants may facilitate the visitation rate, and as a consequence, increase the reproductive success of a given species [
62,
63]. On the other hand, a higher diversity of plant species may increase competition for pollinators when species share pollinators [
62,
63,
64,
65], especially when populations of pollinators are not abundant. Competition for pollination resources can also include intraspecific competition, which may be stronger than interspecies competition, according to niche theory [
66]. The richness of the plant community, and the growth and flowering of particular species, strictly depend on soil conditions. For example, David, et al. [
67] found that a high level of N in soil and a low pH decrease species diversity and the abundance of nectariferous plants. In effect, nectar and pollen resources decline, causing a decrease in pollinators’ assemblages [
68]. Soil properties also shape other plant traits, which influence the level of reproductive success, e.g., the flowering [
69] or quantity and quality of nectar [
27].
Due to the unusual richness of orchids’ flowers and the wide variation of relations with pollinators, orchids are often considered a model system to study plant-pollinator interactions and evolutionary processes. The majority of orchid species are specialists and are connected to only one pollinator species (67% of all orchids; [
70]) or a single functional group [
71,
72,
73,
74]. Others are generalists, and a wide range of animals may pollinate them. For example,
Epipactis palustris is pollinated by more than 100 species [
75], and in
Neottia ovata almost 300 different species were noted as visitors, with about 50 species carrying pollinia [
76]. Specialist orchids are more frequent objects of studies on selection/coevolution between plant and pollinators than generalists. Therefore, it seems interesting to choose the generalist orchid
N. ovata as a model species to test in which way flower traits are adapted to pollination by a wide range of pollinators.
N. ovata was the object of studies on pollination mechanism [
76,
77,
78], demographic processes [
79,
80,
81], genetic variation [
82,
83], and flower anatomy [
84]. So far, there are no published data on nectar composition and floral structure in this orchid and their role in the effectiveness of reproduction. Therefore, the main aim of our study was to determine the floral traits of the generalist orchid
N. ovata and to test for selection on floral morphology and nectar chemistry in populations existing in different habitats. Such studies enrich knowledge about evolutionary factors and processes that underlie the generalization or specialization and consequences at the population and species levels.
3. Discussion
Plants evolved different strategies to achieve reproductive success. In animal pollinated species, the level of RS depends, first of all, on the presence and abundance of pollinators [
85]. Their deficiency is recognized as the main cause of low RS in orchids [
2]. Assemblages of pollinators are strictly connected to the character of vegetation [
60,
61,
86,
87,
88]. Plants being hosts of
N. ovata pollinators are common (e.g., species from the Apiaceae family, and
Alnus,
Crataegus,
Betula,
Salix,
Corylus,
Vaccinium genera) [
76] and were present, more or less frequently, in plant communities in which the studied populations exist. However, vegetation in populations and surrounding areas also showed differences, which certainly influenced insects’ assemblages. Nilsson [
76] found that saw-flies, one of the most important
N. ovata pollinators, were present only in the population near marsh vegetation. This could partially explain the higher level of RS, especially FRS, in populations that existed in BNP on mineral islands among peat bogs in comparison to others (SKA, TUR, and WIS) surrounded by a distinct type of vegetation. Other plants may also decrease RS, competing successfully for pollinators, offering them more and/or better food [
62,
63,
64,
65], especially when populations of pollinators are not abundant. Nilsson [
76] found differentiation of the presence and abundance of visitors and pollinators in distinct Swedish
N. ovata populations. Variability in insect assemblages, and their abundance was probably one of the main factors shaping the levels of RS in populations of this orchid in northeast Poland. Insect assemblages also fluctuate from year to year [
76,
89], which may explain the temporary variation of RS in some
N. ovata populations.
According to Nilsson [
76],
N. ovata may be visited by almost 300 species, representing different systematic groups with a wide spectrum of body sizes, mouth apparatus, and nutritional preferences. Undoubtedly, the main role in the attraction of these insects is played by the scent bouquet, comprised of compounds that are known as general attractants of a wide range of insects [
71,
76]. Numerous insects capable of pollinating flowers of this orchid, together with the easily available nectar on the labellum, create a chance for a high level of RS. In the majority of populations, we observed a higher level of fruiting than those found by Brzosko [
80] and Brys, et al. [
79]. PR in all
N. ovata populations in both years (always above 80% or even above 90% in many cases) suggests that a large number of insects penetrated flowers. On the other hand, FRS was more differentiated (similar to the seven-year studies of Brzosko [
80]). The higher efficiency of pollination we noted in populations from Biebrza National Park, and in remaining FRS was 5–11 times lower than that of PR. This indicates that not all insects that visited flowers (even able to collect pollinia) were effective pollinators. Probably some of the visitors, especially the smallest or the weakest, may suck nectar only from the groove along the labellum and do not penetrate flowers in-depth, omitting in this way the cave at the labellum base, which decreases the probability of contact with the column. Moreover, pollinaria may be attached to different parts of the insect’s body [
76], and the position of the visitor sucking nectar may sometimes be unsuitable for the collection of pollinia and/or to place them on the stigma. The low efficiency of pollination could be explained by Nilsson [
76]. The author found behavioral disturbances of smaller ichneumons (dominant pollinators) if they have big loads of pollinia. Insects that do not penetrate the flowers correctly may occasionally contribute to the pollination of
N. ovata flowers. This indicates that pollination in this species has a haphazard character. The disparity between PR and FRS indicates that pollinia are often lost, as observed by Brys, et al. [
79].
Incorrect flower penetration, causing ineffective pollination, is an effect of mismatch between flower and pollinator. The mechanical fit between partners is one of the essential preconditions of successful pollination [
45,
46,
47] and one of the most important evolutionary mechanisms [
2,
4,
44]. Such a match is generally stronger in specialized systems [
90], which confirms, for example, the results of studies on long-spurred orchids [
41,
42,
43,
51]. Our results suggest the best fit between flowers and pollinators in POG and LUB populations. PR:FRS in LUB in 2019 and in POG in 2020 was close to 1, and additionally, in LUB in 2019, FRS was higher than PR, indicating the presence of effective pollinators and their high efficiency. The high PR:FRS ratio in other populations and the relatively low FRS in some of them denote a mismatch between flowers and visitors and, as a result, a larger loss of pollinia. Weak correlations between flower traits and PR and FRS (five cases among 84 analyzed in both years) confirm this mismatch. In these single cases, distinct flower traits were under selection in particular populations. Nevertheless, four among five flower traits correlated with PR or FRS concerned the sizes of structures (groove and cage), in which nectar is secreted and accumulated. Because we supposed that groove and cavity sizes are the measures of nectar quantity, it could indicate that the amount of nectar is the most important trait influencing RS in
N. ovata. We expected that the labellum in this orchid, as a landing platform and flower part, should be adapted to pollinators’ sizes. Although we did not find an influence of labellum on RS parameters, its length was the most differentiated flower structure between populations, which suggests that it reacts on local insect assemblages. The disparity between the level of PR and FRS in most populations, probably being an effect of structural mismatch, may be explained through the great variation in body sizes of
N. ovata visitors and the differentiation of their behaviors as nectar consumers. Even the main group of pollinators of this species (ichneumonids) includes representatives with a wide range of sizes [
76]. In populations with lower FRS, these were probably predominant insects that more accidentally remove pollinia and less often place them on stigmas of other flowers. Because their main dietary sources are connected to other plant species, and
N. ovata is a marginal part of the food (if only because of small population sizes), they do not need to adapt to its flowers. This suggests that the level of
N. ovata RS depends on accompanying plant species, their diversity, and their abundance. Contrary to Brys, et al. [
79] results, we did not find an influence of floral display on RS in
N. ovata.
In nectariferous plants, the amount and composition of nectar are known to affect plant-pollinator interactions [
9,
10,
12,
13,
16,
19,
22,
23,
27,
28,
34]. Our studies document that
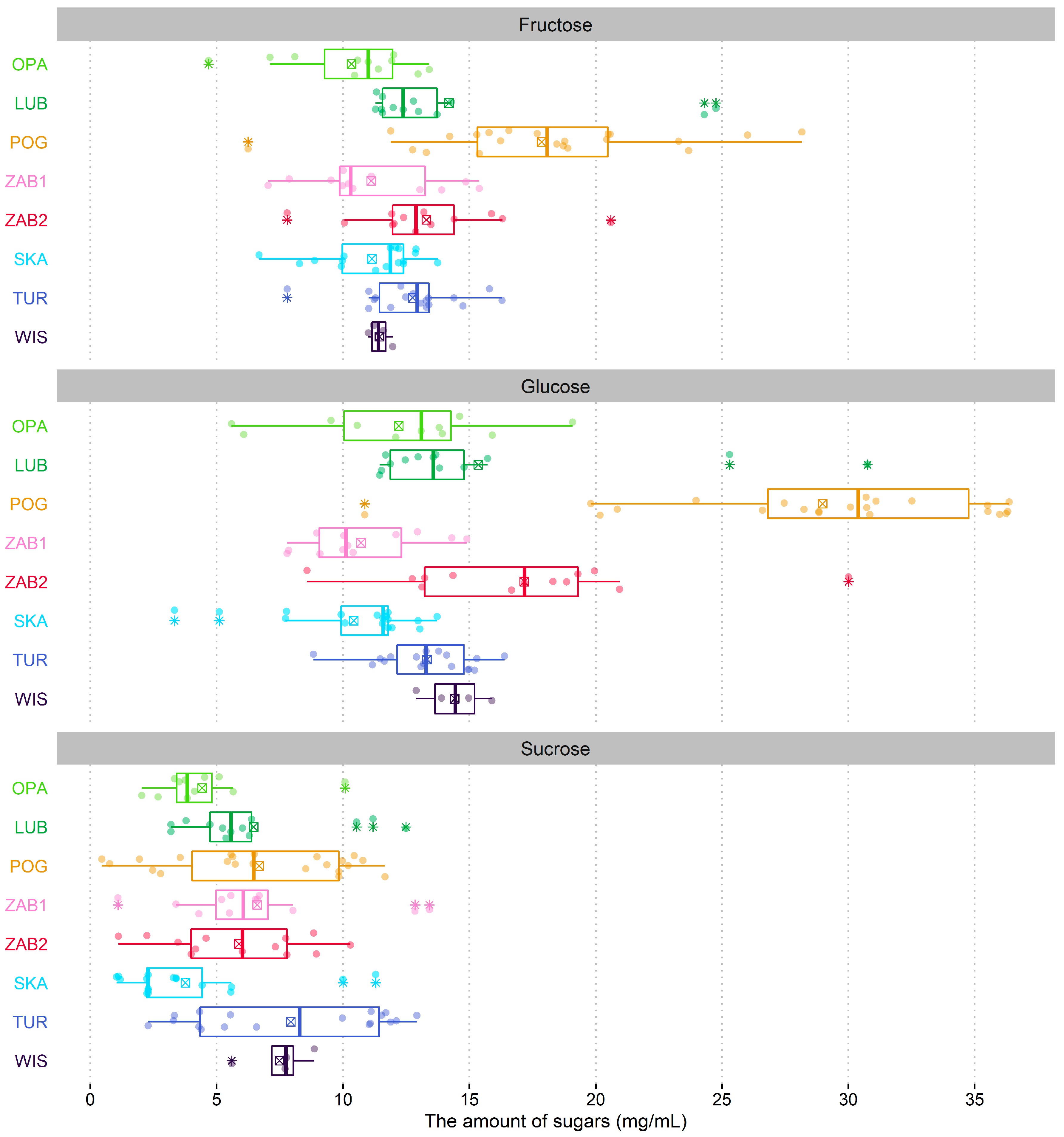
N. ovata is characterized by exceptionally diluted nectar with the lowest sugar concentration among orchids [
17,
18,
27,
28,
29,
30]. To our knowledge, these values are comparable only to the concentration of nectar used by some hummingbirds [
26,
91]. The relatively low sugar concentration was noted for plants pollinated by moths and flies [
17,
23,
24,
26,
92,
93], and only fly-pollinated species have extremely low volume and sugar concentration but high amino acids and hexose content [
12,
34]. Some fly species are also known to pollinate
N. ovata [
76]. The nectar of
N. ovata is dominated by hexoses, which is in agreement with the statements of Gottsberger, et al. [
39] and Pais, et al. [
29] that nectar in flowers with concealed nectaries tends to be dominated by sucrose, while in more open flowers by glucose and fructose. Hexose solution has a higher osmolarity, and therefore lower evaporation rates, than sucrose solution, which can explain the high proportion of hexoses in shallow flowers [
12]. However, the prevalence of hexoses was noted in the nectar of some long-spurred orchids [
17,
18]. On the other hand, contrary to our results, Galetto, et al. [
18], studying nectar in five orchid species, found that nectar located in the spurs in two
Habenaria species was copious and less concentrated (<20%), while in species in which nectar was accumulated in the basal lateral parts of the labellum, it was more concentrated (ca. 50%). Our results are in accordance with the studies of Johnson and Nicolson [
94], who documented a clear distinction between nectar sucrose content of specialized (40–60%) and generalized (0–5%) bird-pollinated species. Nonspecialized insects, i.e., syrphids, flies, and beetles (insects from these groups are
N. ovata pollinators) [
76] preferred monosaccharide nectar of plants from phryganic communities [
22]. Hexose-rich nectar, which is taken up more easily than sucrose, may be an adaptation and advantage for attracting a wide range of nonspecialized pollinators. It is worth noting that low sucrose nectar is also characteristic for species from the Apiaceae family [
22,
95,
96], which are pollinated by the same systematic groups of insects as
N. ovata. The lack of influence of nectar sugars on RS in five populations confirms that these nectar traits are not aimed at any of the pollinator group. Moreover, our results could suggest that insects operated in the three remaining populations did not prefer nectar sugar composition in nectar offered by
N. ovata. With the exception of the TUR population, in which fructose participation benefited FRS, in the remaining cases, statistically significant negative correlations were noted. In studies on two
Platanthera species, we also noted positive selection only on fructose content [
17]. To amount the preferences of insects, experiments should be performed. Hexoses, and especially fructose, are preferred by some pollinators due to their lower viscosity, enabling easier absorption [
25]. Heil [
31] documented that some ants (often observed on
N. ovata in our studies and by Nilsson [
76]) even preferred sucrose-free nectar because they are not able to assimilate this sugar due to lack of invertase. Sucrose-rich nectar may be toxic for some generalists. All the above-mentioned results, at least partially, explain the dominance of hexoses and the high fructose:glucose ratio in the majority of
N. ovata populations. The concentration of sugars in nectar and the sucrose:hexose ratio also depend on water availability [
22].
N. ovata populations exist in relatively wet places, and heavy rainfall in 2020 might additionally decrease sugar concentration.
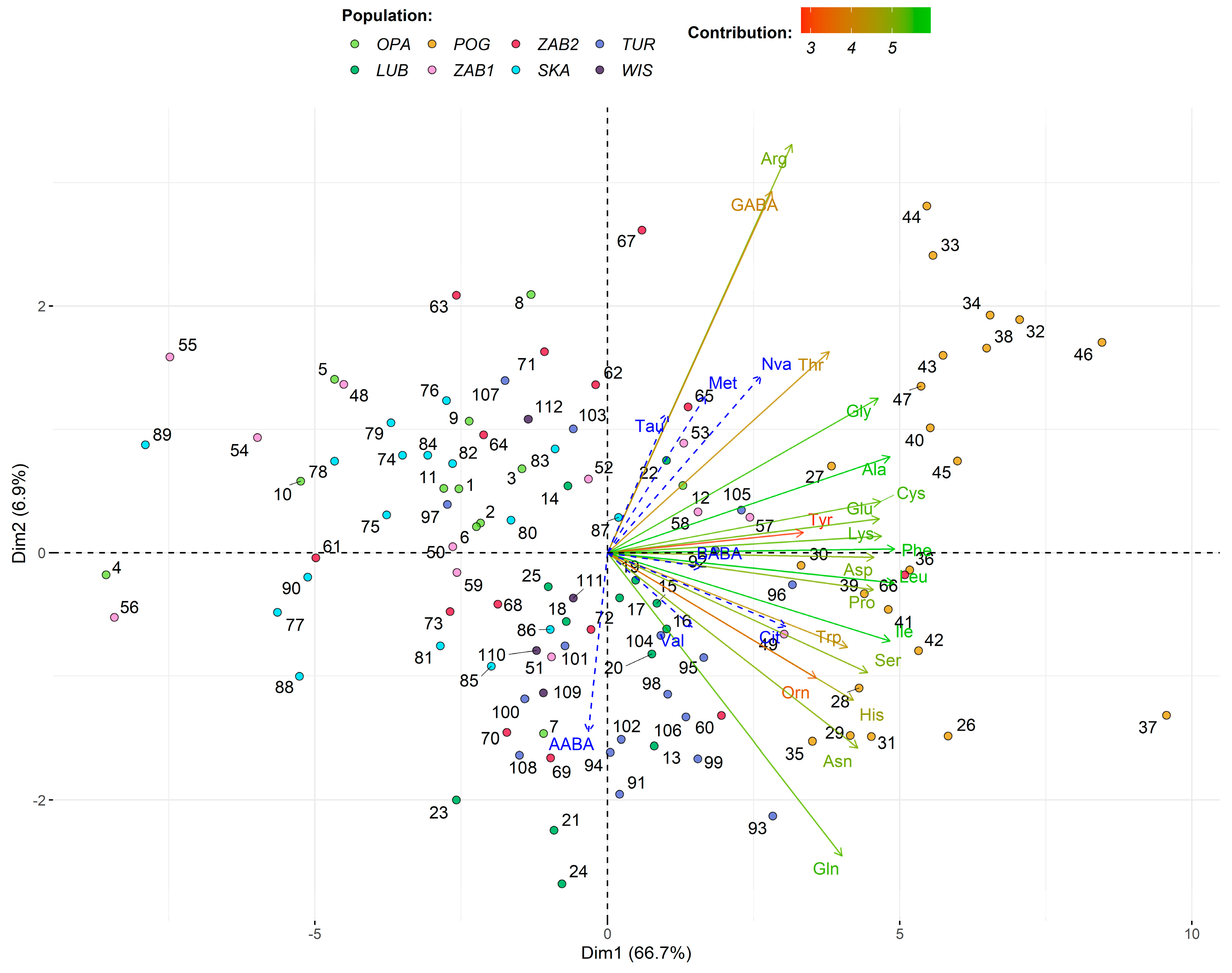
N. ovata nectar is rich in amino acids, we noted 28 distinct AAs (20 proteogenic and eight non-proteogenic), and 26 were common for all populations. In the nectar of specialist orchids from the
Platanthera genus, 23 AAs were found in total, from nine to 20 in each population [
17]. Moreover, the nectar of other orchids was composed of the lower number of AAs—20 in
Gymnadenia conopsea [
28] and 17 in
Limodorum abortivum and
Epipactis atropurpurea [
29]. In three populations (OPA, LUB, and WIS), no relationship between RS and AAs was found, while in two others (TUR and SKA), only single statistically significant correlations were noted—the percentage of Cys positively influenced FRS in TUR, while in SKA negatively influenced FRS. In the last population, an increased amount of Gln increased FRS. In the remaining three populations (POG, ZAB1, and ZAB2) we noted a larger influence of AAs on RS; it concerned mainly FRS and almost all of the correlations were negative. Pollinia removal depended only on Pro in POG, on GABA and Cit in ZAB1, and on Val and Gly in ZAB2. The most abundant in all
N. ovata populations were Ala and Glu, but they weakly affected RS, having only a negative influence on FRS (Glu in SKA and POG, and Ala in ZAB1 and POG). Other AAs with a relatively high amount in
N. ovata nectar were Asp, Cys, Gly, Thr, Asn, Gln, Ile, Phe, and Pro among the group of proteogenic AAs, and Orn and GABA among non-proteogenic ones.
AAs in floral nectar are important for the survival of nectar-feeding animals [
14,
16,
32,
33,
97], although the role of particular AAs is poorly explained. It is known, for example, that one among the most abundant AAs in
N. ovata nectar (Ala) influences insects’ growth, while the second (Glu) affects pollinators’ behavior [
28] similarly to Leu and Met [
12]. On the other hand, Venjakob, et al. [
61] found that Ala and Gly may deter honeybees. In the case of
N. ovata, the second function of Ala is more probable, as we found a negative correlation of this AA with RS. One of the most common AAs in plant nectar is Pro, which rewards pollinators and acts as a propellant for the lift phase of the flight [
98,
99]. It triggers the normal insects’ salt-receptor neurons, which initiates feeding [
19,
23,
97]. Its accumulation is also interpreted as a plant’s answer to stress factors [
98]. Pro was present in all populations studied with a quite high amount, but only in POG did it positively correlated with PR, and in ZAB2 with FRS. Two other AAs (Asp and Thr), which belong to the most abundant AAs in
N. ovata nectar, seem negatively correlated with FRS in ZAB1, and are known as general repellents [
14]. Moreover, Glu, Leu, and Met play a potential role in parasitoid rejection [
12]. One of the two most abundant non-proteogenic AAs in
N. ovata nectar, GABA, influences the insect nervous system and muscle activity [
11,
100]. Its higher amount was connected with a decrease in both PR and FRS in ZAB1. An interesting result was observed in POG, where a higher amount of this AA was negatively correlated with fruiting, while its percentage was positively correlated with FRS. This indicates that not only the amount of a particular AA, but also relationships between them, may be important in shaping plant-pollinator interaction. BABA, although a less common AA in nature than GABA, was present in all
N. ovata populations with a relatively high amount. It contributes to protecting plants from pathogens [
101,
102]. One of the important nectar traits is its taste, which attracts or discourages visitors and depends on some AAs [
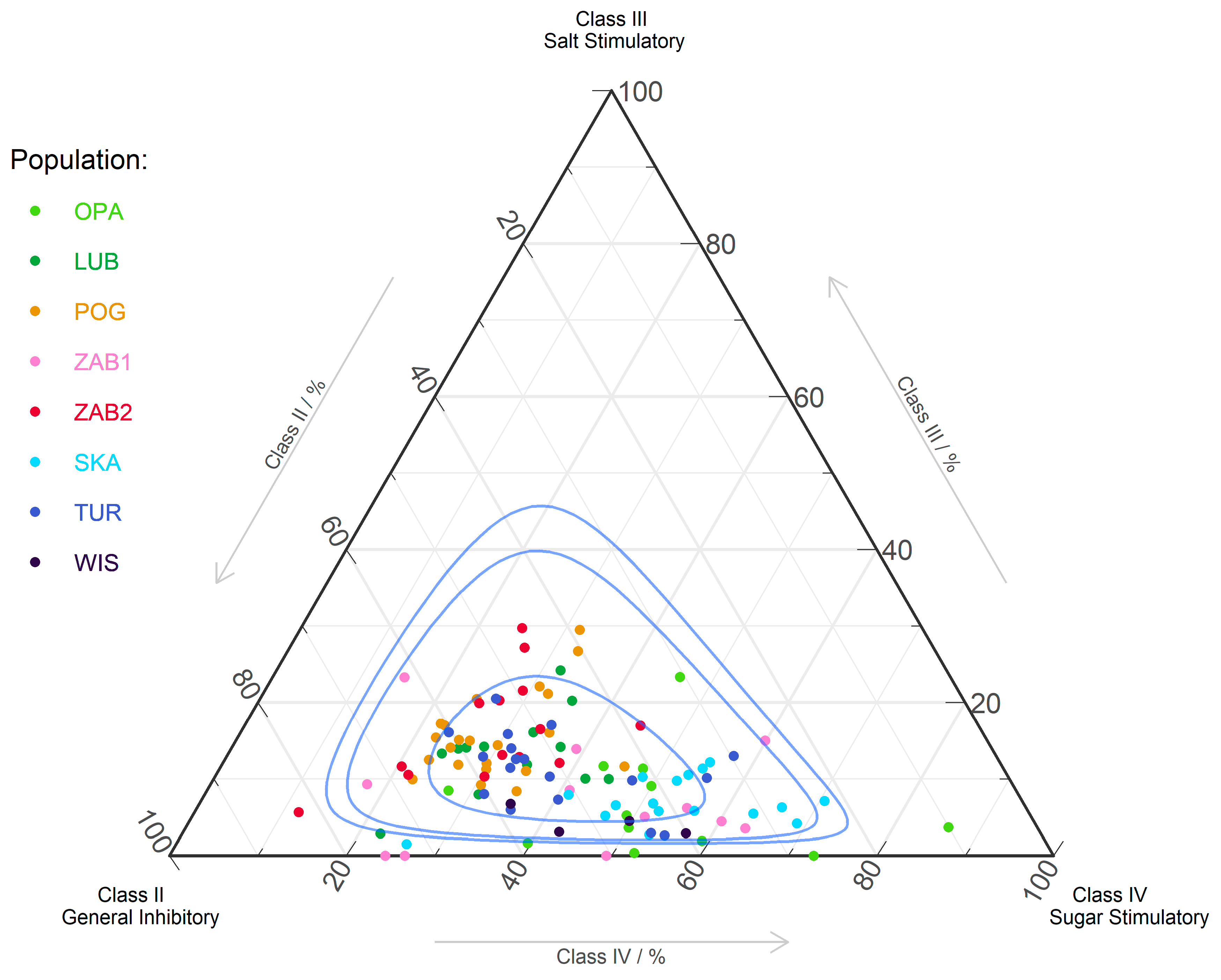
27,
28,
103]. AA compositions influence pollinator taste perception and pollinating behavior through specific neurological or phago-stimulating pathways [
12]. Some authors suggest that the taste function is even more important than the nutritive value [
34,
35]. We observed a potential influence of the amount of AAs from a particular taste group only in POG and ZAB1. The positive correlation between taste group IV and PR was noted only in POG. In the remaining cases, nectar taste could shape FRS, always in a negative way. This may indicate that insects present in the majority of
N. ovata populations were not sensitive to nectar taste or did not prefer this taste. Similar results were obtained for other nectariferous orchids [
17,
27].
We found inter-population variation in flower structure and the amount of particular nectar components, similar to other studies [
19,
27,
41,
55,
56,
58,
79]. One of the sources of this variability is differences in soils in which
N. ovata grows. Soil properties influenced mainly nectar composition. Production of nectar is costly, even to 30% of flower costs [
104]; thus, it requires adequate soil resources. Our studies suggest that more important than the participation of particular chemical elements in the soil is their proportionality. The most important in shaping nectar character were the C:N and N:P ratios. In POG, where C:N was the highest, the sum of sugars was 1.5–2 times higher, and the amount of AAs was three to even almost eight times higher than in other populations. The increase of the C:N ratio in soil caused a higher sum of sugars, glucose, and fructose as well as some AAs (Asp, Glu, Asn, Ser, Trp, and His). Simultaneously, the increase in the same soil characteristic had a negative effect on the percentage of Gly, Thr, Tau, Met, and GABA. A higher N:P ratio negatively influenced the total amount of sugars and fructose and the percentage of Glu and Tyr, while it increased the percentage of Cys and Ile. The importance of soil traits for nectar traits or plant condition was noted by other authors [
27,
28,
34,
39,
69].
It should be noted that the other factors, such as weather conditions, may shape plant properties and pollinators’ assemblages and their activity [
12,
81,
96]. The weather condition in our studies differed between seasons: 2020 was rainier than 2019. It could cause that higher values characterized some plant parameters in the second year of the study. In 19 out of 63 cases, we noted statistically significant changes between years; in 15 cases, we observed an increase of these values (3 cases of floral display and 11 of flower traits). On the other hand, in 2020, in most cases, the level of fruiting was lower. The explanation of year-to-year changes of plant traits due to weather should be undertaken with caution because only some traits and only in some populations differed between seasons. Moreover, in neighboring populations in the Biebrza National Park, the same traits often changed in opposite directions from year to year. It can indicate the greater role of other factors than the weather in these changes.
In our study, we tried to answer the question: In which way is
N. ovata adapted to a wide range of pollinators? As a generalist with reference to pollinators,
N. ovata depends on an exceptionally high number of insects in the pollination process—almost 300 species of visitors, among which at least 50 species attached pollinia [
76]. How can the demands of such a wide range of insects, differing in sizes, mouth apparatus, nutritional needs, and behaviors, be met? The answer is simple: The plants’ offer should also be wide. In the case of
N. ovata, this rich offer includes the wide range of nectar components (e.g., a large number of AAs) and differentiation of their amounts as well as the variability of flower structures. This indicates that this species did not evolve flower traits, which filter flower visitors; thus, they are not dedicated to a certain group of pollinators. Generally, our results fit the generalistic character of
N. ovata, but the level of generalization at the species level seems to be higher than at the population level. The lack of, or poorly matched, interactions between flower structure and nectar chemistry and the levels of RS in
N. ovata populations from northeast Poland confirm this statement. This finding is in agreement with the results of studies on other generalists [
90]. Jacquemyn and Brys [
105] found that a large variation in flower traits in
Orchis purpurea populations is maintained by the lack of strong selection pressures on these traits. Differentiation of flower traits enables pollination by whatever flower visitors have a suitable size and appropriate behavior. The probability that whatever species among almost 300
N. ovata visitors will serve as an effective pollinator is quite high. However, the variation of FRS among populations suggests that despite the high number of potential pollinators of this species, their abundance in particular populations was extremely differentiated. A low level of fruiting in some populations and a high ratio of PR:FRS (especially in SKA) indicates pollinators’ deficiency. In SKA, the problem with pollinators is deeper due to anthropogenic impact. It exists in disturbed and fragmented habitats in a restricted area, less abundant in plants, being hosts of
N. ovata visitors. Significantly lower PR in such populations may reflect unsuitable conditions for insects. The high levels of fruiting in populations from Biebrza National Park resulted from the relatively unchanged environment in this area. Natural habitats are suitable for many plant species connected to insects pollinating
N. ovata flowers. The higher RS in populations from BNP could also be a result of their larger sizes in comparison to others. The minimum population size is often required to attract sufficient pollinators. This assumption is supported by the results of Brys, et al. [
79] study, which found a significant relationship between RS and population size in
N. ovata.
Our results contribute to the knowledge about the reproductive strategy of
N. ovata and fit into studies that explain the causes and consequences of generalization in plants. However, in the course of this study, new questions arose, which required further analysis. For example, why does this orchid invest so many resources into nectar production if it is not an effective allurement of insects, as in other nectariferous species, and which nectar components are the most important for its fitness? The yellow-green color of
N. ovata flowers does not attract pollinators because it does not contrast with the surrounding vegetation. The color purity is typical of many generalist insect-pollinated plants [
71]. In such cases, other flower traits (odor and nectar) play a key role. Floral nectar (its concentration and composition) is rarely detectable by a pollinator at a distance [
11]. The fragrance is a key floral attractant for most wasps and beetles—the insect groups that are pollinators
of N. ovata—and also other generalistic plants accompanying this orchid; also, they often possess the same odor compounds [
76,
95,
96,
106]. A probable scenario is that at the first step insects are attracted by the fragrance emitted by
N. ovata flowers, which can explain the high level of PR; however, after probing nectar, which seems tasteless to the visitors, they do not further penetrate the flowers, thus causing a decrease in FRS. If so, why does
N. ovata produce such ineffectual rewards? The answer partially explains Johnson and Hobbhahn [
71] hypothesis that generalist pollination in orchids comes with high reproductive costs. According to the authors, these higher costs also include pollinia losses and inefficiency of pollination, characteristic of most orchids.
Supposing that, in the evolution process, all acts are intentional/on purpose, these high costs also contribute to N. ovata fitness. Each of the flower traits developed in this orchid is equally important in shaping RS, even those that seem to be negligible in our studies. They may operate side by side as “comprehensive consumer infinity” for pollinators. It seems that the wide flower variability and complexity of their action is the advantage of this species, which enables the maintenance of populations under different environmental conditions. The results of our studies also have conservation implications; protection of this orchid requires the protection of its wide spectrum of insect partners and their hosts and, thus, the entire habitats in which N. ovata exists.



 ,
, 





{kind=link}
{kind=link}
{kind=link}