
MicroRNAs Modulate Signaling Pathways in Osteogenic Differentiation of Mesenchymal Stem Cells

 , , ,
, , ,  , ,
, , 
 ,
, 
Abstract
:1. Introduction
2. MSC Signaling Pathways in Osteogenic Differentiation
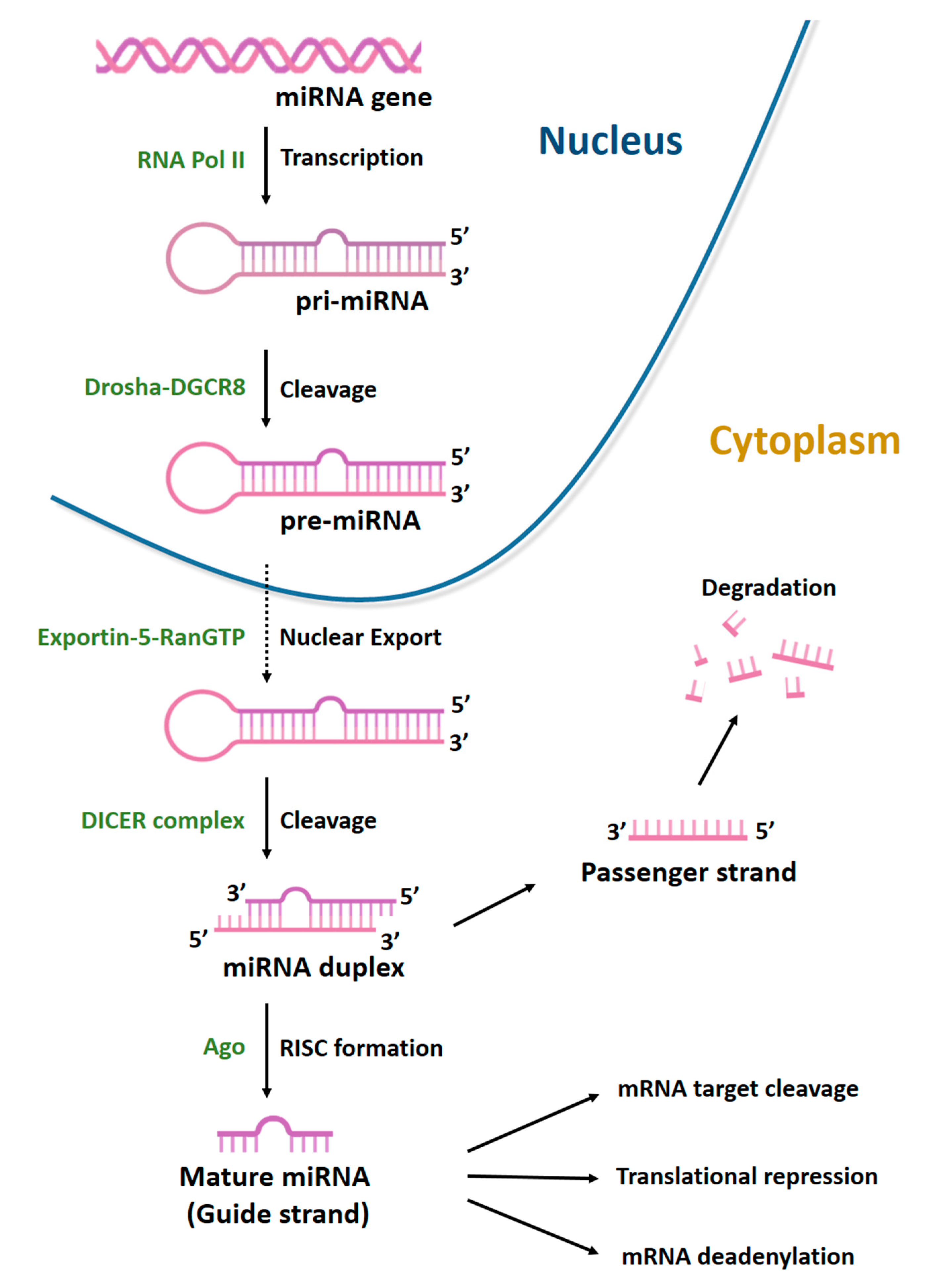
3. MiRNAs
4. Osteogenic Differentiation by miRNA Regulation
4.1. TGF-β/BMP Signaling Pathways
4.1.1. TGF-β/BMP Ligands and Receptors
4.1.2. SMAD Cascade
4.1.3. TGF-β/BMP Signaling Pathway Intracellular and Extracellular Inhibitors: SMAD7, HDAC4, BMP Antagonist Proteins
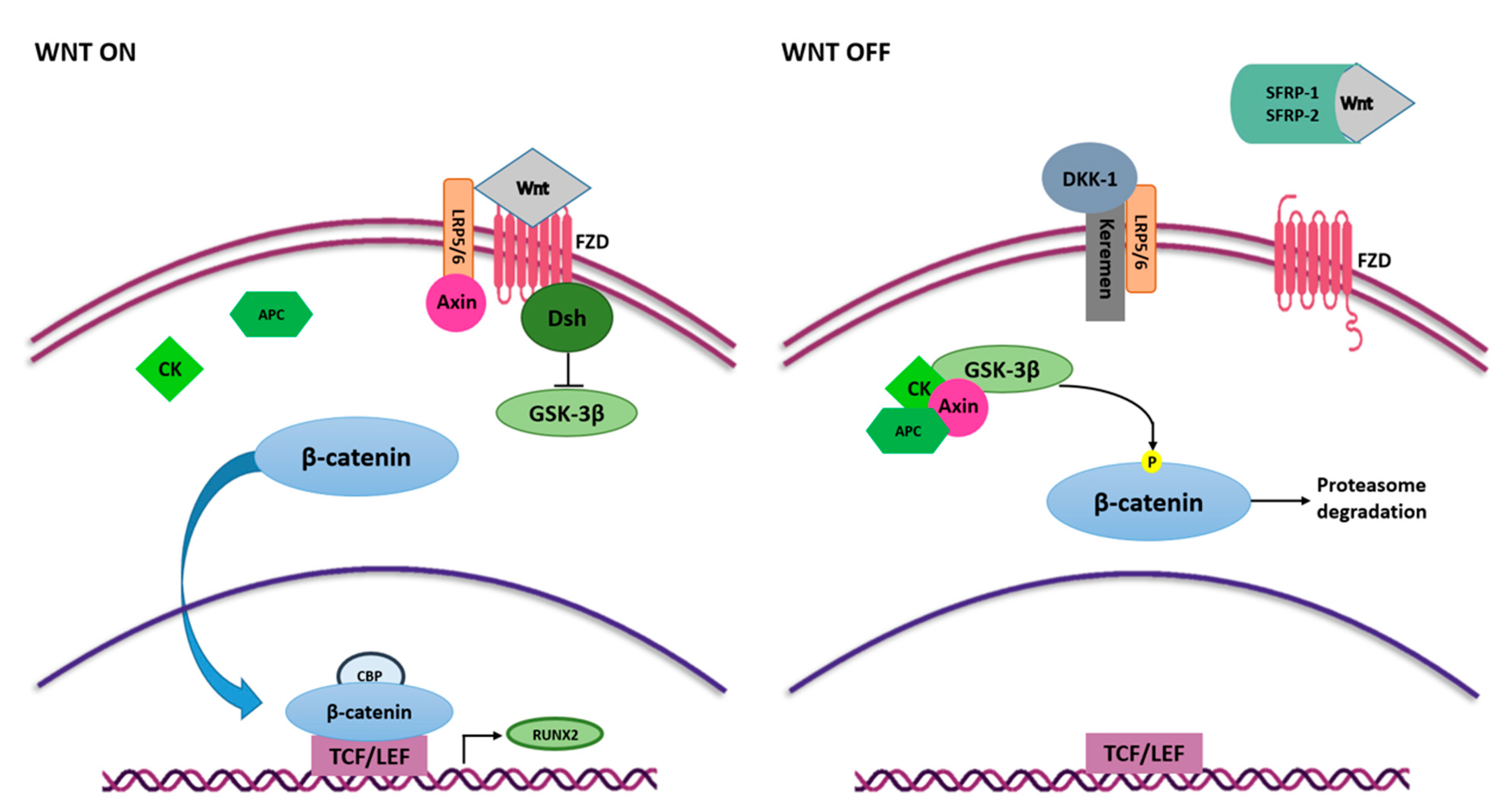
4.2. Wnt/β-Catenin Signaling Pathway
4.2.1. Wnt Ligands and Receptors
4.2.2. Transcription Factors: β-Catenin and TCF/LEF
4.2.3. Wnt Cascade Inhibitors
4.2.4. Transcription Factor RUNX2
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chen, Q.; Shou, P.; Zheng, C.; Jiang, M.; Cao, G.; Yang, Q.; Cao, J.; Xie, N.; Velletri, T.; Zhang, X.; et al. Fate Decision of Mesenchymal Stem Cells: Adipocytes or Osteoblasts? Cell Death Differ. 2016, 23, 1128–1139. [Google Scholar] [CrossRef] [Green Version]
- Mazzoni, E.; D’Agostino, A.; Iaquinta, M.R.; Bononi, I.; Trevisiol, L.; Rotondo, J.C.; Patergnani, S.; Giorgi, C.; Gunson, M.J.; Arnett, G.W.; et al. Hydroxylapatite-Collagen Hybrid Scaffold Induces Human Adipose-Derived Mesenchymal Stem Cells to Osteogenic Differentiation in Vitro and Bone Regrowth in Patients. Stem Cells Transl. Med. 2020, 9, 377–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedenstein, A.J.; Petrakova, K.V.; Kurolesova, A.I.; Frolova, G.P. Heterotopic of Bone Marrow. Analysis of Precursor Cells for Osteogenic and Hematopoietic Tissues. Transplantation 1968, 6, 230–247. [Google Scholar] [CrossRef]
- Mortada, I.; Mortada, R. Epigenetic Changes in Mesenchymal Stem Cells Differentiation. Eur. J. Med. Genet. 2018, 61, 114–118. [Google Scholar] [CrossRef]
- Iaquinta, M.R.; Mazzoni, E.; Bononi, I.; Rotondo, J.C.; Mazziotta, C.; Montesi, M.; Sprio, S.; Tampieri, A.; Tognon, M.; Martini, F. Adult Stem Cells for Bone Regeneration and Repair. Front. Cell Dev. Biol. 2019, 7, 268. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Li, J.; Zou, Q.; Zuo, Y.; Cai, B.; Li, Y. Enhanced Bone Tissue Regeneration of a Biomimetic Cellular Scaffold with Co-Cultured MSCs-Derived Osteogenic and Angiogenic Cells. Cell Prolif. 2019, 52, e12658. [Google Scholar] [CrossRef] [Green Version]
- Di Benedetto, A.; Brunetti, G.; Posa, F.; Ballini, A.; Grassi, F.R.; Colaianni, G.; Colucci, S.; Rossi, E.; Cavalcanti-Adam, E.A.; Lo Muzio, L.; et al. Osteogenic Differentiation of Mesenchymal Stem Cells from Dental Bud: Role of Integrins and Cadherins. Stem Cell Res. 2015, 15, 618–628. [Google Scholar] [CrossRef] [Green Version]
- Klontzas, M.E.; Reakasame, S.; Silva, R.; Morais, J.C.F.; Vernardis, S.; MacFarlane, R.J.; Heliotis, M.; Tsiridis, E.; Panoskaltsis, N.; Boccaccini, A.R.; et al. Oxidized Alginate Hydrogels with the GHK Peptide Enhance Cord Blood Mesenchymal Stem Cell Osteogenesis: A Paradigm for Metabolomics-Based Evaluation of Biomaterial Design. Acta Biomater. 2019, 88, 224–240. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Madaleno, C.; Jatzlau, J.; Knaus, P. BMP Signalling in a Mechanical Context—Implications for Bone Biology. Bone 2020, 137, 115416. [Google Scholar] [CrossRef] [PubMed]
- Varani, K.; Vincenzi, F.; Pasquini, S.; Blo, I.; Salati, S.; Cadossi, M.; De Mattei, M. Pulsed Electromagnetic Field Stimulation in Osteogenesis and Chondrogenesis: Signaling Pathways and Therapeutic Implications. Int. J. Mol. Sci. 2021, 22, 809. [Google Scholar] [CrossRef]
- Barbanti Brodano, G.; Mazzoni, E.; Tognon, M.; Griffoni, C.; Manfrini, M. Human Mesenchymal Stem Cells and Biomaterials Interaction: A Promising Synergy to Improve Spine Fusion. Eur. Spine J. 2012, 21 (Suppl. 1), S3–S9. [Google Scholar] [CrossRef] [Green Version]
- Manfrini, M.; Di Bona, C.; Canella, A.; Lucarelli, E.; Pellati, A.; D’Agostino, A.; Barbanti-Bròdano, G.; Tognon, M. Mesenchymal Stem Cells from Patients to Assay Bone Graft Substitutes. J. Cell. Physiol. 2013, 228, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Manfrini, M.; Mazzoni, E.; Barbanti-Brodano, G.; Nocini, P.; D’agostino, A.; Trombelli, L.; Tognon, M. Osteoconductivity of Complex Biomaterials Assayed by Fluorescent-Engineered Osteoblast-like Cells. Cell Biochem. Biophys. 2015, 71, 1509–1515. [Google Scholar] [CrossRef] [PubMed]
- Barbanti Bròdano, G.; Griffoni, C.; Nataloni, A.; Manfrini, M.; Giavaresi, G.; Bandiera, S.; Gasbarrini, A.; Terzi, S.; Ghermandi, R.; Tedesco, G.; et al. Biomaterials as Bone Graft Substitutes for Spine Surgery: From Preclinical Results to Clinical Study. J. Biol. Regul. Homeost. Agents 2017, 31, 167–181. [Google Scholar]
- Mazzoni, E.; D’Agostino, A.; Manfrini, M.; Maniero, S.; Puozzo, A.; Bassi, E.; Marsico, S.; Fortini, C.; Trevisiol, L.; Patergnani, S.; et al. Human Adipose Stem Cells Induced to Osteogenic Differentiation by an Innovative Collagen/Hydroxylapatite Hybrid Scaffold. FASEB J. 2017, 31, 4555–4565. [Google Scholar] [CrossRef] [Green Version]
- Mazzoni, E.; Mazziotta, C.; Iaquinta, M.R.; Lanzillotti, C.; Fortini, F.; D’Agostino, A.; Trevisiol, L.; Nocini, R.; Barbanti-Brodano, G.; Mescola, A.; et al. Enhanced Osteogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells by a Hybrid Hydroxylapatite/Collagen Scaffold. Front. Cell Dev. Biol. 2021, 8. [Google Scholar] [CrossRef]
- Mazzoni, E.; Iaquinta, M.R.; Lanzillotti, C.; Mazziotta, C.; Maritati, M.; Montesi, M.; Sprio, S.; Tampieri, A.; Tognon, M.; Martini, F. Bioactive Materials for Soft Tissue Repair. Front. Bioeng. Biotechnol. 2021. [Google Scholar] [CrossRef]
- Globig, P.; Willumeit-Römer, R.; Martini, F.; Mazzoni, E.; Luthringer-Feyerabend, B.J.C. Optimizing an Osteosarcoma-Fibroblast Coculture Model to Study Antitumoral Activity of Magnesium-Based Biomaterials. Int. J. Mol. Sci. 2020, 21, 5099. [Google Scholar] [CrossRef] [PubMed]
- Sprio, S.; Dapporto, M.; Preti, L.; Mazzoni, E.; Iaquinta, M.R.; Martini, F.; Tognon, M.; Pugno, N.M.; Restivo, E.; Visai, L.; et al. Enhancement of the Biological and Mechanical Performances of Sintered Hydroxyapatite by Multiple Ions Doping. Front. Mater. 2020, 7. [Google Scholar] [CrossRef]
- Alarçin, E.; Lee, T.Y.; Karuthedom, S.; Mohammadi, M.; Brennan, M.A.; Lee, D.H.; Marrella, A.; Zhang, J.; Syla, D.; Zhang, Y.S.; et al. Injectable Shear-Thinning Hydrogels for Delivering Osteogenic and Angiogenic Cells and Growth Factors. Biomater. Sci. 2018, 6, 1604–1615. [Google Scholar] [CrossRef]
- Yang, C.; Liu, X.; Zhao, K.; Zhu, Y.; Hu, B.; Zhou, Y.; Wang, M.; Wu, Y.; Zhang, C.; Xu, J.; et al. MiRNA-21 Promotes Osteogenesis via the PTEN/PI3K/Akt/HIF-1α Pathway and Enhances Bone Regeneration in Critical Size Defects. Stem Cell Res. Ther. 2019, 10, 65. [Google Scholar] [CrossRef] [PubMed]
- Fassina, L.; Bloise, N.; Montagna, G.; Visai, L.; Mognaschi, M.E.; Benazzo, F.; Magenes, G. Biomaterials and Biophysical Stimuli for Bone Regeneration. J. Biol. Regul. Homeost. Agents 2018, 32, 41–49. [Google Scholar] [PubMed]
- Cakouros, D.; Gronthos, S. Epigenetic Regulators of Mesenchymal Stem/Stromal Cell Lineage Determination. Curr. Osteoporos. Rep. 2020, 18, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Ju, C.; Liu, R.; Zhang, Y.-W.; Zhang, Y.; Zhou, R.; Sun, J.; Lv, X.-B.; Zhang, Z. Mesenchymal Stem Cell-Associated LncRNA in Osteogenic Differentiation. Biomed. Pharmacother. 2019, 115, 108912. [Google Scholar] [CrossRef]
- Martin, E.C.; Qureshi, A.T.; Dasa, V.; Freitas, M.A.; Gimble, J.M.; Davis, T.A. MicroRNA Regulation of Stem Cell Differentiation and Diseases of the Bone and Adipose Tissue: Perspectives on MiRNA Biogenesis and Cellular Transcriptome. Biochimie 2016, 124, 98–111. [Google Scholar] [CrossRef]
- Fu, G.; Ren, A.; Qiu, Y.; Zhang, Y. Epigenetic Regulation of Osteogenic Differentiation of Mesenchymal Stem Cells. Curr. Stem Cell Res. Ther. 2016, 11, 235–246. [Google Scholar] [CrossRef]
- Lanzillotti, C.; De Mattei, M.; Mazziotta, C.; Taraballi, F.; Rotondo, J.C.; Tognon, M.; Martini, F. Interplay between Long Non-Coding RNAs and Micro RNAs in Osteogenic Differentiation of Mesenchymal Stem Cells. Front. Cell Dev. Biol. 2021, in press. [Google Scholar]
- Ostuni, R.; Natoli, G.; Cassatella, M.A.; Tamassia, N. Epigenetic Regulation of Neutrophil Development and Function. Semin. Immunol. 2016, 28, 83–93. [Google Scholar] [CrossRef]
- Chirayil, R.; Kincaid, R.P.; Dahlke, C.; Kuny, C.V.; Dälken, N.; Spohn, M.; Lawson, B.; Grundhoff, A.; Sullivan, C.S. Identification of Virus-Encoded MicroRNAs in Divergent Papillomaviruses. PLoS Pathog. 2018, 14, e1007156. [Google Scholar] [CrossRef] [Green Version]
- Vojtechova, Z.; Tachezy, R. The Role of MiRNAs in Virus-Mediated Oncogenesis. Int. J. Mol. Sci. 2018, 19, 1217. [Google Scholar] [CrossRef] [Green Version]
- Rotondo, J.C.; Mazzoni, E.; Bononi, I.; Tognon, M.; Martini, F. Association Between Simian Virus 40 and Human Tumors. Front. Oncol. 2019, 9, 670. [Google Scholar] [CrossRef]
- Ledda, B.; Ottaggio, L.; Izzotti, A.; Sukkar, S.G.; Miele, M. Small RNAs in Eucaryotes: New Clues for Amplifying MicroRNA Benefits. Cell Biosci. 2020, 10, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hensley, A.P.; McAlinden, A. The Role of MicroRNAs in Bone Development. Bone 2021, 143, 115760. [Google Scholar] [CrossRef]
- Mirzaei, H.; Fathullahzadeh, S.; Khanmohammadi, R.; Darijani, M.; Momeni, F.; Masoudifar, A.; Goodarzi, M.; Mardanshah, O.; Stenvang, J.; Jaafari, M.R.; et al. State of the Art in MicroRNA as Diagnostic and Therapeutic Biomarkers in Chronic Lymphocytic Leukemia. J. Cell. Physiol. 2018, 233, 888–900. [Google Scholar] [CrossRef] [PubMed]
- Rotondo, J.C.; Selvatici, R.; Di Domenico, M.; Marci, R.; Vesce, F.; Tognon, M.; Martini, F. Methylation Loss at H19 Imprinted Gene Correlates with Methylenetetrahydrofolate Reductase Gene Promoter Hypermethylation in Semen Samples from Infertile Males. Epigenetics 2013, 8, 990–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gámez, B.; Rodriguez-Carballo, E.; Ventura, F. MicroRNAs and Post-Transcriptional Regulation of Skeletal Development. J. Mol. Endocrinol. 2014, 52, R179–R197. [Google Scholar] [CrossRef] [Green Version]
- Peng, S.; Gao, D.; Gao, C.; Wei, P.; Niu, M.; Shuai, C. MicroRNAs Regulate Signaling Pathways in Osteogenic Differentiation of Mesenchymal Stem Cells (Review). Mol. Med. Rep. 2016, 14, 623–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Geng, J.; Jiang, S. MicroRNAs in Regulation of Osteogenic Differentiation of Mesenchymal Stem Cells. Cell Tissue Res. 2017, 368, 229–238. [Google Scholar] [CrossRef]
- Riffo-Campos, Á.L.; Riquelme, I.; Brebi-Mieville, P. Tools for Sequence-Based MiRNA Target Prediction: What to Choose? Int. J. Mol. Sci. 2016, 17, 1987. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Yang, L.; Wang, W.; Wang, J.; Wang, J.; Xu, Z. Discovery and Validation of Extracellular/Circulating MicroRNAs during Idiopathic Pulmonary Fibrosis Disease Progression. Gene 2015, 562, 138–144. [Google Scholar] [CrossRef]
- Tomasello, L.; Cluts, L.; Croce, C.M. Experimental Validation of MicroRNA Targets: Analysis of MicroRNA Targets Through Western Blotting. Methods Mol. Biol. 2019, 1970, 341–353. [Google Scholar] [CrossRef]
- Yoshioka, H.; Yoshiko, Y. The Roles of Long Non-Protein-Coding RNAs in Osteo-Adipogenic Lineage Commitment. Int. J. Mol. Sci. 2017, 18, 1236. [Google Scholar] [CrossRef] [Green Version]
- Ferronato, S.; Lira, M.G.; Olivato, S.; Scuro, A.; Veraldi, G.F.; Romanelli, M.G.; Patuzzo, C.; Malerba, G.; Pignatti, P.F.; Mazzucco, S. Upregulated Expression of Toll-like Receptor 4 in Peripheral Blood of Ischaemic Stroke Patients Correlates with Cyclooxygenase 2 Expression. Eur. J. Vasc. Endovasc. Surg. 2011, 41, 358–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Chen, G.; Li, Y.-P. TGF-β and BMP Signaling in Osteoblast, Skeletal Development, and Bone Formation, Homeostasis and Disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef]
- Liu, J.; Dang, L.; Wu, X.; Li, D.; Ren, Q.; Lu, A.; Zhang, G. MicroRNA-Mediated Regulation of Bone Remodeling: A Brief Review. JBMR Plus 2019, 3, e10213. [Google Scholar] [CrossRef]
- Moghaddam, T.; Neshati, Z. Role of MicroRNAs in Osteogenesis of Stem Cells. J. Cell. Biochem. 2019, 120, 14136–14155. [Google Scholar] [CrossRef] [PubMed]
- Majidinia, M.; Sadeghpour, A.; Yousefi, B. The Roles of Signaling Pathways in Bone Repair and Regeneration. J. Cell. Physiol. 2018, 233, 2937–2948. [Google Scholar] [CrossRef] [PubMed]
- Poon, B.; Kha, T.; Tran, S.; Dass, C.R. Bone Morphogenetic Protein-2 and Bone Therapy: Successes and Pitfalls. J. Pharm. Pharmacol. 2016, 68, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Halloran, D.; Durbano, H.W.; Nohe, A. Bone Morphogenetic Protein-2 in Development and Bone Homeostasis. J. Dev. Biol. 2020, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Schupbach, D.; Comeau-Gauthier, M.; Harvey, E.; Merle, G. Wnt Modulation in Bone Healing. Bone 2020, 138, 115491. [Google Scholar] [CrossRef]
- Rahman, M.S.; Akhtar, N.; Jamil, H.M.; Banik, R.S.; Asaduzzaman, S.M. TGF-β/BMP Signaling and Other Molecular Events: Regulation of Osteoblastogenesis and Bone Formation. Bone Res. 2015, 3, 15005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslani, S.; Abhari, A.; Sakhinia, E.; Sanajou, D.; Rajabi, H.; Rahimzadeh, S. Interplay between MicroRNAs and Wnt, Transforming Growth Factor-β, and Bone Morphogenic Protein Signaling Pathways Promote Osteoblastic Differentiation of Mesenchymal Stem Cells. J. Cell. Physiol. 2019, 234, 8082–8093. [Google Scholar] [CrossRef]
- Sampath, T.K.; Reddi, A.H. Discovery of Bone Morphogenetic Proteins—A Historical Perspective. Bone 2020, 140, 115548. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Shi, P.; Tu, M.; Wang, Y.; Liu, M.; Fan, F.; Du, M. Bone Morphogenetic Proteins: Relationship between Molecular Structure and Their Osteogenic Activity. Food Sci. Hum. Wellness 2014, 3, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Martini, F.; Pellati, A.; Mazzoni, E.; Salati, S.; Caruso, G.; Contartese, D.; De Mattei, M. Bone Morphogenetic Protein-2 Signaling in the Osteogenic Differentiation of Human Bone Marrow Mesenchymal Stem Cells Induced by Pulsed Electromagnetic Fields. Int. J. Mol. Sci. 2020, 21, 2104. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Puerto, M.C.; Iyengar, P.V.; García de Vinuesa, A.; Ten Dijke, P.; Sanchez-Duffhues, G. Bone Morphogenetic Protein Receptor Signal Transduction in Human Disease. J. Pathol. 2019, 247, 9–20. [Google Scholar] [CrossRef]
- Li, Z.; Xu, Z.; Duan, C.; Liu, W.; Sun, J.; Han, B. Role of TCF/LEF Transcription Factors in Bone Development and Osteogenesis. Int. J. Med. Sci. 2018, 15, 1415–1422. [Google Scholar] [CrossRef] [Green Version]
- Laxman, N.; Mallmin, H.; Nilsson, O.; Kindmark, A. MiR-203 and MiR-320 Regulate Bone Morphogenetic Protein-2-Induced Osteoblast Differentiation by Targeting Distal-Less Homeobox 5 (Dlx5). Genes 2017, 8, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, B.T.; He, X. Frizzled and LRP5/6 Receptors for Wnt/β-Catenin Signaling. Cold Spring Harb. Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef]
- Houschyar, K.S.; Tapking, C.; Borrelli, M.R.; Popp, D.; Duscher, D.; Maan, Z.N.; Chelliah, M.P.; Li, J.; Harati, K.; Wallner, C.; et al. Wnt Pathway in Bone Repair and Regeneration—What Do We Know So Far. Front. Cell Dev. Biol. 2019, 6. [Google Scholar] [CrossRef]
- Aulicino, F.; Pedone, E.; Sottile, F.; Lluis, F.; Marucci, L.; Cosma, M.P. Canonical Wnt Pathway Controls MESC Self-Renewal Through Inhibition of Spontaneous Differentiation via β-Catenin/TCF/LEF Functions. Stem Cell Rep. 2020, 15, 646–661. [Google Scholar] [CrossRef]
- Chang, J.; Sonoyama, W.; Wang, Z.; Jin, Q.; Zhang, C.; Krebsbach, P.H.; Giannobile, W.; Shi, S.; Wang, C.-Y. Noncanonical Wnt-4 Signaling Enhances Bone Regeneration of Mesenchymal Stem Cells in Craniofacial Defects through Activation of P38 MAPK. J. Biol. Chem. 2007, 282, 30938–30948. [Google Scholar] [CrossRef] [Green Version]
- Cawthorn, W.P.; Bree, A.J.; Yao, Y.; Du, B.; Hemati, N.; Martinez-Santibañez, G.; MacDougald, O.A. Wnt6, Wnt10a and Wnt10b Inhibit Adipogenesis and Stimulate Osteoblastogenesis through a β-Catenin-Dependent Mechanism. Bone 2012, 50, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M.S.; Oyserman, S.M.; Hankenson, K.D. Wnt11 Promotes Osteoblast Maturation and Mineralization through R-Spondin 2. J. Biol. Chem. 2009, 284, 14117–14125. [Google Scholar] [CrossRef] [Green Version]
- Boland, G.M.; Perkins, G.; Hall, D.J.; Tuan, R.S. Wnt 3a Promotes Proliferation and Suppresses Osteogenic Differentiation of Adult Human Mesenchymal Stem Cells. J. Cell. Biochem. 2004, 93, 1210–1230. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Von den Hoff, J.W.; Torensma, R.; Meng, L.; Bian, Z. Wnt16 Is Involved in Intramembranous Ossification and Suppresses Osteoblast Differentiation through the Wnt/β-Catenin Pathway. J. Cell. Physiol. 2014, 229, 384–392. [Google Scholar] [CrossRef]
- Ahmadzadeh, A.; Norozi, F.; Shahrabi, S.; Shahjahani, M.; Saki, N. Wnt/β-Catenin Signaling in Bone Marrow Niche. Cell Tissue Res. 2016, 363, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Rotondo, J.C.; Bosi, S.; Bassi, C.; Ferracin, M.; Lanza, G.; Gafà, R.; Magri, E.; Selvatici, R.; Torresani, S.; Marci, R.; et al. Gene Expression Changes in Progression of Cervical Neoplasia Revealed by Microarray Analysis of Cervical Neoplastic Keratinocytes. J. Cell. Physiol. 2015, 230, 806–812. [Google Scholar] [CrossRef] [PubMed]
- Corazza, M.; Oton-Gonzalez, L.; Scuderi, V.; Rotondo, J.C.; Lanzillotti, C.; Di Mauro, G.; Tognon, M.; Martini, F.; Borghi, A. Tissue Cytokine/Chemokine Profile in Vulvar Lichen Sclerosus: An Observational Study on Keratinocyte and Fibroblast Cultures. J. Dermatol. Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Aulicino, F.; Theka, I.; Ombrato, L.; Lluis, F.; Cosma, M.P. Temporal Perturbation of the Wnt Signaling Pathway in the Control of Cell Reprogramming Is Modulated by TCF1. Stem Cell Rep. 2014, 2, 707–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamos, J.L.; Weis, W.I. The β-Catenin Destruction Complex. Cold Spring Harb. Perspect. Biol. 2013, 5, a007898. [Google Scholar] [CrossRef] [PubMed]
- Holdsworth, G.; Roberts, S.J.; Ke, H.Z. Novel Actions of Sclerostin on Bone. J. Mol. Endocrinol. 2019, 62, R167–R185. [Google Scholar] [CrossRef] [Green Version]
- Brunner, M.; Mandier, N.; Gautier, T.; Chevalier, G.; Ribba, A.-S.; Guardiola, P.; Block, M.R.; Bouvard, D. Β1 Integrins Mediate the BMP2 Dependent Transcriptional Control of Osteoblast Differentiation and Osteogenesis. PLoS ONE 2018, 13, e0196021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finotti, A.; Fabbri, E.; Lampronti, I.; Gasparello, J.; Borgatti, M.; Gambari, R. MicroRNAs and Long Non-Coding RNAs in Genetic Diseases. Mol. Diagn. Ther. 2019, 23, 155–171. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-C.; Xu, Z.; Zhang, T.-F.; Wang, Y.-L. Circulating MicroRNAs as Diagnostic and Prognostic Tools for Hepatocellular Carcinoma. World J. Gastroenterol. 2015, 21, 9853–9862. [Google Scholar] [CrossRef] [PubMed]
- Bär, C.; Thum, T.; Gonzalo-Calvo, D. de Circulating MiRNAs as Mediators in Cell-to-Cell Communication. Epigenomics 2019, 11, 111–113. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating MicroRNAs as Potential Cancer Biomarkers: The Advantage and Disadvantage. Clin. Epigenetics 2018, 10, 59. [Google Scholar] [CrossRef] [Green Version]
- Desantis, V.; Saltarella, I.; Lamanuzzi, A.; Melaccio, A.; Solimando, A.G.; Mariggiò, M.A.; Racanelli, V.; Paradiso, A.; Vacca, A.; Frassanito, M.A. MicroRNAs-Based Nano-Strategies as New Therapeutic Approach in Multiple Myeloma to Overcome Disease Progression and Drug Resistance. Int. J. Mol. Sci. 2020, 21, 3084. [Google Scholar] [CrossRef] [PubMed]
- Arriaga, M.A.; Ding, M.; Gutierrez, A.S.; Chew, S.A. The Application of MicroRNAs in Biomaterial Scaffold-Based Therapies for Bone Tissue Engineering. Biotechnol. J. 2019, 14, 1900084. [Google Scholar] [CrossRef]
- Iaquinta, M.R.; Mazzoni, E.; Manfrini, M.; D’Agostino, A.; Trevisiol, L.; Nocini, R.; Trombelli, L.; Barbanti-Brodano, G.; Martini, F.; Tognon, M. Innovative Biomaterials for Bone Regrowth. Int. J. Mol. Sci. 2019, 20, 618. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Dou, Q.; Ha, X. Let-7a-5p Inhibits BMSCs Osteogenesis in Postmenopausal Osteoporosis Mice. Biochem. Biophys. Res. Commun. 2019, 510, 53–58. [Google Scholar] [CrossRef]
- Zeng, Y.; Qu, X.; Li, H.; Huang, S.; Wang, S.; Xu, Q.; Lin, R.; Han, Q.; Li, J.; Zhao, R.C. MicroRNA-100 Regulates Osteogenic Differentiation of Human Adipose-Derived Mesenchymal Stem Cells by Targeting BMPR2. FEBS Lett. 2012, 586, 2375–2381. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Lv, Q.; Lv, C. MicroRNA-153 Suppresses the Osteogenic Differentiation of Human Mesenchymal Stem Cells by Targeting Bone Morphogenetic Protein Receptor Type II. Int. J. Mol. Med. 2015, 36, 760–766. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhong, L.; Yuan, T.; Chen, S.; Zhou, Y.; An, L.; Guo, Y.; Fan, M.; Li, Y.; Sun, Y.; et al. MicroRNA-155 Inhibits the Osteogenic Differentiation of Mesenchymal Stem Cells Induced by BMP9 via Downregulation of BMP Signaling Pathway. Int. J. Mol. Med. 2018, 41, 3379–3393. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.; Lin, H.; Fu, H.; Wang, B.; Han, G.; Fan, M. MicroRNA-195-5p Regulates Osteogenic Differentiation of Periodontal Ligament Cells Under Mechanical Loading. J. Cell. Physiol. 2017, 232, 3762–3774. [Google Scholar] [CrossRef]
- Wang, H.; Xie, Z.; Hou, T.; Li, Z.; Huang, K.; Gong, J.; Zhou, W.; Tang, K.; Xu, J.; Dong, S. MiR-125b Regulates the Osteogenic Differentiation of Human Mesenchymal Stem Cells by Targeting BMPR1b. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 41, 530–542. [Google Scholar] [CrossRef]
- Arfat, Y.; Basra, M.A.R.; Shahzad, M.; Majeed, K.; Mahmood, N.; Munir, H. MiR-208a-3p Suppresses Osteoblast Differentiation and Inhibits Bone Formation by Targeting ACVR1. Mol. Ther. Nucleic Acids 2018, 11, 323–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.; Park, S.-K.; Lee, H.Y.; Kim, S.W.; Lee, J.S.; Choi, E.K.; You, D.; Kim, C.-S.; Suh, N. MiR-140-5p Suppresses BMP2-Mediated Osteogenesis in Undifferentiated Human Mesenchymal Stem Cells. FEBS Lett. 2014, 588, 2957–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wei, Q.-S.; Ding, W.-B.; Zhang, L.-L.; Wang, H.-C.; Zhu, Y.-J.; He, W.; Chai, Y.-N.; Liu, Y.-W. Increased MicroRNA-93-5p Inhibits Osteogenic Differentiation by Targeting Bone Morphogenetic Protein-2. PLoS ONE 2017, 12, e0182678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.-P.; Zhang, J.; Zhu, C.-H.; Lin, L.; Wang, J.; Zhang, H.-J.; Li, J.; Yu, X.-G.; Zhao, Z.-S.; Dong, W.; et al. MicroRNA-98 Regulates Osteogenic Differentiation of Human Bone Mesenchymal Stromal Cells by Targeting BMP2. J. Cell. Mol. Med. 2017, 21, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Ando, M.; Tsukamasa, Y.; Akao, Y. Expression of BMP-2 and Ets1 in BMP-2-Stimulated Mouse Pre-Osteoblast Differentiation Is Regulated by MicroRNA-370. FEBS Lett. 2012, 586, 1693–1701. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-G.; Liao, Z.; Xiao, H.; Liu, H.; Hu, Y.-H.; Liao, Q.-D.; Zhong, D. LncRNA KCNQ1OT1 Promoted BMP2 Expression to Regulate Osteogenic Differentiation by Sponging MiRNA-214. Exp. Mol. Pathol. 2019, 107, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Zhang, Z.; Peng, T.; Wang, G.; Xu, Q.; Li, G. MiR-204 Inhibits the Osteogenic Differentiation of Mesenchymal Stem Cells by Targeting Bone Morphogenetic Protein 2. Mol. Med. Rep. 2020, 21, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Fan, J.; Fan, L.; Li, T.; Yang, Y.; Xu, H.; Deng, L.; Li, J.; Li, T.; Weng, X.; et al. MiRNA-10b Reciprocally Stimulates Osteogenesis and Inhibits Adipogenesis Partly through the TGF-β/SMAD2 Signaling Pathway. Aging Dis. 2018, 9, 1058–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, F.-Y.; Deng, R.; Lai, S.-H.; Wen, Q.; Zeng, Y.; Gao, L.; Liu, Y.; Kong, P.; Zhong, J.; Su, Y.; et al. Inhibition of MicroRNA-221-5p Induces Osteogenic Differentiation by Directly Targeting Smad3 in Myeloma Bone Disease Mesenchymal Stem Cells. Oncol. Lett. 2019, 18, 6536–6544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, C.; Yang, S.; Xu, W.; Shen, J.K.; Ye, S.; Liu, X.; Dong, Z.; Xiao, B.; Feng, Y. MiR-708 Promotes Steroid-Induced Osteonecrosis of Femoral Head, Suppresses Osteogenic Differentiation by Targeting SMAD3. Sci. Rep. 2016, 6, 22599. [Google Scholar] [CrossRef] [PubMed]
- Dey, B.K.; Gagan, J.; Yan, Z.; Dutta, A. MiR-26a Is Required for Skeletal Muscle Differentiation and Regeneration in Mice. Genes Dev. 2012, 26, 2180–2191. [Google Scholar] [CrossRef] [Green Version]
- Su, X.; Liao, L.; Shuai, Y.; Jing, H.; Liu, S.; Zhou, H.; Liu, Y.; Jin, Y. MiR-26a Functions Oppositely in Osteogenic Differentiation of BMSCs and ADSCs Depending on Distinct Activation and Roles of Wnt and BMP Signaling Pathway. Cell Death Dis. 2015, 6, e1851. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Zhou, H.; Hong, Y.; Li, J.; Jiang, X.; Huang, H. MiR-30 Family Members Negatively Regulate Osteoblast Differentiation. J. Biol. Chem. 2012, 287, 7503–7511. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Guo, D.; Yang, S.; Sun, H.; Wu, B.; Zhou, D. Inhibition of MiR-222-3p Activity Promoted Osteogenic Differentiation of HBMSCs by Regulating Smad5-RUNX2 Signal Axis. Biochem. Biophys. Res. Commun. 2016, 470, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fakhry, M. Molecular Mechanisms of Mesenchymal Stem Cell Differentiation towards Osteoblasts. World J. Stem Cells 2013, 5, 136. [Google Scholar] [CrossRef]
- Iaculli, F.; Di Filippo, E.S.; Piattelli, A.; Mancinelli, R.; Fulle, S. Dental Pulp Stem Cells Grown on Dental Implant Titanium Surfaces: An in Vitro Evaluation of Differentiation and MicroRNAs Expression: MicroRNAS Expression during Osteoblasts Differentiation. J. Biomed. Mater. Res. B Appl. Biomater. 2017, 105, 953–965. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zheng, L.; Zhou, J.; Chen, Y.; Yang, L.; Deng, F.; Hu, Y. MiR-203-3p Participates in the Suppression of Diabetes-associated Osteogenesis in the Jaw Bone through Targeting Smad1. Int. J. Mol. Med. 2018, 41, 1595–1607. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Geng, J.; Wei, X.; Zhang, R.; Jiang, S. MiR-144-3p Regulates Osteogenic Differentiation and Proliferation of Murine Mesenchymal Stem Cells by Specifically Targeting Smad4. FEBS Lett. 2016, 590, 795–807. [Google Scholar] [CrossRef] [Green Version]
- Xie, Q.; Wei, W.; Ruan, J.; Ding, Y.; Zhuang, A.; Bi, X.; Sun, H.; Gu, P.; Wang, Z.; Fan, X. Effects of MiR-146a on the Osteogenesis of Adipose-Derived Mesenchymal Stem Cells and Bone Regeneration. Sci. Rep. 2017, 7, 42840. [Google Scholar] [CrossRef]
- Li, X.; Guo, L.; Liu, Y.; Su, Y.; Xie, Y.; Du, J.; Zhou, J.; Ding, G.; Wang, H.; Bai, Y.; et al. MicroRNA-21 Promotes Osteogenesis of Bone Marrow Mesenchymal Stem Cells via the Smad7-Smad1/5/8-Runx2 Pathway. Biochem. Biophys. Res. Commun. 2017, 493, 928–933. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.-H.; Chen, L.; Chen, H.-H.; Li, Y.-F.; Luo, H.-B.; Hu, D.-Q.; Chen, P. MiR-15b Ameliorates SONFH by Targeting Smad7 and Inhibiting Osteogenic Differentiation of BMSCs. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 9761–9771. [Google Scholar] [CrossRef]
- Fu, Y.; Tang, Y.; Wang, J.; Guo, Z. MicroRNA-181c Suppresses the Biological Progression of Osteosarcoma via Targeting SMAD7 and Regulating Transforming Growth Factor-β (TGF-β) Signaling Pathway. Med. Sci. Monit. 2019, 25, 4801–4810. [Google Scholar] [CrossRef] [PubMed]
- He, G.; Chen, J.; Huang, D. MiR-877-3p Promotes TGF-Β1-Induced Osteoblast Differentiation of MC3T3-E1 Cells by Targeting Smad7. Exp. Ther. Med. 2019, 18, 312–319. [Google Scholar] [CrossRef] [Green Version]
- Jia, J.; Feng, X.; Xu, W.; Yang, S.; Zhang, Q.; Liu, X.; Feng, Y.; Dai, Z. MiR-17-5p Modulates Osteoblastic Differentiation and Cell Proliferation by Targeting SMAD7 in Non-Traumatic Osteonecrosis. Exp. Mol. Med. 2014, 46, e107. [Google Scholar] [CrossRef] [Green Version]
- Smieszek, A.; Marcinkowska, K.; Pielok, A.; Sikora, M.; Valihrach, L.; Marycz, K. The Role of MiR-21 in Osteoblasts-Osteoclasts Coupling In Vitro. Cells 2020, 9, 479. [Google Scholar] [CrossRef] [Green Version]
- Sanjeev, G.; Sidharthan, D.S.; Pranavkrishna, S.; Pranavadithya, S.; Abhinandan, R.; Akshaya, R.L.; Balagangadharan, K.; Siddabathuni, N.; Srinivasan, S.; Selvamurugan, N. An Osteoinductive Effect of Phytol on Mouse Mesenchymal Stem Cells (C3H10T1/2) towards Osteoblasts. Bioorg. Med. Chem. Lett. 2020, 30, 127137. [Google Scholar] [CrossRef] [PubMed]
- Selvamurugan, N.; He, Z.; Rifkin, D.; Dabovic, B.; Partridge, N.C. Pulsed Electromagnetic Field Regulates MicroRNA 21 Expression to Activate TGF-β Signaling in Human Bone Marrow Stromal Cells to Enhance Osteoblast Differentiation. Stem Cells Int. 2017, 2017. [Google Scholar] [CrossRef] [Green Version]
- Valenti, M.T.; Deiana, M.; Cheri, S.; Dotta, M.; Zamboni, F.; Gabbiani, D.; Schena, F.; Dalle Carbonare, L.; Mottes, M. Physical Exercise Modulates MiR-21-5p, MiR-129-5p, MiR-378-5p, and MiR-188-5p Expression in Progenitor Cells Promoting Osteogenesis. Cells 2019, 8, 742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vishal, M.; Vimalraj, S.; Ajeetha, R.; Gokulnath, M.; Keerthana, R.; He, Z.; Partridge, N.C.; Selvamurugan, N. MicroRNA-590-5p Stabilizes Runx2 by Targeting Smad7 During Osteoblast Differentiation: MICRORNA-590-5P STABILIZES RUNX2 BY TARGETING SMAD7. J. Cell. Physiol. 2017, 232, 371–380. [Google Scholar] [CrossRef]
- Chen, Y.; Sun, C.; Lu, J.; Zou, L.; Hu, M.; Yang, Z.; Xu, Y. MicroRNA-590-5p Antagonizes the Inhibitory Effect of High Glucose on Osteoblast Differentiation by Suppressing Smad7 in MC3T3-E1 Cells. J. Int. Med. Res. 2019, 47, 1740–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Xiao, F.; Gan, Y.; Yuan, W.; Zhai, Z.; Jin, T.; Chen, X.; Zhang, X. Improving Bone Regeneration Using Chordin SiRNA Delivered by PH-Responsive and Non-Toxic Polyspermine Imidazole-4,5-Imine. Cell. Physiol. Biochem. 2018, 46, 133–147. [Google Scholar] [CrossRef]
- Gu, C.; Xu, Y.; Zhang, S.; Guan, H.; Song, S.; Wang, X.; Wang, Y.; Li, Y.; Zhao, G. MiR-27a Attenuates Adipogenesis and Promotes Osteogenesis in Steroid-Induced Rat BMSCs by Targeting PPARγ and GREM1. Sci. Rep. 2016, 6, 38491. [Google Scholar] [CrossRef]
- Delcuve, G.P.; Khan, D.H.; Davie, J.R. Roles of Histone Deacetylases in Epigenetic Regulation: Emerging Paradigms from Studies with Inhibitors. Clin. Epigenet. 2012, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Rotondo, J.C.; Borghi, A.; Selvatici, R.; Mazzoni, E.; Bononi, I.; Corazza, M.; Kussini, J.; Montinari, E.; Gafà, R.; Tognon, M.; et al. Association of Retinoic Acid Receptor β Gene with Onset and Progression of Lichen Sclerosus-Associated Vulvar Squamous Cell Carcinoma. JAMA Dermatol. 2018, 154, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Rotondo, J.C.; Oton-Gonzalez, L.; Selvatici, R.; Rizzo, P.; Pavasini, R.; Campo, G.C.; Lanzillotti, C.; Mazziotta, C.; De Mattei, M.; Tognon, M.; et al. SERPINA1 Gene Promoter Is Differentially Methylated in Peripheral Blood Mononuclear Cells of Pregnant Women. Front. Cell Dev. Biol. 2020, 8. [Google Scholar] [CrossRef]
- Rotondo, J.C.; Borghi, A.; Selvatici, R.; Magri, E.; Bianchini, E.; Montinari, E.; Corazza, M.; Virgili, A.; Tognon, M.; Martini, F. Hypermethylation-Induced Inactivation of the IRF6 Gene as a Possible Early Event in Progression of Vulvar Squamous Cell Carcinoma Associated with Lichen Sclerosus. JAMA Dermatol. 2016, 152, 928–933. [Google Scholar] [CrossRef] [PubMed]
- Stomper, J.; Rotondo, J.C.; Greeve, G.; Lübbert, M. Hypomethylating Agents (HMA) for the Treatment of Acute Myeloid Leukemia and Myelodysplastic Syndromes: Mechanisms of Resistance and Novel HMA-Based Therapies. Leukemia 2021, in press. [Google Scholar]
- Kang, J.S.; Alliston, T.; Delston, R.; Derynck, R. Repression of Runx2 Function by TGF-Beta through Recruitment of Class II Histone Deacetylases by Smad3. EMBO J. 2005, 24, 2543–2555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, J.-Y.; Chuang, P.-C.; Ke, H.-J.; Chen, Y.-S.; Sun, Y.-C.; Wang, F.-S. MicroRNA-29a Mitigates Glucocorticoid Induction of Bone Loss and Fatty Marrow by Rescuing Runx2 Acetylation. Bone 2015, 81, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hassan, M.Q.; Jafferji, M.; Aqeilan, R.I.; Garzon, R.; Croce, C.M.; van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; Lian, J.B. Correction: Biological Functions of MiR-29b Contribute to Positive Regulation of Osteoblast Differentiation. J. Biol. Chem. 2019, 294, 10018. [Google Scholar] [CrossRef] [Green Version]
- De Mattei, M.; Grassilli, S.; Pellati, A.; Brugnoli, F.; De Marchi, E.; Contartese, D.; Bertagnolo, V. Pulsed Electromagnetic Fields Modulate MiRNAs During Osteogenic Differentiation of Bone Mesenchymal Stem Cells: A Possible Role in the Osteogenic-Angiogenic Coupling. Stem Cell Rev. Rep. 2020, 16, 1005–1012. [Google Scholar] [CrossRef]
- Duan, L.; Zhao, H.; Xiong, Y.; Tang, X.; Yang, Y.; Hu, Z.; Li, C.; Chen, S.; Yu, X. MiR-16-2* Interferes with WNT5A to Regulate Osteogenesis of Mesenchymal Stem Cells. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 51, 1087–1102. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, C.-H.; Meng, Y. MicroRNA-1297 Promotes the Progression of Osteoporosis through Regulation of Osteogenesis of Bone Marrow Mesenchymal Stem Cells by Targeting WNT5A. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4541–4550. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-G.; Wang, X.-B.; Zhao, H.; Zhou, C.-N. MicroRNA-9-5p Promotes Osteoporosis Development through Inhibiting Osteogenesis and Promoting Adipogenesis via Targeting Wnt3a. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 456–463. [Google Scholar] [CrossRef]
- Long, H.; Zhu, Y.; Lin, Z.; Wan, J.; Cheng, L.; Zeng, M.; Tang, Y.; Zhao, R. MiR-381 Modulates Human Bone Mesenchymal Stromal Cells (BMSCs) Osteogenesis via Suppressing Wnt Signaling Pathway during Atrophic Nonunion Development. Cell Death Dis. 2019, 10, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, H.; Sun, B.; Cheng, L.; Zhao, S.; Zhu, Y.; Zhao, R.; Zhu, J. MiR-139-5p Represses BMSC Osteogenesis via Targeting Wnt/β-Catenin Signaling Pathway. DNA Cell Biol. 2017, 36, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Guan, X.; Guo, F.; Zhou, J.; Chang, A.; Sun, B.; Cai, Y.; Ma, Z.; Dai, C.; Li, X.; et al. MiR-30e Reciprocally Regulates the Differentiation of Adipocytes and Osteoblasts by Directly Targeting Low-Density Lipoprotein Receptor-Related Protein 6. Cell Death Dis. 2013, 4, e845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Li, H.; Wang, Y.; Li, T.; Fan, J.; Xiao, K.; Zhao, R.C.; Weng, X. MicroRNA-23a Inhibits Osteogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells by Targeting LRP5. Int. J. Biochem. Cell Biol. 2016, 72, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Zhang, J.-F.; Shi, L.; Yang, Z.-M.; Wu, T.-Y.; Wang, H.-X.; Lin, W.-P.; Lu, Y.-F.; Lo, J.H.T.; Zhu, D.-H.; et al. MicroRNA-378 Suppressed Osteogenesis of MSCs and Impaired Bone Formation via Inactivating Wnt/β-Catenin Signaling. Mol. Ther. Nucleic Acids 2020, 21, 1017–1028. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, C.; Han, L.; Liu, L.; Jing, W.; Tang, W.; Tian, W.; Long, J. MiR-154-5p Regulates Osteogenic Differentiation of Adipose-Derived Mesenchymal Stem Cells under Tensile Stress through the Wnt/PCP Pathway by Targeting Wnt11. Bone 2015, 78, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, X. Targeting the Wnt/β-Catenin Signaling Pathway in Cancer. J. Hematol. Oncol. 2020, 13, 165. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Zhang, X.; Feng, X.; Fan, X.; Jin, Z. The Crosstalk between MicroRNAs and the Wnt/β-Catenin Signaling Pathway in Cancer. Oncotarget 2017, 8, 14089–14106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Z.-L.; Meng, Y.-L.; Ge, J.-H. Upregulation of MiR-132 Attenuates Osteoblast Differentiation of UC-MSCs. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1580–1587. [Google Scholar] [CrossRef]
- Zhao, W.; Wu, C.; Dong, Y.; Ma, Y.; Jin, Y.; Ji, Y. MicroRNA-24 Regulates Osteogenic Differentiation via Targeting T-Cell Factor-1. Int. J. Mol. Sci. 2015, 16, 11699–11712. [Google Scholar] [CrossRef] [Green Version]
- Yin, C.; Tian, Y.; Yu, Y.; Yang, C.; Su, P.; Zhao, Y.; Wang, X.; Zhang, K.; Pei, J.; Li, D.; et al. MiR-129-5p Inhibits Bone Formation Through TCF4. Front. Cell Dev. Biol. 2020, 8, 600641. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Zhao, C.; Zhang, P.; Liu, Y.; Jiang, Y.; Wu, E.; Xue, H.; Liu, C.; Li, Z. MiR-26b Modulates OA Induced BMSC Osteogenesis through Regulating GSK3β/β-Catenin Pathway. Exp. Mol. Pathol. 2019, 107, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xu, Z. MiR-27 Promotes Osteoblast Differentiation by Modulating Wnt Signaling. Biochem. Biophys. Res. Commun. 2010, 402, 186–189. [Google Scholar] [CrossRef]
- Hu, W.; Ye, Y.; Zhang, W.; Wang, J.; Chen, A.; Guo, F. MiR-142-3p Promotes Osteoblast Differentiation by Modulating Wnt Signaling. Mol. Med. Rep. 2013, 7, 689–693. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Cai, J.; Cai, X.-H.; Chen, L. MiR-346 Regulates Osteogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells by Targeting the Wnt/β-Catenin Pathway. PLoS ONE 2013, 8, e72266. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Li, Y.; Lin, Z.; Wan, J.; Xu, C.; Zeng, Y.; Zhu, Y. MiR-199b-5p Modulates BMSC Osteogenesis via Suppressing GSK-3β/β-Catenin Signaling Pathway. Biochem. Biophys. Res. Commun. 2016, 477, 749–754. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Tu, Q.; Bonewald, L.F.; He, X.; Stein, G.; Lian, J.; Chen, J. Effects of MiR-335-5p in Modulating Osteogenic Differentiation by Specifically Downregulating Wnt Antagonist DKK1. J. Bone Miner. Res. 2011, 26, 1953–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Xu, H.; Kou, J.; Wang, Q.; Zheng, X.; Yu, T. MiR-9 Promotes Osteoblast Differentiation of Mesenchymal Stem Cells by Inhibiting DKK1 Gene Expression. Mol. Biol. Rep. 2016, 43, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Lin, J.; Wang, G.; Lu, J. MicroRNA-433-3p Promotes Osteoblast Differentiation through Targeting DKK1 Expression. PLoS ONE 2017, 12, e0179860. [Google Scholar] [CrossRef] [Green Version]
- Dai, Z.; Jin, Y.; Zheng, J.; Liu, K.; Zhao, J.; Zhang, S.; Wu, F.; Sun, Z. MiR-217 Promotes Cell Proliferation and Osteogenic Differentiation of BMSCs by Targeting DKK1 in Steroid-Associated Osteonecrosis. Biomed. Pharmacother. 2019, 109, 1112–1119. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Tang, Y.; Yang, Y.; Wang, M.; Yu, H. Low-Magnitude, High-Frequency Vibration Promotes Osteogenic Differentiation via Intensifying MiRNA-335-5p Expression. J. Environ. Pathol. Toxicol. Oncol. 2019, 38, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Di, G.; Kong, L.; Zhao, Q.; Ding, T. MicroRNA-146a Knockdown Suppresses the Progression of Ankylosing Spondylitis by Targeting Dickkopf 1. Biomed. Pharmacother. 2018, 97, 1243–1249. [Google Scholar] [CrossRef]
- Hassan, M.Q.; Maeda, Y.; Taipaleenmaki, H.; Zhang, W.; Jafferji, M.; Gordon, J.A.R.; Li, Z.; Croce, C.M.; van Wijnen, A.J.; Stein, J.L.; et al. MiR-218 Directs a Wnt Signaling Circuit to Promote Differentiation of Osteoblasts and Osteomimicry of Metastatic Cancer Cells. J. Biol. Chem. 2012, 287, 42084–42092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.-B.; Zhong, W.-J.; Wang, L. A Signal-Amplification Circuit between MiR-218 and Wnt/β-Catenin Signal Promotes Human Adipose Tissue-Derived Stem Cells Osteogenic Differentiation. Bone 2014, 58, 59–66. [Google Scholar] [CrossRef]
- Ma, S.; Wang, D.-D.; Ma, C.-Y.; Zhang, Y.-D. MicroRNA-96 Promotes Osteoblast Differentiation and Bone Formation in Ankylosing Spondylitis Mice through Activating the Wnt Signaling Pathway by Binding to SOST. J. Cell. Biochem. 2019, 120, 15429–15442. [Google Scholar] [CrossRef]
- Guo, D.; Li, Q.; Lv, Q.; Wei, Q.; Cao, S.; Gu, J. MiR-27a Targets SFRP1 in HFOB Cells to Regulate Proliferation, Apoptosis and Differentiation. PLoS ONE 2014, 9, e91354. [Google Scholar] [CrossRef]
- Almalki, S.G.; Agrawal, D.K. Key Transcription Factors in the Differentiation of Mesenchymal Stem Cells. Differ. Res. Biol. Divers. 2016, 92, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Narayanan, A.; Srinaath, N.; Rohini, M.; Selvamurugan, N. Regulation of Runx2 by MicroRNAs in Osteoblast Differentiation. Life Sci. 2019, 232, 116676. [Google Scholar] [CrossRef] [PubMed]
- Mevel, R.; Draper, J.E.; Lie-A-Ling, M.; Kouskoff, V.; Lacaud, G. RUNX Transcription Factors: Orchestrators of Development. Development 2019, 146. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Liu, S.; Li, J.; Zhao, S.; Yi, Z. Roles for MiRNAs in Osteogenic Differentiation of Bone Marrow Mesenchymal Stem Cells. Stem Cell Res. Ther. 2019, 10, 197. [Google Scholar] [CrossRef]
- Zhang, Y.; Xie, R.-L.; Croce, C.M.; Stein, J.L.; Lian, J.B.; van Wijnen, A.J.; Stein, G.S. A Program of MicroRNAs Controls Osteogenic Lineage Progression by Targeting Transcription Factor Runx2. Proc. Natl. Acad. Sci. USA 2011, 108, 9863–9868. [Google Scholar] [CrossRef] [Green Version]
- Gay, I.; Cavender, A.; Peto, D.; Sun, Z.; Speer, A.; Cao, H.; Amendt, B.A. Differentiation of Human Dental Stem Cells Reveals a Role for MicroRNA-218. J. Periodontal Res. 2014, 49, 110–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seenprachawong, K.; Nuchnoi, P.; Nantasenamat, C.; Prachayasittikul, V.; Supokawej, A. Computational Identification of MiRNAs That Modulate the Differentiation of Mesenchymal Stem Cells to Osteoblasts. PeerJ 2016, 4, e1976. [Google Scholar] [CrossRef] [Green Version]
- Deng, L.; Hu, G.; Jin, L.; Wang, C.; Niu, H. Involvement of MicroRNA-23b in TNF-α-Reduced BMSC Osteogenic Differentiation via Targeting Runx2. J. Bone Miner. Metab. 2018, 36, 648–660. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Liu, D.; Li, C.-G.; Wang, Y.-J. MiR-203 Is Essential for the Shift from Osteogenic Differentiation to Adipogenic Differentiation of Mesenchymal Stem Cells in Postmenopausal Osteoporosis. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5804–5814. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Lu, Y.; Mai, Z.; Liu, R.; Peng, Z.; Chen, L.; Chen, Z.; Wang, R.; Ai, H. Suppressing MicroRNA-30b by Estrogen Promotes Osteogenesis in Bone Marrow Mesenchymal Stem Cells. Stem Cells Int. 2019, 2019, 7547506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhou, K.; Wu, L.; Gu, H.; Huang, Z.; Xu, J. Downregulation of MicroRNA-143 Promotes Osteogenic Differentiation of Human Adipose-derived Mesenchymal Stem Cells through the K-Ras/MEK/ERK Signaling Pathway. Int. J. Mol. Med. 2020, 46, 965–976. Available online: https://pubmed.ncbi.nlm.nih.gov/32582994/ (accessed on 20 October 2020). [CrossRef]
- Hassan, M.Q.; Gordon, J.A.R.; Beloti, M.M.; Croce, C.M.; van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; Lian, J.B. A Network Connecting Runx2, SATB2, and the MiR-23a~27a~24-2 Cluster Regulates the Osteoblast Differentiation Program. Proc. Natl. Acad. Sci. USA 2010, 107, 19879–19884. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, S.; Yuan, S.; Zhang, H.; Liu, J. MicroRNA-23a Inhibits Osteogenesis of Periodontal Mesenchymal Stem Cells by Targeting Bone Morphogenetic Protein Signaling. Arch. Oral Biol. 2019, 102, 93–100. [Google Scholar] [CrossRef]
- Park, J.; Wada, S.; Ushida, T.; Akimoto, T. The MicroRNA-23a Has Limited Roles in Bone Formation and Homeostasis In Vivo. Physiol. Res. 2015, 64, 711–719. [Google Scholar] [CrossRef]
- Zhang, R.; Weng, Y.; Li, B.; Jiang, Y.; Yan, S.; He, F.; Chen, X.; Deng, F.; Wang, J.; Shi, Q. BMP9-Induced Osteogenic Differentiation Is Partially Inhibited by MiR-30a in the Mesenchymal Stem Cell Line C3H10T1/2. J. Mol. Histol. 2015, 46, 399–407. [Google Scholar] [CrossRef]
- Yi, J.; Liu, D.; Xiao, J. LncRNA MALAT1 Sponges MiR-30 to Promote Osteoblast Differentiation of Adipose-Derived Mesenchymal Stem Cells by Promotion of Runx2 Expression. Cell Tissue Res. 2019, 376, 113–121. [Google Scholar] [CrossRef]
- Li, Z.; Hassan, M.Q.; Volinia, S.; van Wijnen, A.J.; Stein, J.L.; Croce, C.M.; Lian, J.B.; Stein, G.S. A MicroRNA Signature for a BMP2-Induced Osteoblast Lineage Commitment Program. Proc. Natl. Acad. Sci. USA 2008, 105, 13906–13911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Wu, Y.; Shiozaki, Y.; Sugimoto, Y.; Takigawa, T.; Tanaka, M.; Matsukawa, A.; Ozaki, T. MiRNA-133a-5p Inhibits the Expression of Osteoblast Differentiation-Associated Markers by Targeting the 3’ UTR of RUNX2. DNA Cell Biol. 2018, 37, 199–209. [Google Scholar] [CrossRef]
- Shi, X.; Zhang, Z. MicroRNA-135a-5p Is Involved in Osteoporosis Progression through Regulation of Osteogenic Differentiation by Targeting RUNX2. Exp. Ther. Med. 2019, 18, 2393–2400. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Sun, Q.; Wan, C.; Li, L.; Zhang, L.; Chen, Z. MicroRNA-338-3p Regulates Osteogenic Differentiation of Mouse Bone Marrow Stromal Stem Cells by Targeting Runx2 and Fgfr2. J. Cell. Physiol. 2014, 229, 1494–1502. [Google Scholar] [CrossRef] [PubMed]
- Qiao, W.; Chen, L.; Zhang, M. MicroRNA-205 Regulates the Calcification and Osteoblastic Differentiation of Vascular Smooth Muscle Cells. Cell. Physiol. Biochem. 2014, 33, 1945–1953. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Li, Y.; Xu, J.; Xiong, Z. Advances in the Role of MiRNAs in the Occurrence and Development of Osteosarcoma. Open Med. 2020, 15, 1003–1011. [Google Scholar] [CrossRef]
- Hu, N.; Feng, C.; Jiang, Y.; Miao, Q.; Liu, H. Regulative Effect of Mir-205 on Osteogenic Differentiation of Bone Mesenchymal Stem Cells (BMSCs): Possible Role of SATB2/Runx2 and ERK/MAPK Pathway. Int. J. Mol. Sci. 2015, 16, 10491–10506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, L.; Zuo, R.; Wang, M.; Wang, W.; Xu, J.; Chai, Y.; Guan, J.; Kang, Q. Silencing MicroRNA-137-3p, Which Targets RUNX2 and CXCL12 Prevents Steroid-Induced Osteonecrosis of the Femoral Head by Facilitating Osteogenesis and Angiogenesis. Int. J. Biol. Sci. 2020, 16, 655–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, R.; Zhai, Y.; Ao, L.; Fullerton, D.A.; Meng, X. MicroRNA-204 Deficiency in Human Aortic Valves Elevates Valvular Osteogenic Activity. Int. J. Mol. Sci. 2019, 21, 76. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Zhao, L.; Xing, L.; Chen, D. MicroRNA-204 Regulates Runx2 Protein Expression and Mesenchymal Progenitor Cell Differentiation. Stem Cells 2010, 28, 357–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Ji, X.; She, F.; Gao, Y.; Tang, P. MiR-628-3p Regulates Osteoblast Differentiation by Targeting RUNX2: Possible Role in Atrophic Non-Union. Int. J. Mol. Med. 2017, 39, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, T.; Ochi, H.; Sunamura, S.; Haiden, A.; Bando, W.; Inose, H.; Okawa, A.; Asou, Y.; Takeda, S. MicroRNA-145 Regulates Osteoblastic Differentiation by Targeting the Transcription Factor Cbfb. FEBS Lett. 2015, 589, 3302–3308. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Chen, Z.; Cai, C.; Li, G.; Wang, X.; Shi, Z. MicroRNA-505 Is Involved in the Regulation of Osteogenic Differentiation of MC3T3-E1 Cells Partially by Targeting RUNX2. J. Orthop. Surg. 2020, 15, 143. [Google Scholar] [CrossRef] [Green Version]
- Bellavia, D.; Salamanna, F.; Raimondi, L.; De Luca, A.; Carina, V.; Costa, V.; Alessandro, R.; Fini, M.; Giavaresi, G. Deregulated MiRNAs in Osteoporosis: Effects in Bone Metastasis. Cell. Mol. Life Sci. 2019, 76, 3723–3744. [Google Scholar] [CrossRef] [PubMed]
- Han, T.-S.; Hur, K.; Cho, H.-S.; Ban, H.S. Epigenetic Associations between LncRNA/CircRNA and MiRNA in Hepatocellular Carcinoma. Cancers 2020, 12, 2622. [Google Scholar] [CrossRef]
- Wu, D.; Ma, L. Downregulating MicroRNA-152-3p Promotes the Viability and Osteogenic Differentiation of Periodontal Ligament Stem Cells via Targeting Integrin Alpha 5. Arch. Oral Biol. 2020, 120, 104930. [Google Scholar] [CrossRef]
- Zhao, H.; Yan, P.; Wang, J.; Zhang, Y.; Zhang, M.; Wang, Z.; Fu, Q.; Liang, W. Clinical Significance of Tumor MiR-21, MiR-221, MiR-143, and MiR-106a as Biomarkers in Patients with Osteosarcoma. Int. J. Biol. Markers 2019, 34, 184–193. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Cell Type | Target Gene | Effect on Osteogenesis |
|---|---|---|---|
| let-7a-5p | PMOP BM-MSCs | TGFβR1 | − |
| miR-9 | MSCs | DKK1 | + |
| miR-9-5p | MSCs | Wnt3a | − |
| miR-10b | hASCs | SMAD2 | + |
| miR-16-2-3p | hBM-MSCs | Wnt5a | − |
| miR-17-5p | hBM-MSCs | SMAD7 | + |
| miR-21 | BM-MSCs | SMAD7 | + |
| miR-21-5p | BM-MSCs | SMAD7 | + |
| miR-21a | C3H10T1/2 | SMAD7 | + |
| miR-23a | MSCs, PDLSCs, MC3T3-E1, hBM-MSCs | RUNX2, LRP6 | − |
| miR-23b | MSCs | RUNX2 | − |
| miR-24 | BM-MSCs, MC3T3-E1 | TCF-1 | − |
| miR-26a | BM-MSCs, hASCs | SMAD1, GSK3β | −/+ |
| miR-26b | BM-MSCs | GSK3β | + |
| miR-27a | hFOB1.19, BM-MSCs | APC, SFRP1, GREM1 | −/+ |
| miR-29a | BM-MSCs, MC3TC-E1 | HDAC4 | + |
| miR-29b | BM-MSCs, MC3TC-E1 | HDAC4 | + |
| miR-30 | MC3T3-E1, MSCs, hASCs, C3H10T1/2 | RUNX2, SMAD1 | − |
| miR-30a | MC3T3-E1 | RUNX2, SMAD1 | − |
| miR-30b | MSCs, MC3T3-E1 | RUNX2, SMAD1 | − |
| miR-30c | MSCs, hASCs, MC3T3-E1 | RUNX2, SMAD1 | − |
| miR-30d | MC3T3-E1 | RUNX2, SMAD1 | − |
| miR-30e | hBM-MSCs | LRP6 | − |
| miR-34c | MSCs, MC3T3-E1 | RUNX2 | − |
| miR-93-5p | MSCs, BM-MSCs | BMP2 | − |
| miR-96 | Mice AS cells | SOST | + |
| miR-98 | hMSCs | BMP2 | − |
| miR-100 | hMSCs | BMPR2 | − |
| miR-125b | hBM-MSCs | BMPR1B | − |
| miR-129-5p | MC3T3-E1, C57BL6 | TCF-4 | − |
| miR-132 | UC-MSCs | β-catenin | − |
| miR-133 | C2C12 | RUNX2 | − |
| miR-133a | DPSCs, C2C12 | RUNX2, SMAD5 | − |
| miR-133a-5p | MC3T3-E1 | RUNX2 | − |
| miR-133b | DPSCs | RUNX2, SMAD5 | − |
| miR-135 | C2C12 | SMAD1, SMAD5 | − |
| miR-135a | MSCs, MC3T3-E1, ATDC5, C2C12, DPSCs | RUNX2, SMAD5 | − |
| miR-135a-5p | C2C12 | RUNX2 | − |
| miR-137 | MSCs | RUNX2 | − |
| miR-137-3p | BM-MSCs | RUNX2 | − |
| miR-139-5p | hBM-MSCs | β-catenin, FZD4 | − |
| miR-140-5p | hMSCs | BMP2 | − |
| miR-142-3p | hFOB1.19 | APC | + |
| miR-143 | MSCs | RUNX2 | − |
| miR-144-3p | C3H10T1/2 | SMAD4 | − |
| miR-145 | MC3T3-E1 | CBFB | − |
| miR-146a | hASCs, AS fibroblast | SMAD4, DKK1 | −/+ |
| miR-153 | hMSCs | BMPR2 | − |
| miR-154-5p | hASCs | Wnt11 | − |
| miR-155 | C2C12, MEF | BMPR2 | − |
| miR-195-5p | PDLSCs | BMPR1A | − |
| miR-199b-5p | hBM-MSCs | GSK3β | + |
| miR-203 | MSCs | RUNX2 | − |
| miR-203-3p | MSCs | SMAD1 | − |
| miR-204 | C3H10T1/2, C2C12, ST2, BM-MSCs | RUNX2, BMP2 | − |
| miR-205 | MSCs, BM-MSCs | RUNX2, SATB2 | − |
| miR-208a-3p | MC3T3-E1 | ACVR1/Alk2 | − |
| miR-214 | BM-MSCs | BMP2 | − |
| miR-217 | MSCs | RUNX2, DKK1 | −/+ |
| miR-218 | hASCs, BM-MSCs, MC3T3, hDPSCs | DKK2, SFRP2, SOST, RUNX2 | +/− |
| miR-221 | MSCs | RUNX2 | − |
| miR-221-5p | MB D-MSCs | SMAD3 | − |
| miR-222-3p | BM-MSCs | RUNX2, SMAD5 | − |
| miR-335-5p | MSCs, C3H10T-1/2, MC3T3-E1, MLO-A5, -Y4 | DKK1 | + |
| miR-338 | MSCs | RUNX2 | − |
| miR-338-3p | BM-MSCs mice | RUNX2, FGFR2 | − |
| miR-346 | hBM-MSCs | GSK3β | + |
| miR-370 | MC3T3-E1 | BMP2 | − |
| miR-378 | MSCs | Wnt6, Wnt10a | − |
| miR-381 | hBM-MSCs | Wnt5a, FZD3 | − |
| miR-433-3p | MSCs, hFOB1.19, ROS17/2.8 | DKK1 | + |
| miR-505 | MC3T3-E1 | RUNX2 | − |
| miR-590-3p | hMSCs | APC | + |
| miR-590-5p | C3H10T1/2, MG63, MC3T3-E1 | SMAD7 | + |
| miR-628-3p | MG63 | RUNX2 | − |
| miR-708 | MSCs | SMAD3 | − |
| miR-1297 | hBM-MSCs | Wnt5a | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazziotta, C.; Lanzillotti, C.; Iaquinta, M.R.; Taraballi, F.; Torreggiani, E.; Rotondo, J.C.; Otòn-Gonzalez, L.; Mazzoni, E.; Frontini, F.; Bononi, I.; et al. MicroRNAs Modulate Signaling Pathways in Osteogenic Differentiation of Mesenchymal Stem Cells. Int. J. Mol. Sci. 2021, 22, 2362. https://doi.org/10.3390/ijms22052362
Mazziotta C, Lanzillotti C, Iaquinta MR, Taraballi F, Torreggiani E, Rotondo JC, Otòn-Gonzalez L, Mazzoni E, Frontini F, Bononi I, et al. MicroRNAs Modulate Signaling Pathways in Osteogenic Differentiation of Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2021; 22(5):2362. https://doi.org/10.3390/ijms22052362
Chicago/Turabian StyleMazziotta, Chiara, Carmen Lanzillotti, Maria Rosa Iaquinta, Francesca Taraballi, Elena Torreggiani, John Charles Rotondo, Lucia Otòn-Gonzalez, Elisa Mazzoni, Francesca Frontini, Ilaria Bononi, and et al. 2021. "MicroRNAs Modulate Signaling Pathways in Osteogenic Differentiation of Mesenchymal Stem Cells" International Journal of Molecular Sciences 22, no. 5: 2362. https://doi.org/10.3390/ijms22052362
APA StyleMazziotta, C., Lanzillotti, C., Iaquinta, M. R., Taraballi, F., Torreggiani, E., Rotondo, J. C., Otòn-Gonzalez, L., Mazzoni, E., Frontini, F., Bononi, I., De Mattei, M., Tognon, M., & Martini, F. (2021). MicroRNAs Modulate Signaling Pathways in Osteogenic Differentiation of Mesenchymal Stem Cells. International Journal of Molecular Sciences, 22(5), 2362. https://doi.org/10.3390/ijms22052362