
Teneurins: Role in Cancer and Potential Role as Diagnostic Biomarkers and Targets for Therapy

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Common Structural and Functional Features of Mammalian Teneurins
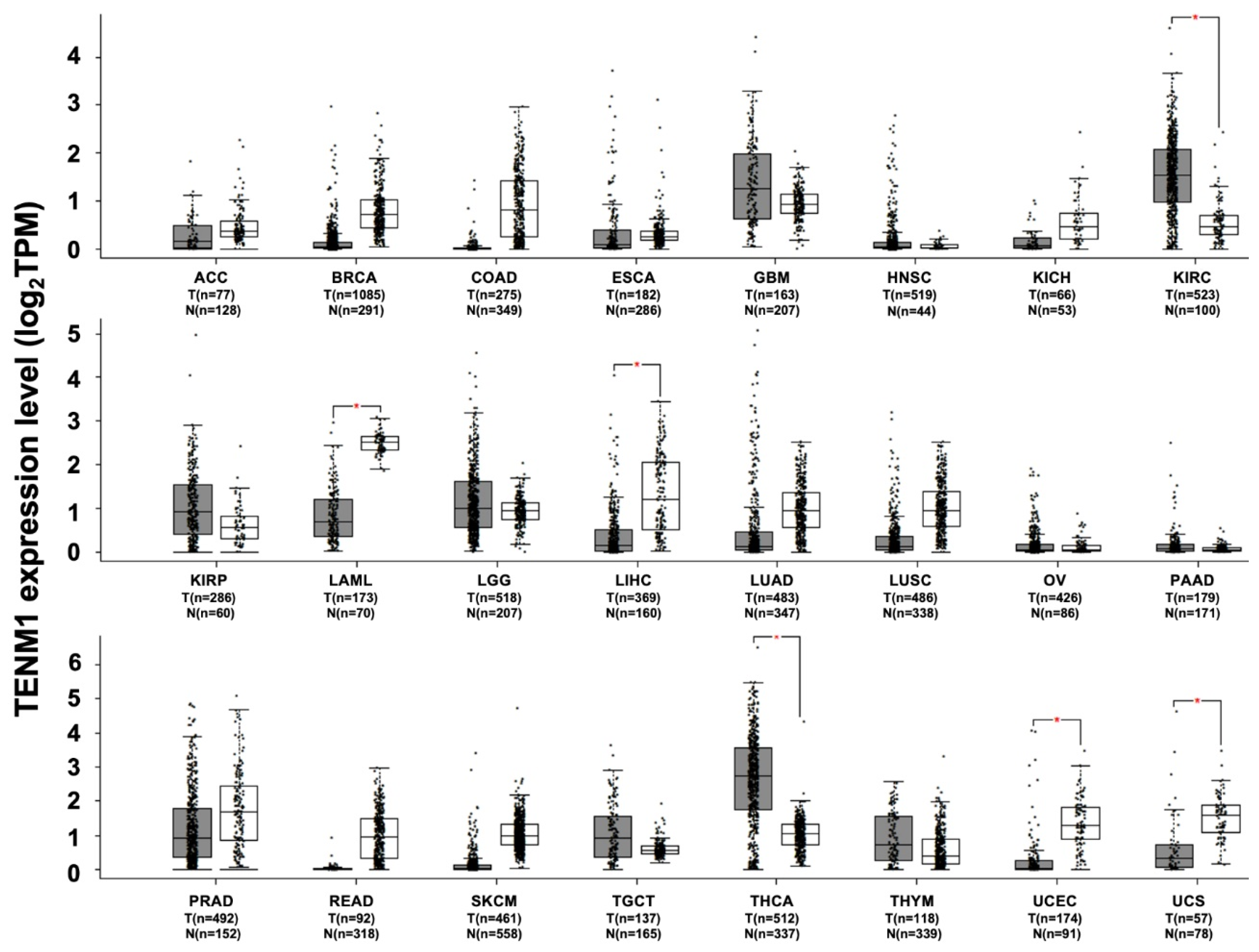
3. TENM1
TENM1 and Cancer
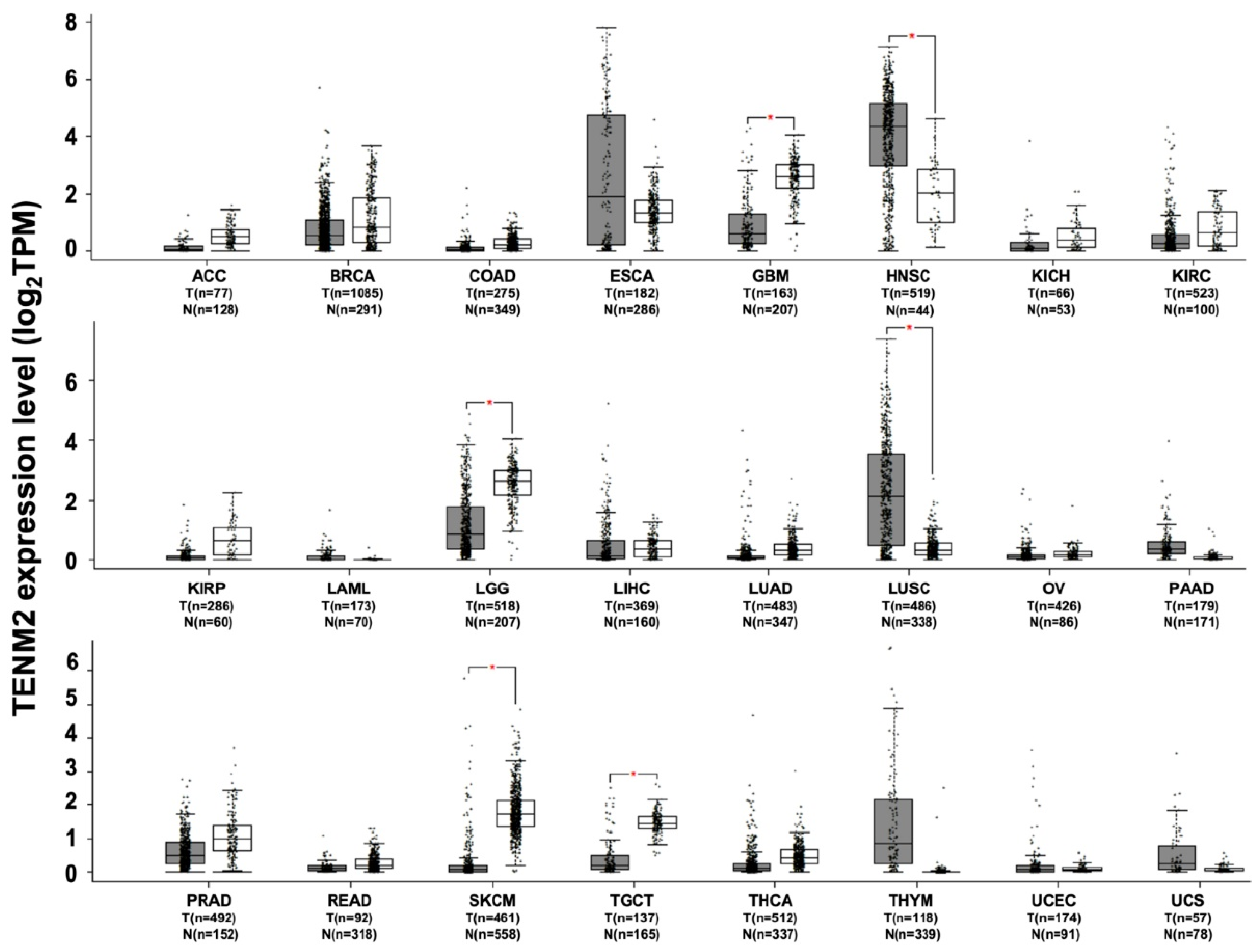
4. TENM2
TENM2 and Cancer
5. TENM3
TENM3 and Cancer
6. TENM4
TENM4 and Cancer
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Young, T.R.; Leamey, C.A. Teneurins: Important regulators of neural circuitry. Int. J. Biochem. Cell Biol. 2009, 41, 990–993. [Google Scholar] [CrossRef]
- Rubin, B.P.; Tucker, R.P.; Martin, D.; Chiquet-Ehrismann, R. Teneurins: A novel family of neuronal cell surface proteins in vertebrates, homologous to the Drosophila pair-rule gene product Ten-m. Dev. Biol. 1999, 216, 195–209. [Google Scholar] [CrossRef] [Green Version]
- Bagutti, C.; Forro, G.; Ferralli, J.; Rubin, B.; Chiquet-Ehrismann, R. The intracellular domain of teneurin-2 has a nuclear function and represses zic-1-mediated transcription. J. Cell Sci. 2003, 116 Pt 14, 2957–2966. [Google Scholar] [CrossRef] [Green Version]
- Nunes, S.M.; Ferralli, J.; Choi, K.; Brown-Luedi, M.; Minet, A.D.; Chiquet-Ehrismann, R. The intracellular domain of teneurin-1 interacts with MBD1 and CAP/ponsin resulting in subcellular codistribution and translocation to the nuclear matrix. Exp. Cell Res. 2005, 305, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Kenzelmann, D.; Chiquet-Ehrismann, R.; Leachman, N.T.; Tucker, R.P. Teneurin-1 is expressed in interconnected regions of the developing brain and is processed in vivo. BMC Dev. Biol. 2008, 8, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.Z.; Kuroda, M.; Sok, J.; Batchvarova, N.; Kimmel, R.; Chung, P.; Zinszner, H.; Ron, D. Identification of novel stress-induced genes downstream of chop. EMBO J. 1998, 17, 3619–3630. [Google Scholar] [CrossRef] [Green Version]
- Baumgartner, S.; Martin, D.; Hagios, C.; Chiquet-Ehrismann, R. Tenm, a Drosophila gene related to tenascin, is a new pair-rule gene. EMBO J. 1994, 13, 3728–3740. [Google Scholar] [CrossRef]
- Levine, A.; Bashan-Ahrend, A.; Budai-Hadrian, O.; Gartenberg, D.; Menasherow, S.; Wides, R. Odd Oz: A novel Drosophila pair rule gene. Cell 1994, 77, 587–598. [Google Scholar] [CrossRef]
- Zheng, L.; Michelson, Y.; Freger, V.; Avraham, Z.; Venken, K.J.; Bellen, H.J.; Justice, M.J.; Wides, R. Drosophila Ten-m and filamin affect motor neuron growth cone guidance. PLoS ONE 2011, 6, e22956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fascetti, N.; Baumgartner, S. Expression of Drosophila Ten-a, a dimeric receptor during embryonic development. Mech. Dev. 2002, 114, 197–200. [Google Scholar] [CrossRef]
- Hunding, A.; Baumgartner, S. Ancient role of ten-m/odz in segmentation and the transition from sequential to syncytial segmentation. Hereditas 2017, 154, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Zur, T.; Feige, E.; Motro, B.; Wides, R. The mammalian Odz gene family: Homologs of a Drosophila pair-rule gene with expression implying distinct yet overlapping developmental roles. Dev. Biol. 2000, 217, 107–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Bishop, K.M.; O’Leary, D.D. Potential target genes of EMX2 include Odz/Ten-M and other gene families with implications for cortical patterning. Mol. Cell Neurosci. 2006, 33, 136–149. [Google Scholar] [CrossRef]
- Zhou, X.H.; Brandau, O.; Feng, K.; Oohashi, T.; Ninomiya, Y.; Rauch, U.; Fassler, R. The murine Ten-m/Odz genes show distinct but overlapping expression patterns during development and in adult brain. Gene Expr. Patterns 2003, 3, 397–405. [Google Scholar] [CrossRef]
- Tran, H.; Sawatari, A.; Leamey, C.A. The glycoprotein Ten-m3 mediates topography and patterning of thalamostriatal projections from the parafascicular nucleus in mice. Eur. J. Neurosci. 2015, 41, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Bibollet-Bahena, O.; Okafuji, T.; Hokamp, K.; Tear, G.; Mitchell, K.J. A dual-strategy expression screen for candidate connectivity labels in the developing thalamus. PLoS ONE 2017, 12, e0177977. [Google Scholar] [CrossRef] [PubMed]
- Woelfle, R.; D’Aquila, A.L.; Pavlovic, T.; Husic, M.; Lovejoy, D.A. Ancient interaction between the teneurin C-terminal associated peptides (TCAP) and latrophilin ligand-receptor coupling: A role in behavior. Front. Neurosci. 2015, 9, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferralli, J.; Tucker, R.P.; Chiquet-Ehrismann, R. The teneurin C-terminal domain possesses nuclease activity and is apoptogenic. Biol. Open 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Tucker, R.P. Teneurins: Domain Architecture, Evolutionary Origins, and Patterns of Expression. Front. Neurosci. 2018, 12, 938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oohashi, T.; Zhou, X.H.; Feng, K.; Richter, B.; Morgelin, M.; Perez, M.T.; Su, W.D.; Chiquet-Ehrismann, R.; Rauch, U.; Fassler, R. Mouse ten-m/Odz is a new family of dimeric type II transmembrane proteins expressed in many tissues. J. Cell Biol. 1999, 145, 563–577. [Google Scholar] [CrossRef]
- Arac, D.; Li, J. Teneurins and latrophilins: Two giants meet at the synapse. Curr. Opin. Struct. Biol. 2019, 54, 141–151. [Google Scholar] [CrossRef]
- Hong, W.; Mosca, T.J.; Luo, L. Teneurins instruct synaptic partner matching in an olfactory map. Nature 2012, 484, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Mosca, T.J.; Hong, W.; Dani, V.S.; Favaloro, V.; Luo, L. Trans-synaptic Teneurin signalling in neuromuscular synapse organization and target choice. Nature 2012, 484, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Mosca, T.J. On the Teneurin track: A new synaptic organization molecule emerges. Front. Cell Neurosci. 2015, 9, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenzelmann, D.; Chiquet-Ehrismann, R.; Tucker, R.P. Teneurins, a transmembrane protein family involved in cell communication during neuronal development. Cell Mol. Life Sci. 2007, 64, 1452–1456. [Google Scholar] [CrossRef]
- Feng, K.; Zhou, X.H.; Oohashi, T.; Morgelin, M.; Lustig, A.; Hirakawa, S.; Ninomiya, Y.; Engel, J.; Rauch, U.; Fassler, R. All four members of the Ten-m/Odz family of transmembrane proteins form dimers. J. Biol. Chem. 2002, 277, 26128–26135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, J.P.; Lelianova, V.G.; Ermolyuk, Y.S.; Vysokov, N.; Hitchen, P.G.; Berninghausen, O.; Rahman, M.A.; Zangrandi, A.; Fidalgo, S.; Tonevitsky, A.G.; et al. Latrophilin 1 and its endogenous ligand Lasso/teneurin-2 form a high-affinity transsynaptic receptor pair with signaling capabilities. Proc. Natl. Acad. Sci. USA 2011, 108, 12113–12118. [Google Scholar] [CrossRef] [Green Version]
- Boucard, A.A.; Maxeiner, S.; Sudhof, T.C. Latrophilins function as heterophilic cell-adhesion molecules by binding to teneurins: Regulation by alternative splicing. J. Biol. Chem. 2014, 289, 387–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Shalev-Benami, M.; Sando, R.; Jiang, X.; Kibrom, A.; Wang, J.; Leon, K.; Katanski, C.; Nazarko, O.; Lu, Y.C.; et al. Structural Basis for Teneurin Function in Circuit-Wiring: A Toxin Motif at the Synapse. Cell 2018, 173, 735–748.e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Toro, D.; Carrasquero-Ordaz, M.A.; Chu, A.; Ruff, T.; Shahin, M.; Jackson, V.A.; Chavent, M.; Berbeira-Santana, M.; Seyit-Bremer, G.; Brignani, S.; et al. Structural Basis of Teneurin-Latrophilin Interaction in Repulsive Guidance of Migrating Neurons. Cell 2020, 180, 323–339.e19. [Google Scholar] [CrossRef] [Green Version]
- Berns, D.S.; DeNardo, L.A.; Pederick, D.T.; Luo, L. Teneurin-3 controls topographic circuit assembly in the hippocampus. Nature 2018, 554, 328–333. [Google Scholar] [CrossRef]
- Tucker, R.P.; Chiquet-Ehrismann, R.; Chevron, M.P.; Martin, D.; Hall, R.J.; Rubin, B.P. Teneurin-2 is expressed in tissues that regulate limb and somite pattern formation and is induced in vitro and in situ by FGF8. Dev. Dyn. 2001, 220, 27–39. [Google Scholar] [CrossRef]
- Qian, X.; Barsyte-Lovejoy, D.; Wang, L.; Chewpoy, B.; Gautam, N.; Al Chawaf, A.; Lovejoy, D.A. Cloning and characterization of teneurin C-terminus associated peptide (TCAP)-3 from the hypothalamus of an adult rainbow trout (Oncorhynchus mykiss). Gen. Comp. Endocrinol. 2004, 137, 205–216. [Google Scholar] [CrossRef]
- Ziegler, A.; Corvalan, A.; Roa, I.; Branes, J.A.; Wollscheid, B. Teneurin protein family: An emerging role in human tumorigenesis and drug resistance. Cancer Lett. 2012, 326, 1–7. [Google Scholar] [CrossRef]
- Rebolledo-Jaramillo, B.; Ziegler, A. Teneurins: An Integrative Molecular, Functional, and Biomedical Overview of Their Role in Cancer. Front. Neurosci. 2018, 12, 937. [Google Scholar] [CrossRef] [PubMed]
- Alkelai, A.; Olender, T.; Haffner-Krausz, R.; Tsoory, M.M.; Boyko, V.; Tatarskyy, P.; Gross-Isseroff, R.; Milgrom, R.; Shushan, S.; Blau, I.; et al. A role for TENM1 mutations in congenital general anosmia. Clin. Genet. 2016, 90, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Kioka, N. A novel adaptor protein family regulating cytoskeletal organization and signal transduction—Vinexin, CAP/ponsin, ArgBP2. Seikagaku 2002, 74, 1356–1360. [Google Scholar] [CrossRef] [Green Version]
- Hogg, D.W.; Husic, M.; Wosnick, D.; Dodsworth, T.; D’Aquila, A.L.; Lovejoy, D.A. Activity of the Carboxy-Terminal Peptide Region of the Teneurins and Its Role in Neuronal Function and Behavior in Mammals. Front. Neurosci. 2019, 13, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trubiani, G.; Al Chawaf, A.; Belsham, D.D.; Barsyte-Lovejoy, D.; Lovejoy, D.A. Teneurin carboxy (C)-terminal associated peptide-1 inhibits alkalosis-associated necrotic neuronal death by stimulating superoxide dismutase and catalase activity in immortalized mouse hypothalamic cells. Brain Res. 2007, 1176, 27–36. [Google Scholar] [CrossRef]
- Chand, D.; Song, L.; deLannoy, L.; Barsyte-Lovejoy, D.; Ackloo, S.; Boutros, P.C.; Evans, K.; Belsham, D.D.; Lovejoy, D.A. C-Terminal region of teneurin-1 co-localizes with dystroglycan and modulates cytoskeletal organization through an extracellular signal-regulated kinase-dependent stathmin- and filamin A-mediated mechanism in hippocampal cells. Neuroscience 2012, 219, 255–270. [Google Scholar] [CrossRef]
- Husic, M.; Barsyte-Lovejoy, D.; Lovejoy, D.A. Teneurin C-Terminal Associated Peptide (TCAP)-1 and Latrophilin Interaction in HEK293 Cells: Evidence for Modulation of Intercellular Adhesion. Front. Endocrinol. 2019, 10, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Chawaf, A.; St Amant, K.; Belsham, D.; Lovejoy, D.A. Regulation of neurite growth in immortalized mouse hypothalamic neurons and rat hippocampal primary cultures by teneurin C-terminal-associated peptide-1. Neuroscience 2007, 144, 1241–1254. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [Green Version]
- Mann, M.B.; Black, M.A.; Jones, D.J.; Ward, J.M.; Yew, C.C.; Newberg, J.Y.; Dupuy, A.J.; Rust, A.G.; Bosenberg, M.W.; McMahon, M.; et al. Transposon mutagenesis identifies genetic drivers of Braf(V600E) melanoma. Nat. Genet. 2015, 47, 486–495. [Google Scholar] [CrossRef] [Green Version]
- Mouly, E.; Ghamlouch, H.; Della-Valle, V.; Scourzic, L.; Quivoron, C.; Roos-Weil, D.; Pawlikowska, P.; Saada, V.; Diop, M.K.; Lopez, C.K.; et al. B-cell tumor development in Tet2-deficient mice. Blood Adv. 2018, 2, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Encinas, G.; Sabelnykova, V.Y.; de Lyra, E.C.; Hirata Katayama, M.L.; Maistro, S.; de Vasconcellos Valle, P.W.M.; de Lima Pereira, G.F.; Rodrigues, L.M.; de Menezes Pacheco Serio, P.A.; de Gouvea, A.; et al. Somatic mutations in early onset luminal breast cancer. Oncotarget 2018, 9, 22460–22479. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Burnett, M.G.; Shpak, M. A Comparative Study of the Molecular Characteristics of Familial Gliomas and Other Cancers. Cancer Genomics Proteomics 2016, 13, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Petrini, I.; Rajan, A.; Pham, T.; Voeller, D.; Davis, S.; Gao, J.; Wang, Y.; Giaccone, G. Whole genome and transcriptome sequencing of a B3 thymoma. PLoS ONE 2013, 8, e60572. [Google Scholar] [CrossRef]
- Yoshihara, K.; Wang, Q.; Torres-Garcia, W.; Zheng, S.; Vegesna, R.; Kim, H.; Verhaak, R.G. The landscape and therapeutic relevance of cancer-associated transcript fusions. Oncogene 2015, 34, 4845–4854. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Lee, K.; Yu, N.; Jang, I.; Choi, I.; Kim, P.; Jang, Y.E.; Kim, B.; Kim, S.; Lee, B.; et al. ChimerDB 3.0: An enhanced database for fusion genes from cancer transcriptome and literature data mining. Nucleic Acids Res. 2017, 45, D784–D789. [Google Scholar]
- Huang, Y.; Prasad, M.; Lemon, W.J.; Hampel, H.; Wright, F.A.; Kornacker, K.; LiVolsi, V.; Frankel, W.; Kloos, R.T.; Eng, C.; et al. Gene expression in papillary thyroid carcinoma reveals highly consistent profiles. Proc. Natl. Acad. Sci. USA 2001, 98, 15044–15049. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.P.; Chen, M.J.; Chien, M.N.; Lin, C.H.; Lee, J.J.; Liu, C.L. Overexpression of teneurin transmembrane protein 1 is a potential marker of disease progression in papillary thyroid carcinoma. Clin. Exp. Med. 2017, 17, 555–564. [Google Scholar] [CrossRef]
- Sun, Y.H.; Liu, Z.F.; Yang, B.B.; Yu, B. MicroRNA-486 inhibits cell proliferation, invasion and migration via down-regulating the TENM1 expressions and affecting ERK and Akt signaling pathways and epithelial-to-mesenchymal transition in papillary thyroid carcinoma. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8429–8439. [Google Scholar] [PubMed]
- Giannini, R.; Ugolini, C.; Poma, A.M.; Urpi, M.; Niccoli, C.; Elisei, R.; Chiarugi, M.; Vitti, P.; Miccoli, P.; Basolo, F. Identification of Two Distinct Molecular Subtypes of Non-Invasive Follicular Neoplasm with Papillary-Like Nuclear Features by Digital RNA Counting. Thyroid 2017, 27, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Hu, S.; Hou, Y.; He, Y.; Liu, S. Identification of potential novel biomarkers to differentiate malignant thyroid nodules with cytological indeterminate. BMC Cancer 2020, 20, 199. [Google Scholar] [CrossRef]
- Zhang, W.; Zang, Z.; Song, Y.; Yang, H.; Yin, Q. Co-expression network analysis of differentially expressed genes associated with metastasis in prolactin pituaitary tumors. Mol. Med. Rep. 2014, 10, 113–118. [Google Scholar] [CrossRef]
- Talamillo, A.; Grande, L.; Ruiz-Ontanon, P.; Velasquez, C.; Mollinedo, P.; Torices, S.; Sanchez-Gomez, P.; Aznar, A.; Esparis-Ogando, A.; Lopez-Lopez, C.; et al. ODZ1 allows glioblastoma to sustain invasiveness through a Myc-dependent transcriptional upregulation of RhoA. Oncogene 2017, 36, 1733–1744. [Google Scholar] [CrossRef] [Green Version]
- Velasquez, C.; Mansouri, S.; Gutierrez, O.; Mamatjan, Y.; Mollinedo, P.; Karimi, S.; Singh, O.; Teran, N.; Martino, J.; Zadeh, G.; et al. Hypoxia Can Induce Migration of Glioblastoma Cells Through a Methylation-Dependent Control of ODZ1 Gene Expression. Front. Oncol. 2019, 9, 1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, J.; Liu, X.; Laffin, B.; Chen, X.; Choy, G.; Jeter, C.R.; Calhoun-Davis, T.; Li, H.; Palapattu, G.S.; Pang, S.; et al. The PSA(-/lo) prostate cancer cell population harbors self-renewing long-term tumor-propagating cells that resist castration. Cell Stem Cell 2012, 10, 556–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tajima, T.; Kito, F.; Yoshida, A.; Kawai, A.; Kondo, T. Calreticulin as A Novel Potential Metastasis-Associated Protein in Myxoid Liposarcoma, as Revealed by Two-Dimensional Difference Gel Electrophoresis. Proteomes 2019, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Ni, X.; Zhuo, M.; Su, Z.; Duan, J.; Gao, Y.; Wang, Z.; Zong, C.; Bai, H.; Chapman, A.R.; Zhao, J.; et al. Reproducible copy number variation patterns among single circulating tumor cells of lung cancer patients. Proc. Natl. Acad. Sci. USA 2013, 110, 21083–21088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leamey, C.A.; Merlin, S.; Lattouf, P.; Sawatari, A.; Zhou, X.; Demel, N.; Glendining, K.A.; Oohashi, T.; Sur, M.; Fassler, R. Ten_m3 regulates eye-specific patterning in the mammalian visual pathway and is required for binocular vision. PLoS Biol. 2007, 5, e241. [Google Scholar] [CrossRef]
- Young, T.R.; Bourke, M.; Zhou, X.; Oohashi, T.; Sawatari, A.; Fassler, R.; Leamey, C.A. Ten-m2 is required for the generation of binocular visual circuits. J. NeuroSci. 2013, 33, 12490–12509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-da-Silva, K.R.; Tessarin, G.W.L.; Dias, C.A.; Guiati, I.Z.; Ervolino, E.; Goncalves, A.; Beneti, I.M.; Lovejoy, D.A.; Casatti, C.A. Teneurin-2 presence in rat and human odontoblasts. PLoS ONE 2017, 12, e0184794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tews, D.; Schwar, V.; Scheithauer, M.; Weber, T.; Fromme, T.; Klingenspor, M.; Barth, T.F.; Moller, P.; Holzmann, K.; Debatin, K.M.; et al. Comparative gene array analysis of progenitor cells from human paired deep neck and subcutaneous adipose tissue. Mol. Cell Endocrinol. 2014, 395, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Kneller, J.M.; Ehlen, T.; Matisic, J.P.; Miller, D.; Van Niekerk, D.; Lam, W.L.; Marra, M.; Richards-Kortum, R.; Follen, M.; Macaulay, C.; et al. Using LongSAGE to Detect Biomarkers of Cervical Cancer Potentially Amenable to Optical Contrast Agent Labelling. Biomark. Insights 2007, 2, 447–461. [Google Scholar] [CrossRef]
- Kan, T.; Yamasaki, S.; Kondo, K.; Teratani, N.; Kawabe, A.; Kaganoi, J.; Meltzer, S.J.; Imamura, M.; Shimada, Y. A new specific gene expression in squamous cell carcinoma of the esophagus detected using representational difference analysis and cDNA microarray. Oncology 2006, 70, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, A.; Cerciello, F.; Bigosch, C.; Bausch-Fluck, D.; Felley-Bosco, E.; Ossola, R.; Soltermann, A.; Stahel, R.A.; Wollscheid, B. Proteomic surfaceome analysis of mesothelioma. Lung Cancer 2012, 75, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Minami, M.; Daimon, Y.; Mori, K.; Takashima, H.; Nakajima, T.; Itoh, Y.; Okanoue, T. Hepatitis B virus-related insertional mutagenesis in chronic hepatitis B patients as an early drastic genetic change leading to hepatocarcinogenesis. Oncogene 2005, 24, 4340–4348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varambally, S.; Yu, J.; Laxman, B.; Rhodes, D.R.; Mehra, R.; Tomlins, S.A.; Shah, R.B.; Chandran, U.; Monzon, F.A.; Becich, M.J.; et al. Integrative genomic and proteomic analysis of prostate cancer reveals signatures of metastatic progression. Cancer Cell 2005, 8, 393–406. [Google Scholar] [CrossRef] [Green Version]
- Noordhuis, M.G.; Fehrmann, R.S.; Wisman, G.B.; Nijhuis, E.R.; van Zanden, J.J.; Moerland, P.D.; Ver Loren van Themaat, E.; Volders, H.H.; Kok, M.; ten Hoor, K.A.; et al. Involvement of the TGF-beta and beta-catenin pathways in pelvic lymph node metastasis in early-stage cervical cancer. Clin. Cancer Res. 2011, 17, 1317–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Medina, D.; Tsimelzon, A.; Mohsin, S.K.; Mao, S.; Wu, Y.; Allred, D.C. Alterations of gene expression in the development of early hyperplastic precursors of breast cancer. Am. J. Pathol. 2007, 171, 252–262. [Google Scholar] [CrossRef] [Green Version]
- Graumann, R.; Di Capua, G.A.; Oyarzun, J.E.; Vasquez, M.A.; Liao, C.; Branes, J.A.; Roa, I.; Casanello, P.; Corvalan, A.H.; Owen, G.I.; et al. Expression of teneurins is associated with tumor differentiation and patient survival in ovarian cancer. PLoS ONE 2017, 12, e0177244. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, L.M.; Frederiksen, K.S.; Din, N.; Galsgaard, E.; Christensen, L.; Berchtold, M.W.; Panina, S. Prolactin and oestrogen synergistically regulate gene expression and proliferation of breast cancer cells. Endocr. Relat. Cancer 2010, 17, 809–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maturi, V.; Enroth, S.; Heldin, C.H.; Moustakas, A. Genome-wide binding of transcription factor ZEB1 in triple-negative breast cancer cells. J. Cell Physiol. 2018, 233, 7113–7127. [Google Scholar] [CrossRef] [Green Version]
- Buys, T.P.; Chari, R.; Lee, E.H.; Zhang, M.; MacAulay, C.; Lam, S.; Lam, W.L.; Ling, V. Genetic changes in the evolution of multidrug resistance for cultured human ovarian cancer cells. Genes Chromosomes Cancer 2007, 46, 1069–1079. [Google Scholar] [CrossRef]
- Mizuuchi, H.; Suda, K.; Sato, K.; Tomida, S.; Fujita, Y.; Kobayashi, Y.; Maehara, Y.; Sekido, Y.; Nishio, K.; Mitsudomi, T. Collateral chemoresistance to anti-microtubule agents in a lung cancer cell line with acquired resistance to erlotinib. PLoS ONE 2015, 10, e0123901. [Google Scholar] [CrossRef] [Green Version]
- Huret, J.L.; Ahmad, M.; Arsaban, M.; Bernheim, A.; Cigna, J.; Desangles, F.; Guignard, J.C.; Jacquemot-Perbal, M.C.; Labarussias, M.; Leberre, V.; et al. Atlas of genetics and cytogenetics in oncology and haematology in 2013. Nucleic Acids Res. 2013, 41, D920–D924. [Google Scholar] [CrossRef]
- Pugh, T.J.; Morozova, O.; Attiyeh, E.F.; Asgharzadeh, S.; Wei, J.S.; Auclair, D.; Carter, S.L.; Cibulskis, K.; Hanna, M.; Kiezun, A.; et al. The genetic landscape of high-risk neuroblastoma. Nat. Genet. 2013, 45, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Banerji, S.; Cibulskis, K.; Rangel-Escareno, C.; Brown, K.K.; Carter, S.L.; Frederick, A.M.; Lawrence, M.S.; Sivachenko, A.Y.; Sougnez, C.; Zou, L.; et al. Sequence analysis of mutations and translocations across breast cancer subtypes. Nature 2012, 486, 405–409. [Google Scholar] [CrossRef]
- Mercier, I.; Casimiro, M.C.; Wang, C.; Rosenberg, A.L.; Quong, J.; Minkeu, A.; Allen, K.G.; Danilo, C.; Sotgia, F.; Bonuccelli, G.; et al. Human breast cancer-associated fibroblasts (CAFs) show caveolin-1 downregulation and RB tumor suppressor functional inactivation: Implications for the response to hormonal therapy. Cancer Biol. Ther. 2008, 7, 1212–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGuire, T.F.; Sajithlal, G.B.; Lu, J.; Nicholls, R.D.; Prochownik, E.V. In vivo evolution of tumor-derived endothelial cells. PLoS ONE 2012, 7, e37138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leamey, C.A.; Sawatari, A. Teneurins: Mediators of Complex Neural Circuit Assembly in Mammals. Front. Neurosci. 2019, 13, 580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antinucci, P.; Nikolaou, N.; Meyer, M.P.; Hindges, R. Teneurin-3 specifies morphological and functional connectivity of retinal ganglion cells in the vertebrate visual system. Cell Rep. 2013, 5, 582–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Zur, T.; Wides, R. Mapping homologs of Drosophila odd Oz (odz): Doc4/Odz4 to mouse chromosome 7, Odz1 to mouse chromosome 11; and ODZ3 to human chromosome Xq25. Genomics 1999, 58, 102–103. [Google Scholar] [CrossRef]
- Bastias-Candia, S.; Martinez, M.; Zolezzi, J.M.; Inestrosa, N.C. Wnt Signaling Upregulates Teneurin-3 Expression via Canonical and Non-canonical Wnt Pathway Crosstalk. Front. Neurosci. 2019, 13, 505. [Google Scholar] [CrossRef]
- Niell, N.; Larriba, M.J.; Ferrer-Mayorga, G.; Sanchez-Perez, I.; Cantero, R.; Real, F.X.; Del Peso, L.; Munoz, A.; Gonzalez-Sancho, J.M. The human PKP2/plakophilin-2 gene is induced by Wnt/beta-catenin in normal and colon cancer-associated fibroblasts. Int. J. Cancer 2018, 142, 792–804. [Google Scholar] [CrossRef] [Green Version]
- Jackson, V.A.; Meijer, D.H.; Carrasquero, M.; van Bezouwen, L.S.; Lowe, E.D.; Kleanthous, C.; Janssen, B.J.C.; Seiradake, E. Structures of Teneurin adhesion receptors reveal an ancient fold for cell-cell interaction. Nat. Commun. 2018, 9, 1079. [Google Scholar] [CrossRef] [Green Version]
- Lovejoy, D.A.; Barsyte-Lovejoy, D. Systems approaches to genomic and epigenetic inter-regulation of peptide hormones in stress and reproduction. Prog. Biophys. Mol. Biol. 2013, 113, 375–386. [Google Scholar] [CrossRef]
- Orentas, R.J.; Yang, J.J.; Wen, X.; Wei, J.S.; Mackall, C.L.; Khan, J. Identification of cell surface proteins as potential immunotherapy targets in 12 pediatric cancers. Front. Oncol. 2012, 2, 194. [Google Scholar] [CrossRef] [Green Version]
- Garcia, J.; Faca, V.; Jarzembowski, J.; Zhang, Q.; Park, J.; Hanash, S. Comprehensive profiling of the cell surface proteome of Sy5Y neuroblastoma cells yields a subset of proteins associated with tumor differentiation. J. Proteome Res. 2009, 8, 3791–3796. [Google Scholar] [CrossRef] [PubMed]
- Balogh, G.A.; Heulings, R.; Mailo, D.A.; Russo, P.A.; Sheriff, F.; Russo, I.H.; Moral, R.; Russo, J. Genomic signature induced by pregnancy in the human breast. Int. J. Oncol. 2006, 28, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Russo, J.; Mailo, D.; Hu, Y.F.; Balogh, G.; Sheriff, F.; Russo, I.H. Breast differentiation and its implication in cancer prevention. Clin. Cancer Res. 2005, 11 Pt 2, 931s–936s. [Google Scholar]
- Caruso, J.A.; Campana, R.; Wei, C.; Su, C.H.; Hanks, A.M.; Bornmann, W.G.; Keyomarsi, K. Indole-3-carbinol and its N-alkoxy derivatives preferentially target ERalpha-positive breast cancer cells. Cell Cycle 2014, 13, 2587–2599. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.Y.; Oskarsson, T.; Acharyya, S.; Nguyen, D.X.; Zhang, X.H.; Norton, L.; Massague, J. Tumor self-seeding by circulating cancer cells. Cell 2009, 139, 1315–1326. [Google Scholar] [CrossRef] [Green Version]
- Fleron, M.; Greffe, Y.; Musmeci, D.; Massart, A.C.; Hennequiere, V.; Mazzucchelli, G.; Waltregny, D.; De Pauw-Gillet, M.C.; Castronovo, V.; De Pauw, E.; et al. Novel post-digest isotope coded protein labeling method for phospho- and glycoproteome analysis. J. Proteomics 2010, 73, 1986–2005. [Google Scholar] [CrossRef]
- You, H.J.; Park, H.Y.; Kim, J.; Lee, I.H.; Seol, H.J.; Lee, J.I.; Kim, S.T.; Kong, D.S.; Nam, D.H. Integrative radiogenomic analysis for genomic signatures in glioblastomas presenting leptomeningeal dissemination. Medicine 2016, 95, e4109. [Google Scholar] [CrossRef]
- Molenaar, J.J.; Koster, J.; Zwijnenburg, D.A.; van Sluis, P.; Valentijn, L.J.; van der Ploeg, I.; Hamdi, M.; van Nes, J.; Westerman, B.A.; van Arkel, J.; et al. Sequencing of neuroblastoma identifies chromothripsis and defects in neuritogenesis genes. Nature 2012, 483, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.K.; Shen, K.; McBride, A.A. Papillomavirus genomes associate with BRD4 to replicate at fragile sites in the host genome. PLoS Pathog. 2014, 10, e1004117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexandrescu, S.; Korshunov, A.; Lai, S.H.; Dabiri, S.; Patil, S.; Li, R.; Shih, C.S.; Bonnin, J.M.; Baker, J.A.; Du, E.; et al. Epithelioid Glioblastomas and Anaplastic Epithelioid Pleomorphic Xanthoastrocytomas--Same Entity or First Cousins? Brain Pathol. 2016, 26, 215–223. [Google Scholar] [CrossRef]
- Hu, W.; Li, X.; Wang, T.; Zheng, S. Association mining of mutated cancer genes in different clinical stages across 11 cancer types. Oncotarget 2016, 7, 68270–68277. [Google Scholar] [CrossRef] [Green Version]
- Cannataro, V.L.; Gaffney, S.G.; Townsend, J.P. Effect Sizes of Somatic Mutations in Cancer. J. Natl. Cancer Inst. 2018, 110, 1171–1177. [Google Scholar] [CrossRef] [Green Version]
- Tanyi, J.L.; Bobisse, S.; Ophir, E.; Tuyaerts, S.; Roberti, A.; Genolet, R.; Baumgartner, P.; Stevenson, B.J.; Iseli, C.; Dangaj, D.; et al. Personalized cancer vaccine effectively mobilizes antitumor T cell immunity in ovarian cancer. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- Cobb, J.; Duboule, D. Comparative analysis of genes downstream of the Hoxd cluster in developing digits and external genitalia. Development 2005, 132, 3055–3067. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Numakawa, T.; Chou, J.; de Vega, S.; Mizuniwa, C.; Sekimoto, K.; Adachi, N.; Kunugi, H.; Arikawa-Hirasawa, E.; Yamada, Y.; et al. Teneurin-4 promotes cellular protrusion formation and neurite outgrowth through focal adhesion kinase signaling. FASEB J. 2014, 28, 1386–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, N.; Fukushi, M.; Kosaki, K.; Doyle, A.D.; de Vega, S.; Yoshizaki, K.; Akazawa, C.; Arikawa-Hirasawa, E.; Yamada, Y. Teneurin-4 is a novel regulator of oligodendrocyte differentiation and myelination of small-diameter axons in the CNS. J. Neurosci. 2012, 32, 11586–11599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hor, H.; Francescatto, L.; Bartesaghi, L.; Ortega-Cubero, S.; Kousi, M.; Lorenzo-Betancor, O.; Jimenez-Jimenez, F.J.; Gironell, A.; Clarimon, J.; Drechsel, O.; et al. Missense mutations in TENM4, a regulator of axon guidance and central myelination, cause essential tremor. Hum. Mol. Genet. 2015, 24, 5677–5686. [Google Scholar] [CrossRef] [Green Version]
- Houle, G.; Schmouth, J.F.; Leblond, C.S.; Ambalavanan, A.; Spiegelman, D.; Laurent, S.B.; Bourassa, C.V.; Panisset, M.; Chouinard, S.; Dupre, N.; et al. Teneurin transmembrane protein 4 is not a cause for essential tremor in a Canadian population. Mov. Disord. 2017, 32, 292–295. [Google Scholar] [CrossRef]
- Suzuki, N.; Mizuniwa, C.; Ishii, K.; Nakagawa, Y.; Tsuji, K.; Muneta, T.; Sekiya, I.; Akazawa, C. Teneurin-4, a transmembrane protein, is a novel regulator that suppresses chondrogenic differentiation. J. Orthop. Res. 2014, 32, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Brohl, D.; Vasyutina, E.; Czajkowski, M.T.; Griger, J.; Rassek, C.; Rahn, H.P.; Purfurst, B.; Wende, H.; Birchmeier, C. Colonization of the satellite cell niche by skeletal muscle progenitor cells depends on Notch signals. Dev. Cell 2012, 23, 469–481. [Google Scholar] [CrossRef] [Green Version]
- Ishii, K.; Suzuki, N.; Mabuchi, Y.; Ito, N.; Kikura, N.; Fukada, S.; Okano, H.; Takeda, S.; Akazawa, C. Muscle Satellite Cell Protein Teneurin-4 Regulates Differentiation During Muscle Regeneration. Stem Cells 2015, 33, 3017–3027. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Wang, X.; Chen, Y.; Wang, X.; Zhu, J.; Lu, L. Expression quantitative trait loci and genetic regulatory network analysis reveals that Gabra2 is involved in stress responses in the mouse. Stress 2009, 12, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Agurto, R.; Dickson, A.J. Multiplexed Digital mRNA Expression Analysis Profiles System-Wide Changes in mRNA Abundance and Responsiveness of UPR-Specific Gene Expression Changes during Batch Culture of Recombinant Chinese Hamster Ovary Cells. Biotechnol. J. 2018, 13, e1700429. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Davis, N.A.; White, B.C.; Pajewski, N.M.; Savitz, J.; Drevets, W.C.; McKinney, B.A. Epistasis network centrality analysis yields pathway replication across two GWAS cohorts for bipolar disorder. Transl. Psychiatry 2012, 2, e154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivorra, J.L.; Rivero, O.; Costas, J.; Iniesta, R.; Arrojo, M.; Ramos-Rios, R.; Carracedo, A.; Palomo, T.; Rodriguez-Jimenez, R.; Cervilla, J.; et al. Replication of previous genome-wide association studies of psychiatric diseases in a large schizophrenia case-control sample from Spain. Schizophr. Res. 2014, 159, 107–113. [Google Scholar] [CrossRef]
- Xue, C.B.; Xu, Z.H.; Zhu, J.; Wu, Y.; Zhuang, X.H.; Chen, Q.L.; Wu, C.R.; Hu, J.T.; Zhou, H.S.; Xie, W.H.; et al. Exome Sequencing Identifies TENM4 as a Novel Candidate Gene for Schizophrenia in the SCZD2 Locus at 11q14-21. Front. Genet. 2018, 9, 725. [Google Scholar] [CrossRef]
- Heinrich, A.; Lourdusamy, A.; Tzschoppe, J.; Vollstadt-Klein, S.; Buhler, M.; Steiner, S.; Bach, C.; Poustka, L.; Banaschewski, T.; Barker, G.; et al. The risk variant in ODZ4 for bipolar disorder impacts on amygdala activation during reward processing. Bipolar Disord. 2013, 15, 440–445. [Google Scholar] [CrossRef]
- Croarkin, P.E.; Luby, J.L.; Cercy, K.; Geske, J.R.; Veldic, M.; Simonson, M.; Joshi, P.T.; Wagner, K.D.; Walkup, J.T.; Nassan, M.M.; et al. Genetic Risk Score Analysis in Early-Onset Bipolar Disorder. J. Clin. Psychiatry 2017, 78, 1337–1343. [Google Scholar] [CrossRef] [Green Version]
- De Robertis, M.; Arigoni, M.; Loiacono, L.; Riccardo, F.; Calogero, R.A.; Feodorova, Y.; Tashkova, D.; Belovejdov, V.; Sarafian, V.; Cavallo, F.; et al. Novel insights into Notum and glypicans regulation in colorectal cancer. Oncotarget 2015, 6, 41237–41257. [Google Scholar] [CrossRef] [PubMed]
- Quaglino, E.; Conti, L.; Cavallo, F. Breast cancer stem cell antigens as targets for immunotherapy. Semin. Immunol. 2020, 47, 101386. [Google Scholar] [CrossRef]
- Ruiu, R.; Barutello, G.; Arigoni, M.; Riccardo, F.; Conti, L.; Peppino, G.; Annaratone, L.; Marchiò, C.; Mengozzi, G.; Calogero, R.A.; et al. Identification of TENM4 as a Novel Cancer Stem Cell-Associated Molecule and Potential Target in Triple Negative Breast Cancer. Cancers 2021, 13, 894. [Google Scholar] [CrossRef]
- Wang, X.Z.; Jolicoeur, E.M.; Conte, N.; Chaffanet, M.; Zhang, Y.; Mozziconacci, M.J.; Feiner, H.; Birnbaum, D.; Pebusque, M.J.; Ron, D. gamma-heregulin is the product of a chromosomal translocation fusing the DOC4 and HGL/NRG1 genes in the MDA-MB-175 breast cancer cell line. Oncogene 1999, 18, 5718–5721. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Baker, E.; Eyre, H.J.; Sutherland, G.R.; Zhou, M. Gamma-heregulin: A fusion gene of DOC-4 and neuregulin-1 derived from a chromosome translocation. Oncogene 1999, 18, 7110–7114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, G.; Fitzpatrick, V.D.; Sliwkowski, M.X. Gamma-heregulin: A novel heregulin isoform that is an autocrine growth factor for the human breast cancer cell line, MDA-MB-175. Oncogene 1997, 15, 1385–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, W.E.; Sliwkowski, M.X.; Akita, R.W.; Henzel, W.J.; Lee, J.; Park, J.W.; Yansura, D.; Abadi, N.; Raab, H.; Lewis, G.D.; et al. Identification of heregulin, a specific activator of p185erbB2. Science 1992, 256, 1205–1210. [Google Scholar] [CrossRef] [PubMed]
- George, J.; Lim, J.S.; Jang, S.J.; Cun, Y.; Ozretic, L.; Kong, G.; Leenders, F.; Lu, X.; Fernandez-Cuesta, L.; Bosco, G.; et al. Comprehensive genomic profiles of small cell lung cancer. Nature 2015, 524, 47–53. [Google Scholar] [CrossRef]
- Puente, X.S.; Bea, S.; Valdes-Mas, R.; Villamor, N.; Gutierrez-Abril, J.; Martin-Subero, J.I.; Munar, M.; Rubio-Perez, C.; Jares, P.; Aymerich, M.; et al. Non-coding recurrent mutations in chronic lymphocytic leukaemia. Nature 2015, 526, 519–524. [Google Scholar] [CrossRef]
- Marimuthu, A.; O’Meally, R.N.; Chaerkady, R.; Subbannayya, Y.; Nanjappa, V.; Kumar, P.; Kelkar, D.S.; Pinto, S.M.; Sharma, R.; Renuse, S.; et al. A comprehensive map of the human urinary proteome. J. Proteome Res. 2011, 10, 2734–2743. [Google Scholar] [CrossRef] [Green Version]
- Gangoda, L.; Keerthikumar, S.; Fonseka, P.; Edgington, L.E.; Ang, C.S.; Ozcitti, C.; Bogyo, M.; Parker, B.S.; Mathivanan, S. Inhibition of cathepsin proteases attenuates migration and sensitizes aggressive N-Myc amplified human neuroblastoma cells to doxorubicin. Oncotarget 2015, 6, 11175–11190. [Google Scholar] [CrossRef] [Green Version]
- Keerthikumar, S.; Gangoda, L.; Liem, M.; Fonseka, P.; Atukorala, I.; Ozcitti, C.; Mechler, A.; Adda, C.G.; Ang, C.S.; Mathivanan, S. Proteogenomic analysis reveals exosomes are more oncogenic than ectosomes. Oncotarget 2015, 6, 15375–15396. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peppino, G.; Ruiu, R.; Arigoni, M.; Riccardo, F.; Iacoviello, A.; Barutello, G.; Quaglino, E. Teneurins: Role in Cancer and Potential Role as Diagnostic Biomarkers and Targets for Therapy. Int. J. Mol. Sci. 2021, 22, 2321. https://doi.org/10.3390/ijms22052321
Peppino G, Ruiu R, Arigoni M, Riccardo F, Iacoviello A, Barutello G, Quaglino E. Teneurins: Role in Cancer and Potential Role as Diagnostic Biomarkers and Targets for Therapy. International Journal of Molecular Sciences. 2021; 22(5):2321. https://doi.org/10.3390/ijms22052321
Chicago/Turabian StylePeppino, Giulia, Roberto Ruiu, Maddalena Arigoni, Federica Riccardo, Antonella Iacoviello, Giuseppina Barutello, and Elena Quaglino. 2021. "Teneurins: Role in Cancer and Potential Role as Diagnostic Biomarkers and Targets for Therapy" International Journal of Molecular Sciences 22, no. 5: 2321. https://doi.org/10.3390/ijms22052321
APA StylePeppino, G., Ruiu, R., Arigoni, M., Riccardo, F., Iacoviello, A., Barutello, G., & Quaglino, E. (2021). Teneurins: Role in Cancer and Potential Role as Diagnostic Biomarkers and Targets for Therapy. International Journal of Molecular Sciences, 22(5), 2321. https://doi.org/10.3390/ijms22052321