
Overexpression of the Zygophyllum xanthoxylum Aquaporin, ZxPIP1;3, Promotes Plant Growth and Stress Tolerance

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
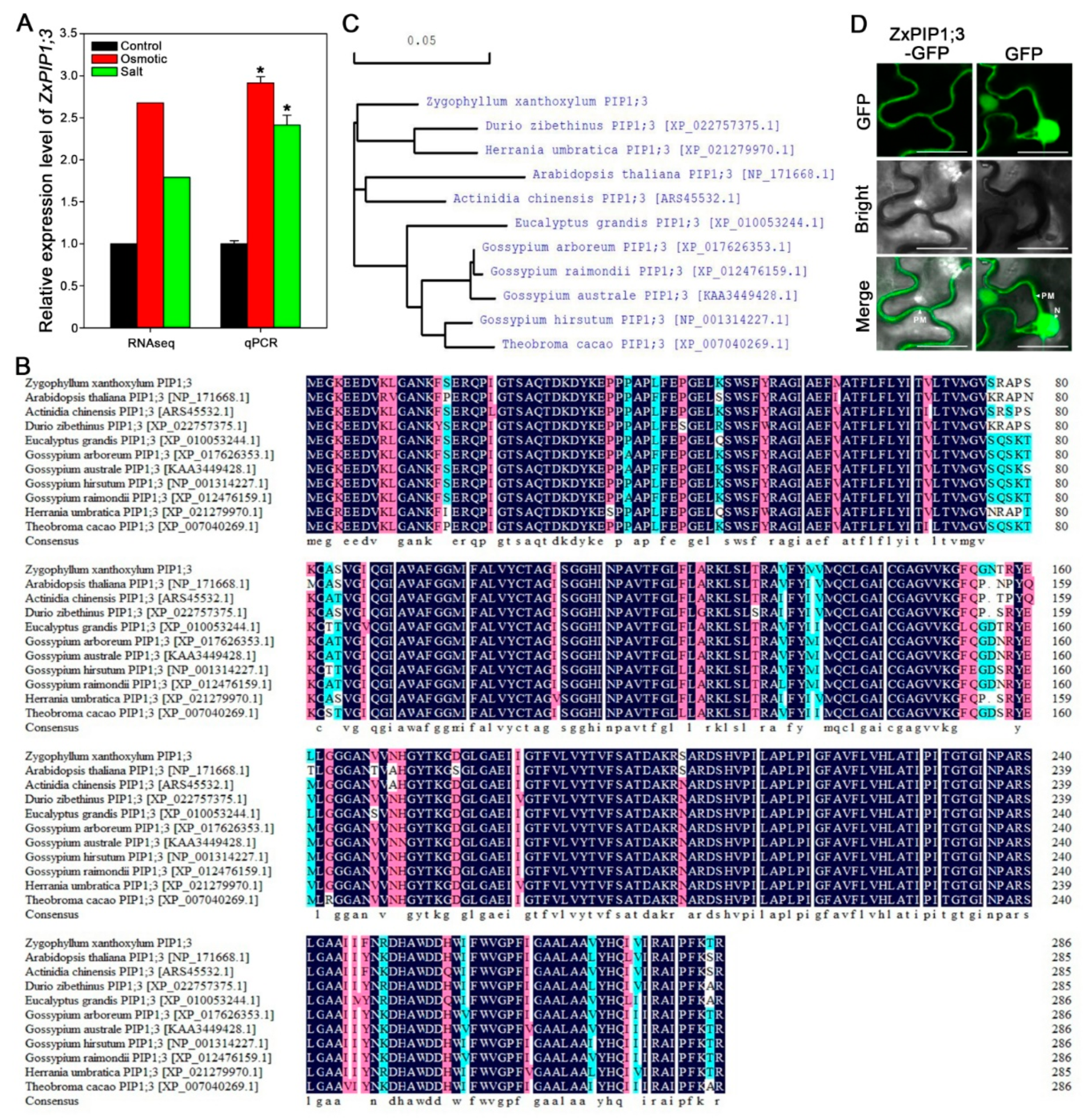
2.1. ZxPIP1;3 Is Induced under Osmotic and Salt Treatment
2.2. ZxPIP1;3 Encodes an AQP of PIP1 Subgroup
2.3. Overexpression of ZxPIP1;3 Promotes Plant Growth
2.4. ZxPIP1;3 Overexpression Improves Salt Tolerance of Transgenic Arabidopsis
2.5. ZxPIP1;3 Overexpression Confers Drought Tolerance of Transgenic Plants
2.6. Expression Level of Stress-Related Genes Is Increased in ZxPIP1;3 Transgenic Plants under Stress Treatment Compared with Wild-Type Plants
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Expression Pattern Analysis
4.3. Cloning of ZxPIP1;3 and Sequence Analysis
4.4. ZxPIP1;3 Expression Vector Construction
4.5. Subcellular Localization
4.6. Transgenic Plants Generation
4.7. Root and Shoot Growth Analysis
4.8. Salt Tolerance Analysis of Transgenic Plants
4.9. Drought Tolerance Analysis of Transgenic Plants
4.10. Expression Pattern Analysis of Stress-Related Genes
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. Regulation of root water uptake under abiotic stress conditions. J. Exp. Bot. 2011, 63, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front. Plant Sci. 2014, 5, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.H.; Saier, M.H.J. Phylogenetic characterization of the MIP family of transmembrane channel proteins. J. Membr. Biol. 1996, 153, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Kozono, D.; Ding, X.; Iwasaki, I.; Meng, X.; Kamagata, Y.; Agre, P.; Kitagawa, Y. Functional Expression and Characterization of an Archaeal Aquaporin. J. Biol. Chem. 2003, 278, 10649–10656. [Google Scholar] [CrossRef] [Green Version]
- Maurel, C.; Boursiac, Y.; Luu, D.-T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in Plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef]
- Kaldenhoff, R.; Fischer, M. Functional aquaporin diversity in plants. Biochim. Biophys. Acta (BBA) Biomembr. 2006, 1758, 1134–1141. [Google Scholar] [CrossRef] [Green Version]
- Benga, G. Water channel proteins (later called aquaporins) and relatives: Past, present, and future. IUBMB Life 2009, 61, 112–133. [Google Scholar] [CrossRef]
- Li, G.; Santoni, V.; Maurel, C. Plant aquaporins: Roles in plant physiology. Biochim. Biophys. Acta (BBA) Gen. Subj. 2014, 1840, 1574–1582. [Google Scholar] [CrossRef] [PubMed]
- Matsui, H.; Hopkinson, B.M.; Nakajima, K.; Matsuda, Y. Plasma-membrane-type aquaporins from marine diatoms function as CO2/NH3 channels and provide photoprotection. Plant Physiol. 2018, 178, 345–357. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Wang, Z.Y.; Lin, H.; Cui, W.E.; Chen, J.; Liu, M.; Chen, Z.L.; Qu, L.J.; Gu, H. Expression and functional analysis of the rice plasma-membrane intrinsic protein gene family. Cell Res. 2006, 16, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, Q.-T.; Lei, Q.; Feng, C.; Zheng, X.; Zhou, F.; Li, L.; Liu, X.; Wang, Z.; Kong, J. Ectopically expressing MdPIP1;3, an aquaporin gene, increased fruit size and enhanced drought tolerance of transgenic tomatoes. BMC Plant Biol. 2017, 17, 246. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Yuan, Q.; Wang, Y.; Cai, R.; Deng, X.; Wang, J.; Zhou, S.; Chen, M.; Chen, L.; Huang, C.; et al. Overexpression of a Wheat Aquaporin Gene, TaAQP8, Enhances Salt Stress Tolerance in Transgenic Tobacco. Plant Cell Physiol. 2012, 53, 2127–2141. [Google Scholar] [CrossRef] [Green Version]
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Ballesta, M.D.C.; Carvajal, M. New challenges in plant aquaporin biotechnology. Plant Sci. 2014, 217–218, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.-J.; Ma, Q.; Wu, G.-Q.; Wang, P.; Hu, J.; Wang, S.-M. ZxNHX controls Na+ and K+ homeostasis at the whole-plant level in Zygophyllum xanthoxylum through feedback regulation of the expression of genes involved in their transport. Ann. Bot. 2014, 115, 495–507. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Zhou, Z.; Wu, C. The Research of the breeding characters of Zygophyllum xanthoxylum. Pratacult. Sci. 2006, 23, 38–41. [Google Scholar]
- Ma, Q.; Yue, L.-J.; Zhang, J.-L.; Wu, G.-Q.; Bao, A.-K.; Wang, S.-M. Sodium chloride improves photosynthesis and water status in the succulent xerophyte Zygophyllum xanthoxylum. Tree Physiol. 2011, 32, 4–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, L.; Li, S.; Ma, Q.; Zhou, X.; Wu, G.; Bao, A.; Zhang, J.; Wang, S. NaCl stimulates growth and alleviates water stress in the xerophyte Zygophyllum xanthoxylum. J. Arid. Environ. 2012, 87, 153–160. [Google Scholar] [CrossRef]
- Ma, Q.; Hu, J.; Zhou, X.-R.; Yuan, H.-J.; Kumar, T.; Luan, S.; Wang, S.-M. ZxAKT1 is essential for K+ uptake and K+/Na+ ho-meostasis in the succulent xerophyte Zygophyllum xanthoxylum. Plant J. 2017, 90, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Li, M.; Li, D.; Khan, S.-A.; Hepworth, S.R.; Wang, S.-M. Transcriptome analysis reveals regulatory framework for salt and osmotic tolerance in a succulent xerophyte. BMC Plant Biol. 2019, 19, 88. [Google Scholar] [CrossRef]
- Ma, Q.; Bao, A.-K.; Chai, W.-W.; Wang, W.-Y.; Zhang, J.-L.; Li, Y.-X.; Wang, S.-M. Transcriptomic analysis of the succulent xerophyte Zygophyllum xanthoxylum in response to salt treatment and osmotic stress. Plant Soil 2016, 402, 343–361. [Google Scholar] [CrossRef]
- Golldack, D.; Lüking, I.; Yang, O. Plant tolerance to drought and salinity: Stress regulating transcription factors and their functional significance in the cellular transcriptional network. Plant Cell Rep. 2011, 30, 1383–1391. [Google Scholar] [CrossRef]
- Moore, K.; Roberts, L.J. Measurement of Lipid Peroxidation. Free. Radic. Res. 1998, 28, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozak, K. A nove1 cis-acting element in an Arabidopsis gene 1s involved in responsiveness to drought, low temperature, or high-salt stress. Plant Cell 1994, 6, 251–264. [Google Scholar]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought-and low temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Wan, C.; Wang, Y.; Chen, H.; Zhou, Z.; Fu, H.; Sosebee, R.E. The characteristics of Na+, K+ and free proline distribution in several drought-resistant plants of the Alxa Desert, China. J. Arid. Environ. 2004, 56, 525–539. [Google Scholar] [CrossRef]
- Flowers, T.J. Improving crop salt tolerance. J. Exp. Bot. 2004, 55, 307–319. [Google Scholar] [CrossRef]
- Yang, S.; Vanderbeld, B.; Wan, J.; Huang, Y. Narrowing Down the Targets: Towards Successful Genetic Engineering of Drought-Tolerant Crops. Mol. Plant 2010, 3, 469–490. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.H.; Lee, J.T.; Charng, Y.Y.; Chan, M.T. Tomato plants ectopically expressing Arabidopsis CBF1 show enhanced re-sistance to water deficit stress. Plant Physiol. 2002, 130, 618–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, T.-H.; Lee, J.-T.; Yang, P.-T.; Chiu, L.-H.; Charng, Y.-Y.; Wang, Y.-C.; Chan, M.-T. Heterology Expression of the Arabidopsis C-Repeat/Dehydration Response Element Binding Factor 1 Gene Confers Elevated Tolerance to Chilling and Oxidative Stresses in Transgenic Tomato. Plant Physiol. 2002, 129, 1086–1094. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.-G.; Kim, J.; Kim, B.; Jeong, H.; Choi, S.H.; Kim, E.K.; Lee, H.-Y.; Lim, P.O. Overexpression of FTL1/DDF1, an AP2 transcription factor, enhances tolerance to cold, drought, and heat stresses in Arabidopsis thaliana. Plant Sci. 2011, 180, 634–641. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Z.; Liu, F.; Sun, L.; Hao, F. Versatile Roles of Aquaporins in Plant Growth and Development. Int. J. Mol. Sci. 2020, 21, 9485. [Google Scholar] [CrossRef]
- Xu, Y.; Hu, W.; Liu, J.; Zhang, J.; Jia, C.; Miao, H.; Xu, B.; Jin, Z. A banana aquaporin gene, MaPIP1;1, is involved in tolerance to drought and salt stresses. BMC Plant Biol. 2014, 14, 59. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Liu, Y.; Feng, S.; Yang, J.; Li, D.; Zhang, J. Roles of Plasmalemma Aquaporin Gene StPIP1 in Enhancing Drought Tolerance in Potato. Front. Plant Sci. 2017, 8, 616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alavilli, H.; Awasthi, J.P.; Rout, G.R.; Sahoo, L.; Lee, B.-H.; Panda, S.K. Overexpression of a Barley Aquaporin Gene, HvPIP2;5 Confers Salt and Osmotic Stress Tolerance in Yeast and Plants. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Qiang, X.-J.; Han, X.-R.; Jiang, L.-L.; Zhang, S.-H.; Han, J.; He, R.; Cheng, X.-G. Ectopic Expression of a Thellungiella salsuginea Aquaporin Gene, TsPIP1;1, Increased the Salt Tolerance of Rice. Int. J. Mol. Sci. 2018, 19, 2229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Zhou, J.; Xiong, Y.; Liu, C.; Wang, J.; Wang, G.; Cai, Y. Overexpression of a maize plasma membrane intrinsic protein ZmPIP1;1 confers drought and salt tolerance in Arabidopsis. PLoS ONE 2018, 13, e0198639. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Gao, F.; Bing, J.; Sun, W.; Feng, X.; Ma, X.; Zhou, Y.; Zhang, G. Overexpression of the Jojoba Aquaporin Gene, ScPIP1, Enhances Drought and Salt Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2019, 20, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Hu, W.; Deng, X.; Ma, Z.; Chen, L.; Huang, C.; Wang, C.; Wang, J.; He, Y.; Yang, G.; et al. Overexpression of the Wheat Aquaporin Gene, TaAQP7, Enhances Drought Tolerance in Transgenic Tobacco. PLoS ONE 2012, 7, e52439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayadi, M.; Brini, F.; Masmoudi, K. Overexpression of a Wheat Aquaporin Gene, TdPIP2;1, Enhances Salt and Drought Tolerance in Transgenic Durum Wheat cv. Maali. Int. J. Mol. Sci. 2019, 20, 2389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apse, M.P.; Blumwald, E. Na+ transport in plants. FEBS Lett. 2007, 581, 2247–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.; He, X.; Zhao, B.; Zhou, C.; Liang, Y.; Ge, R.; Shen, Y.; Huang, Z. Overexpressing a Putative Aquaporin Gene from Wheat, TaNIP, Enhances Salt Tolerance in Transgenic Arabidopsis. Plant Cell Physiol. 2010, 51, 767–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Li, L.; Lou, Y.; Zhao, H.; Yang, Y.; Wang, S.; Gao, Z. The bamboo aquaporin gene PeTIP4;1–1 confers drought and salinity tolerance in transgenic Arabidopsis. Plant Cell Rep. 2017, 36, 597–609. [Google Scholar] [CrossRef]
- Hill, A.E.; Shachar-Hill, B. What Are Aquaporins For? J. Membr. Biol. 2004, 197, 1–32. [Google Scholar] [CrossRef]
- Macrobbie, E.A.C. Osmotic effects on vacuolar ion release in guard cells. Proc. Natl. Acad. Sci. USA 2006, 103, 1135–1140. [Google Scholar] [CrossRef] [Green Version]
- Ismail, A.; El-Sharkawy, I.; Sherif, S. Salt Stress Signals on Demand: Cellular Events in the Right Context. Int. J. Mol. Sci. 2020, 21, 3918. [Google Scholar] [CrossRef] [PubMed]
- Dang, Z.-H.; Zheng, L.-L.; Wang, J.; Gao, Z.; Wu, S.-B.; Qi, Z.; Wang, Y.-C. Transcriptomic profiling of the salt-stress response in the wild recretohalophyte Reaumuria trigyna. BMC Genom. 2013, 14, 29. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, N.; Watada, A.E. Effectiveness of various phenolic compounds in degradation of chlorophyll by In Vitro peroxi-dase-hydrogen peroxide system. J. Jpn. Soc. Hortic. Sci. 1994, 3, 439–444. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Yang, S.; Kong, X.; Wang, C.; Xiang, N.; Yang, Y.; Yang, Y. Molecular cloning of a plasma membrane aquaporin in Stipa purpurea, and exploration of its role in drought stress tolerance. Gene 2018, 665, 41–48. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Li, M.; Li, D.; Wang, S.-M.; Yin, H. Overexpression of the Zygophyllum xanthoxylum Aquaporin, ZxPIP1;3, Promotes Plant Growth and Stress Tolerance. Int. J. Mol. Sci. 2021, 22, 2112. https://doi.org/10.3390/ijms22042112
Li M, Li M, Li D, Wang S-M, Yin H. Overexpression of the Zygophyllum xanthoxylum Aquaporin, ZxPIP1;3, Promotes Plant Growth and Stress Tolerance. International Journal of Molecular Sciences. 2021; 22(4):2112. https://doi.org/10.3390/ijms22042112
Chicago/Turabian StyleLi, Mengzhan, Mingfa Li, Dingding Li, Suo-Min Wang, and Hongju Yin. 2021. "Overexpression of the Zygophyllum xanthoxylum Aquaporin, ZxPIP1;3, Promotes Plant Growth and Stress Tolerance" International Journal of Molecular Sciences 22, no. 4: 2112. https://doi.org/10.3390/ijms22042112
APA StyleLi, M., Li, M., Li, D., Wang, S. -M., & Yin, H. (2021). Overexpression of the Zygophyllum xanthoxylum Aquaporin, ZxPIP1;3, Promotes Plant Growth and Stress Tolerance. International Journal of Molecular Sciences, 22(4), 2112. https://doi.org/10.3390/ijms22042112