
Advances in Sensing, Response and Regulation Mechanism of Salt Tolerance in Rice



Abstract
:1. Background
2. Salt Stress Sensing
2.1. Osmosensing
2.2. Na+ Sensing
3. Signal Transduction
3.1. CaM/CML
3.2. CDPK
3.3. Calcineurin B-Like Protein (CBL)/CIPK
3.4. MAPK
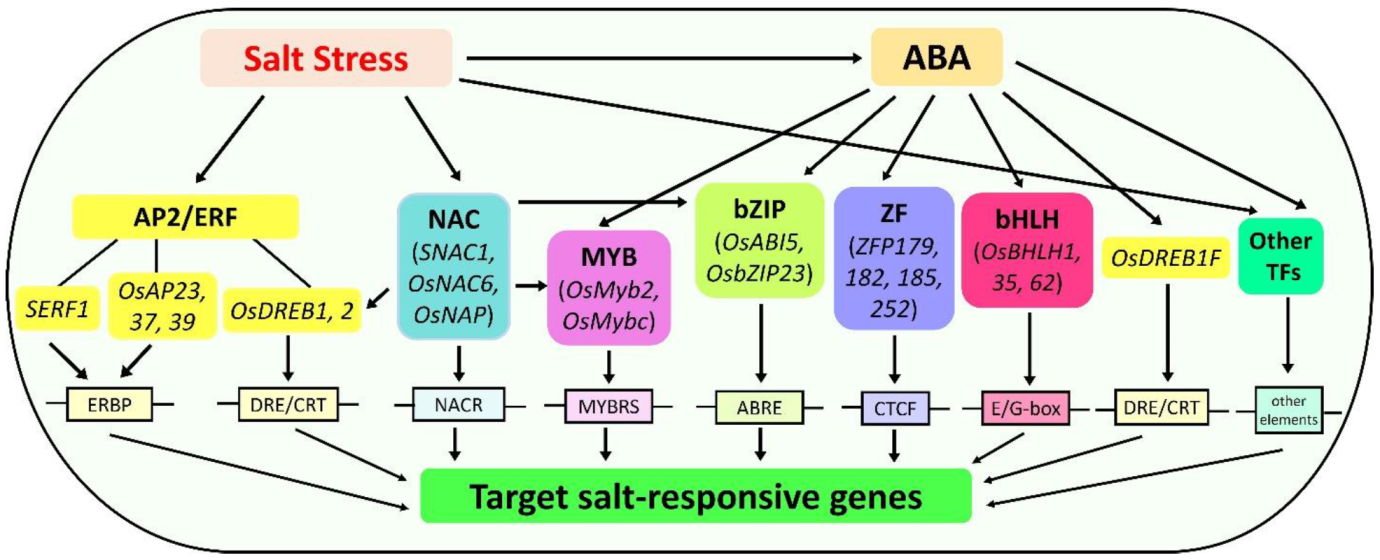
4. Transcriptional Regulation
4.1. APETALA2/Ethylene Responsive Factor (AP2/ERF) Regulation
4.2. NAC Regulation
4.3. MYB Regulation
4.4. bZIP Regulation
4.5. ZF Regulation
4.6. bHLH Regulation
4.7. Other TFs Involved in Salt Tolerance
5. Salt Tolerance Adaptive Mechanisms
5.1. Osmoprotection and Osmoregulation
5.1.1. Osmolytes
5.1.2. Water Channel Aquaporins
5.2. Stomatal Closure
5.3. Antioxidation
5.3.1. Superoxide Dismutase (SOD)
5.3.2. Catalase (CAT)
5.3.3. Ascorbate Peroxidase (APX)
5.3.4. Glutathione Reductase (GR)
5.3.5. Thioredoxin (TRX) and Glutaredoxin (GRX)
5.4. Na+ Exclusion and Sequestration
5.4.1. Na+ Efflux
5.4.2. Na+ Loading and Unloading
5.4.3. Vacuolar Na+ Sequestration
5.5. Suberin Deposition
5.6. K+ Uptake
6. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AP2/ERF | APETALA2/ethylene responsive factor |
| APX AT | Ascorbate peroxidase Adenosine-thymine |
| bHLH | Basic-helix-loop-helix |
| Bzip CAMK | Basic leucine-zipper Calcium/calmodulin-dependent protein kinase |
| CAT | Catalase |
| CBL CDA | Calcineurin B-like protein Cytidine deaminase |
| CDPK | Calcium-dependent protein kinase |
| CIPK | CBL-interacting protein kinase |
| CaM | Calmodulin |
| CML | Calmodulin-like protein |
| CPP | Cysteine-rich poly comb-like protein |
| DRE/CRT | Dehydration-responsive element/c-repeat |
| DST | Drought and salt tolerance |
| DCA1 | DST co-activator 1 |
| GB | Glycine betaine |
| GR GUS | Glutathione reductase β-glucoronidase |
| HAK | High-affinity potassium transporter |
| HD-Zip | Homeodomain-leucine zipper |
| HKT | High-affinity K+ transporter |
| HpT | Histidine-containing phosphotransfer protein |
| LEA | Late embryogenesis abundant |
| LRR-RLK | Leucine-rich repeat-receptor-like kinase |
| MAPK | Mitogen-activated protein kinase |
| MKK | MAPK kinase |
| MKKK | MKK kinase |
| MKP | Mitogen-activated protein kinase phosphatase |
| MYB | Myeloblastosis |
| NAC | NAM, ATAF and CUC |
| NAM | No apical meristem |
| NHX | Na+/H+ antiporter |
| NIN | Nodule inception |
| OSCA | Ca2+ permeable stress-gated cation channels |
| PA | Phosphatidic acid |
| PIP QTL | Plasma membrane intrinsic protein Quantitative trait locus |
| RAV | Related to ABI3 and VP1 |
| RLK | Receptor-like kinase |
| ROS | Reactive oxygen species |
| SIT | Salt intolerance |
| SOD | Superoxide dismutase |
| SOS | Salt overly sensitive |
| TCP | Teosinte branched 1/cycloidea/proliferating cell |
| TF | Transcription factor |
| TIP | Tonoplast intrinsic |
| TPS | Trehalose-6-phosphate phosphatase |
| TPP | Trehalose-6-phosphate synthase |
| TRX | Thioredoxin |
| VP | Vacuolar-type H+-pyrophosphatase |
| ZF | Zinc finger |
References
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.; Shannon, M.C.; Grieve, C.M. Evaluation of salt tolerance in rice genotypes by multiple agronomic parameters. Euphytica 2002, 127, 235–245. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. Effects of salt stress on growth mineral nutrition and proline accumulation in relation to osmotic adjustment in rice (Oryza sativa L.) cultivars differing in salinity resistance. Plant Growth Regul. 1996, 19, 207–218. [Google Scholar] [CrossRef]
- Cui, H.; Takeoka, Y.; Wada, T. Effect of sodium chloride on the panicle and spikelet morphogenesis in rice. Jpn. J. Crop. Sci. 1995, 64, 593–600. [Google Scholar] [CrossRef] [Green Version]
- Khatun, S.; Flowers, T.J. Effects of salinity on seed set in rice. Plant Cell. Environ. 1995, 18, 61–67. [Google Scholar] [CrossRef]
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef]
- Tester, M.; Davenport, R.J. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, M.; Guo, R.; Shi, D.; Liu, B.; Lin, X.; Yang, C. Effects of salt stress on ion balance and nitrogen metabolism of old and young leaves in rice (Oryza sativa L.). BMC Plant Biol. 2012, 12, 194. [Google Scholar] [CrossRef] [Green Version]
- Li, C.H.; Wang, G.; Zhao, J.L.; Zhang, L.Q.; Ai, L.F.; Han, Y.F.; Sun, D.Y.; Zhang, S.W.; Sun, Y. The receptor-like kinase SIT1 mediates salt sensitivity by activating MAPK3/6 and regulating ethylene homeostasis in rice. Plant Cell 2014, 26, 2538–2553. [Google Scholar] [CrossRef] [Green Version]
- Boonburapong, B.; Buaboocha, T. Genome-wide identification and analyses of the rice calmodulin and related potential calcium sensor proteins. BMC Plant Biol. 2007, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Campo, S.; Baldrich, P.; Messeguer, J.; Lalanne, E.; Coca, M.; San Segundo, B. Overexpression of a calcium-dependent protein kinase confers salt and drought tolerance in rice by preventing membrane lipid peroxidation. Plant Physiol. 2014, 165, 688–704. [Google Scholar] [CrossRef] [Green Version]
- Saijo, Y.; Hata, S.; Kyozuka, J.; Shimamoto, S.; Izui, K. Over-expression of a single Ca2+-dependent protein kinase confers both cold and salt/drought tolerance on rice plants. Plant J. 2000, 23, 319–327. [Google Scholar] [CrossRef]
- Saijo, Y.; Kinoshita, N.; Ishiyama, K.; Hata, S.; Kyozuka, J.; Hayakawa, T.; Nakamura, T.; Shimamoto, K.; Yamaya, T.; Izui, K. A Ca2+-dependent protein kinase that endows rice plants with cold- and salt-stress tolerance functions in vascular bundles. Plant Cell. Physiol. 2001, 42, 1228–1233. [Google Scholar] [CrossRef] [Green Version]
- Asano, T.; Hayashi, N.; Kobayashi, M.; Aoki, N.; Miyao, A.; Mitsuhara, I.; Ichikawa, H.; Komatsu, S.; Hirochika HKikuchi, S.; Ohsugi, R. A rice calcium-dependent protein kinase OsCPK12 oppositely modulates salt-stress tolerance and blast disease resistance. Plant J. Cell. Mol. Biol. 2011, 69, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Hakata, M.; Nakamura, H.; Aoki, N.; Ichikawa, H.; Komatsu, S.; Hirochika, H.; Kikuchi, S.; Ohsugi, R. Functional characterisation of OsCPK21 a calcium-dependent protein kinase that confers salt tolerance in rice. Plant Mol. Biol. 2011, 75, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.X.; Zhou, X.J.; Chang, S.; Chu, Z.L.; Wang, H.M.; Han, S.C.; Wang, Y.D. Calcium-dependent protein kinase 21 phosphorylates 14-3-3 proteins in response to ABA signaling and salt stress in rice. Biochem. Biophy. Res. Commun. 2017, 4, 1450–1456. [Google Scholar] [CrossRef]
- Xiang, Y.; Huang, Y.M.; Xiong, L.Z. Characterization of stress-responsive CIPK genes in rice for stress tolerance improvement. Plant Physiol. 2007, 144, 1416–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piao, H.L.; Xuan, Y.H.; Park, S.H.; Je, B.I.I.; Park, S.J.; Park, S.H.; Kim, C.H.; Huang, J.; Wang, G.K.; Kim, M.J.; et al. OsCIPK31 a CBL-interacting protein kinase is involved in germination and seedling growth under abiotic stress conditions in rice plants. Mol. Cells 2010, 30, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.Z.; Yang, Y.N. Disease resistance and abiotic stress tolerance in rice are inversely modulated by an abscisic acid–inducible mitogen-activated protein kinase. Plant Cell 2003, 15, 745–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.K.; Kim, B.G.; Kwon, T.R.; Jeong, M.J.; Park, S.R.; Lee, J.W.; Byun, M.O.; Kwon, H.B.; Matthews, B.F.; Hong, C.B.; et al. Overexpression of the mitogen-activated protein kinase gene OsMAPK33 enhances sensitivity to salt stress in rice (Oryza sativa L.). J. Biosci. 2011, 36, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.Z.; Jing, W.; Zhang, W.H. The mitogen-activated protein kinase cascade MKK1–MPK4 mediates salt signaling in rice. Plant Sci. 2014, 227, 181–189. [Google Scholar] [CrossRef]
- Kumar, K.; Sinha, A.K. Overexpression of constitutively active mitogen activated protein kinase kinase 6 enhances tolerance to salt stress in rice. Rice 2013, 6, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Na, Y.J.; Choi, H.K.; Park, M.Y.; Choi, S.W.; Vo, K.T.X.; Jeon, J.S.; Kim, S.Y. OsMAPKKK63 is involved in salt stress response and seed dormancy control. Plant Signal Behav. 2019, 14, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubouzet, J.G.; Sakuma, Y.; Ito, Y.; Kasuga, M.; Dubouzet, E.G.; Miura, S.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. OsDREB genes in rice Oryza sativa L., encode transcription activators that function in drought-, high-salt- and cold-responsive gene expression. Plant J. 2003, 33, 751–763. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, C.; Jin, X.F.; Xiong, A.S.; Peng, R.H.; Hong, Y.H.; Yao, Q.H.; Chen, J.M.; Chen, J.M. Expression of a rice DREB1 gene, OsDREB1D, enhances cold and high-salt tolerance in transgenic Arabidopsis. BMB Rep. 2009, 42, 486–492. [Google Scholar] [CrossRef]
- Wang, Q.; Guan, Y.; Wu, Y.; Chen, H.; Chen, F.; Chu, C. Overexpression of a rice OsDREB1F gene increases salt drought and low temperature tolerance in both Arabidopsis and rice. Plant Mol. Biol. 2008, 67, 589–602. [Google Scholar] [CrossRef]
- Cui, M.; Zhang, W.; Zhang, Q.; Xu, Z.; Zhu, Z.; Duan, F.; Wu, R. Induced over-expression of the transcription factor OsDREB2A improves drought tolerance in rice. Plant Physiol. Biochem. 2011, 49, 1384–1391. [Google Scholar] [CrossRef]
- Mallikarjuna, G.; Mallikarjuna, K.; Reddy, M.K.; Kaul, T. Expression of OsDREB2A transcription factor confers enhanced dehydration and salt stress tolerance in rice (Oryza sativa L.). Biotechnol. Lett. 2011, 33, 1689–1697. [Google Scholar] [CrossRef]
- Matsukura, S.; Mizoi, J.; Yoshida, T.; Todaka, D.; Ito, Y.; Maruyama, K.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Comprehensive analysis of rice DREB2-type genes that encode transcription factors involved in the expression of abiotic stress-responsive genes. Mol. Genet. Genom. 2010, 283, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.; Jiang, H.H.; Wang, F.; Peng, R.H.; Yao, Q.H.; Xiong, A.S. A Rice OsAP23, functioning as an AP2/ERF transcription factor, reduces salt tolerance in transgenic Arabidopsis. Plant Mol. Biol. Rep. 2013, 31, 1336–1345. [Google Scholar] [CrossRef]
- Oh, S.J.; Kim, Y.S.; Kwon, C.W.; Park, H.K.; Jeong, J.S.; Kim, J.K. Overexpression of the transcription factor AP37 in rice improves grain yield under drought conditions. Plant Physiol. 2009, 150, 1368–1379. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.X.; Wang, J.; Zhao, X.Q.; Yang, S.; Huang, L.Y.; Du, F.P.; Li, Z.K.; Zhao, X.Q.; Fu, B.Y.; Wang, W. Overexpression of the transcription factor gene OsSTAP1 increases salt tolerance in rice. Rice 2020, 50, 1–12. [Google Scholar] [CrossRef]
- Ke, Y.G.; Yang, Z.J.; Yu, S.W.; Li, T.F.; Wu, J.H.; Gao, H.; Fu, Y.P.; Luo, L.J. Characterization of OsDREB6 responsive to osmotic and cold stresses in rice. J. Plant Biol. 2014, 57, 150–161. [Google Scholar] [CrossRef]
- Schmidt, R.; Mieulet, D.; Hubberten, H.M.; Obata, T.; Hoefgen, R.; Fernie, A.R.; Fisahn, J.; San Segundo, B.; Guiderdoni, E.; Schippers, J.H.M.; et al. Salt-responsive ERF1 regulates reactive oxygen species-dependent signaling during the initial response to salt stress in rice. Plant Cell 2013, 25, 2115–2131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.F.; Chen, X.J.; Liu, J.Q.; Ye, J.C.; Guo, Z.J. The rice ERF transcription factor OsERF922 negatively regulates resistance to Magnaporthe oryzae and salt tolerance. J. Exp. Bot. 2012, 63, 3899–3911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Y.B.; Li, J.; Qin, R.Y.; Xu, R.F.; Li, H.; Yang, Y.C.; Ma, H.; Li, L.; Weng, P.C.; Yang, J.B. Identification of a regulatory element responsible for salt induction of rice OsRAV2 through ex situ and in situ promoter analysis. Plant Mol. Biol. 2016, 90, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Y.F.; Lv, B.; Li, J.; Luo, L.Q.; Lu, S.C.; Zhang, X.; Ma, H.; Ming, F. The NAC family transcription factor OsNAP confers abiotic stress response through the ABA pathway. Plant Cell. Physiol. 2016, 55, 604–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Y.B.; Zhang, H.J.; Huang, L.; Li, D.Y.; Song, F.M. Overexpression of a stress-responsive NAC transcription factor gene ONAC022 improves drought and salt tolerance in rice. Front. Plant Sci. 2016, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.N.; Chen, B.; Lu, G.J.; Han, B. Overexpression of a NAC transcription factor enhances rice drought and salt tolerance. Biochem. Biophys. Res. Commun. 2009, 379, 985–989. [Google Scholar] [CrossRef]
- Yokotani, N.; Ichikawa, T.; Kondou, Y.; Matsui, M.; Hirochika, H.; Iwabuchi, M.; Oda, K. Tolerance to various environmental stresses conferred by the salt-responsive rice gene ONAC063 in transgenic Arabidopsis. Planta 2009, 229, 1065–1075. [Google Scholar] [CrossRef] [PubMed]
- Sarukaba, Y.; Piao, W.L.; Lim, J.H.; Han, S.H.; Kim, Y.S.; An, G.; Paek, N.C. Rice ONAC106 inhibits leaf senescence and increases salt tolerance and tiller angle. Plant Cell. Physiol. 2015, 56, 2325–2339. [Google Scholar] [CrossRef]
- Mao, C.J.; Ding, J.L.; Zhang, B.; Xi, D.D.; Ming, F. OsNAC2 positively affects salt-induced cell death and binds to the OsAP37 and OsCOX11 promoters. Plant J. 2018, 94, 454–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, D.G.; Zhou, L.Y.; Chen, W.T.; Ye, N.H.; Xia, J.X.; Zhuang, C.X. Overexpression of a microRNA-targeted NAC transcription factor improves drought and salt tolerance in rice via ABA-mediated pathways. Rice 2019, 12, 76. [Google Scholar] [CrossRef] [PubMed]
- Takasaki, H.; Maruyama, K.; Kidokor, S.; Ito, Y.; Fujita, Y.; Shinozaki, K.; Yamaguchi-Shinozaki, K.; Nakashima, K. The abiotic stress-responsive NAC-type transcription factor OsNAC5 regulates stress-inducible genes and stress tolerance in rice. Mol. Genet. Genom. 2010, 284, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Song, S.Y.; Chen, Y.; Chen, J.; Dai, X.Y.; Zhang, W.H. Physiological mechanisms underlying OsNAC5-dependent tolerance of rice plants to abiotic stress. Planta 2011, 234, 331–345. [Google Scholar] [CrossRef]
- Nakashima, K.; Tran, L.S.P.; Nguyen, D.V.; Fujita, M.; Maruyama, K.; Todaka, D.; Ito, Y.; Hayashi, N.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of a NAC-type transcription factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice. Plant J. 2007, 51, 617–630. [Google Scholar] [CrossRef]
- Hu, H.H.; You, J.; Fang, Y.J.; Zhu, X.Y.; Qi, Z.Y.; Xiong, L.Z. Characterization of transcription factor gene SNAC2 conferring cold and salt tolerance in rice. Plant Mol. Biol. 2010, 67, 169–181. [Google Scholar] [CrossRef]
- Hu, H.H.; Dai, M.Q.; Yao, J.L.; Xiao, B.Z.; Li, X.H.; Zhang, Q.F.; Xiong, L.Z. Overexpressing a NAM ATAF and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc. Natl. Acad. Sci. USA 2006, 103, 12987–12992. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.S.; Kim, Y.K.; Baek, K.H.; Jung, H.; Ha, S.H.; Choi, Y.D.; Kim, M.; Reuzeau, C.; Kim, J.K. Root-specific expression of OsNAC10 improves drought tolerance and grain yield in rice under field drought conditions. Plant Physiol. 2010, 153, 185–197. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Zhong, Z.H.; Hang, H.H.; Wang, X.; Liu, B.L.; Yang, L.J.; Han, X.Y.; Yu, D.S.; Zheng, X.L.; Wang, C.G.; et al. Targeted mutagenesis of NAC transcription factor gene OsNAC041 leading to salt sensitivity in rice. Rice 2019, 26, 98–108. [Google Scholar] [CrossRef]
- Yang, A.; Dai, X.Y.; Zhang, W.H. A R2R3-type MYB gene, OsMYB2, is involved in salt, cold, and dehydration tolerance in rice. J. Exp. Bot. 2012, 63, 2541–2556. [Google Scholar] [CrossRef]
- Dai, X.; Xu, Y.; Ma, Q.; Xu, W.; Wang, T.; Xue, Y.; Chong, K. Overexpression of an R1R2R3 MYB gene, OsMYB3R-2, increases tolerance to freezing, drought, and salt stress in transgenic Arabidopsis. Plant Physiol. 2007, 143, 1739–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, H.Y.; Li, J.J.; Liu, P.L.; Duan, J.Z.; Zhao, Y.; Guo, X.; Li, Y.; Zhang, H.L.; Ali, J.; Li, Z.C. Overexpression of OsMYB48-1 a novel MYB-related transcription factor enhances drought and salinity tolerance in rice. PLoS ONE 2014, 9, e92913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, R.; Schippers, J.H.M.; Mieulet, D.; Obata, T.; Fernie, A.R.; Guiderdoni, E.; Mueller-Roeber, B. MULTIPASS, a rice R2R3-type MYB transcription factor, regulates adaptive growth by integrating multiple hormonal pathways. Plant J. 2013, 76, 258–273. [Google Scholar] [CrossRef]
- Zhu, N.; Cheng, S.F.; Liu, X.Y.; Du, H.; Dai, M.Q.; Zhou, D.X.; Yang, W.J.; Zhao, Y. The R2R3-type MYB gene OsMYB91 has a function in coordinating plant growth and salt stress tolerance in rice. Plant Sci. 2015, 236, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Jing, W.; Xiao, L.Y.; Jin, Y.K.; Shen, L.K.; Zhang, W.H. The rice High-Affinity Potassium Transporter1;1 is involved in salt tolerance and regulated by an MYB-type transcription factor. Plant Physiol. 2015, 168, 1076–1090. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.A.; Cho, J.; Han, M.; Ahn, C.H.; Jeon, J.S.; An, G.; Park, P.B. The ABRE-binding bZIP transcription factor OsABF2 is a positive regulator of abiotic stress and ABA signaling in rice. J. Plant Physiol. 2010, 167, 1512–1520. [Google Scholar] [CrossRef]
- Zou, M.J.; Guan, Y.C.; Ren, H.B.; Zhang, F.; Chen, F. A bZIP transcription factor, OsABI5, is involved in rice fertility and stress tolerance. Plant Mol. Biol. 2008, 66, 675–683. [Google Scholar] [CrossRef]
- Xiang, Y.; Tang, N.; Du, H.; Ye, H.Y.; Xiong, L.Z. Characterization of OsbZIP23 as a key player of the basic leucine zipper transcription factor family for conferring abscisic acid sensitivity and salinity and drought tolerance in rice. Plant Physiol. 2008, 148, 1938–1952. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.T.; Mao, B.G.; Ou, S.J.; Wang, W.; Liu, L.C.; Wu, Y.B.; Chu, C.C.; Wang, X.P. OsbZIP71, a bZIP transcription factor, confers salinity and drought tolerance in rice. Plant Mol. Biol. 2014, 89, 19–36. [Google Scholar] [CrossRef]
- Das, P.; Lakra, N.; Nutan, K.K.; Singla-Pareek, S.L.; Pareek, A. A unique bZIP transcription factor imparting multiple stress tolerance in rice. Rice 2019, 12, 58. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.Y.; Chao, D.Y.; Gao, P.J.; Zhu, M.Z.; Shi, M.Z.; Lin, H.X. A previously unknown zinc finger protein, DST, regulates drought and salt tolerance in rice via stomatal aperture control. Genes Dev. 2009, 23, 1805–1817. [Google Scholar] [CrossRef] [Green Version]
- Jan, A.M.K.; Todaka, D.; Kidokoro, S.; Abo, M.; Yoshimura, E.; Shinozaki, K.; Nakashima, K.; Yamaguchi-Shinozaki, K. OsTZF1, a CCCH-tandem zinc finger protein, confers delayed senescence and stress tolerance in rice by regulating stress-related genes. Plant Physiol. 2013, 161, 1202–1216. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.J.; Guo, S.Q.; Yang, X.; Bao, Y.M.; Tang, H.J.; Sun, H.; Huang, J.; Zhang, H.S. Functional analysis of a novel Cys2/His2-type zinc finger protein involved in salt tolerance in rice. J. Exp. Bot. 2010, 61, 2807–2818. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Sun, S.J.; Xu, D.Q.; Lan, H.X.; Sun, H.; Wang, Z.F.; Bao, Y.M.; Wang, J.F.; Tang, H.J.; Zhang, H.S. A TFIIIA-type zinc finger protein confers multiple abiotic stress tolerances in transgenic rice (Oryza sativa L.). Plant Mol. Biol. 2012, 80, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lan, H.X.; Shao, Q.L.; Wang, R.Q.; Chen, H.; Tang, H.J.; Zhang, H.S.; Huang, J. An A20/AN1-type zinc finger protein modulates gibberellins and abscisic acid contents and increases sensitivity to abiotic stress in rice (Oryza sativa). J. Exp. Bot. 2016, 67, 315–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.Q.; Huang, J.; Guo, S.Q.; Yang, X.; Bao, Y.M.; Tang, H.J.; Zhang, H.S. Overexpression of a TFIIIA-type zinc finger protein gene ZFP252 enhances drought and salt tolerance in rice (Oryza sativa L.). FEBS Lett. 2008, 582, 1037–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, Q.J.; Ma, H.Y.; Zhang, Z.J.; Wang, Z.Y.; Bu, Q.Y.; Liu, S.K. A rice LSD1-like-type ZFP gene OsLOL5 enhances saline-alkaline tolerance in transgenic Arabidopsis thaliana, yeast and rice. BMC Genom. 2016, 17, 142. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Li, F.; Ma, Y.; Ching, K.; Xu, Y. Overexpression of OrbHLH001, a putative helix-loop-helix transcription factor, causes increased expression of AKT1 and maintains ionic balance under salt stress in rice. J. Plant Physiol. 2013, 170, 93–100. [Google Scholar] [CrossRef]
- Chen, H.C.; Cheng, W.H.; Hong, C.Y.; Chang, Y.S.; Chang, M.C. The transcription factor OsbHLH035 mediates seed germination and enables seedling recovery from salt stress through ABA-dependent and ABA-independent pathways respectively. Rice 2018, 11, 50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.X.; Haider, I.; Kohlen, W.; Jiang, L.; Bouwmeester, H.; Meijer, A.H.; Schluepmann, H.; Liu, C.M.; Ouwerkerk, P.B.F. Function of the HD-Zip I gene Oshox22 in ABA-mediated drought and salt tolerances in rice. Plant Mol. Biol. 2012, 80, 571–585. [Google Scholar] [CrossRef]
- Bang, S.W.; Lee, D.K.; Jung, H.; Chung, P.J.; Kim, Y.S.; Choi, Y.D.; Suh, J.W.; Kim, J.K. Overexpression of OsTF1L a rice HD-Zip transcription factor promotes lignin biosynthesis and stomatal closure that improves drought tolerance. Plant Biotechnol. J. 2019, 17, 118–131. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.Y.; Yu, C.Y.; Huang, L.L.; Wu, M.J.; Liu, B.H.; Liu, Y.H.; Song, G.; Liu, D.D.; Gan, Y.B. Overexpression of MADS-box transcription factor OsMADS25 enhances salt stress tolerance in Rice and Arabidopsis. Plant Growth Regul. 2020, 90, 163–171. [Google Scholar] [CrossRef]
- Tao, Z.; Kou, Y.J.; Liu, H.B.; Li, X.H.; Xiao, J.H.; Wang, S.P. OsWRKY45 alleles play different roles in abscisic acid signalling and salt stress tolerance but similar roles in drought and cold tolerance in rice. J. Exp. Bot. 2011, 62, 4863–4874. [Google Scholar] [CrossRef]
- Hasthanssombut, S.; Supaibulwatana, K.; Mii, M.; Nakamura, I. Genetic manipulation of Japonica rice using the OsBADH1 from Indica rice to improve salinity tolerance. Plant Cell Tissue Organ. Cult. 2011, 104, 79–89. [Google Scholar] [CrossRef]
- Tang, W.; Sun, J.Q.; Liu, J.; Liu, F.F.; Yan, J.; Guo, X.J.; Lu, B.R.; Liu, Y.S. RNAi-directed downregulation of betaine aldehyde dehydrogenase 1 (OsBADH1) results in decreased stress tolerance and increased oxidative markers without affecting glycine betaine biosynthesis in rice (Oryza sativa). Plant Mol. Biol. 2014, 86, 443–454. [Google Scholar] [CrossRef]
- Ge, L.F.; Chao, D.Y.; Shi, M.; Zhu, M.Z.; Gao, J.P.; Lin, H.X. Overexpression of the trehalose-6-phosphate phosphatase gene OsTPP1 confers stress tolerance in rice and results in the activation of stress responsive genes. Planta 2008, 228, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Li, H.W.; Zhang, B.S.; Deng, X.W.; Wang, X.P. Overexpression of the trehalose-6-phosphate synthase gene OsTPS1 enhances abiotic stress tolerance in rice. Planta 2011, 234, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Vishal, B.; Krishnamurthy, P.; Ramamoorthy, R.; Kumar, P.P. OsTPS8 controls yield-related traits and confers salt stress tolerance in rice by enhancing suberin deposition. New Phytol. 2018, 221, 1369–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Wang, Z.Y.; Lin, H.; Cui, W.E.; Chen, J.; Liu, M.; Chen, Z.L.; Qu, L.J.; Gu, H. Expression and functional analysis of the rice plasma-membrane intrinsic protein gene family. Cell Res. 2006, 16, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.W.; Fukumoto, T.; Matsumoto, T.; Gena, P.; Frascaria, D.; Kaneko, T.; Katsuhara, M.; Zhong, S.H.; Sun, X.L.; Zhu, Y.M.; et al. Aquaporin OsPIP1;1 promotes rice salt resistance and seed germination. Plant Physiol. Biochem. 2013, 63, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.Q.; Sheng, P.K.; Tan, J.J.; Chen, X.L.; Lu, G.W.; Ma, W.W.; Heng, Y.Q.; Lin, Q.B.; Zhu, S.S.; Wang, J.L.; et al. Plasma membrane receptor-like kinase leaf panicle 2 acts downstream of the DROUGHT AND SALT TOLERANCE transcription factor to regulate drought sensitivity in rice. J. Exp. Bot. 2015, 66, 271–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, J.; Zong, W.; Li, X.K.; Ning, J.; Hu, H.H.; Li, X.H.; Xiao, J.H.; Xiong, L.Z. The SNAC1-targeted gene OsSRO1c modulates stomatal closure and oxidative stress tolerance by regulating hydrogen peroxide in rice. J. Exp. Bot. 2013, 64, 569–583. [Google Scholar] [CrossRef] [Green Version]
- Guan, Q.; Liao, X.; He, M.; Li, X.; Wang, Z.; Ma, H.; Yu, S.; Liu, S. Tolerance analysis of chloroplast OsCu/Zn-SOD overexpressing rice under NaCl and NaHCO3 stress. PLoS ONE 2017, 12, e0186052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, Y.; Hibino, T.; Tanaka, A.; Kishitani, S.; Takabe, T.; Yokota, S.; Takabe, T. Salt tolerance of transgenic rice overexpressing yeast mitochondrial Mn-SOD in chloroplasts. Plant Sci. 1999, 148, 131–138. [Google Scholar] [CrossRef]
- Lu, Z.Q.; Liu, D.L.; Liu, S.K. Two rice cytosolic ascorbate peroxidases differentially improve salt tolerance in transgenic Arabidopsis. Plant Cell Rep. 2007, 26, 1909–1917. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.G.; Zhang, Q.A.; Wu, J.X.; Zheng, X.; Zheng, S.; Sun, X.H.; Qiu, Q.S.; Lu, T.G. Gene knockout study reveals that cytosolic ascorbate peroxidase 2 (OsAPX2) plays a critical role in growth and reproduction in rice under drought, salt and cold stresses. PLoS ONE 2013, 8, e57472. [Google Scholar] [CrossRef]
- Wu, T.M.; Lin, W.R.; Kao, Y.T.; Hsu, Y.T.; Yeh, C.H.; Hong, C.Y.; Kao, C.H. Identification and characterization of a novel chloroplast/mitochondria co-localized glutathione reductase 3 involved in salt stress response in rice. Plant Mol. Biol. 2013, 83, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.J.; Guo, Y. OsTRXh1 regulates the redox state of the apoplast and influences stress responses in rice. Plant Signal Behav. 2012, 7, 440–442. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Priya, P.; Jain, M. Modified expression of an auxin-responsive rice CC-type glutaredoxin gene affects multiple abiotic stress responses. Planta 2013, 238, 871–884. [Google Scholar] [CrossRef]
- Ning, X.; Sun, Y.; Wang, C.C.; Zhang, W.L.; Sun, M.H.; Hu, H.T.; Liu, J.Z.; Yang, L. A rice CPYC-type glutaredoxin OsGRX20 in protection against bacterial blight methyl viologen and salt stresses. Front. Plant Sci. 2018, 9, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, M.T.; Bandillo, N.; Razzaq, F.; Shiblawi, A.; Sharma, S.; Liu, K.; Schmitz, A.J.; Zhang, C.; Véry, A.A.; Lorenz, A.J.; et al. Allelic variants of OsHKT1;1 underlie the divergence between indica and japonica subspecies of rice (Oryza sativa) for root sodium content. PloS Genet. 2017, 13, e1006823. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Kobayashi, N.I.; Tanoi, K.; Ma, J.F.; Itou, Y.; Katsuhara, M.; Itou, T.; Horie, T. T-DNA tagging-based gain-of-function of OsHKT1;4 reinforces Na+ exclusion from leaves and stems but triggers Na toxicity in roots of rice under salt stress. Int. J. Mol. Sci. 2018, 19, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Z.H.; Gao, J.P.; Li, L.G.; Cai, X.L.; Huang, W.; Chao, D.Y.; Zhu, M.Z.; Wang, Z.Y.; Luan, S.; Lin, H.X. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef]
- El Mahi, H.; Pérez-Hormaeche, J.; De Luca, A.; Villalta, I.; Espartero, J.; Gámez-Arjona, F.; Fernández, J.L.; Bundó, M.; Mendoza, I.; Mieulet, D.; et al. A critical role of sodium flux via the plasma membrane Na+/H+ exchanger SOS1 in the salt tolerance of rice. Plant Physiol. 2019, 180, 1046–1065. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.P.; Zheng, L.Q.; Xue, Y.H.; Zhang, Q.; Wang, L.; Shuo, H.X. Overexpression of OsVP1 and OsNHX1 increases tolerance to drought and salinity in rice. J. Plant Biol. 2010, 53, 444–452. [Google Scholar] [CrossRef]
- Chen, G.; Hu, Q.; Luo, L.; Yang, T.; Zhang, S.; Hu, Y.; Xu, G. Rice potassium transporter OsHAK1 is essential for maintaining potassium-mediated growth and functions in salt tolerance over low and high potassium concentration ranges. Plant Cell. Environ. 2009, 38, 2747–2765. [Google Scholar] [CrossRef]
- Yang, T.Y.; Zhang, S.; Hu, Y.B.; Wu, F.C.; Hu, Q.D.; Chen, G.; Cai, J.; Wu, T.; Moran, N.; Yu, L.; et al. The role of a potassium transporter OsHAK5 in potassium acquisition and transport from roots to shoots in rice at low potassium supply levels. Plant Physiol. 2014, 166, 945–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, H.M.; Tang, Q.; Cai, J.; Xu, B.C.; Xu, G.H.; Yu, L. Rice OsHAK16 functions in potassium uptake and translocation in shoot maintaining potassium homeostasis and salt tolerance. Planta 2019, 250, 549–561. [Google Scholar] [CrossRef]
- Shen, Y.; Shen, L.; Shen, Z.J.W.; Ge, H.; Zhao, J.; Zhang, W. The potassium transporter OsHAK 21 functions in the maintenance of ion homeostasis and tolerance to salt stress in rice. Plant Cell. Environ. 2015, 38, 2766–2779. [Google Scholar] [CrossRef] [PubMed]
- Nongpiur, R.; Soni, P.; Karan, R.; Singla-Pareek, S.L.; Pareek, A. Histidine kinases in plants. Plant Signal Behav. 2012, 7, 1230–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, L.S.P.; Urao, T.; Qin, F.; Maruyama, K.; Kakimoto, T.; Shonozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of AHK1/ATHK1 and cytokinin receptor histidine kinases in response to abscisic acid drought and salt stress in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 20623–20628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.N.; Jane, W.N.; Verslues, P.E. Role of the putative osmosensor Arabidopsis histidine kinase1 in dehydration avoidance and low-water-potential response. Plant Physiol. 2013, 161, 942–953. [Google Scholar] [CrossRef] [Green Version]
- Kushwaha, H.R.; Singla-Pareek, S.L.; Pareek, A. Putative osmosensor–OsHK3b–a histidine kinase protein from rice shows high structural conservation with its ortholog AtHK1 from Arabidopsis. J. Biomol. Struct. Dyn. 2014, 32, 1318–1332. [Google Scholar] [CrossRef] [Green Version]
- Ozakabe, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S.P. Sensing the environment: Key roles of membrane-localized kinases in plant perception and response to abiotic stress. J. Exp. Bot. 2013, 64, 445–458. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.L.; Zhang, L.Q.; Liu, N.; Xu, S.L.; Yue, Z.L.; Zhang, L.L.; Deng, Z.P.; Burlingame, A.L.; Sun, D.Y.; Wang, Z.Y.; et al. Mutual regulation of receptor-like kinase SIT1 and B’κ-PP2A shapes the early response of rice to salt stress. Plant Cell 2019, 31, 2131–2151. [Google Scholar] [CrossRef]
- Yuan, F.; Yang, H.; Xue, Y.; Kong, D.; Ye, R.; Li, C.; Zhang, J.; Theprungsirikul, L.; Shrift, T.; Krichilsky, B. OSCA1 mediates osmotic-stress-evoked Ca2+ increases vital for osmosensing in Arabidopsis. Nature 2014, 514, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.S.; Yuan, F.; Wen, Z.H.; Li, Y.H.; Wang, F.; Zhu, T.; Zhou, W.Q.; Jin, X.; Wang, Y.D.; Zhao, H.P.; et al. Genome-wide survey and expression analysis of the OSCA gene family in rice. BMC Plant Biol. 2015, 15, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.Z.; Ishitani, M.; Kim, C.S.; Zhu, J.K. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. USA 2000, 97, 6896–6901. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Kader, M.A.; Lindberg, S.; Seidel, T.; Golldack, D.; Yemelyanov, V. Sodium sensing induces different changes in free cytosolic calcium concentration and pH in salt-tolerant and -sensitive rice (Oryza sativa) cultivars. Physiol. Plantarum. 2007, 130, 99–111. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism oxidative stress and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Batistic, O.; Kudla, J. Integration and channeling of calcium signaling through the CBL calcium sensor/CIPK protein kinase network. Planta 2004, 219, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Bouché, N.; Yellin, A.; Snedded, W.A.; Fromm, H. Plant-specific calmodulin-binding proteins. Annu. Rev. Plant Sci. 2005, 56, 435–466. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.Q.; Xu, L.Q.; Singh, A.; Wang, H.Z.; Du, L.Q.; Poovaiah, B.W. Involvement of calmodulin and calmodulin-like proteins in plant responses to abiotic stresses. Front. Plant Sci. 2015, 6, 600. [Google Scholar] [CrossRef] [Green Version]
- Yuenyong, W.; Chinpongpanich, A.; Comai, L.; Chadchawan, S.; Buaboocha, T. Downstream components of the calmodulin signaling pathway in the rice salt stress response revealed by transcriptome profiling and target identification. BMC Plant Biol. 2018, 18, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katou, S.; Kuroda, K.; Seo, S.; Yanagawa, Y.; Tsuge, T.; Yamazaki, M.; Miyao, A.; Hirochika, H.; Ohashi, Y. A calmodulin-binding mitogen-activated protein kinase phosphatase is induced by wounding and regulates the activities of stress-related mitogen-activated protein kinases in rice. Plant Cell. Physiol. 2007, 48, 332–344. [Google Scholar] [CrossRef]
- Yang, J.; Liu, S.; Ji, L.X.; Tang, X.Y.; Zhu, Y.S.; Xie, G.S. Identification of novel OsCML16 target proteins and differential expression analysis under abiotic stresses in rice. J. Plant Physiol. 2020, 249, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.Y.; Li, S.Z.; Xie, K.; Zhang, Q.; Wang, Y.; Tang, Y.; Hong, Y.G.; He, C.Y.; Liu, Y.L. Plant ERD2-like proteins function as endoplasmic reticulum luminal protein receptors and participate in programmed cell death during innate immunity. Plant J. 2012, 72, 57–69. [Google Scholar] [CrossRef]
- Asano, T.; Tanaka, N.; Yang, G.X.; Hayashi, N.; Komatsu, S. Genome-wide Identification of the Rice Calcium-dependent Protein Kinase and its Closely Related Kinase Gene Families: Comprehensive Analysis of the CDPKs Gene Family in Rice. Plant Cell. Physiol. 2005, 46, 356–366. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.L.; Jiang, S.F.; Liang, J.S.; Chen, M. Roles of plant CBL-CIPK systems in abiotic stress responses. Turk. J. Bot. 2019, 43, 271–280. [Google Scholar] [CrossRef]
- Martínez-Atienza, J.; Jiang, X.; Garciadeblas, B.; Mendoza, I.; Zhu, J.K.; Pardo, J.M.; Quintero, F.J. Conservation of the salt overly sensitive pathway in rice. Plant Physiol. 2007, 143, 1001–1012. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.W.; Zhang, T.Y.; Wang, H.; Zhao, N.; Liu, B. Heritable alteration in salt tolerance in rice induced by introgression from wild rice (Zizania latifolia). Rice 2012, 5, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanwar, P.; Sanyal, S.K.; Tokas, I.; Yadav, A.K.; Pandey, A.; Kapoor, S.; Pandey, G.K. Comprehensive structural interaction and expression analysis of CBL and CIPK complement during abiotic stresses and development in rice. Cell Calcium 2014, 56, 81–95. [Google Scholar] [CrossRef]
- Chen, X.; Gu, Z.; Liu, F.; Ma, B.; Zhang, H. Molecular analysis of rice CIPKs Involved in both biotic and abiotic stress responses. Rice Sci. 2016, 18, 1–9. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Rohila, J.S.; Yang, Y. Rice mitogen-activated protein kinase gene family and its role in biotic and abiotic stress response. J. Integr. Plant Biol. 2007, 49, 751–759. [Google Scholar] [CrossRef]
- Rodriguez, M.C.; Petersen, M.; Mundy, J. Mitogen-activated protein kinase signaling in plants. Annu. Rev. Plant Biol. 2010, 61, 621–649. [Google Scholar] [CrossRef]
- MAPK Group. Mitogen-activated protein kinase cascades in plants: A new nomenclature. Trends Plant Sci. 2002, 7, 301–308. [Google Scholar] [CrossRef]
- Sinha, A.K.; Jaggi, M.; Raghuram, B.; Tuteja, N. Mitogen-activated protein kinase signalling in plants under abiotic stress. Plant Signal Behav. 2011, 16, 196–203. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.; Wankhede, D.P.; Sinha, A.K. Signal convergence through the lenses of MAP kinases: Paradigms of stress and hormone signaling in plants. Front. Biol. 2013, 8, 109–118. [Google Scholar] [CrossRef]
- Yoo, S.J.; Kim, S.H.; Kim, M.J.; Ryu, C.M.; Kim, Y.C.; Cho, B.H.; Yang, K.Y. Involvement of the OsMKK4-OsMPK1 cascade and its downstream transcription factor OsWRKY53 in the wounding response in rice. Plant Path J. 2014, 30, 168–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohnert, H.J.; Ayoubi, P.; Borchert, C.; Bressan, R.A.; Burnap, R.L.; Cushman, J.C.; Deyholos MFischer RGalbraith, D.W.; Hasegawa, P.M.; Jenks, M.; Kawasaki, M.; et al. A genomics approach towards salt stress tolerance. Plant Physiol. Biochem. 2001, 39, 295–311. [Google Scholar] [CrossRef]
- Zhu, J.K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Seki, M.; Narusaka, M.; Abe, H.; Kasuga, M.; Yamaguchi-Shinozaki, K.; Carninci, P.; Hayashizaki, Y.; Shinozaki, K. Monitoring the expression pattern of 1300 Arabidopsis genes under drought and cold stresses by using a full-length cDNA microarray. Plant Cell 2001, 13, 61–72. [Google Scholar] [CrossRef] [Green Version]
- Seki, M.; Ishida, J.; Narusaka, M.; Fujita, M.; Nanjo, T.; Umezawa, T.; Kamiya, A.; Nakajima, M.; Enju, A.; Sakurai, T.; et al. Monitoring the expression pattern of 7000 Arabidopsis genes under ABA treatments by using a full-length cDNA microarray. Plant J. 2020, 2, 282–291. [Google Scholar] [CrossRef]
- Gao, G.; Zhong, Y.F.; Guo, A.; Zhu, Q.; Tang, W.; Zheng, W. DRTF: A database of rice transcription factors. Bioinformatics 2006, 22, 1286–1287. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.X.; Liu, D.; Pan, Y.; Gong, W.; Ma, L.G.; Luo, J.C.; Deng, X.W.; Zhu, Y. An annotation update via cDNA sequence analysis and comprehensive profiling of developmental hormonal or environmental responsiveness of the Arabidopsis AP2/EREBP transcription factor gene family. Plant Mol. Biol. 2005, 59, 853–868. [Google Scholar] [CrossRef]
- Agarwal, P.K.; Gupta, K.; Lopato, S.; Agarwal, P. Dehydration responsive element binding transcription factors and their applications for the engineering of stress tolerance. J. Exp. Bot. 2017, 68, 2135–2148. [Google Scholar] [CrossRef]
- Ito, Y.; Katsura, K.; Maruyama, K.; Taji, T.; Kobayashi, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of rice DREB1/CBF-type transcription factors involved in cold-responsive gene expression in transgenic rice. Plant Cell. Physiol. 2006, 47, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.Q.; Meng, X.P.; Zhang, Y.; Xia, M.; Wang, X.P. Over-expression of OsDREB genes lead to enhanced drought tolerance in rice. Biotechnol. Lett. 2008, 30, 2191–2198. [Google Scholar] [CrossRef]
- Danyluk, J.; Perron, A.; Houde, M.; Limin, A.; Fowler, B.; Benhamou, N.; Sarhan, F. Accumulation of an acidic dehydrin in the vicinity of the plasma membrane during cold acclimation of wheat. Plant Cell 1998, 10, 623–638. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.S. The role of DREB transcription factors in abiotic stress tolerance of plants. Agric. Environ. Biotechnol. 2011, 25, 2433–2442. [Google Scholar] [CrossRef] [Green Version]
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; Prasad, M. NAC proteins: Regulation and role in stress tolerance. Trends Plant Sci. 2007, 17, 369–381. [Google Scholar] [CrossRef]
- Li, C.N.; Ng, C.K.Y.; Fan, L.M. MYB transcription factors active players in abiotic stress signaling. Environ. Exp. Bot. 2015, 114, 80–91. [Google Scholar] [CrossRef]
- Li, J.; Han, G.; Sun, C.; Sui, N. Research advances of MYB transcription factors in plant stress resistance and breeding. Trends Plant Sci. 2019, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lakra, N.; Nutan, K.K.; Das, P.; Anwar KSingla-Pareek, S.L.; Pareek, A. A nuclear-localized histone-gene binding protein from rice (OsHBP1b) functions in salinity and drought stress tolerance by maintaining chlorophyll content and improving the antioxidant machinery. J. Plant Physiol. 2015, 176, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Li, W.T.; He, M.; Wang, J.; Wang, Y.P. Zinc finger protein (ZFP) in plants-A review. Plant Omics J. 2013, 6, 474–480. [Google Scholar]
- Murre, C.; Bain, G.; Dijk, M.A.V.; Engel, I.; Furnari, B.A.; Massari, M.E.; Matthews, J.R.; Quong, M.W.; Rivera, R.R.; Stuiver, M.H. Structure and function of helix–loop–helix proteins. BBA Biomembr. 1994, 1218, 129–135. [Google Scholar] [CrossRef]
- Wu, H.; Ye, H.Y.; Yao, R.F.; Zhang, T.; Xiong, L.Z. OsJAZ9 acts as a transcriptional regulator in jasmonate signaling and modulates salt stress tolerance in rice. Plant Sci. 2015, 232, 1–12. [Google Scholar] [CrossRef]
- Sun, X.; Wang, Y.; Sui, N. Transcriptional regulation of bHLH during plant response to stress. Biochem. Biophys. Res. Commun. 2018, 503, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.M.; Gregorio, G.; Oliveira, M.M.; Saibo, N.J.M. Five novel transcription factors as potential regulators of OsNHX1 gene expression in a salt tolerant rice genotype. Plant Mol. Biol. 2017, 93, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Rontein, D.; Basset, G.; Hanson, A.D. Metabolic engineering of osmoprotectant accumulation in plants. Metab. Eng. 2002, 4, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdallah, M.M.S.; Abdelgawad, Z.A.; El-Bassiuny, H.M.S. Alleviation of the adverse effects of salinity stress using trehalose in two rice varieties. South Afr. J. Bot. 2016, 103, 275–282. [Google Scholar] [CrossRef]
- Bhusan, D.; Das, D.K.; Hossain, M.; Murata, Y.; Hoque, M.A. Improvement of salt tolerance in rice (Oryza sativa L.) by increasing antioxidant defense systems using exogenous application of proline. Aus. J. Crop. Sci. 2016, 10, 50–56. [Google Scholar]
- Demiral, T.; Türkan, I. Exogenous glycine betaine affects growth and proline accumulation and retards senescence in two rice cultivars under NaCl stress. Environ. Exp. Bot. 2005, 56, 72–79. [Google Scholar] [CrossRef]
- Harinasut, P.; Tsutsui, K.; Takabe, T.; Nomura, M.; Takabe, T.; Kishitani, S. Exogenous glycinebetaine accumulation and increased salt-tolerance in rice seedlings. Biosci. Biotech. Bioch. 1996, 60, 366–368. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Ding, T.; Zuo, J.; Gao, L.; Fan, L. Amelioration of postharvest chilling injury in sweet pepper by glycine betaine. Postharvest Biol. Technol. 2016, 112, 114–120. [Google Scholar] [CrossRef]
- Chen, T.H.H.; Murata, N. Glycine betaine protects plants against abiotic stress: Mechanisms and biotechnological applications. Plant Cell. Environ. 2010, 34, 1–20. [Google Scholar] [CrossRef]
- Chaumont, F.; Tyerman, S.D. Aquaporins: Highly regulated channels controlling plant water relations. Plant Physiol. 2014, 164, 1600–1618. [Google Scholar] [CrossRef] [Green Version]
- Maurel, C.; Boursiac, Y.; Luu, D.T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, J.; Ishikawa, F.; Yamaguchi, T.; Uemura, M.; Maeshima, M. Identification of 33 rice aquaporin genes and analysis of their expression and function. Plant Cell. Physiol. 2005, 46, 1568–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawasaki, S.; Borchert, C.; Deyholos, M.; Wang, H.; Brazille, S.; Kawai, K.; Galbraith, D.; Bohnert, H.J. Gene expression profiles during the initial phase of salt stress in rice. Plant Cell 2001, 13, 889–905. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Miao, Y.; Song, C.P. Behind the scenes: The roles of reactive oxygen species in guard cells. New Phytol. 2014, 201, 1121–1140. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.G.; Shan, J.X.; Shi, M.; Gao, J.P.; Lin, H.X. DCA1 acts as a transcriptional co-activator of DST and contributes to drought and salt tolerance in rice. PLoS Genet. 2015, 11, e1005617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef]
- Mishra, P.; Bhoomika, K.; Dubey, R.S. Differential responses of antioxidative defense system to prolonged salinity stress in salt-tolerant and salt-sensitive Indica rice (Oryza sativa L.) seedlings. Protoplasma 2013, 250, 3–19. [Google Scholar] [CrossRef]
- Rossatto, T.; do Amaral, M.N.; Benitz, L.C.; Vighi, I.L.; Braga, E.J.B.; Júnior, A.M.D.; Maia, M.A.C.; Pinto, L.D. Gene expression and activity of antioxidant enzymes in rice plants, cv. BRS AG, under saline stress. Physiol. Mol. Biol. Plants 2017, 23, 865–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Ahsan, N.; Lee, K.W.; Kim, D.H.; Lee, D.G.; Kwak, S.S. Simultaneous overexpression of both Cu/Zn superoxide dismutase and ascorbate peroxidase in transgenic tall fescue plants confers increased tolerance to a wide range of abiotic stresses. J. Plant Physiol. 2007, 164, 1626–1638. [Google Scholar] [CrossRef]
- Wang, Y.; Ying, Y.; Chen, J.; Wang, X. Transgenic Arabidopsis, overexpressing Mn-SOD enhanced salt-tolerance. Plant Sci. 2004, 167, 671–677. [Google Scholar] [CrossRef]
- Cheng, Y.W.; Kong, X.W.; Wang, N.; Wang, T.T.; Chen, J.; Shi, Z.Q. Thymol confers tolerance to salt stress by activating anti-oxidative defense and modulating Na+ homeostasis in rice root. Ecotoxicol. Environ. Saf. 2020, 188, 109894. [Google Scholar] [CrossRef] [PubMed]
- Mhamdi, A.; Queval, G.; Chaouch, S.; Vanderauwera, S.; Breusegem, F.V.; Noctor, G. Catalase function in plants: A focus on Arabidopsis mutants as stress-mimic models. J. Exp. Bot. 2010, 61, 4197–4220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Poovaiah, B.W. Hydrogen peroxide homeostasis: Activation of plant catalases by calcium/calmodulin. Proc. Natl. Acad. Sci. USA 2002, 99, 4097–4102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, J.; Lee, Y.H.; Song, S.I. Rice CatA, CatB, and CatC are involved in environmental stress response, root growth, and photorespiration, respectively. J. Plant Biol. 2002, 57, 375–382. [Google Scholar] [CrossRef]
- Zhou, Y.B.; Liu, C.; Tang, D.Y.; Yan, L.; Wang, D.; Yang, Y.Z.; Gui, J.S.; Zhao, X.Y.; Li, L.G.; Tang, X.D.; et al. The receptor-like cytoplasmic kinase STRK1 phosphorylates and activates CatC, thereby regulating H2O2 homeostasis and improving salt tolerance in rice. Plant Cell 2018, 30, 1100–1118. [Google Scholar] [CrossRef] [Green Version]
- Kibria, M.G.; Hossain, M.; Murata, Y.; Hoque, M.A. Antioxidant defense mechanisms of salinity tolerance in rice genotypes. Rice Sci. 2017, 24, 155–162. [Google Scholar] [CrossRef]
- Yamane, K.; Mitsuya, S.; Taniguchi, M.; Miyake, H. Transcription profiles of genes encoding catalase and ascorbate peroxidase in the rice leaf tissues under salinity. Plant Prod. Sci. 2010, 13, 164–168. [Google Scholar] [CrossRef]
- Wutipraditkul, N.; Boonkomrat, S.; Buaboocha, T. Cloning and characterization of catalases from rice, Oryza sativa L. Biosci Biotechnol Biochem 2011, 75, 1900–1906. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, F.K.; Menezes-Benzvente, L.; Margis, R.; Margis-Pinheiro, M. Analysis of the molecular evolutionary history of the ascorbate peroxidase gene family: Inferences from the rice genome. J. Mol. Evol. 2004, 59, 761–770. [Google Scholar] [CrossRef]
- Hong, C.Y.; Hsu, Y.T.; Tsai, Y.C.; Kao, C.H. Expression of Ascorbate Peroxidase 8 in roots of rice (Oryza sativa L) seedlings in response to NaCl. J. Exp. Bot. 2007, 58, 3273–3283. [Google Scholar] [CrossRef]
- Bonifacio, A.; Martins, M.O.; Ribeiro, C.V.; Fontenele, A.V.; Carvalho, F.E.; Margis-Pinheiro, M.; Silveira, J.G. Role of peroxidases in the compensation of cytosolic ascorbate peroxidase knockdown in rice plants under abiotic stress. Plant Cell. Environ. 2011, 34, 1705–1722. [Google Scholar] [CrossRef]
- Rosa, S.B.; Caverzan, A.; Teixeira, F.K.; Lazzarotto, F.; Silveira, J.A.G.; Ferreira-Silva, S.L.; Abreu-Neto, J.; Margis, R.; Margis-Pinheiro, M. Cytosolic APx knockdown indicates an ambiguous redox response in rice. Phytochem 2010, 71, 548–558. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.M.; Lin, W.R.; Kao, C.H.; Hong, C.Y. Gene knockout of glutathione reductase 3 results in increased sensitivity to salt stress in rice. Plant Mol. Biol. 2015, 87, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.Y.; Chao, Y.Y.; Yang, M.Y.; Cheng, S.Y.; Cho, S.C.; Kao, C.H. NaCl-induced expression of glutathione reductase in roots of rice (Oryza sativa L.) seedlings is mediated through hydrogen peroxide but not abscisic acid. Plant Soil 2009, 320, 103–115. [Google Scholar] [CrossRef]
- Tsai, Y.C.; Hong, Y.C.; Kiu, L.F.; Kao, C.H. Expression of ascorbate peroxidase and glutathione reductase in roots of rice seedlings in response to NaCl and H2O2. J. Plant Physiol. 2005, 162, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A. Thioredoxin and glutaredoxin systems. J. Biol. Chem. 1989, 264, 13963–13966. [Google Scholar] [CrossRef]
- Garg, R.; Jhanwar, S.; Tyagi, A.K.; Jain, M. Genome-wide survey and expression analysis suggest diverse roles of glutaredoxin gene family members during development and response to various stimuli in rice. DNA Res. 2010, 17, 353–367. [Google Scholar] [CrossRef] [Green Version]
- Nuruzzaman, M.; Gupta, M.; Zhang, C.; Wang, L.; Xie, W.; Xiong, L.; Zhang, Q.; Lian, X. Sequence and expression analysis of the thioredoxin protein gene family in rice. Mol. Genet. Genom. 2008, 280, 139–151. [Google Scholar] [CrossRef]
- Zhang, C.J.; Zhao, B.C.; Ge, W.N.; Zhang, F.Y.; Song, Y.; Sun, D.Y.; Guo, Y. An apoplastic H-type thioredoxin is involved in the stress response through regulation of the apoplastic reactive oxygen species in rice. Plant Physiol. 2011, 157, 1884–1899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, G.S.; Kato, H.; Sasaki, K.; Imai, R. A cold-induced thioredoxin h of rice OsTrx23 negatively regulates kinase activities of OsMPK3 and OsMPK6 in vitro. Febs Lett. 2009, 583, 2734–2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blumwald, E.; Aharon, G.S.; Apse, M.P. Sodium transport in plant cells. Biochem. Biophys. Acta 2000, 1465, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.J.; Ding, L.; Zhu, J.K. SOS1, a genetic locus essential for salt tolerance and potassium acquisition. Plant Cell. 1996, 8, 617–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garciadeblas, B.; Senn, M.E.; Banuelos, M.A.; Rodriguez-Navarro, A. Sodium transport and HKT transporters: The rice model. Plant J. 2003, 34, 788–801. [Google Scholar] [CrossRef]
- Horie, T.; Yoshida, K.; Nakayama, H.; Yamada, K.; Oiki, S.; Shinmyo, A. Two types of HKT transporters with different properties of Na+ and K+ transport in Oryza sativa. Plant J. 2001, 27, 129–138. [Google Scholar] [CrossRef]
- Suzuki, K.; Yamaji, N.; Costa, A.; Okuma, E.; Kobayashi, N.I.; Kashiwagi, T.; Katsuhara, M.; Wang, C.; Tanoi, K.; Murata, Y.; et al. OsHKT1;4-mediated Na+ transport in stems contributes to Na+ exclusion from leaf blades of rice at the reproductive growth stage upon salt stress. BMC Plant Biol. 2016, 16, 22. [Google Scholar] [CrossRef] [Green Version]
- Imran, S.; Horie, T.; Katsuhara, M. Expression and ion transport activity of rice OsHKT1;1 variants. Plants 2020, 9, 16. [Google Scholar] [CrossRef] [Green Version]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Glenn, E.P.; Brown, J.J.; Blumwald, E. Salt tolerance and crop potential of halophytes. Crit. Rev. Plant Sci. 1999, 18, 227–255. [Google Scholar] [CrossRef]
- Kader, M.A.; Lindberg, S. Cytosolic calcium and pH signaling in plants under salinity stress. Plant Signal Behav. 2010, 5, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, A.; Nakamura, A.; Hara, N.; Toki, S.; Tanaka, Y. Molecular and functional analyses of rice NHX-type Na+/H+ antiporter genes. Planta 2011, 233, 175–188. [Google Scholar] [CrossRef]
- Amin, U.S.M.; Biswas, S.; Elias, S.M.; Razzaque, S.; Haque, T.; Malo, R.; Seraj, Z.I. Enhanced salt tolerance conferred by the complete 23 kb cDNA of the rice vacuolar Na+/H+ antiporter gene compared to 19 kb coding region with 5’ UTR in transgenic lines of rice. Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brini, F.; Hanin, M.; Mezghani, I.; Berkowitz, G.A.; Masmou, K. Overexpression of wheat Na+/H+ antiporter TNHX1 and H+-pyrophosphatase TVP1 improve salt- and drought-stress tolerance in Arabidopsis thaliana plants. J. Exp. Bot. 2007, 58, 301–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaxiola, R.A.; Li, J.S.; Undurraga, S.; Dang, L.M.; Allen, G.J.; Alper, S.L.; Fink, G.R. Drought- and salt-tolerant plants result from overexpression of the AVP1 H+-pump. Proc. Natl. Acad. Sci. USA 2005, 98, 11444–11449. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.Y.; Zhang, X.J.; Li, P.H.; Zhao, Y.X.; Zhang, H. Co-expression of the Suaeda salsa SsNHX1 and Arabidopsis AVP1 confer greater salt tolerance to transgenic rice than the single SsNHX1. Mol. Breed. 2006, 17, 341–353. [Google Scholar] [CrossRef]
- Gong, H.J.; Randall, D.P.; Flowers, T.J. Silicon deposition in the root reduces sodium uptake in rice (Oryza sativa L.) seedlings by reducing bypass flow. Plant Cell. Environ. 2006, 1273, 433. [Google Scholar] [CrossRef]
- Krishnamurthy, P.; Ranathunge, K.; Franke, R.; Prakash, H.S.; Schreiber, L.; Mathew, M.K. The role of root apoplastic transport barriers in salt tolerance of rice (Oryza sativa L.). Planta 2009, 230, 119–134. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Functional Annotation | Method of Validation | * Regulation Role | References |
|---|---|---|---|---|---|
| Osmosensing | |||||
| SIT1 | LOC_Os02g42780 | lectin receptor-type protein kinase | Knockdown Overexpression | – | [9] |
| Signaling | |||||
| OsCam1-1 | LOC_Os03g20370 | Calmodulin | Overexpression | + | [10] |
| OsCPK4 | LOC_Os02g03410 | CAMK_CAMK_like.12—Ca2+/calmodulin-dependent protein kinase(CAMK) includes calcium/calmodulin dependent protein kinases | Knockdown Overexpression | + | [11] |
| OsCDPK7 | LOC_Os04g49510 | CAMK_CAMK_like.27—CAMK includes calcium/calmodulin dependent protein kinases | Overexpression | + | [12,13] |
| OsCPK12 | LOC_Os04g47300 | CAMK_CAMK_like.26—CAMK includes calcium/calmodulin dependent protein kinases | Overexpression | + | [14] |
| OsCPK21 | LOC_Os08g42750 | CAMK_CAMK_like.37—CAMK includes calcium/calmodulin dependent protein kinases | Overexpression | + | [15,16] |
| OsCIPK15 | LOC_Os11g02240 | CAMK_Nim1_like.4—CAMK includes calcium/calmodulin dependent protein kinases | Overexpression | + | [17] |
| OsCIPK31 | LOC_Os03g20380 | CAMK_Nim1_like.2—CAMK includes calcium/calmodulin dependent protein kinases | Mutant | + | [18] |
| OsMAPK5 | LOC_Os03g17700 | CGMC_MAPKCGMC_2_ERK.2—CGMC includes CDA, MAPK, GSK3, and CLKC kinases | Knockdown Overexpression | + | [19] |
| OsMAPK33 | LOC_Os02g05480 | CGMC_MAPKCMGC_2_SLT2y_ERK.1—includes cytidine deaminase (CDA), glycogen synthase kinase 3 (GSK3), mitogen-activated protein kinase (MAPK), and CLKC kinases | Knockdown Overexpression | – | [20] |
| OsMKK1 | LOC_Os06g05520 | MAPK | Knockdown | + | [21] |
| OsMKK6 | LOC_Os01g32660 | STE_MEK_ste7_MAP2K.2—STE kinases | Overexpression | + | [22] |
| OsMaPKKK63 | LOC_Os01g50370 | STE_MEKK_ste11_MAP3K.4—STE kinases | Knockdown | – | [23] |
| Transcriptional regulation | |||||
| OsDREB1A | LOC_Os09g35030 | Dehydration-responsive element (DRE)--binding protein | Overexpression | + | [24] |
| OsDREB1D | LOC_Os06g06970 | DRE--binding protein | Overexpression | + | [25] |
| OsDREB1F | LOC_Os01g73770 | DRE--binding protein | Overexpression | + | [26] |
| OsDREB2A | LOC_Os01g07120 | APETALA2 (AP2) domain containing protein | Overexpression | + | [27,28] |
| OsDREB2B | LOC_Os05g27930 | AP2 domain containing protein | Overexpression | + | [29] |
| OsAP23 | LOC_Os03g05590 | AP2 domain containing protein | Overexpression | – | [30] |
| OsAP37 | LOC_Os01g58420 | AP2 domain containing protein | Overexpression | + | [31] |
| OsSTAP1 | LOC_Os03g08470 | APETALA2/ethylene responsive factor (AP2/ERF)-type transcription factor | Overexpression | + | [32] |
| OsDREB6 | LOC_Os09g20350 | ERF transcription factor | Knockdown Overexpression | + | [33] |
| SERF1 | LOC_Os05g34730 | ERF020- transcription factor | Knockdown | + | [34] |
| OsERF922 | LOC_Os01g54890 | Ethylene-responsive transcription factor 2 | Knockdown Overexpression | – | [35] |
| OsRAV2 | LOC_Os01g04800 | B3 DNA binding domain containing protein | Mutant | + | [36] |
| OsNAP | LOC_Os03g21060 | No apical meristem (NAM)protein | Overexpression | + | [37] |
| ONAC022 | LOC_Os03g04070 | NAM protein | Overexpression | + | [38] |
| ONAC045 | LOC_Os11g03370 | NAM protein | Overexpression | + | [39] |
| ONAC063 | LOC_Os08g33910 | NAM protein | Overexpression | + | [40] |
| ONAC106 | LOC_Os08g33670 | NAM protein | Overexpression | + | [41] |
| OsNAC2 | LOC_Os04g38720 | NAM protein | Overexpression | + | [42,43] |
| OsNAC5 | LOC_Os11g08210 | NAM protein | Knockdown Overexpression | + | [44,45] |
| OsNAC6/SNAC2 | LOC_Os01g66120 | NAM protein | Overexpression | + | [46,47] |
| SNAC1 | LOC_Os03g60080 | NAM, ATAF and CUC (NAC) domain-containing protein 67 | Overexpression | + | [48] |
| OsNAC10 | LOC_Os11g03300 | NAC domain transcription factor | Overexpression | + | [49] |
| OsNAC041 | - | - | Knockdown | + | [50] |
| OsMYB2 | LOC_Os03g20090 | Myeloblastosis (MYB) family transcription factor | Overexpression | + | [51] |
| OsMYB3R-2 | LOC_Os01g62410 | MYB family transcription factor | Overexpression | + | [52] |
| OsMYB48-1 | LOC_Os01g74410 | MYB family transcription factor | Overexpression | + | [53] |
| OsMPS | LOC_Os02g40530 | MYB family transcription factor | Overexpression | + | [54] |
| OsMYB91 | LOC_Os12g38400 | MYB family transcription factor | Knockdown Overexpression | + | [55] |
| OsMYBc | LOC_Os09g12770 | Adenosine-thymine (AT) hook motif domain containing protein | Mutant | + | [56] |
| OsABF2 | LOC_Os06g10880 | Basic leucine-zipper (bZIP) transcription factor | Mutant | + | [57] |
| OsABI5 | LOC_Os01g64000 | bZIP transcription factor | Overexpression | – | [58] |
| OsbZIP23 | LOC_Os02g52780 | bZIP transcription factor | Overexpression | + | [59] |
| OsbZIP71 | LOC_Os09g13570 | CPuORF2—conserved peptide uORF-containing transcript | Knockdown Overexpression | + | [60] |
| OsHBP1b | LOC_Os01g17260 | Transcription factor | Overexpression | + | [61] |
| DST | LOC_Os03g57240 | ZOS3-19—C2H2 zinc finger (ZF) protein | Mutant | – | [62] |
| OsTZF1 | LOC_Os05g10670 | ZF CCCH type family protein | Knockdown Overexpression | + | [63] |
| ZFP179 | LOC_Os01g62190 | ZOS1-15—C2H2 ZF protein | Overexpression | + | [64] |
| ZFP182 | LOC_Os03g60560 | ZOS3-21—C2H2 ZF protein | Overexpression | + | [65] |
| ZFP185 | LOC_Os02g10200 | ZF A20 and AN1 domain-containing stress-associated protein | Knockdown Overexpression | – | [66] |
| ZFP252 | LOC_Os12g39400 | ZOS12-09—C2H2 ZF protein | Knockdown Overexpression | + | [67] |
| OsLOL5 | LOC_Os01g42710 | LSD1-like-type ZF protein | Overexpression | + | [68] |
| OrbHLH001 | LOC_Os01g70310 | Inducer of CBF expression 2 | Overexpression | + | [69] |
| OsbHLH035 | LOC_Os01g06640 | Basic helix-loop-helix (bHLH) | Mutant | + | [70] |
| Oshox22 | LOC_Os04g45810 | Homeobox associated leucine zipper | Mutant Overexpression | – | [71] |
| OsTF1L | LOC_Os08g19590 | Homeobox domain containing protein | Knockdown Overexpression | + | [72] |
| OsMADS25 | LOC_Os04g23910 | MADS-box family gene with MIKCc type-box | Knockdown Overexpression | + | [73] |
| OsWRKY45 | LOC_Os05g25770 | WRKY45 | Knockdown Overexpression | – | [74] |
| Osmoprotection | |||||
| OsBADH1 | LOC_Os04g39020 | Aldehyde dehydrogenase | Knockdown; Overexpression | + | [75,76] |
| OsTPP1 | LOC_Os02g44230 | CPuORF22—conserved peptide uORF-containing transcript | Overexpression | + | [77] |
| OsTPS1 | LOC_Os05g44210 | Trehalose-6-phosphate synthase | Overexpression | + | [78] |
| OsTPS8 | LOC_Os08g34580 | Trehalose-6-phosphate synthase | Mutant | + | [79] |
| Overexpression | |||||
| Osmoregulation | |||||
| OsPIP1;1 | LOC_Os02g44630 | Aquaporin protein | Overexpression | + | [80,81] |
| OsPIP2;2 | LOC_Os02g41860 | Aquaporin protein | Overexpression | + | [80] |
| Stomatal Closure | |||||
| LP2 | LOC_Os02g40240 | Receptor kinase | Overexpression | + | [82] |
| OsSRO1c | LOC_Os03g12820 | ATP8 | Mutant Overexpression | + | [83] |
| Antioxidation | |||||
| OsCu/Zn-SOD | LOC_Os08g44770 | Copper/zinc superoxide dismutase | Overexpression | + | [84] |
| OsMn-SOD | LOC_Os05g25850 | Manganese superoxide dismutase | Overexpression | + | [85] |
| OsAPx1 | LOC_Os03g17690 | Cytosolic Ascorbate Peroxidase encoding gene 1-8 | Overexpression | + | [86] |
| OsAPx2 | LOC_Os07g49400 | Cytosolic Ascorbate Peroxidase encoding gene 4,5,6,8 | Knockdown | + | [87] |
| Overexpression | |||||
| OsGR3 | LOC_Os10g28000 | Glutathione reductase | Knockdown | + | [88] |
| OsTRXh1/OsTrx23 | LOC_Os07g08840 | Thioredoxin | Knockdown; Overexpression | – | [89] |
| OsGRX8 | LOC_Os02g30850 | OsGrx_C8—Glutaredoxin subgroup III | Knockdown; Overexpression | + | [90] |
| OsGRX20 | LOC_Os08g44400 | Glutathione S-transferase | Knockdown; Overexpression | + | [91] |
| Na+ exclusion | |||||
| OsHKT1;1 | LOC_Os04g51820 | Na+ transporter | Natural variation | + | [92] |
| OsHKT1;4 | LOC_Os04g51830 | Na+ transporter | Mutant | – | [93] |
| OsHKT1;5/SKC1 | LOC_Os01g20160 | Na+ transporter | Natural variation | + | [94] |
| OsSOS1 | LOC_Os12g44360 | Sodium/hydrogen exchanger 7 | Mutant | + | [95] |
| Na+ compartmentation | |||||
| OsNHX1 | LOC_Os07g47100 | transporter, monovalent cation:proton antiporter-2 family | Overexpression | + | [96] |
| OsVP1 | LOC_Os01g68370 | B3 DNA binding domain containing protein | Overexpression | + | [96] |
| K+ uptake | |||||
| OsHAK1 | LOC_Os04g32920 | Potassium transporter | Mutant and overexpression | + | [97] |
| OsHAK5 | LOC_Os01g70490 | Potassium transporter | Knockdown overexpression | + | [98] |
| OsHAK16 | LOC_Os03g37840 | Potassium transporter | Overexpression | + | [99] |
| OsHAK21 | LOC_Os03g37930 | Potassium transporter | Knockdown | + | [100] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ponce, K.S.; Guo, L.; Leng, Y.; Meng, L.; Ye, G. Advances in Sensing, Response and Regulation Mechanism of Salt Tolerance in Rice. Int. J. Mol. Sci. 2021, 22, 2254. https://doi.org/10.3390/ijms22052254
Ponce KS, Guo L, Leng Y, Meng L, Ye G. Advances in Sensing, Response and Regulation Mechanism of Salt Tolerance in Rice. International Journal of Molecular Sciences. 2021; 22(5):2254. https://doi.org/10.3390/ijms22052254
Chicago/Turabian StylePonce, Kimberly S., Longbiao Guo, Yujia Leng, Lijun Meng, and Guoyou Ye. 2021. "Advances in Sensing, Response and Regulation Mechanism of Salt Tolerance in Rice" International Journal of Molecular Sciences 22, no. 5: 2254. https://doi.org/10.3390/ijms22052254
APA StylePonce, K. S., Guo, L., Leng, Y., Meng, L., & Ye, G. (2021). Advances in Sensing, Response and Regulation Mechanism of Salt Tolerance in Rice. International Journal of Molecular Sciences, 22(5), 2254. https://doi.org/10.3390/ijms22052254